
Effects of Five Coumarins and Standardized Extracts from Tagetes lucida Cav. on Motor Impairment and Neuroinflammation Induced with Cuprizone

 , ,
, ,  , ,
, ,  and
and
Abstract
:1. Introduction
2. Results
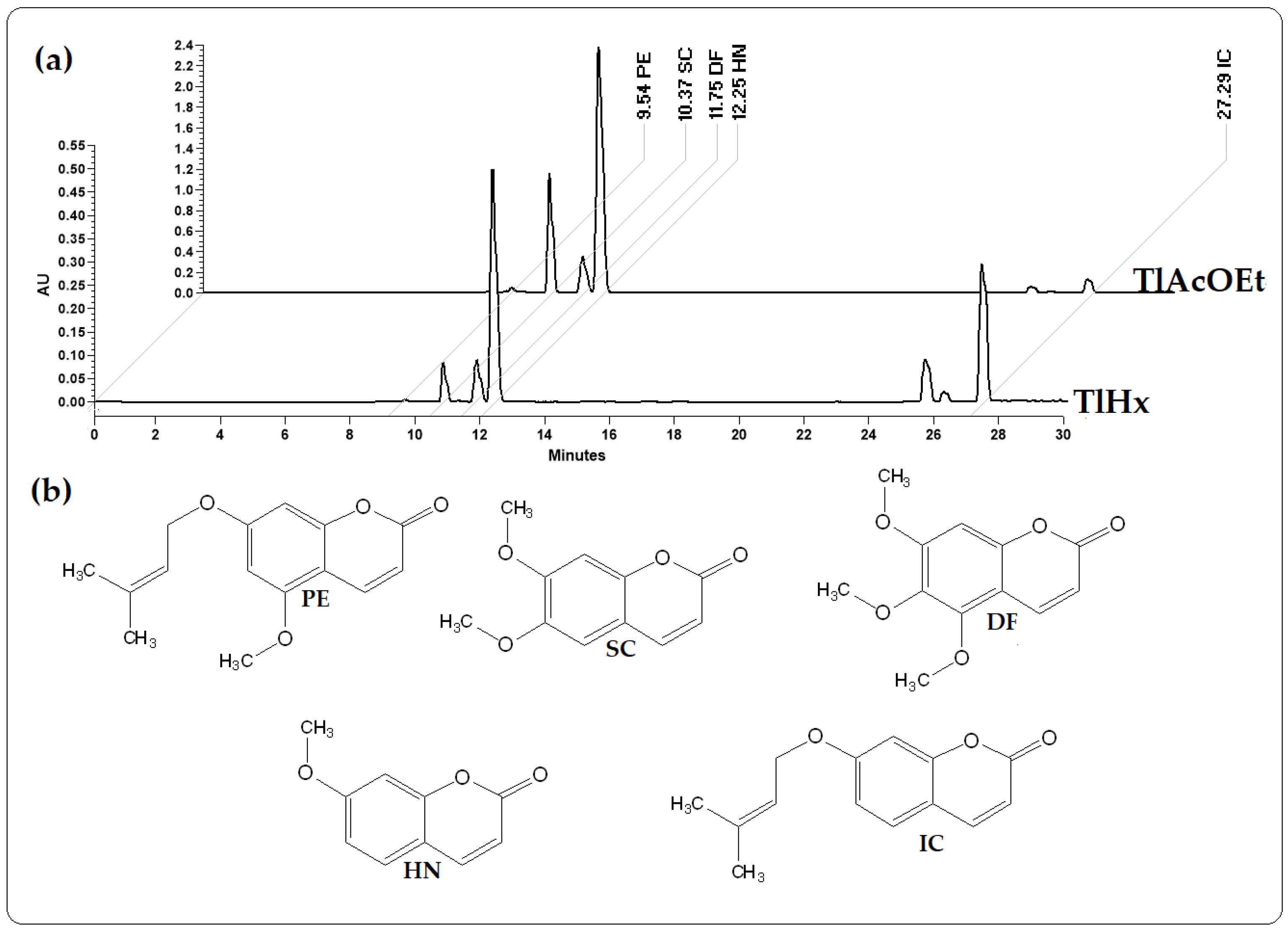
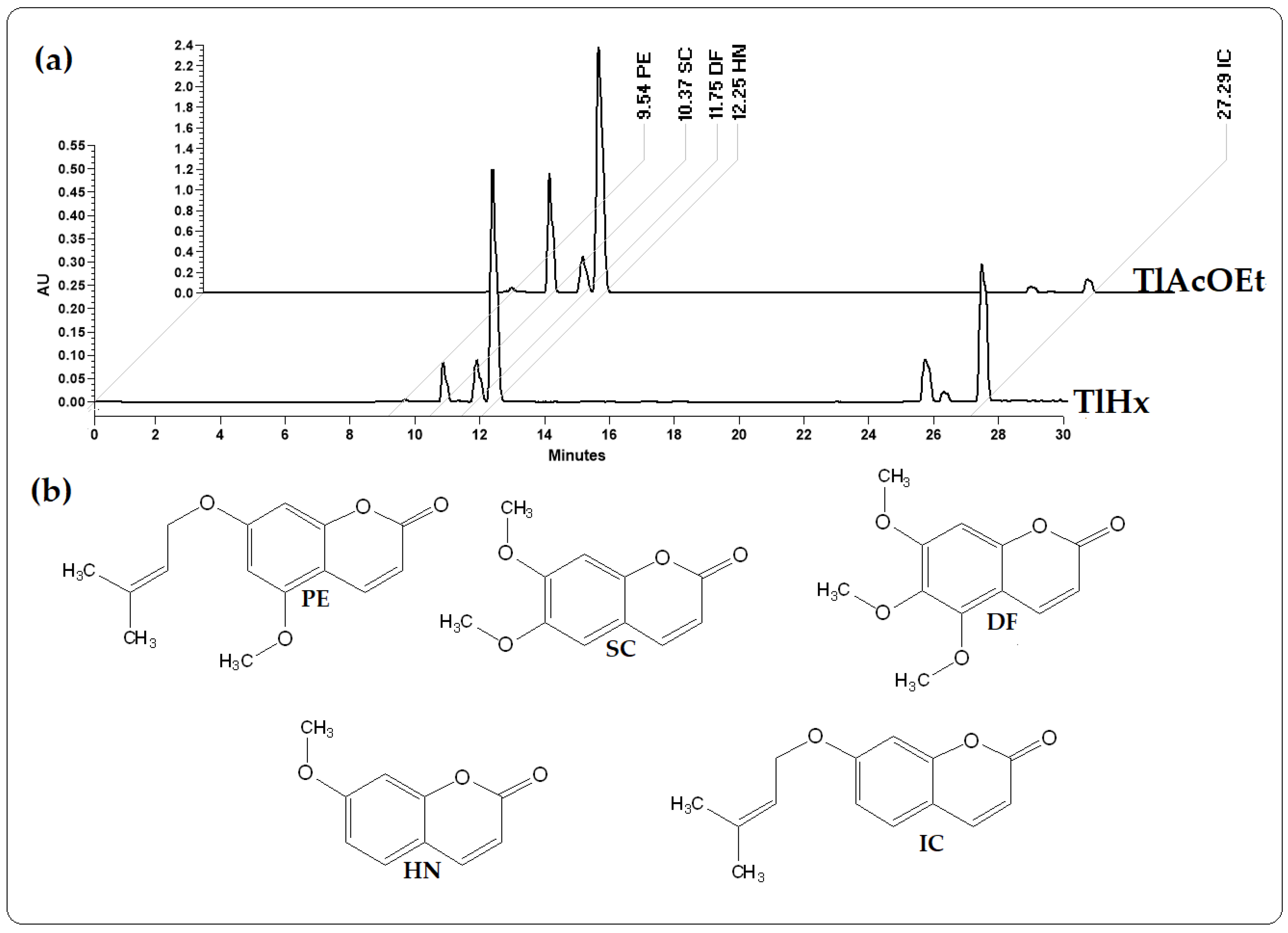
2.1. Quantification and Chemical Identification of Five Coumarins
2.2. Behavioral Tests
