
A Novel PDK1/MEK Dual Inhibitor Induces Cytoprotective Autophagy via the PDK1/Akt Signaling Pathway in Non-Small Cell Lung Cancer

 ,
,
Abstract
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
1. Introduction
2. Results
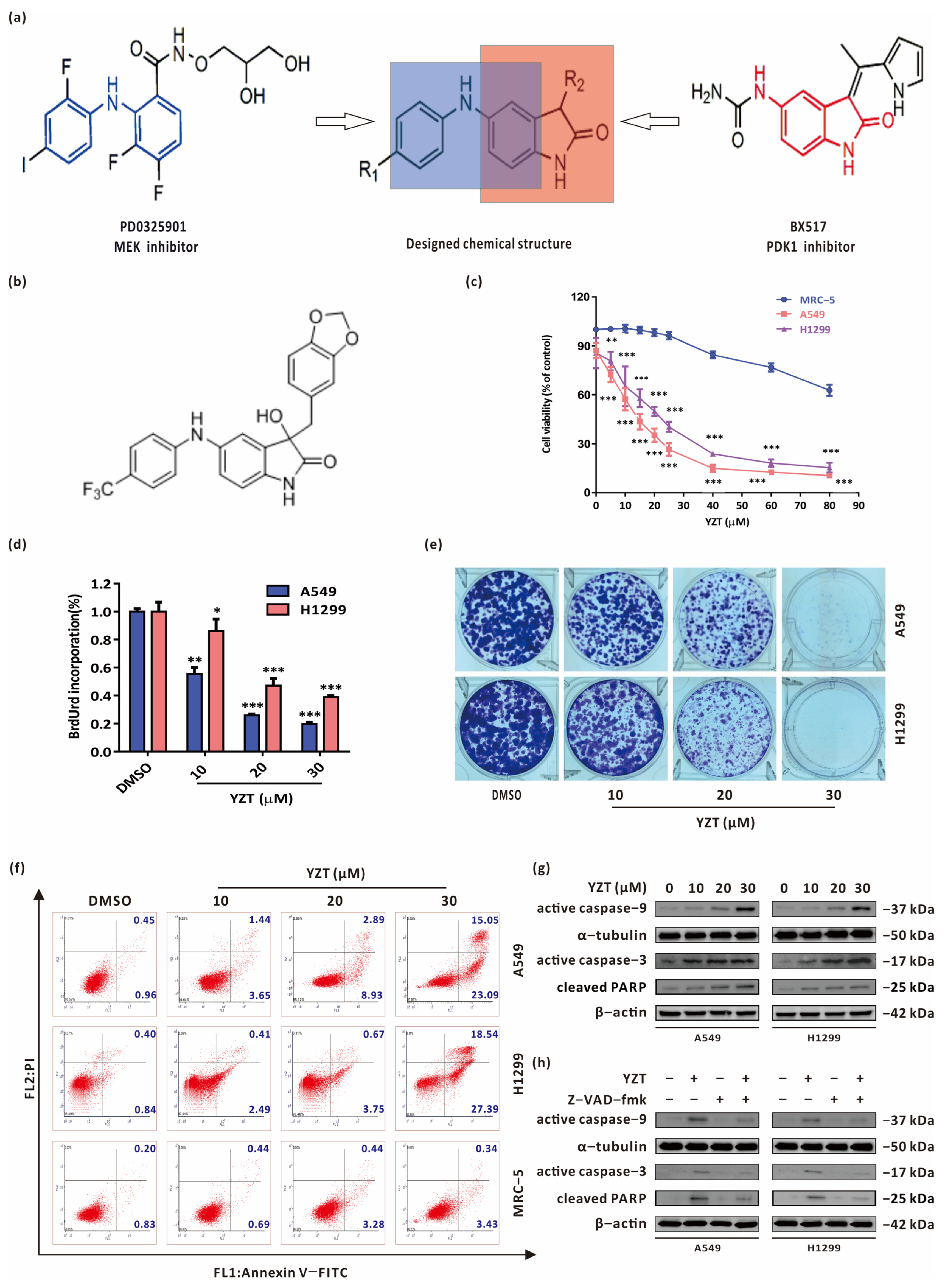
2.1. Compound YZT Suppresses NSCLC Cell Viability and Growth
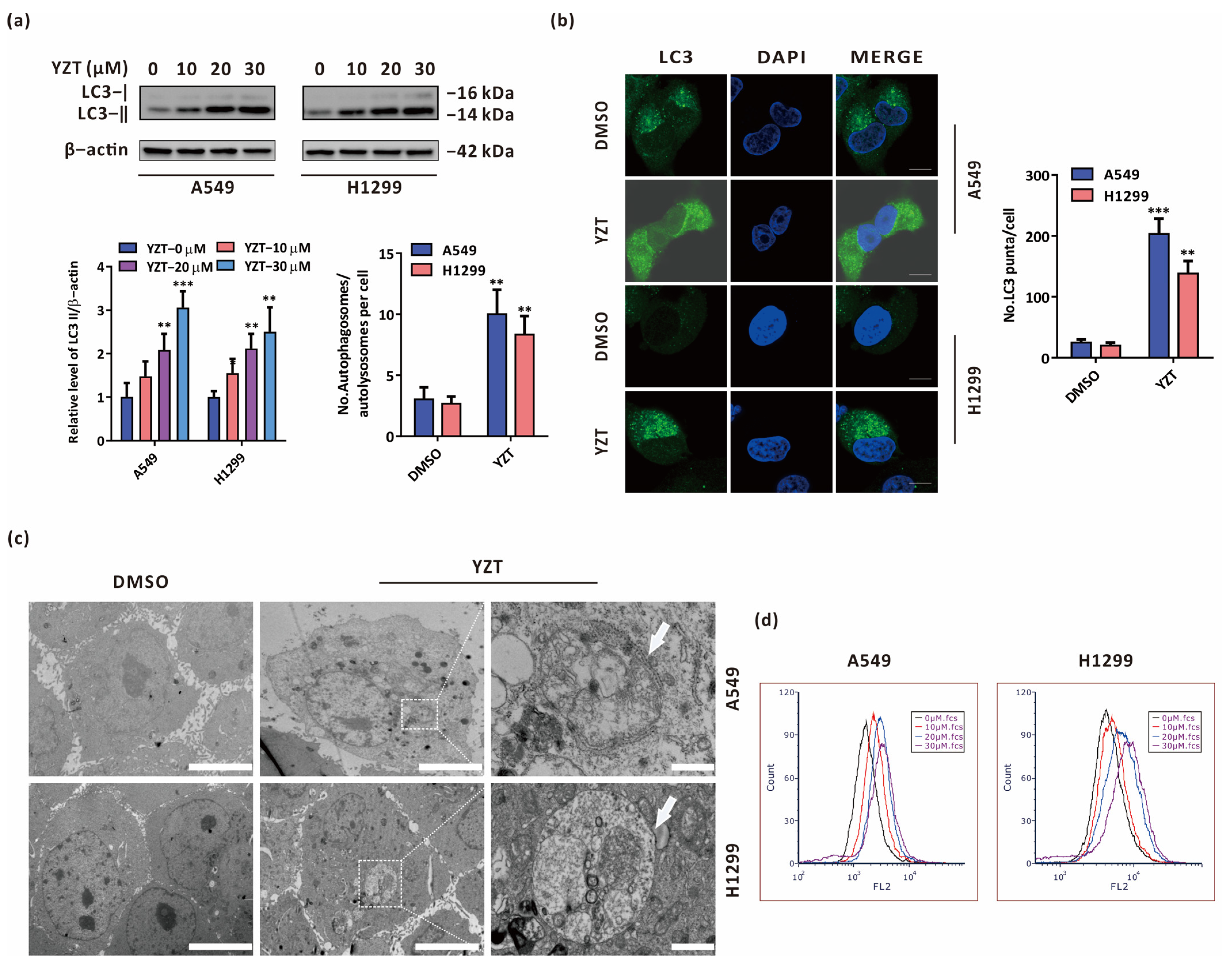
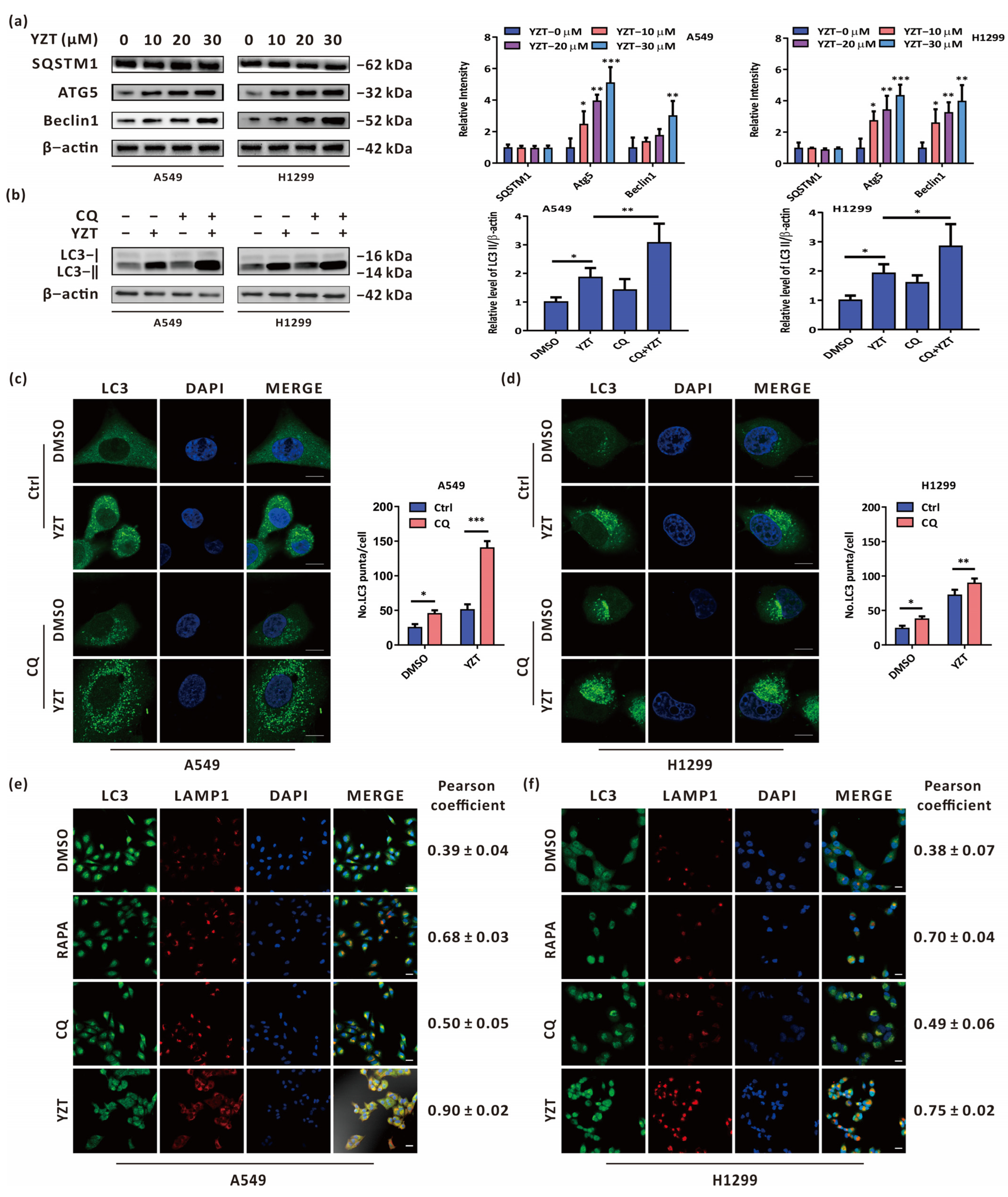
2.2. Compound YZT Provokes Autophagy in NSCLC Cells
2.3. Compound YZT Induces Intact Autophagic Flux
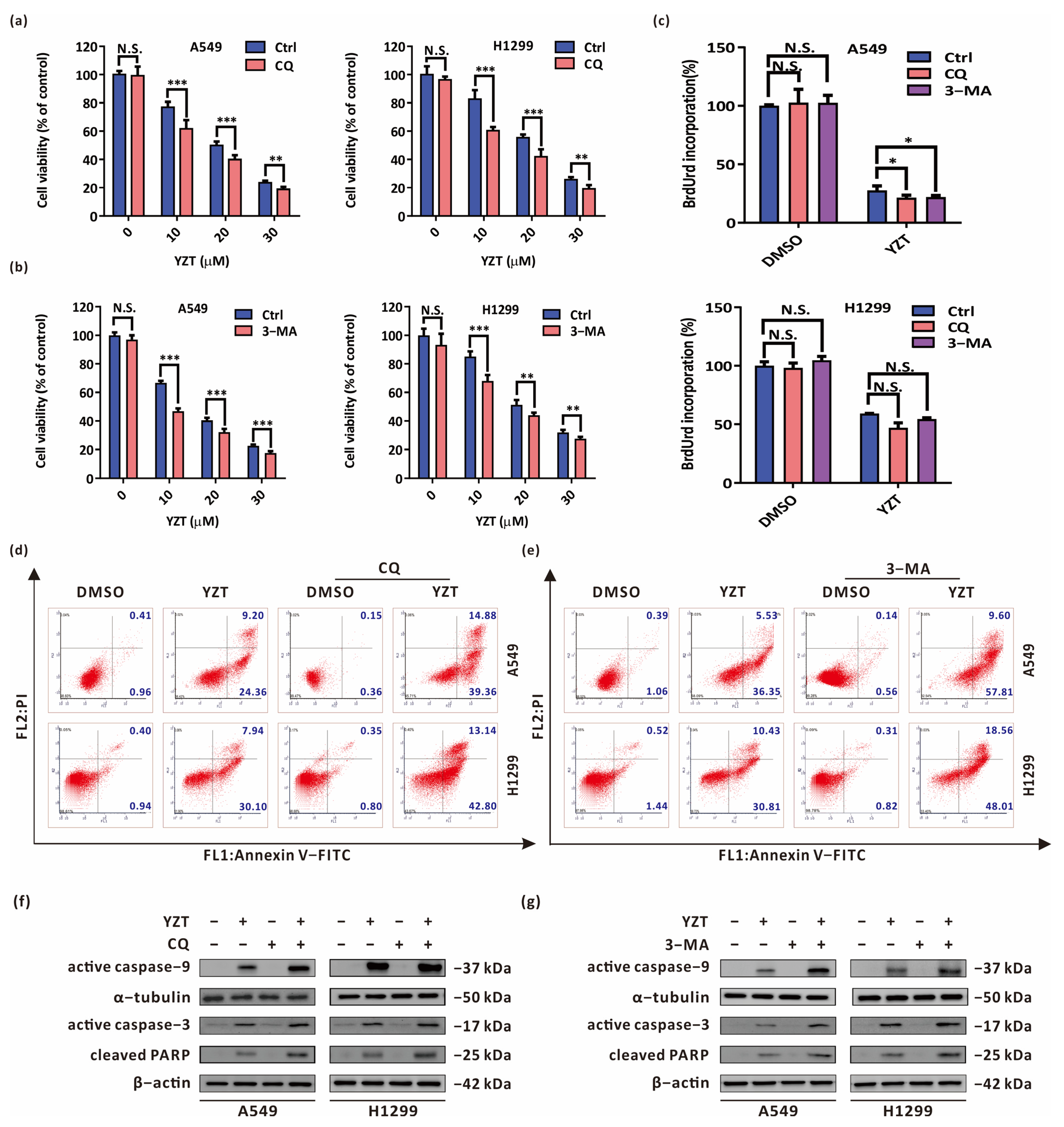
2.4. Autophagy Blockade Strengthens the Anticancer Activities of Compound YZT
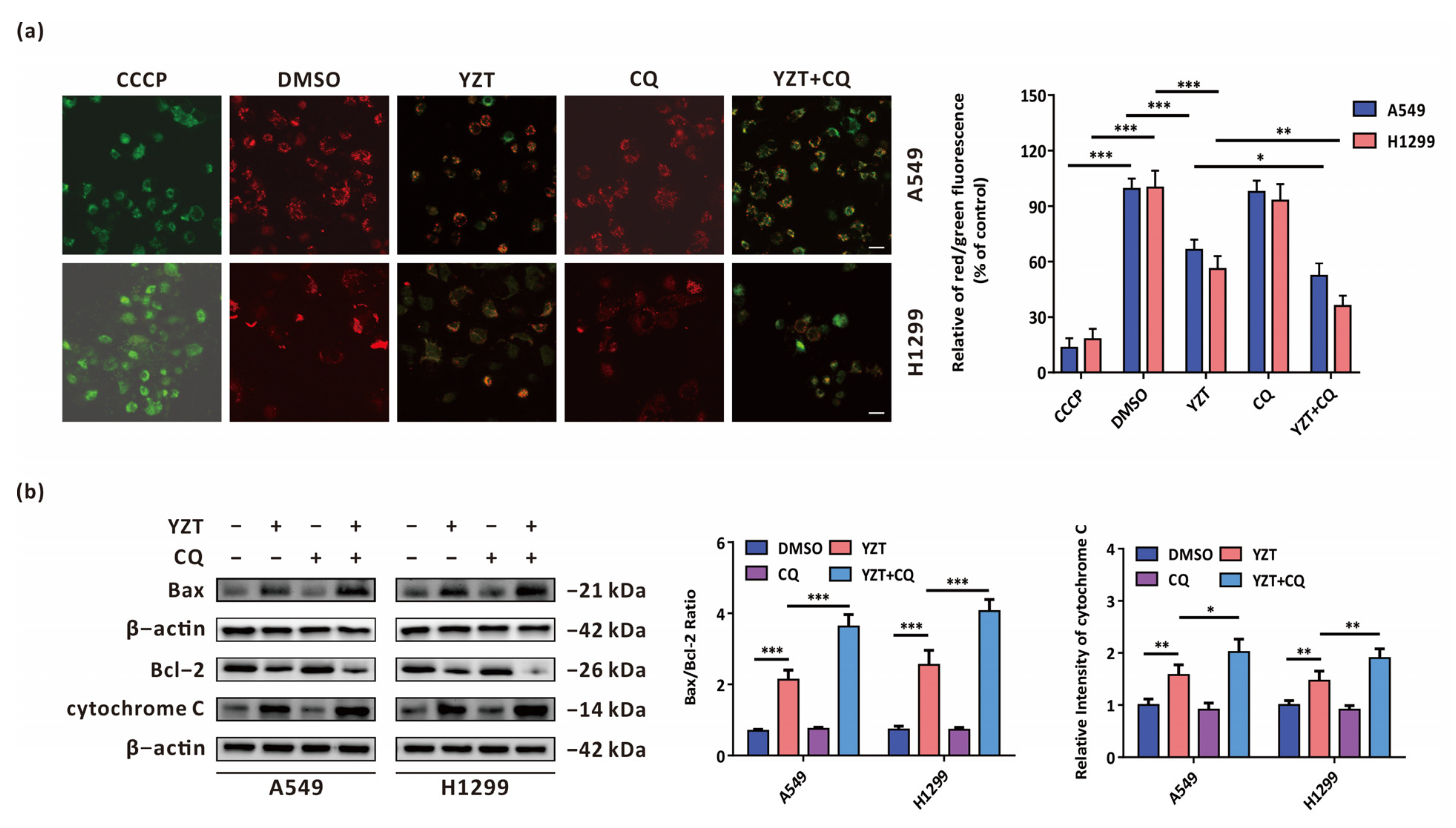
2.5. Autophagy Inhibition Exacerbates YZT-Mediated Mitochondrial Apoptosis
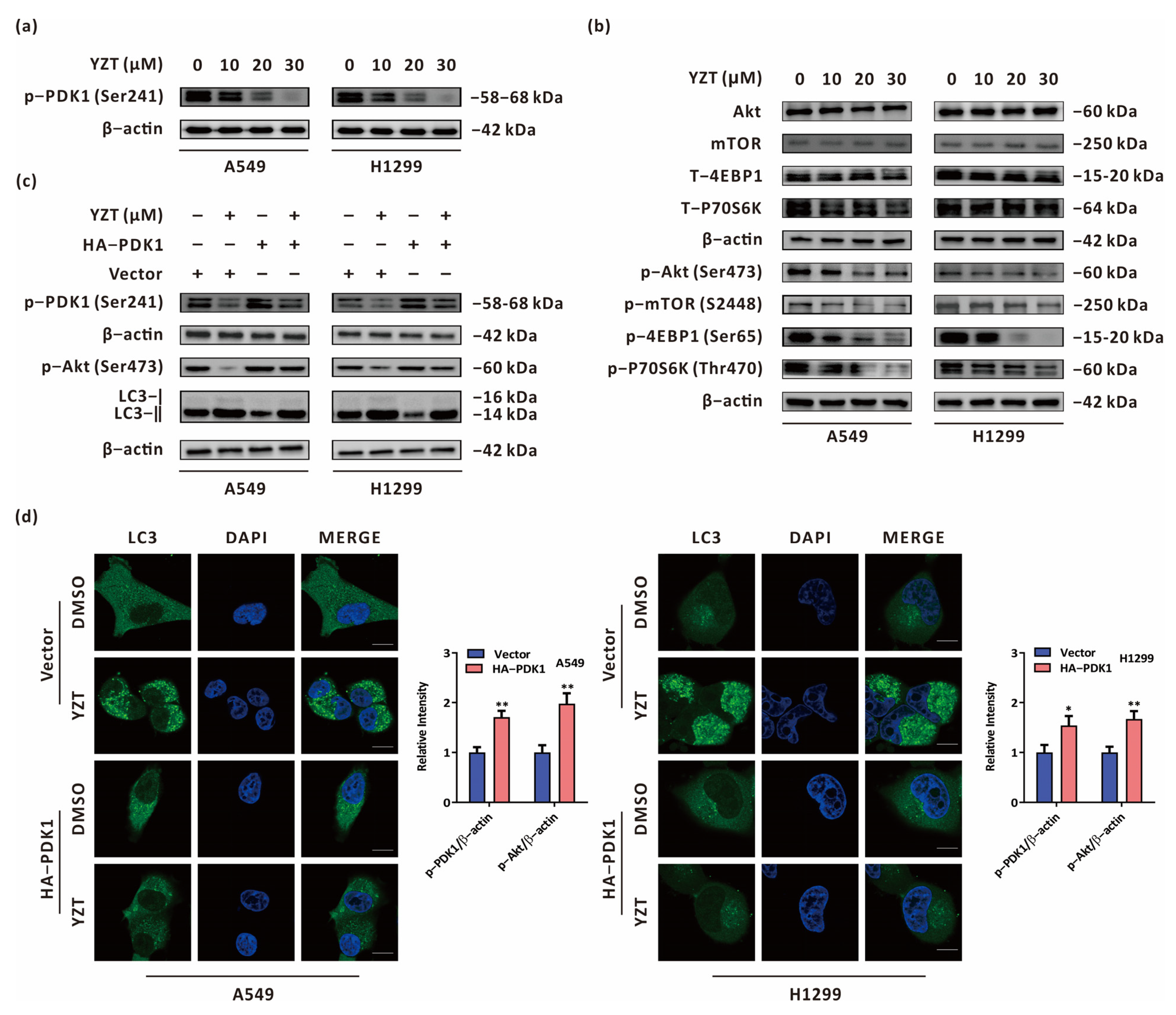
2.6. Compound YZT Initiates Autophagy through the PDK1/Akt Axis
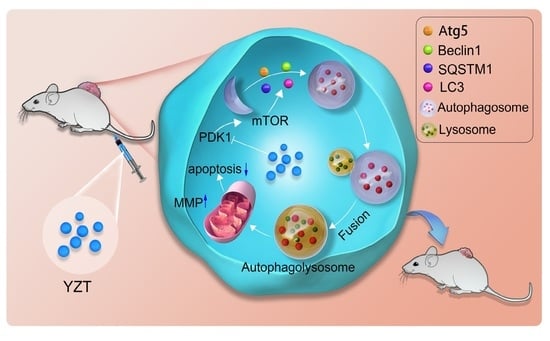
2.7. Combined Administration with Chloroquine Strengthens the Antitumor Activities of Compound YZT In Vivo
3. Discussion
4. Materials and Methods
4.1. Cell Preparation
4.2. Antibodies and Reagents
4.3. CCK-8
4.4. BrdU Labeling
4.5. Colony Formation
4.6. Annexin V-FITC/PI Apoptosis Detection
4.7. Western Blotting
4.8. Immunofluorescence
4.9. Transmission Electron Microscopy (TEM)
