


Chronic Effects of Apelin on Cardiovascular Regulation and Angiotensin II-Induced Hypertension

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
1. Introduction
2. Results
2.1. Effect of Chronic Subcutaneous Infusion of Apelin on BP, HR, and Cardiac Morphology
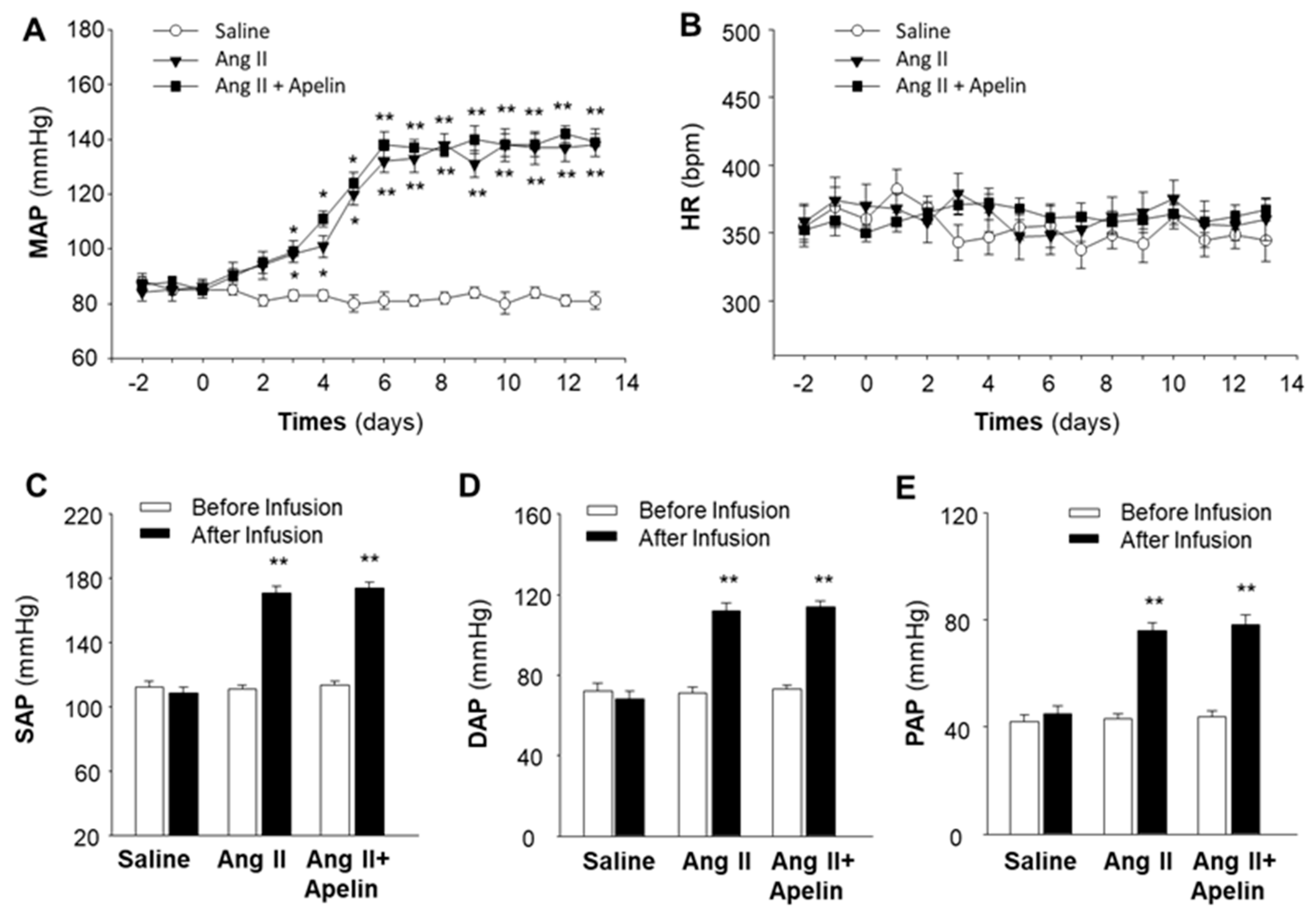
2.2. Chronic Effect of Apelin and Ang II on BP and HR
2.3. Chronic Effect of Apelin and Ang II on Cardiac Morphology
2.4. Chronic Effect of Apelin and Ang II on Cardiac Fibrosis
3. Discussion
4. Material and Methods
4.1. Animals and Materials
4.2. Chronic Recording of BP and HR
4.3. Chronic Administration of Apelin
4.4. Heart Histology and mRNA Detection
4.5. Statistical Analyses
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Tsao, C.W.; Aday, A.W.; Almarzooq, Z.I.; Anderson, C.A.; Arora, P.; Avery, C.L.; Baker-Smith, C.M.; Beaton, A.Z.; Boehme, A.K.; Buxton, A.E.; et al. Heart Disease and Stroke Statistics-2023 Update: A Report from the American Heart Association. Circulation 2023, 147, e93–e621. [Google Scholar] [CrossRef]
- Touyz, R.M.; Schiffrin, E.L. A Compendium on Hypertension: New Advances and Future Impact. Circ. Res. 2021, 128, 803–807. [Google Scholar] [CrossRef] [PubMed]
- Carey, R.M.; Moran, A.E.; Whelton, P.K. Treatment of Hypertension: A Review. JAMA 2022, 328, 1849–1861. [Google Scholar] [CrossRef] [PubMed]
- Acelajado, M.C.; Hughes, Z.H.; Oparil, S.; Calhoun, D.A. Treatment of Resistant and Refractory Hypertension. Circ. Res. 2019, 124, 1061–1070. [Google Scholar] [CrossRef] [PubMed]
- Tatemoto, K.; Hosoya, M.; Habata, Y.; Fujii, R.; Kakegawa, T.; Zou, M.X.; Kawamata, Y.; Fukusumi, S.; Hinuma, S.; Kitada, C.; et al. Isolation and characterization of a novel endogenous peptide ligand for the human APJ receptor. Biochem. Biophys. Res. Commun. 1998, 251, 471–476. [Google Scholar] [CrossRef]
- O’Dowd, B.F.; Heiber, M.; Chan, A.; Heng, H.H.; Tsui, L.C.; Kennedy, J.L.; Shi, X.; Petronis, A.; George, S.R.; Nguyen, T. A human gene that shows identity with the gene encoding the angiotensin receptor is located on chromosome 11. Gene 1993, 136, 355–360. [Google Scholar] [CrossRef] [PubMed]
