
DP2, a Carbohydrate Derivative, Enhances In Vitro Osteoblast Mineralisation

 , , , , ,
, , , , ,
Abstract
:1. Introduction
2. Results
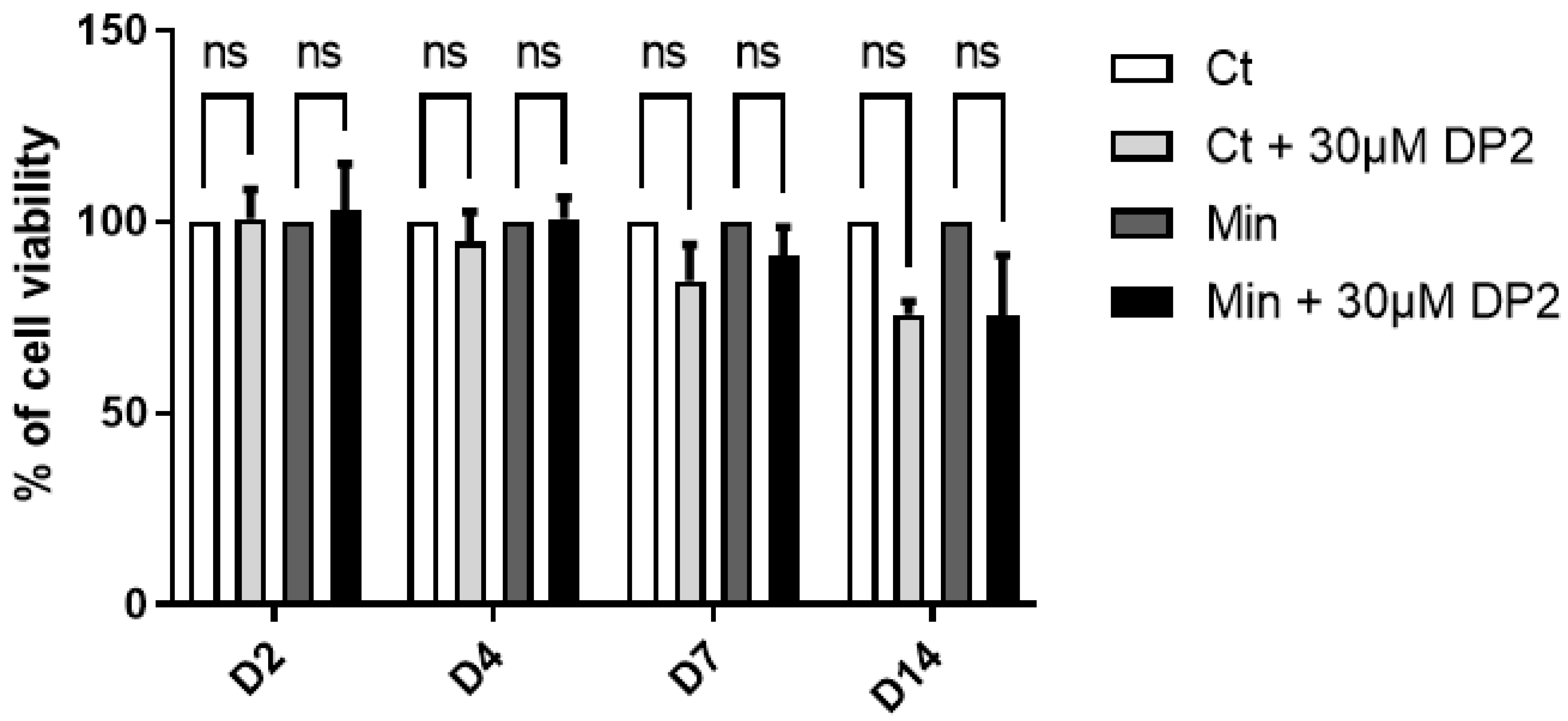
2.1. DP2 Does Not Affect the Viability of HOb Cells
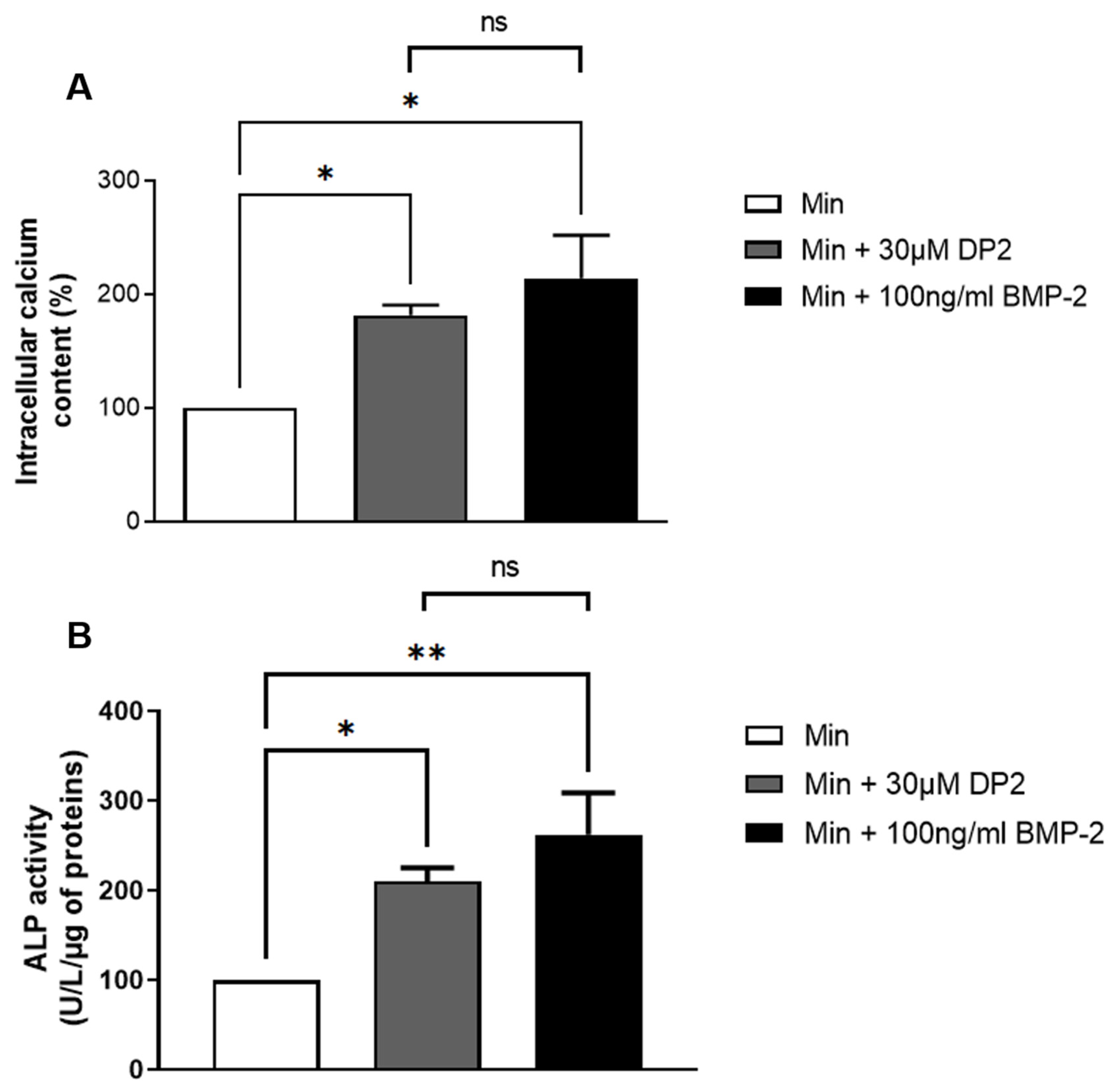
2.2. DP2 Promotes Calcification and Induces Alkaline Phosphatase (ALP) Activity in HOb Cells
2.3. DP2 Induces Calcification Marker Expression Earlier Than BMP-2
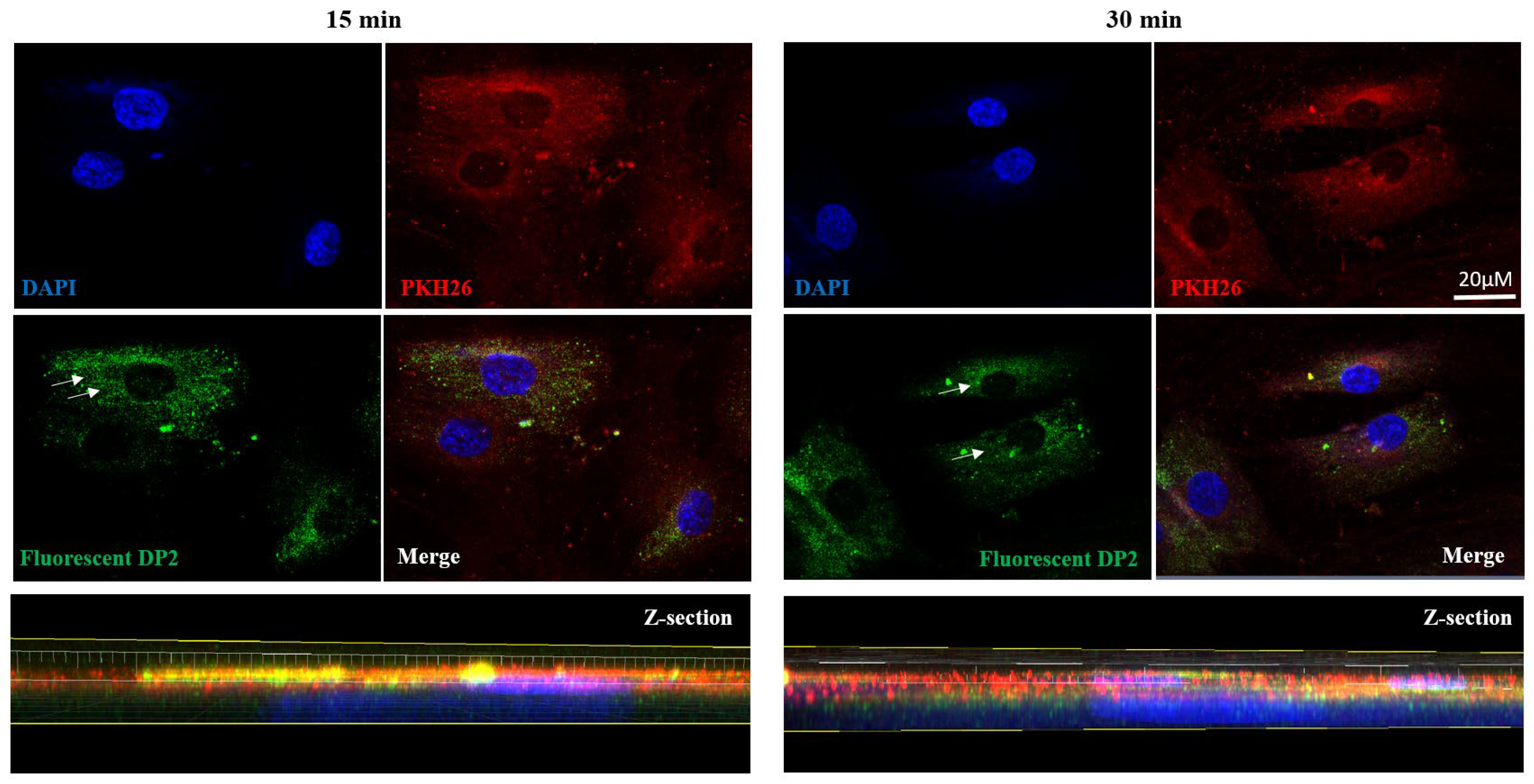
2.4. DP2 Is Internalized in HOb Cells
2.5. DP2-Induced Calcification Could Be Mediated at Least in Part by PKC Signalling
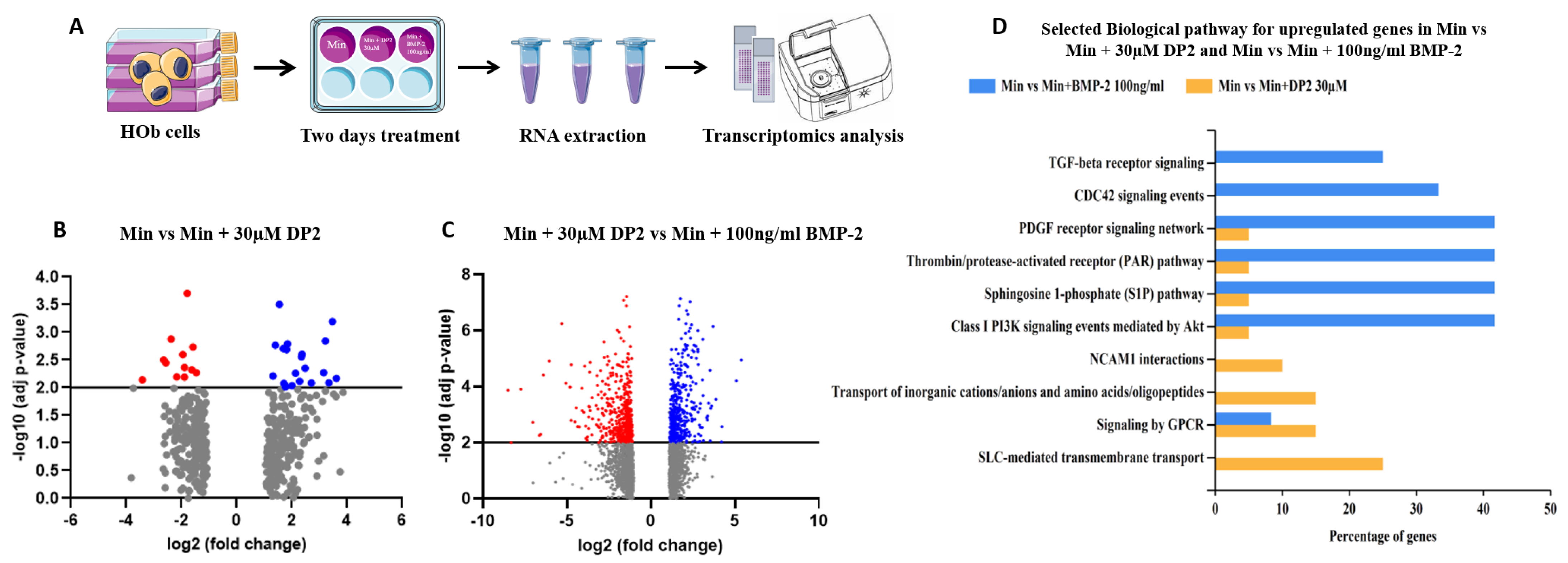
2.6. Signalling Pathways Implicated in DP2-Induced Mineralisation
3. Discussion
4. Materials and Methods
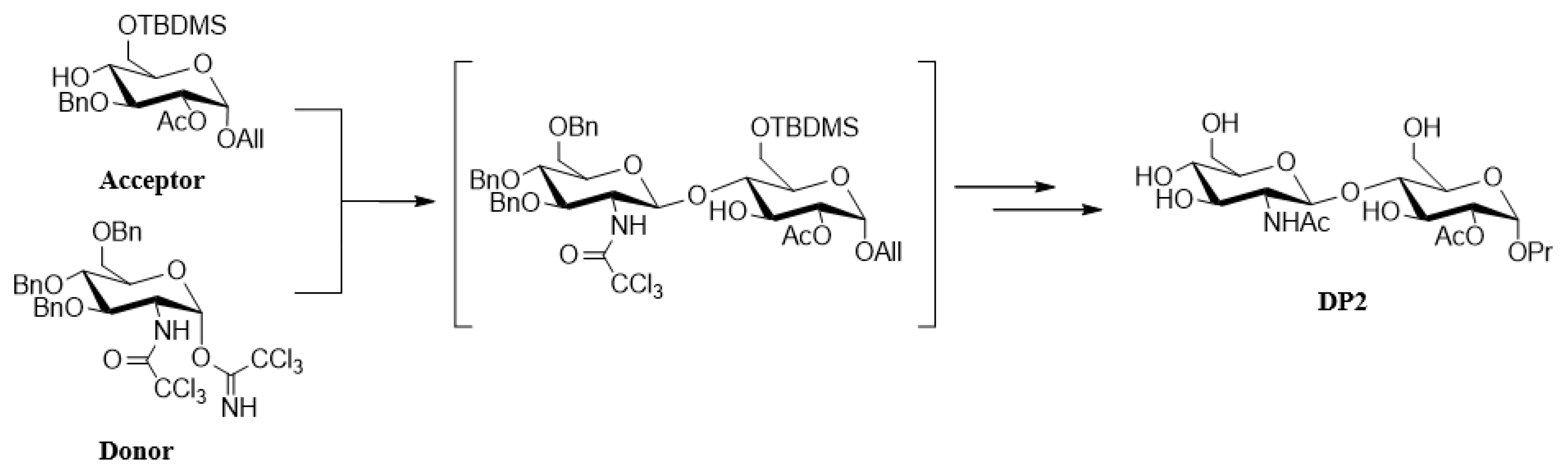
4.1. DP2 Disaccharide Synthesis
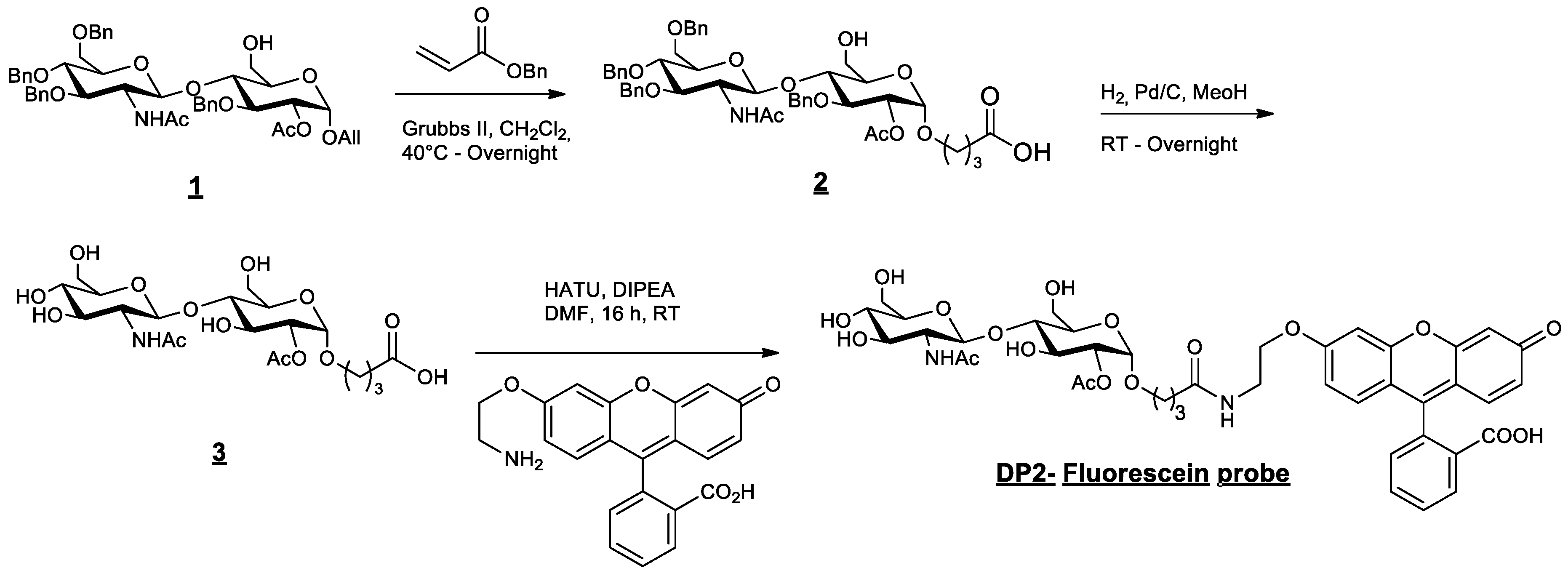
4.2. Fluorescent DP2 Synthesis
4.3. Cell Culture
4.4. Cell Viability Assay
4.5. Calcification Assay
4.6. Alkaline Phosphatase Activity
4.7. Real-Time PCR Analysis
4.8. Transcriptomic Analysis
4.9. Western Blot
4.10. Fluorescence Microscopy
4.11. Statistical Analysis
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Kostenuik, P.; Mirza, F.M. Fracture Healing Physiology and the Quest for Therapies for Delayed Healing and Nonunion. J. Orthop. Res. 2017, 35, 213–223. [Google Scholar] [CrossRef] [PubMed]
- Kanis, J.A.; Norton, N.; Harvey, N.C.; Jacobson, T.; Johansson, H.; Lorentzon, M.; McCloskey, E.V.; Willers, C.; Borgström, F. SCOPE 2021: A New Scorecard for Osteoporosis in Europe. Arch. Osteoporos. 2021, 16, 82. [Google Scholar] [CrossRef] [PubMed]