2.2.1. The Irwin Test
2.2.2. Open Field Test (OFT)
2.2.3. Rota-Rod Test (Rr)
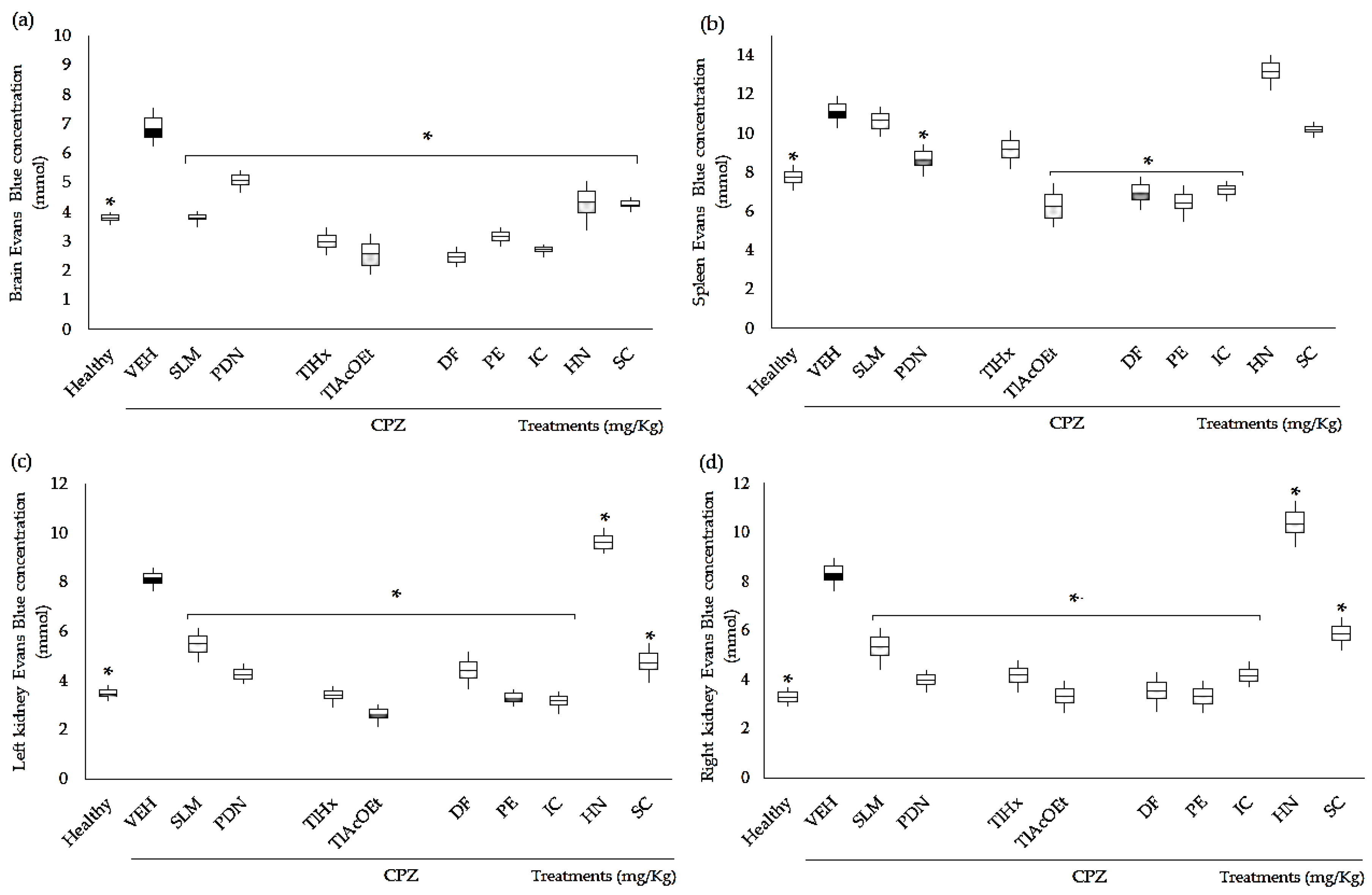
2.3. Effect of Extracts and Coumarins from T. lucida on Vascular Permeability
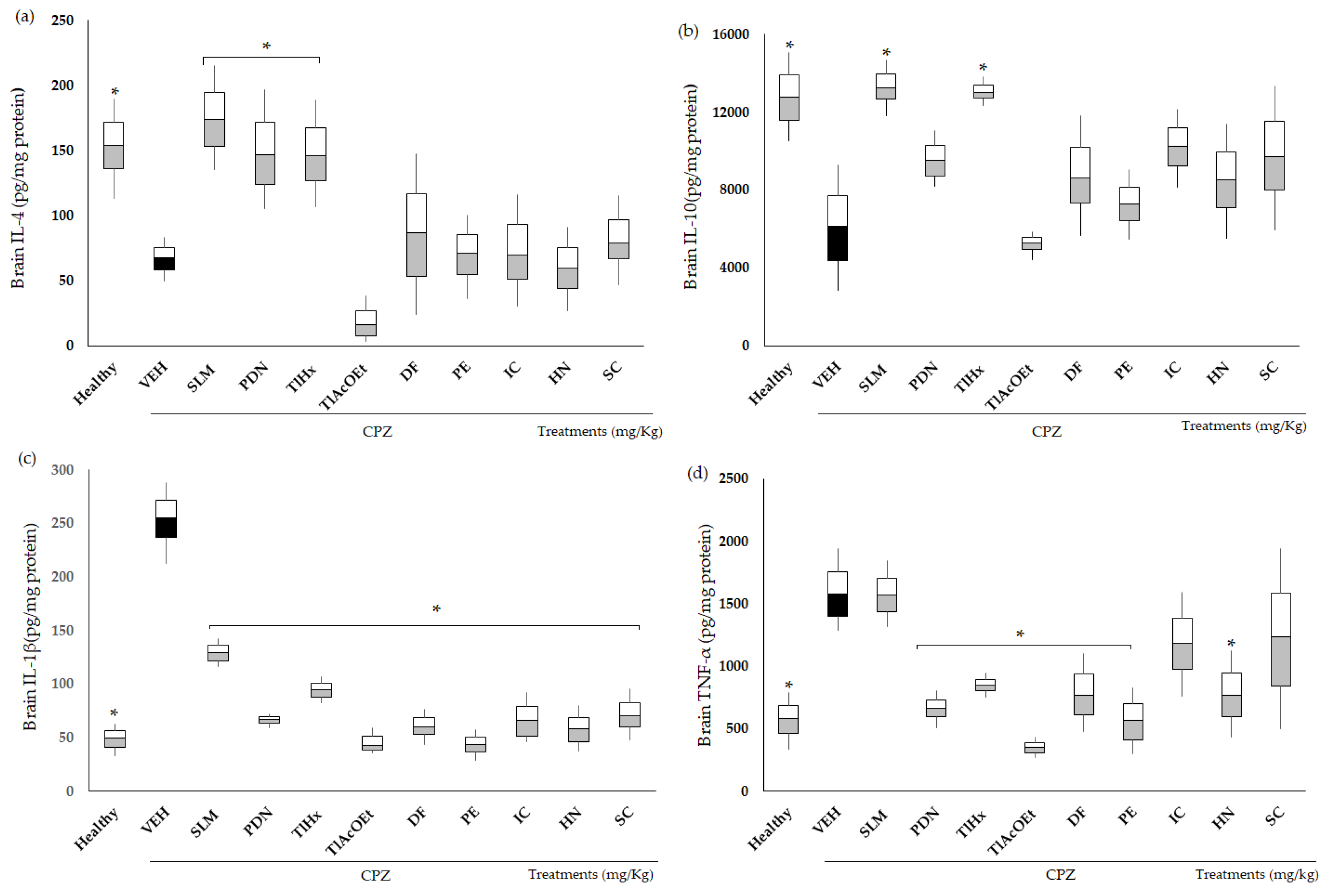
2.4. Effect of Extracts and Coumarins from T. lucida on Cytokine Concentrations
3. Discussion
3.1. Quantification and Chemical Identification of Five Coumarins
3.2. Evaluation of Two Extracts and Five Coumarins against the Motor Damage Induced by CPZ
3.3. Behavioral Tests
3.4. Evans Blue Extravasation
3.5. Cytokines as a Measure of Neuroinflammation
4. Materials and Methods
4.1. Plant Material
4.2. Preparation of Extracts
4.3. Isolation and Identification of Five Coumarins
4.4. HPLC Analysis of Extracts and Coumarins of T. lucida
4.5. Animals
4.6. Model of Deteriorous Behavior, Motor Damage and Neuroinflamation Induced with Cuprizone
- Healthy mice were treated with Tween 20 at 1% only. The remaining groups were treated with CPZ (Bis(cyclohexanone)oxaldihydrazone, daily for five weeks), either alone or co-administered at the end of week 2 with the respective treatments by the oral pathway.