- Lee, D.K.; Cheng, R.; Nguyen, T.; Fan, T.; Kariyawasam, A.P.; Liu, Y.; Osmond, D.H.; George, S.R.; O’Dowd, B.F. Characterization of apelin, the ligand for the APJ receptor. J. Neurochem. 2000, 74, 34–41. [Google Scholar] [CrossRef] [PubMed]
- Sato, T.; Kadowaki, A.; Suzuki, T.; Ito, H.; Watanabe, H.; Imai, Y.; Kuba1, K. Loss of Apelin Augments Angiotensin II-Induced Cardiac Dysfunction and Pathological Remodeling. Int. J. Mol. Sci. 2019, 20, 239. [Google Scholar] [CrossRef]
- Wysocka, M.B.; Pietraszek-Gremplewicz, K.; Nowak, D. The role of apelin in cardiovascular diseases, obesity and cancer. Front. Physiol. 2018, 9, 557. [Google Scholar] [CrossRef]
- Katugampola, S.D.; Maguire, J.J.; Matthewson, S.R.; Davenport, A.P. [(125)I]-(Pyr(1))Apelin-13 is a novel radioligand for localizing the APJ orphan receptor in human and rat tissues with evidence for a vasoconstrictor role in man. Br. J. Pharmacol. 2001, 132, 1255–1260. [Google Scholar] [CrossRef]
- Maguire, J.J.; Kleinz, M.J.; Pitkin, S.L.; Davenport, A.P. [Pyr1]apelin-13 identified as the predominant apelin isoform in the human heart: Vasoactive mechanisms and inotropic action in disease. Hypertension 2009, 54, 598–604. [Google Scholar] [CrossRef] [PubMed]
- Mughal, A.; O’Rourke, S.T. Vascular effects of apelin: Mechanisms and therapeutic potential. Pharmacol. Ther. 2018, 190, 139–147. [Google Scholar] [CrossRef] [PubMed]
- Xie, F.; Liu, W.; Feng, F.; Li, X.; He, L.; Lv, D.; Qin, X.; Li, L.; Li, L.; Chen, L. Apelin-13 promotes cardiomyocyte hypertrophy via PI3K-Akt-ERK1/2-p70S6K and PI3K-induced autophagy. Acta Biochim. Biophys. Sin. 2015, 47, 969–980. [Google Scholar] [CrossRef] [PubMed]
- Besserer-Offroy, É.; Bérubé, P.; Côté, J.; Murza, A.; Longpré, J.M.; Dumaine, R.; Lesur, O.; Auger-Messier, M.; Leduc, R.; Marsault, É.; et al. The hypotensive effect of activated apelin receptor is correlated with β-arrestin recruitment. Pharmacol. Res. 2018, 131, 7–16. [Google Scholar] [CrossRef]
- Anto, S.; Sathish, V.; Sun, C.; O’Rourke, S.T. Apelin-Induced Relaxation of Coronary Arteries is Impaired in a Model of Second-Hand Cigarette Smoke Exposure. J. Cardiovasc. Pharmacol. 2022, 80, 842–851. [Google Scholar] [CrossRef]
