
Nifuratel Induces Triple-Negative Breast Cancer Cell G2/M Phase Block and Apoptosis by Regulating GADD45A
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
1. Introduction
2. Results
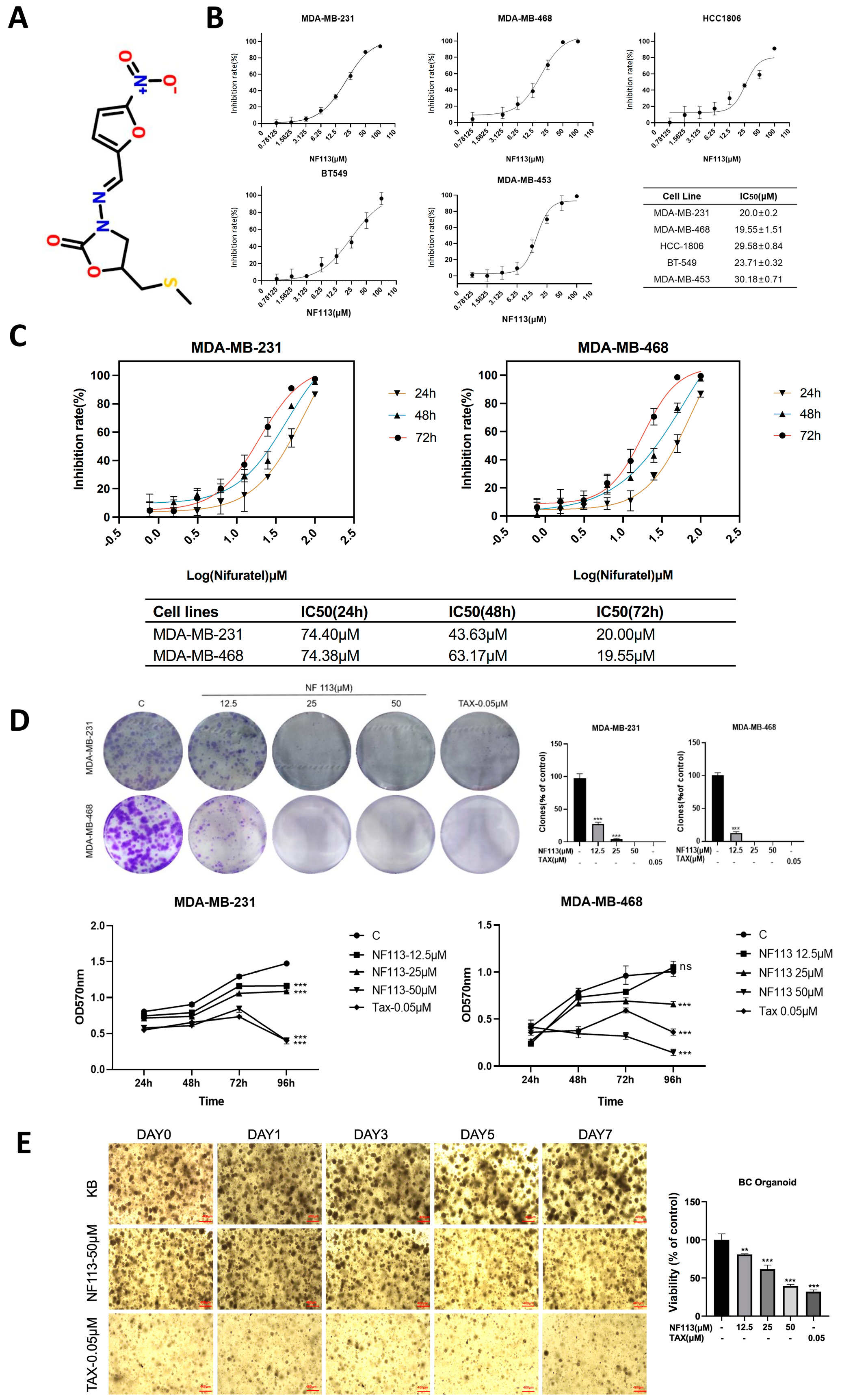
2.1. Nifuratel Has Viability Inhibition on Triple-Negative Breast Cancer Cells
2.2. Nifuratel Induced Apoptosis of TNBC Cells
2.3. Nifuratel Induced Phase Arrest of TNBC Cells
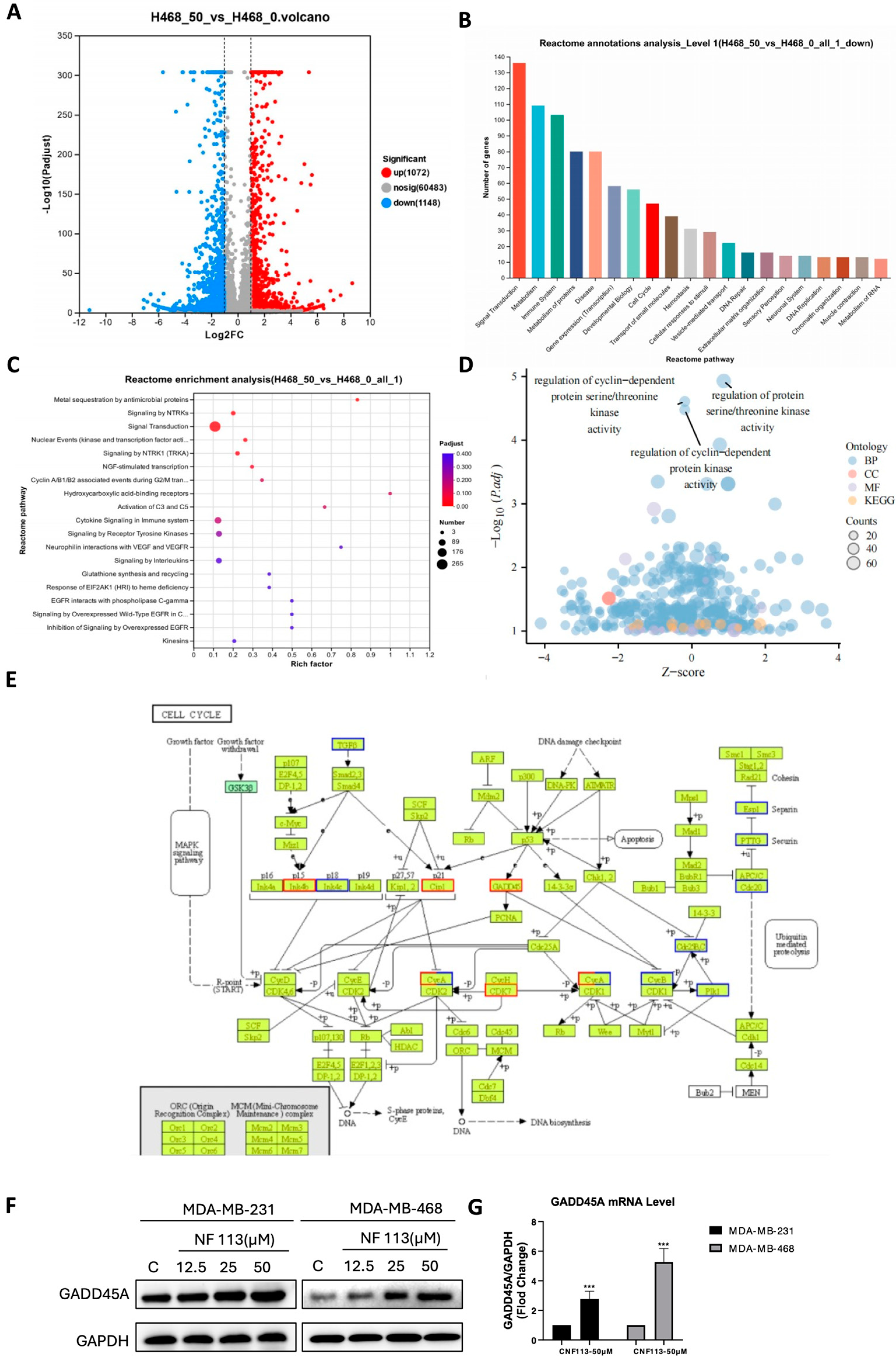
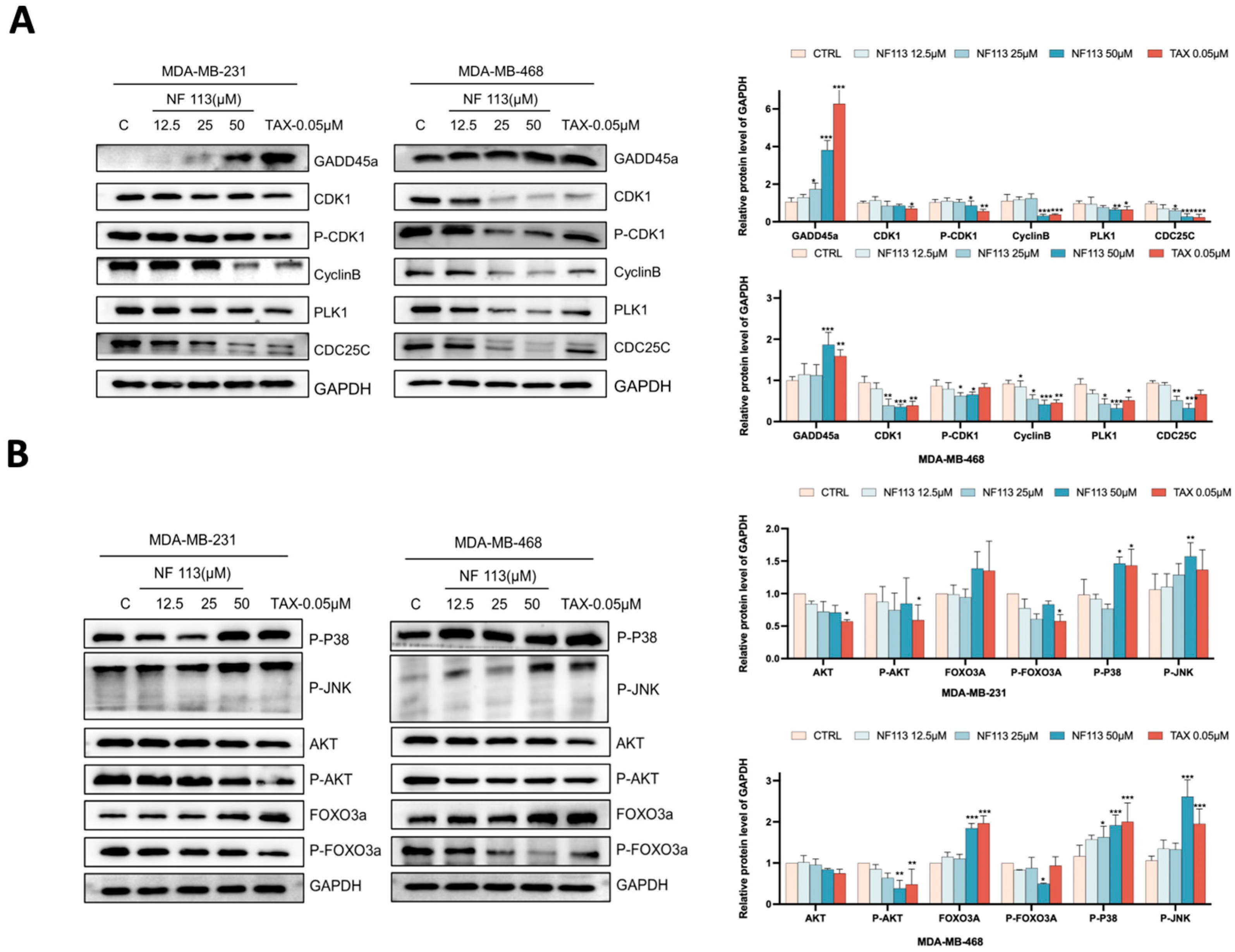
2.4. Nifuratel Inhibited Cell Proliferation Arrest by Increasing GADD45A Levels
2.5. Nifuratel Regulated the Expression of GADD45A and Its Downstream Signaling Pathways
2.6. Nifuratel Inhibited TNBC Growth In Vivo
3. Discussion
4. Materials and Methods
4.1. Chemicals and Reagents
4.2. Cell Culture
4.3. Cell Viability Assay
4.4. Colony Formation Assay
4.5. Cell Cycle Analysis
4.6. Apoptosis Analysis
4.7. Immunoblotting Assay
4.8. MDA-MB-468 Subcutaneous Tumor Model
4.9. Organoid Model
4.10. Statistical Analysis
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Bray, F.; Laversanne, M.; Sung, H.; Ferlay, J.; Siegel, R.L.; Soerjomataram, I.; Jemal, A. Global Cancer Statistics 2022: GLOBOCAN Estimates of Incidence and Mortality Worldwide for 36 Cancers in 185 Countries. CA Cancer J. Clin. 2024, 74, 229–263. [Google Scholar] [CrossRef]
- de Ruijter, T.C.; Veeck, J.; de Hoon, J.P.J.; van Engeland, M.; Tjan-Heijnen, V.C. Characteristics of Triple-Negative Breast Cancer. J. Cancer Res. Clin. Oncol. 2011, 137, 183–192. [Google Scholar] [CrossRef] [PubMed]