- Willers, C.; Norton, N.; Harvey, N.C.; Jacobson, T.; Johansson, H.; Lorentzon, M.; McCloskey, E.V.; Borgström, F.; Kanis, J.A. The SCOPE review panel of the IOF Osteoporosis in Europe: A Compendium of Country-Specific Reports. Arch. Osteoporos. 2022, 17, 23. [Google Scholar] [CrossRef]
- Wu, A.C.; Raggatt, L.J.; Alexander, K.A.; Pettit, A.R. Unraveling Macrophage Contributions to Bone Repair. Bonekey Rep. 2013, 2, 373. [Google Scholar] [CrossRef] [PubMed]
- Marsell, R.; Einhorn, T.A. The Biology of Fracture Healing. Injury 2011, 42, 551–555. [Google Scholar] [CrossRef]
- Schell, H.; Duda, G.N.; Peters, A.; Tsitsilonis, S.; Johnson, K.A.; Schmidt-Bleek, K. The Haematoma and Its Role in Bone Healing. J. Exp. Orthop. 2017, 4, 5. [Google Scholar] [CrossRef]
- Gerstenfeld, L.C.; Cullinane, D.M.; Barnes, G.L.; Graves, D.T.; Einhorn, T.A. Fracture Healing as a Post-Natal Developmental Process: Molecular, Spatial, and Temporal Aspects of Its Regulation. J. Cell. Biochem. 2003, 88, 873–884. [Google Scholar] [CrossRef]
- Campana, V.; Milano, G.; Pagano, E.; Barba, M.; Cicione, C.; Salonna, G.; Lattanzi, W.; Logroscino, G. Bone Substitutes in Orthopaedic Surgery: From Basic Science to Clinical Practice. J. Mat. Sci. Mater. Med. 2014, 25, 2445–2461. [Google Scholar] [CrossRef]
- Perez, J.R.; Kouroupis, D.; Li, D.J.; Best, T.M.; Kaplan, L.; Correa, D. Tissue Engineering and Cell-Based Therapies for Fractures and Bone Defects. Front. Bioeng. Biotechnol. 2018, 6, 105. [Google Scholar] [CrossRef]