- Scimia, M.C.; Hurtado, C.; Ray, S.; Metzler, S.; Wei, K.; Wang, J.; Woods, C.E.; Purcell, N.H.; Catalucci, D.; Akasaka, T.; et al. APJ acts as a dual receptor in cardiac hypertrophy. Nature 2012, 488, 394–398. [Google Scholar] [CrossRef]
- Ban, T.; Li, X.; Ma, X.; Yang, H.; Song, Y.; Sun, Y.; Shen, M.; Li, N.; Zhang, M.Y.; Ma, Y.; et al. GPCR structure and function relationship: Identification of a biased apelin receptor mutant. Biochem. J. 2018, 475, 3813–3826. [Google Scholar] [CrossRef]
- McAnally, D.; Siddiquee, K.; Sharir, H.; Qi, F.; Phatak, S.; Li, J.L.; Berg, E.; Fishman, J.; Smith, L. A Systematic Approach to Identify Biased Agonists of the Apelin Receptor through High-Throughput Screening. SLAS Discov. 2017, 22, 867–878. [Google Scholar] [CrossRef]
- Portilla-Martínez, A.; Ortiz-Flores, M.Á.; Meaney, E.; Villarreal, F.; Nájera, N.; Ceballos, G. (-)-Epicatechin Is a Biased Ligand of Apelin Receptor. Int. J. Mol. Sci. 2022, 23, 8962. [Google Scholar] [CrossRef]
- Yamaleyeva, L.M.; Brosnihan, K.B.; Elsangeedy, E.; McGee, C.; Shi, S.; Caudell, D.; Miller, C.; Varagic, J.; Bader, M.; Dechend, R.; et al. Systemic Outcomes of (Pyr1)-Apelin-13 Infusion at Mid-Late Pregnancy in a Rat Model with Preeclamptic Features. Sci. Rep. 2019, 9, 8579. [Google Scholar] [CrossRef]
- Liu, H.; Shi, Q.; Tang, L.; Wang, H.; Wang, D. Apelin-13 Ameliorates LPS-Induced Endothelial-to-Mesenchymal Transition and Post-Acute Lung Injury Pulmonary Fibrosis by Suppressing Transforming Growth Factor-B1 Signaling. Shock 2023, 59, 108–117. [Google Scholar] [CrossRef] [PubMed]
- Li, L.; Zeng, H.; Chen, J.X. Apelin-13 increases myocardial progenitor cells and improves repair postmyocardial infarction. Am. J. Physiol. Heart Circ. Physiol. 2012, 303, H605–H618. [Google Scholar] [CrossRef]
- Young, B.C.; Levine, R.J.; Karumanchi, S.A. Pathogenesis of preeclampsia. Annu. Rev. Pathol. 2010, 5, 173–192. [Google Scholar] [CrossRef] [PubMed]
- Kojima, H.; Tokunou, T.; Takahara, Y.; Sunagawa, K.; Hirooka, Y.; Ichiki, T.; Tsutsui, H. Hypoxia-inducible Factor-1 α Deletion in Myeloid Lineage Attenuates Hypoxia-induced Pulmonary Hypertension. Physiol. Rep. 2019, 7, e14025. [Google Scholar] [CrossRef] [PubMed]