4.10. Acridine Orange (AO) Staining
4.11. JC-1 Staining Assay
4.12. Overexpression Plasmid Transfection
4.13. Pharmacodynamics Experiment In Vivo
4.14. Immunohistochemistry
4.15. Statistical Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Siegel, R.L.; Miller, K.D.; Fuchs, H.E.; Jemal, A. Cancer Statistics, 2021. CA Cancer J. Clin. 2021, 71, 7–33. [Google Scholar] [CrossRef] [PubMed]
- Oun, R.; Moussa, Y.E.; Wheate, N.J. The side effects of platinum-based chemotherapy drugs: A review for chemists. Dalton Trans. 2018, 47, 6645–6653. [Google Scholar] [CrossRef] [PubMed]
- Qu, G.P.; Shi, M.; Wang, D.; Wu, J.H.; Wang, P.; Gong, M.L.; Zhang, Z.J. Dual targeting of MEK and PI3K effectively controls the proliferation of human EGFR-TKI resistant non-small cell lung carcinoma cell lines with different genetic backgrounds. BMC Pulm. Med. 2021, 21, 208. [Google Scholar] [CrossRef] [PubMed]
- Marqués, M.; Tranchant, R.; Risa-Ebrí, B.; Suárez-Solís, M.L.; Fernández, L.C.; Carrillo-de-Santa-Pau, E.; Del Pozo, N.; Martínez de Villarreal, J.; Meiller, C.; Allory, Y.; et al. Combined MEK and PI3K/p110b inhibition as a novel targeted therapy for malignant mesothelioma displaying sarcomatoid features. Cancer Res. 2020, 80, 843–856. [Google Scholar] [CrossRef] [PubMed]