- Wang, W.; Yeung, K.W.K. Bone Grafts and Biomaterials Substitutes for Bone Defect Repair: A Review. Bioact. Mater. 2017, 2, 224–247. [Google Scholar] [CrossRef]
- Oliveira, É.R.; Nie, L.; Podstawczyk, D.; Allahbakhsh, A.; Ratnayake, J.; Brasil, D.L.; Shavandi, A. Advances in Growth Factor Delivery for Bone Tissue Engineering. Int. J. Mol. Sci. 2021, 22, 903. [Google Scholar] [CrossRef] [PubMed]
- Ducy, P.; Karsenty, G. The Family of Bone Morphogenetic Proteins. Kidney Int. 2000, 57, 2207–2214. [Google Scholar] [CrossRef] [PubMed]
- Dumic-Cule, I.; Peric, M.; Kucko, L.; Grgurevic, L.; Pecina, M.; Vukicevic, S. Bone Morphogenetic Proteins in Fracture Repair. Inter. Orthop. 2018, 42, 2619–2626. [Google Scholar] [CrossRef] [PubMed]
- Fischerauer, E.E.; Manninger, M.; Seles, M.; Janezic, G.; Pichler, K.; Ebner, B.; Weinberg, A.M. BMP-6 and BMPR-1a Are up-Regulated in the Growth Plate of the Fractured Tibia. J. Orthop. Res. 2013, 31, 357–363. [Google Scholar] [CrossRef] [PubMed]
- Sheikh, Z.; Javaid, M.A.; Hamdan, N.; Hashmi, R. Bone Regeneration Using Bone Morphogenetic Proteins and Various Biomaterial Carriers. Materials 2015, 8, 1778–1816. [Google Scholar] [CrossRef]
- Cahill, K.S.; McCormick, P.C.; Levi, A.D. A Comprehensive Assessment of the Risk of Bone Morphogenetic Protein Use in Spinal Fusion Surgery and Postoperative Cancer Diagnosis. J. Neurosurg. Spine 2015, 23, 86–93. [Google Scholar] [CrossRef]
- Carragee, E.J.; Hurwitz, E.L.; Weiner, B.K. A Critical Review of Recombinant Human Bone Morphogenetic Protein-2 Trials in Spinal Surgery: Emerging Safety Concerns and Lessons Learned. Spine J. 2011, 11, 471–491. [Google Scholar] [CrossRef]