- Hou, J.; Wang, L.; Long, H.; Wu, H.; Wu, Q.; Zhong, T.; Chen, X.; Zhou, C.; Guo, T.; Wang, T. Hypoxia Preconditioning Promotes Cardiac Stem Cell Survival and Cardiogenic Differentiation in vitro Involving Activation of the HIF-1α/apelin/APJ axis. Stem Cell Res. Ther. 2017, 8, 215. [Google Scholar] [CrossRef]
- Shah, A.K.; Bhullar, S.K.; Elimban, V.; Dhalla, N.S. Oxidative Stress as A Mechanism for Functional Alterations in Cardiac Hypertrophy and Heart Failure. Antioxidants 2021, 10, 931. [Google Scholar] [CrossRef]
- Zhang, Q.; Yao, F.; Raizada, M.K.; O’Rourke, S.T.; Sun, C. Apelin gene transfer into the rostral ventrolateral medulla induces chronic blood pressure elevation in normotensive rats. Circ. Res. 2009, 104, 1421–1428. [Google Scholar] [CrossRef]
- Guo, L.; Yin, A.; Zhang, Q.; Zhong, T.; O’Rourke, S.T.; Sun, C. Angiotensin-(1-7) attenuates angiotensin II-induced cardiac hypertrophy via a Sirt3-dependent mechanism. Am. J. Physiol. Heart Circ. Physiol. 2017, 312, H980–H991. [Google Scholar] [CrossRef]
- Yao, F.; Sun, C.; Chang, S.K. Lentil polyphenol extract prevents angiotensin II-induced hypertension, vascular remodeling and perivascular fibrosis. Food Funct. 2012, 3, 127–133. [Google Scholar] [CrossRef]





Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zhang, Q.; Shen, Y.; Niloy, S.I.; O'Rourke, S.T.; Sun, C. Chronic Effects of Apelin on Cardiovascular Regulation and Angiotensin II-Induced Hypertension. Pharmaceuticals 2023, 16, 600. https://doi.org/10.3390/ph16040600
Zhang Q, Shen Y, Niloy SI, O'Rourke ST, Sun C. Chronic Effects of Apelin on Cardiovascular Regulation and Angiotensin II-Induced Hypertension. Pharmaceuticals. 2023; 16(4):600. https://doi.org/10.3390/ph16040600
Chicago/Turabian StyleZhang, Qi, Yue Shen, Sayeman Islam Niloy, Stephen T. O'Rourke, and Chengwen Sun. 2023. "Chronic Effects of Apelin on Cardiovascular Regulation and Angiotensin II-Induced Hypertension" Pharmaceuticals 16, no. 4: 600. https://doi.org/10.3390/ph16040600
APA StyleZhang, Q., Shen, Y., Niloy, S. I., O'Rourke, S. T., & Sun, C. (2023). Chronic Effects of Apelin on Cardiovascular Regulation and Angiotensin II-Induced Hypertension. Pharmaceuticals, 16(4), 600. https://doi.org/10.3390/ph16040600