- Damage group (VEH, Tween 20 at 1%).
- Sylimarin (SLM, 200 mg/kg).
- Prednisone (PDN, 2 mg/kg).
- 5.
- Isoprenyloxycumarin (IC, 25 mg/kg).
- 6.
- Herniarin (HN, 25 mg/kg).
- 7.
- 7-O-prenyl-scopoletin (PE, 25 mg/kg).
- 8.
- Dimethylfraxetin (DF, 25 mg/kg).
- 9.
- Scoporone (SC, 25 mg/kg).
- 10.
- TlHx (25 mg/kg).
- 11.
- TlAcOEt (25 mg/kg).
4.7. Behavioral Activity
4.7.1. The Irwin Test
4.7.2. Rota-Rod Test
4.7.3. Open Field Test (OFT)
4.8. Measure of Vascular Permeability with Extravasation of Evans Blue Test
4.9. Cytokine Quantification by ELISA
4.10. Statistics
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Peng, Y.; Zhu, F.Z.; Chen, Z.X.; Zhou, J.X.; Gan, L.; Yang, S.S.; Gao, S.; Liu, Q.Q. Characterization of myelin oligodendrocyte glycoprotein (MOG)35-55-specific CD8+ T cells in experimental autoimmune encephalomyelitis. Chin. Med. J. 2019, 132, 2934–2940. [Google Scholar] [CrossRef] [PubMed]
- Baecher-allan, C.; Kaskow, B.J.; Weiner, H.L. Review Multiple Sclerosis: Mechanisms and Immunotherapy. Neuron 2018, 97, 742–768. [Google Scholar] [CrossRef]
- Zirngibl, M.; Assinck, P.; Sizov, A.; Caprariello, A.V.; Plemel, J.R. Oligodendrocyte death and myelin loss in the cuprizone model: An updated overview of the intrinsic and extrinsic causes of cuprizone demyelination. Mol. Neurodegener. 2022, 17, 34. [Google Scholar] [CrossRef]
- Castillo-Álvarez, F.; Marzo-Sola, M.E. Role of intestinal microbiota in the development of multiple sclerosis. Neurología 2017, 32, 175–184. [Google Scholar] [CrossRef] [PubMed]
- Terry, R.L.; Ifergan, I.; Miller, S.D. Experimental Autoimmune Encephalomyelitis in Mice. Methods Mol. Biol. 2016, 1304, 145–160. [Google Scholar] [CrossRef] [PubMed]
- LaMothe, R.A.; Kolte, P.N.; Vo, T.; Ferrari, J.D.; Gelsinger, T.C.; Wong, J.; Chan, V.T.; Ahmed, S.; Srinivasan, A.; Deitemeyer, P.; et al. Tolerogenic nanoparticles induce antigen-specific regulatory T cells and provide therapeutic efficacy and transferrable tolerance against experimental autoimmune encephalomyelitis. Front. Immunol. 2018, 9, 281. [Google Scholar] [CrossRef]
- Macías-Cosme, K.; Cervantes-Llanos, M.; Marín-Prida, J.; Falcón-Cama, V.; Pentón-Arias, E.; Pentón-Rol, G. Generation of a murine chronic progressive experimental autoimmune encephalomyelitis model for molecular pharmacology studies in multiple sclerosis. Biotecnol. Apl. 2012, 29, 162–168. [Google Scholar]
- Constantinescu, C.S.; Farooqi, N.; O’Brien, K.; Gran, B. Experimental autoimmune encephalomyelitis (EAE) as a model for multiple sclerosis (MS). Br. J. Pharmacol. 2011, 164, 1079–1106. [Google Scholar] [CrossRef]
- Blaschek, A.; Karenfort, M.; Schimmel, M.; Pritsch, M.; Storm, K.; Gravesande, V.; Weber, M.; Schmoeger, M.; Seidl, R.; Prayer, D.; et al. Clinical and magnetic resonance imaging features of children, adolescents, and adults with a clinically isolated syndrome. SC. Eur. J. Paediatr. Neurol. 2018, 22, 1087–1094. [Google Scholar] [CrossRef]