- Coussy, F.; El Botty, R.; Lavigne, M.; Gu, C.; Fuhrmann, L.; Briaux, A.; de Koning, L.; Dahmani, A.; Montaudon, E.; Morisset, L.; et al. Combination of PI3K and MEK inhibitors yields durable remission in PDX models of PIK3CA-mutated metaplastic breast cancers. J. Hematol. Oncol. 2020, 13, 13. [Google Scholar] [CrossRef] [PubMed]
- Yu, Z.; Chen, Z.; Su, Q.; Ye, S.; Yuan, H.; Kuai, M.; Lv, M.; Tu, Z.; Yang, X.; Liu, R.; et al. Dual inhibitors of RAF-MEK-ERK and PI3K-PDK1-AKT pathways: Design, synthesis and preliminary anticancer activity studies of 3-substituted-5-(phenylamino) indolone derivatives. Bioorg. Med. Chem. 2019, 27, 944–954. [Google Scholar] [CrossRef]
- Mizushima, N. Autophagy: Process and function. Genes Dev. 2007, 21, 2861–2873. [Google Scholar] [CrossRef]
- Kroemer, G.; Marino, G.; Levine, B. Autophagy and the integrated stress response. Mol. Cell 2010, 40, 280–293. [Google Scholar] [CrossRef]
- Sun, Y.; Huang, Y.H.; Huang, F.Y.; Mei, W.L.; Liu, Q.; Wang, C.C.; Lin, Y.Y.; Huang, C.; Li, Y.N.; Dai, H.F.; et al. 3′-epi-12beta-hydroxyfroside, a new cardenolide, induces cytoprotective autophagy via blocking the Hsp90/Akt/mTOR axis in lung cancer cells. Theranostics 2018, 8, 2044–2060. [Google Scholar] [CrossRef]