- Dent, R.; Trudeau, M.; Pritchard, K.I.; Hanna, W.M.; Kahn, H.K.; Sawka, C.A.; Lickley, L.A.; Rawlinson, E.; Sun, P.; Narod, S.A. Triple-Negative Breast Cancer: Clinical Features and Patterns of Recurrence. Clin. Cancer Res. 2007, 13, 4429–4434. [Google Scholar] [CrossRef]
- Huang, X.; Qiao, Y.; Brady, S.W.; Factor, R.E.; Downs-Kelly, E.; Farrell, A.; McQuerry, J.A.; Shrestha, G.; Jenkins, D.; Johnson, W.E.; et al. Novel Temporal and Spatial Patterns of Metastatic Colonization from Breast Cancer Rapid-Autopsy Tumor Biopsies. Genome Med. 2021, 13, 170. [Google Scholar] [CrossRef] [PubMed]
- Marra, A.; Curigliano, G. Adjuvant and Neoadjuvant Treatment of Triple-Negative Breast Cancer with Chemotherapy. Cancer J. 2021, 27, 41–49. [Google Scholar] [CrossRef] [PubMed]
- Li, Y.; Zhang, H.; Merkher, Y.; Chen, L.; Liu, N.; Leonov, S.; Chen, Y. Recent Advances in Therapeutic Strategies for Triple-Negative Breast Cancer. J. Hematol. Oncol. 2022, 15, 121. [Google Scholar] [CrossRef] [PubMed]
- Küng, E.; Fürnkranz, U.; Walochnik, J. Chemotherapeutic Options for the Treatment of Human Trichomoniasis. Int. J. Antimicrob. Agents 2019, 53, 116–127. [Google Scholar] [CrossRef]
- Polatti, F. Bacterial Vaginosis, Atopobium Vaginae and Nifuratel. Curr. Clin. Pharmacol. 2012, 7, 36–40. [Google Scholar] [CrossRef]
- Evans, B.A.; Catterall, R.D. Nifuratel Compared with Metronidazole in the Treatment of Trichomonal Vaginitis. Br. Med. J. 1970, 2, 335–336. [Google Scholar] [CrossRef]