- Lee, J.W.; Lee, S.; Lee, S.H.; Yang, H.S.; Im, G.-I.; Kim, C.-S.; Park, J.-H.; Kim, B.S. Improved Spinal Fusion Efficacy by Long-Term Delivery of Bone Morphogenetic Protein-2 in a Rabbit Model. Acta. Orthop. 2011, 82, 756–760. [Google Scholar] [CrossRef]
- Fu, R.; Selph, S.; McDonagh, M.; Peterson, K.; Tiwari, A.; Chou, R.; Helfand, M. Effectiveness and Harms of Recombinant Human Bone Morphogenetic Protein-2 in Spine Fusion. Ann. Intern. Med. 2013, 158, 890–902. [Google Scholar] [CrossRef]
- Nguyen, V.; Meyers, C.A.; Yan, N.; Agarwal, S.; Levi, B.; James, A.W. BMP-2-Induced Bone Formation and Neural Inflammation. J. Orthop. 2017, 14, 252–256. [Google Scholar] [CrossRef]
- Tannoury, C.A.; An, H.S. Complications with the Use of Bone Morphogenetic Protein 2 (BMP-2) in Spine Surgery. Spine J. 2014, 14, 552–559. [Google Scholar] [CrossRef] [PubMed]
- Suzuki, M.F.; Oliveira, J.E.; Damiani, R.; Lima, E.R.; Amaral, K.C.; Santos, A.M.d.S.; Magalhães, G.S.; Faverani, L.P.; Pereira, L.A.V.D.; Silva, F.M.; et al. Human Bone Morphogenetic Protein-2 (HBMP-2) Characterization by Physical–Chemical, Immunological and Biological Assays. AMB Express 2020, 10, 34. [Google Scholar] [CrossRef] [PubMed]
- Dussouy, C.; Bultel, L.; Saguez, J.; Cherqui, A.; Khelifa, M.; Grand, E.; Giordanengo, P.; Kovensky, J. Strong Aphicidal Activity of GlcNAc(Β1→4)Glc Disaccharides: Synthesis, Physiological Effects, and Chitinase Inhibition. Chem. Eur. J. 2012, 18, 10021–10028. [Google Scholar] [CrossRef] [PubMed]