- Zhou, J.; Zhang, L.; Wang, M.; Zhou, L.; Feng, X.; Yu, L.; Lan, J.; Gao, W.; Zhang, C.; Bu, Y.; et al. CPX Targeting DJ-1 Triggers ROS-induced Cell Death and Protective Autophagy in Colorectal Cancer. Theranostics 2019, 9, 5577–5594. [Google Scholar] [CrossRef]
- Pellerito, C.; Emanuele, S.; Ferrante, F.; Celesia, A.; Giuliano, M.; Fiore, T. Tributyltin (IV) ferulate, a novel synthetic ferulic acid derivative, induces autophagic cell death in colon cancer cells: From chemical synthesis to biochemical effects. J. Inorg. Biochem. 2020, 205, 110999. [Google Scholar] [CrossRef] [PubMed]
- Sala de Oyanguren, F.J.; Rainey, N.E.; Moustapha, A.; Saric, A.; Sureau, F.; O’Connor, J.E.; Petit, P.X. Highlighting Curcumin-Induced Crosstalk between Autophagy and Apoptosis as Supported by Its Specific Subcellular Localization. Cells 2020, 9, 361. [Google Scholar] [CrossRef] [PubMed]
- Veeran, S.; Shu, B.; Cui, G.; Fu, S.; Zhong, G. Curcumin induces autophagic cell death in Spodoptera frugiperda cells. Pestic. Biochem. Physiol. 2017, 139, 79–86. [Google Scholar] [CrossRef] [PubMed]
- Zheng, X.; Li, W.; Xu, H.; Liu, J.; Ren, L.; Yang, Y.; Li, S.; Wang, J.; Ji, T.; Du, G. Sinomenine ester derivative inhibits glioblastoma by inducing mitochondria-dependent apoptosis and autophagy by PI3K/AKT/mTOR and AMPK/mTOR pathway. Acta Pharm. Sin. B 2021, 11, 3465–3480. [Google Scholar] [CrossRef]