- Pushpakom, S.; Iorio, F.; Eyers, P.A.; Escott, K.J.; Hopper, S.; Wells, A.; Doig, A.; Guilliams, T.; Latimer, J.; McNamee, C.; et al. Drug Repurposing: Progress, Challenges and Recommendations. Nat. Rev. Drug Discov. 2019, 18, 41–58. [Google Scholar] [CrossRef]
- Sleire, L.; Førde, H.E.; Netland, I.A.; Leiss, L.; Skeie, B.S.; Enger, P.Ø. Drug Repurposing in Cancer. Pharmacol. Res. 2017, 124, 74–91. [Google Scholar] [CrossRef] [PubMed]
- Pantziarka, P.; Bouche, G.; Meheus, L.; Sukhatme, V.; Sukhatme, V.P.; Vikas, P. The Repurposing Drugs in Oncology (ReDO) Project. Ecancermedicalscience 2014, 8, 442. [Google Scholar] [CrossRef] [PubMed]
- Zheng, H.; Hong, H.; Zhang, L.; Cai, X.; Hu, M.; Cai, Y.; Zhou, B.; Lin, J.; Zhao, C.; Hu, W. Nifuratel, a Novel STAT3 Inhibitor with Potent Activity against Human Gastric Cancer Cells. Cancer Manag. Res. 2017, 9, 565–572. [Google Scholar] [CrossRef]
- Patel, K.; Murray, M.G.; Whelan, K.A. Roles for GADD45 in Development and Cancer. Adv. Exp. Med. Biol. 2022, 1360, 23–39. [Google Scholar] [CrossRef]
- Pietrasik, S.; Zajac, G.; Morawiec, J.; Soszynski, M.; Fila, M.; Blasiak, J. Interplay between BRCA1 and GADD45A and Its Potential for Nucleotide Excision Repair in Breast Cancer Pathogenesis. Int. J. Mol. Sci. 2020, 21, 870. [Google Scholar] [CrossRef]
- Hollander, M.C.; Sheikh, M.S.; Bulavin, D.V.; Lundgren, K.; Augeri-Henmueller, L.; Shehee, R.; Molinaro, T.A.; Kim, K.E.; Tolosa, E.; Ashwell, J.D.; et al. Genomic Instability in Gadd45a-Deficient Mice. Nat. Genet. 1999, 23, 176–184. [Google Scholar] [CrossRef]