- Churchill, D.G.; Sikirić, M.D.; Čolović, B.; Milhofer, H.F. (Eds.) Pinki Dey Bone Mineralisation. In Contemporary Topics about Phosphorus in Biology and Materials; IntechOpen: Rijeka, Croatia, 2020; Chapter 3; ISBN 978-1-78985-040-6. [Google Scholar]
- Zamurovic, N.; Cappellen, D.; Rohner, D.; Susa, M. Coordinated Activation of Notch, Wnt, and Transforming Growth Factor-β Signaling Pathways in Bone Morphogenic Protein 2-Induced Osteogenesis: Notch Target Gene Hey1 Inhibits Mineralization and Runx2 Transcriptional Activity. J. Biol. Chem. 2004, 279, 37704–37715. [Google Scholar] [CrossRef]
- Suzuki, A.; Ghayor, C.; Guicheux, J.; Magne, D.; Quillard, S.; Kakita, A.; Ono, Y.; Miura, Y.; Oiso, Y.; Itoh, M.; et al. Enhanced Expression of the Inorganic Phosphate Transporter Pit-1 Is Involved in BMP-2–Induced Matrix Mineralization in Osteoblast-Like Cells. J. Bone Miner. Rese. 2006, 21, 674–683. [Google Scholar] [CrossRef]
- Fujioka-Kobayashi, M.; Abd El Raouf, M.; Saulacic, N.; Kobayashi, E.; Zhang, Y.; Schaller, B.; Miron, R.J. Superior Bone-Inducing Potential of RhBMP9 Compared to RhBMP2. J. Biomed. Mater. Rese. A 2018, 106, 1561–1574. [Google Scholar] [CrossRef]
- O’Neill, W.C. Pyrophosphate, Alkaline Phosphatase, and Vascular Calcification. Circ. Res. 2006, 99, e2. [Google Scholar] [CrossRef]
- Khotib, J.; Gani, M.A.; Budiatin, A.S.; Lestari, M.L.; Rahadiansyah, E.; Ardianto, C. Signaling Pathway and Transcriptional Regulation in Osteoblasts during Bone Healing: Direct Involvement of Hydroxyapatite as a Biomaterial. Pharmaceuticals 2021, 14, 615. [Google Scholar] [CrossRef]