- Jiang, J.; Zhang, L.; Chen, H.; Lei, Y.; Zhang, T.; Wang, Y.; Jin, P.; Lan, J.; Zhou, L.; Huang, Z.; et al. Regorafenib induces lethal autophagy arrest by stabilizing PSAT1 in glioblastoma. Autophagy 2020, 16, 106–122. [Google Scholar] [CrossRef]
- Zhou, L.; Gao, W.; Wang, K.; Huang, Z.; Zhang, L.; Zhang, Z.; Zhou, J.; Nice, E.C.; Huang, C. Brefeldin A inhibits colorectal cancer growth by triggering Bip/Akt-regulated autophagy. FASEB J. 2019, 33, 5520–5534. [Google Scholar] [CrossRef]
- Jin, P.; Jiang, J.; Xie, N.; Zhou, L.; Huang, Z.; Zhang, L.; Qin, S.; Fu, S.; Peng, L.; Gao, W.; et al. MCT1 relieves osimertinib-induced CRC suppression by promoting autophagy through the LKB1/AMPK signaling. Cell Death Dis. 2019, 10, 615. [Google Scholar] [CrossRef]
- Liu, R.; Yu, Z.; Chen, Z.; Liu, D.; Huang, F.; Li, Q.; Hu, G.; Yi, X.; Li, X.; Zhou, H.; et al. A novel dual MEK/PDK1 inhibitor 9za retards the cell cycle at G0/G1 phase and induces mitochondrial apoptosis in non-small cell lung cancer cells. PeerJ 2020, 8, e9981. [Google Scholar] [CrossRef]