- Tamura, R.E.; de Vasconcellos, J.F.; Sarkar, D.; Libermann, T.A.; Fisher, P.B.; Zerbini, L.F. GADD45 Proteins: Central Players in Tumorigenesis. Curr. Mol. Med. 2012, 12, 634–651. [Google Scholar] [CrossRef]
- Salvador, J.M.; Brown-Clay, J.D.; Fornace, A.J. Gadd45 in Stress Signaling, Cell Cycle Control, and Apoptosis. Adv. Exp. Med. Biol. 2013, 793, 1–19. [Google Scholar] [CrossRef] [PubMed]
- Camilleri-Robles, C.; Serras, F.; Corominas, M. Role of D-GADD45 in JNK-Dependent Apoptosis and Regeneration in Drosophila. Genes 2019, 10, 378. [Google Scholar] [CrossRef]
- Palomer, X.; Salvador, J.M.; Griñán-Ferré, C.; Barroso, E.; Pallàs, M.; Vázquez-Carrera, M. GADD45A: With or without You. Med. Res. Rev. 2024, 44, 1375–1403. [Google Scholar] [CrossRef]
- Takekawa, M.; Saito, H. A Family of Stress-Inducible GADD45-like Proteins Mediate Activation of the Stress-Responsive MTK1/MEKK4 MAPKKK. Cell 1998, 95, 521–530. [Google Scholar] [CrossRef] [PubMed]
- Libson, S.; Lippman, M. A Review of Clinical Aspects of Breast Cancer. Int. Rev. Psychiatry 2014, 26, 4–15. [Google Scholar] [CrossRef] [PubMed]
- Zheng, H.; Chen, Z.; Cai, A.; Lin, X.; Jiang, X.; Zhou, B.; Wang, J.; Yao, Q.; Chen, R.; Kou, L. Nanoparticle Mediated Codelivery of Nifuratel and Doxorubicin for Synergistic Anticancer Therapy through STAT3 Inhibition. Colloids Surf. B Biointerfaces 2020, 193, 111109. [Google Scholar] [CrossRef] [PubMed]
- Jang, H.-J.; Yang, J.H.; Hong, E.; Jo, E.; Lee, S.; Lee, S.; Choi, J.S.; Yoo, H.S.; Kang, H. Chelidonine Induces Apoptosis via GADD45a-P53 Regulation in Human Pancreatic Cancer Cells. Integr. Cancer Ther. 2021, 20, 15347354211006191. [Google Scholar] [CrossRef]