- Kazarian, A.A.; Smith, J.A.; Hilder, E.F.; Breadmore, M.C.; Quirino, J.P.; Suttil, J. Development of a Novel Fluorescent Tag O-2-[Aminoethyl]Fluorescein for the Electrophoretic Separation of Oligosaccharides. Anal. Chim. Acta 2010, 662, 206–213. [Google Scholar] [CrossRef]
- Vimalraj, S. Alkaline Phosphatase: Structure, Expression and Its Function in Bone Mineralization. Gene 2020, 754, 144855. [Google Scholar] [CrossRef]
- Kim, Y.-J.; Lee, M.-H.; Wozney, J.M.; Cho, J.-Y.; Ryoo, H.-M. Bone Morphogenetic Protein-2-Induced Alkaline Phosphatase Expression Is Stimulated by Dlx5 and Repressed by Msx2. J. Biol. Chem. 2004, 279, 50773–50780. [Google Scholar] [CrossRef] [PubMed]
- Rodríguez-Carballo, E.; Gámez, B.; Ventura, F. P38 MAPK Signaling in Osteoblast Differentiation. Front. Cell Dev. Biol. 2016, 4, 40. [Google Scholar] [CrossRef] [PubMed]
- Delannoy, P.; Lemonnier, J.; Haÿ, E.; Modrowski, D.; Marie, P.J. Protein Kinase C-Dependent Upregulation of N-Cadherin Expression by Phorbol Ester in Human Calvaria Osteoblasts. Exp. Cell Res. 2001, 269, 154–161. [Google Scholar] [CrossRef] [PubMed]
- Lampasso, J.D.; Chen, W.; Marzec, N. The Expression Profile of PKC Isoforms during MC3T3-E1 Differentiation. Int. J. Mol. Med. 2006, 17, 1125–1131. [Google Scholar]
- Richardson, J.A.; Amantea, C.M.; Kianmahd, B.; Tetradis, S.; Lieberman, J.R.; Hahn, T.J.; Parhami, F. Oxysterol-Induced Osteoblastic Differentiation of Pluripotent Mesenchymal Cells Is Mediated through a PKC- and PKA-Dependent Pathway. J. Cell. Biochem. 2007, 100, 1131–1145. [Google Scholar] [CrossRef]