- Guo, L.; Gong, H.; Tang, T.L.; Zhang, B.K.; Zhang, L.Y.; Yan, M. Crizotinib and Sunitinib Induce Hepatotoxicity and Mitochondrial Apoptosis in L02 Cells via ROS and Nrf2 Signaling Pathway. Front. Pharmacol. 2021, 12, 620934. [Google Scholar] [CrossRef]
- Zhou, J.; Jiang, Y.Y.; Chen, H.; Wu, Y.C.; Zhang, L. Tanshinone I attenuates the malignant biological properties of ovarian cancer by inducing apoptosis and autophagy via the inactivation of PI3K/AKT/mTOR pathway. Cell Prolif. 2020, 53, e12739. [Google Scholar] [CrossRef]
- Fang, S.; Wan, X.; Zou, X.; Sun, S.; Hao, X.; Liang, C.; Zhang, Z.; Zhang, F.; Sun, B.; Li, H.; et al. Arsenic trioxide induces macrophage autophagy and atheroprotection by regulating ROS-dependent TFEB nuclear translocation and AKT/mTOR pathway. Cell Death Dis. 2021, 12, 88. [Google Scholar] [CrossRef]
- Paulo, J.A.; McAllister, F.E.; Everley, R.A.; Beausoleil, S.A.; Banks, A.S.; Gygi, S.P. Effects of MEK inhibitors GSK1120212 and PD0325901 in vivo using 10-plex quantitative proteomics and phosphoproteomics. Proteomics 2015, 15, 462–473. [Google Scholar] [CrossRef]
- Akula, S.M.; Abrams, S.L.; Steelman, L.S.; Emma, M.R.; Augello, G.; Cusimano, A.; Azzolina, A.; Montalto, G.; Cervello, M.; McCubrey, J.A. RAS/RAF/MEK/ERK, PI3K/PTEN/AKT/mTORC1 and TP53 pathways and regulatory miRs as therapeutic targets in hepatocellular carcinoma. Expert Opin. Ther. Targets 2019, 23, 915–929. [Google Scholar] [CrossRef] [PubMed]