- Schneider, G.; Weber, A.; Zechner, U.; Oswald, F.; Friess, H.M.; Schmid, R.M.; Liptay, S. GADD45alpha Is Highly Expressed in Pancreatic Ductal Adenocarcinoma Cells and Required for Tumor Cell Viability. Int. J. Cancer 2006, 118, 2405–2411. [Google Scholar] [CrossRef]
- Yu, K.-D.; Di, G.-H.; Li, W.-F.; Rao, N.-Y.; Fan, L.; Yuan, W.-T.; Hu, Z.; Wu, J.; Shen, Z.-Z.; Huang, W.; et al. Genetic Contribution of GADD45A to Susceptibility to Sporadic and Non-BRCA1/2 Familial Breast Cancers: A Systematic Evaluation in Chinese Populations. Breast Cancer Res. Treat. 2010, 121, 157–167. [Google Scholar] [CrossRef]
- Xie, L.; Jia, L.; Qu, J.; Chen, D.; Lv, Y.; Li, H.; Zheng, J. Expression and Prognostic Significance of the P53-Related DNA Damage Repair Proteins Checkpoint Kinase 1 (CHK1) and Growth Arrest and DNA-Damage-Inducible 45 Alpha (GADD45A) in Human Oral Squamous Cell Carcinoma. Eur. J. Oral Sci. 2020, 128, 128–135. [Google Scholar] [CrossRef]
- Takahashi, S.; Saito, S.; Ohtani, N.; Sakai, T. Involvement of the Oct-1 Regulatory Element of the Gadd45 Promoter in the P53-Independent Response to Ultraviolet Irradiation. Cancer Res. 2001, 61, 1187–1195. [Google Scholar]
- Qi, L.; Wang, Y.; Su, S.; Wang, M.; Jablonska, E.; Jia, Y.; Wang, R.; Hao, S.; Feng, C.; Li, G.; et al. Sodium Selenite Inhibits Cervical Cancer Growth via ROS Mediated AMPK/FOXO3a/GADD45a Axis. Chem. Biol. Interact. 2022, 367, 110171. [Google Scholar] [CrossRef]