- Jeong, H.M.; Jin, Y.-H.; Choi, Y.H.; Yum, J.; Choi, J.-K.; Yeo, C.-Y.; Lee, K.-Y. PKC Signaling Inhibits Osteogenic Differentiation through the Regulation of Msx2 Function. Biochimi. Biophys. Acta BBA Mol. Cell Res. 2012, 1823, 1225–1232. [Google Scholar] [CrossRef]
- Cheng, B.-F.; Feng, X.; Gao, Y.-X.; Jian, S.-Q.; Liu, S.-R.; Wang, M.; Xie, Y.-F.; Wang, L.; Feng, Z.-W.; Yang, H.-J. Neural Cell Adhesion Molecule Regulates Osteoblastic Differentiation Through Wnt/β-Catenin and PI3K-Akt Signaling Pathways in MC3T3-E1 Cells. Front. Endocrinol. 2021, 12, 657953. [Google Scholar] [CrossRef]
- Park, K.H.; Han, D.I.; Rhee, Y.-H.; Jeong, S.-J.; Kim, S.-H.; Park, Y.G. Protein Kinase C ΒII and δ/θ Play Critical Roles in Bone Morphogenic Protein-4-Stimulated Osteoblastic Differentiation of MC3T3-E1 Cells. Biochemi. Biophys. Rese. Commun. 2010, 403, 7–12. [Google Scholar] [CrossRef]
- Sarkar, B.C.; Chauhan, U.P.S. A New Method for Determining Micro Quantities of Calcium in Biological Materials. Anal. Biochem. 1967, 20, 155–166. [Google Scholar] [CrossRef]
- Peterson, G.L. A Simplification of the Protein Assay Method of Lowry et al. Which Is More Generally Applicable. Anal. Biochem. 1977, 83, 346–356. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Gene | Forward | Reverse |
|---|---|---|
| hGAPDH | 5′-TCGGAGTCAACGGATTTGG-3′ | 5′-GCAACAATATCCACTTTACCAGAGTTAA-3′ |
| hRunx2 | 5′-GGCCCACAAATCTCAGATCGTT-3′ | 5′-CACTGGCGCTGCAACAAGAC-3 |
| hOCN | 5′-TGCAGAGTCCAGCAAAGGTGCA-3′ | 5′-ATAGGCCTCCTGAAAGCCGATGT-3′ |
| hOPN | 5′-TCACAGCCATGAAGATATGCTGG-3′ | 5′-TACAGGGAGTTTCCATGAAGCCAC-3′ |
| Antibody | Molecular Weight | Catalogue Number | Dilution | Ref. |