- Mendoza, M.C.; Er, E.E.; Blenis, J. The Ras-ERK and PI3K-mTOR pathways: Cross-talk and compensation. Trends Biochem. Sci. 2011, 36, 320–328. [Google Scholar] [CrossRef] [PubMed]
- Temraz, S.; Mukherji, D.; Shamseddine, A. Dual Inhibition of MEK and PI3K Pathway in KRAS and BRAF Mutated Colorectal Cancers. Int. J. Mol. Sci. 2015, 16, 22976–22988. [Google Scholar] [CrossRef] [PubMed]
- Gewirtz, D.A. The four faces of autophagy: Implications for cancer therapy. Cancer Res. 2014, 74, 647–651. [Google Scholar] [CrossRef]
- Dou, Q.; Chen, H.N.; Wang, K.; Yuan, K.; Lei, Y.; Li, K.; Lan, J.; Chen, Y.; Huang, Z.; Xie, N.; et al. Ivermectin Induces Cytostatic Autophagy by Blocking the PAK1/Akt Axis in Breast Cancer. Cancer Res. 2016, 76, 4457–4469. [Google Scholar] [CrossRef]
- Maiuri, M.C.; Zalckvar, E.; Kimchi, A.; Kroemer, G. Self-eating and self-killing: Crosstalk between autophagy and apoptosis. Nat. Rev. Mol. Cell Biol. 2007, 8, 741–752. [Google Scholar] [CrossRef]
- Thorburn, J.; Andrysik, Z.; Staskiewicz, L.; Gump, J.; Maycotte, P.; Oberst, A.; Green, D.R.; Espinosa, J.M.; Thorburn, A. Autophagy controls the kinetics and extent of mitochondrial apoptosis by regulating PUMA levels. Cell Rep. 2014, 7, 45–52. [Google Scholar] [CrossRef]







Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Liu, R.; Chen, Z.; Hu, G.; Yu, Z.; Li, Q.; Liu, D.; Li, L.; Liu, Z. A Novel PDK1/MEK Dual Inhibitor Induces Cytoprotective Autophagy via the PDK1/Akt Signaling Pathway in Non-Small Cell Lung Cancer. Pharmaceuticals 2023, 16, 244. https://doi.org/10.3390/ph16020244
Liu R, Chen Z, Hu G, Yu Z, Li Q, Liu D, Li L, Liu Z. A Novel PDK1/MEK Dual Inhibitor Induces Cytoprotective Autophagy via the PDK1/Akt Signaling Pathway in Non-Small Cell Lung Cancer. Pharmaceuticals. 2023; 16(2):244. https://doi.org/10.3390/ph16020244
Chicago/Turabian StyleLiu, Rangru, Zhuo Chen, Gaoyun Hu, Zutao Yu, Qianbin Li, Danqi Liu, Ling Li, and Zhaoqian Liu. 2023. "A Novel PDK1/MEK Dual Inhibitor Induces Cytoprotective Autophagy via the PDK1/Akt Signaling Pathway in Non-Small Cell Lung Cancer" Pharmaceuticals 16, no. 2: 244. https://doi.org/10.3390/ph16020244
APA StyleLiu, R., Chen, Z., Hu, G., Yu, Z., Li, Q., Liu, D., Li, L., & Liu, Z. (2023). A Novel PDK1/MEK Dual Inhibitor Induces Cytoprotective Autophagy via the PDK1/Akt Signaling Pathway in Non-Small Cell Lung Cancer. Pharmaceuticals, 16(2), 244. https://doi.org/10.3390/ph16020244