- Zhan, Q.; Antinore, M.J.; Wang, X.W.; Carrier, F.; Smith, M.L.; Harris, C.C.; Fornace, A.J. Association with Cdc2 and Inhibition of Cdc2/Cyclin B1 Kinase Activity by the P53-Regulated Protein Gadd45. Oncogene 1999, 18, 2892–2900. [Google Scholar] [CrossRef]
- Mendling, W.; Mailland, F. Microbiological and Pharmaco-Toxicological Profile of Nifuratel and Its Favourable Risk/Benefit Ratio for the Treatment of Vulvo-Vaginal Infections. A Review. Arzneimittelforschung 2002, 52, 8–13. [Google Scholar] [CrossRef] [PubMed]



Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Hou, Y.; Hao, H.; Yuan, Y.; Zhang, J.; Liu, Z.; Nie, Y.; Zhang, S.; Yuan, S.; Yang, M. Nifuratel Induces Triple-Negative Breast Cancer Cell G2/M Phase Block and Apoptosis by Regulating GADD45A. Pharmaceuticals 2024, 17, 1269. https://doi.org/10.3390/ph17101269
Hou Y, Hao H, Yuan Y, Zhang J, Liu Z, Nie Y, Zhang S, Yuan S, Yang M. Nifuratel Induces Triple-Negative Breast Cancer Cell G2/M Phase Block and Apoptosis by Regulating GADD45A. Pharmaceuticals. 2024; 17(10):1269. https://doi.org/10.3390/ph17101269
Chicago/Turabian StyleHou, Yuhang, Hongyun Hao, Yan Yuan, Jing Zhang, Zhengrui Liu, Yimin Nie, Shichang Zhang, Shengtao Yuan, and Mei Yang. 2024. "Nifuratel Induces Triple-Negative Breast Cancer Cell G2/M Phase Block and Apoptosis by Regulating GADD45A" Pharmaceuticals 17, no. 10: 1269. https://doi.org/10.3390/ph17101269
APA StyleHou, Y., Hao, H., Yuan, Y., Zhang, J., Liu, Z., Nie, Y., Zhang, S., Yuan, S., & Yang, M. (2024). Nifuratel Induces Triple-Negative Breast Cancer Cell G2/M Phase Block and Apoptosis by Regulating GADD45A. Pharmaceuticals, 17(10), 1269. https://doi.org/10.3390/ph17101269