|---|---|---|---|---|
| Rabbit anti-protein kinase c (PKC) | 80 kDa | 2058 | 1/1000 | Cell signalling (Leiden, The Netherlands) |
| Rabbit anti-phospho PKC | 80, 82 kDa | 9374 | ||
| Rabbit anti-glyceraldehyde-3-phosphate dehydrogenase (GAPDH) | 37 kDa | 2118 | ||
| Goat anti-rabbit HRP | 7074 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ballout, N.; Boullier, A.; Darwiche, W.; Ait-Mohand, K.; Trécherel, E.; Gallégo, T.; Gomila, C.; Yaker, L.; Gennero, I.; Kovensky, J.; et al. DP2, a Carbohydrate Derivative, Enhances In Vitro Osteoblast Mineralisation. Pharmaceuticals 2023, 16, 1512. https://doi.org/10.3390/ph16111512
Ballout N, Boullier A, Darwiche W, Ait-Mohand K, Trécherel E, Gallégo T, Gomila C, Yaker L, Gennero I, Kovensky J, et al. DP2, a Carbohydrate Derivative, Enhances In Vitro Osteoblast Mineralisation. Pharmaceuticals. 2023; 16(11):1512. https://doi.org/10.3390/ph16111512
Chicago/Turabian StyleBallout, Nissrine, Agnès Boullier, Walaa Darwiche, Katia Ait-Mohand, Eric Trécherel, Théo Gallégo, Cathy Gomila, Linda Yaker, Isabelle Gennero, José Kovensky, and et al. 2023. "DP2, a Carbohydrate Derivative, Enhances In Vitro Osteoblast Mineralisation" Pharmaceuticals 16, no. 11: 1512. https://doi.org/10.3390/ph16111512
APA StyleBallout, N., Boullier, A., Darwiche, W., Ait-Mohand, K., Trécherel, E., Gallégo, T., Gomila, C., Yaker, L., Gennero, I., Kovensky, J., Ausseil, J., & Toumieux, S. (2023). DP2, a Carbohydrate Derivative, Enhances In Vitro Osteoblast Mineralisation. Pharmaceuticals, 16(11), 1512. https://doi.org/10.3390/ph16111512