- García-Díaz, B.; Estivill-Torrús, G. Modelos de experimentación animal para la investigación en esclerosis múltiple. Rev. Española Escler. Mult. 2008, 1, 5–9. [Google Scholar]
- Wootla, B.; Eriguchi, M.; Rodriguez, M. Is multiple sclerosis an autoimmune disease? Autoimmune Dis. 2012, 2012, 969657. [Google Scholar] [CrossRef]
- Barthelmes, J.; Tafferner, N.; Kurz, J.; de Bruin, N.; Parnham, M.J.; Geisslinger, G.; Schiffmann, S. Induction of experimental autoimmune encephalomyelitis in mice and evaluation of the disease-dependent distribution of immune cells in various tissues. J. Vis. Exp. 2016, 2016, 53933. [Google Scholar] [CrossRef]
- Torre-Fuentes, L.; Moreno-Jiménez, L.; Pytel, V.; Matías-Guiu, J.A.; Gómez-Pinedo, U.; Matías-Guiu, J. Experimental models of demyelination and remyelination. Neurología 2019, 35, 32–39. [Google Scholar] [CrossRef] [PubMed]
- Zhan, J.; Mann, T.; Joost, S.; Behrangi, N.; Frank, M.; Kipp, M. The Cuprizone Model: Dos and Do Nots. Cells 2020, 9, 843. [Google Scholar] [CrossRef] [PubMed]
- Gudi, V.; Gingele, S.; Skripuletz, T.; Stangel, M. Glial response during cuprizone-induced de- and remyelination in the CNS: Lessons learned. Front. Cell. Neurosci. 2014, 8, 73. [Google Scholar] [CrossRef]
- Torkildsen, Ø.; Brunborg, L.A.; Myhr, K.M.; Bø, L. The cuprizone model for demyelination. Acta Neurol. Scand. 2008, 117, 72–76. [Google Scholar] [CrossRef]
- González-Trujano, M.E.; Gutiérrez-Valentino, C.; Hernández-Arámburo, M.Y.; Díaz-Reval, M.I.; Pellicer, F. Identification of some bioactive metabolites and inhibitory receptors in the antinociceptive activity of Tagetes lucida Cav. Life Sci. 2019, 231, 116523. [Google Scholar] [CrossRef] [PubMed]
- Pérez-Ortega, G.; González-Trujano, M.E.; Ángeles-López, G.E.; Brindis, F.; Vibrans, H.; Reyes-Chilpa, R. Tagetes lucida Cav.: Ethnobotany, phytochemistry and pharmacology of its tranquilizing properties. J. Ethnopharmacol. 2016, 181, 221–228. [Google Scholar] [CrossRef] [PubMed]
- Capunzo, M.; Brunetti, L.; Cavallo, P.; Boccia, G.; Caro, F.D.E.; Ieluzzi, M. Antimicrobial activity of dry extracts of Tagetes lucida from Guatemala. J. Prev. Mesicine Hyg. 2003, 44, 85–87. [Google Scholar] [CrossRef]
- Adams, M.; Gmünder, F.; Hamburger, M. Plants traditionally used in age related brain disorders-A survey of ethnobotanical literature. J. Ethnopharmacol. 2007, 113, 363–381. [Google Scholar] [CrossRef]
- Guadarrama-Cruz, G.; Alarcon-Aguilar, F.J.; Lezama-Velasco, R.; Vazquez-Palacios, G.; Bonilla-Jaime, H. Antidepressant-like effects of Tagetes lucida Cav. in the forced swimming test. J. Ethnopharmacol. 2008, 120, 277–281. [Google Scholar] [CrossRef]
- Guzmán Gutiérrez, S.L.G.; Reyes Chilpa, R.R.; Bonilla Jaime, H. Medicinal plants for the treatment of “nervios”, anxiety, and depression in Mexican Traditional Medicine. Rev. Bras. Farmacogn. 2014, 24, 591–608. [Google Scholar] [CrossRef]
- Bonilla-Jaime, H.; Guadarrama-Cruz, G.; Alarcon-Aguilar, F.J.; Limón-Morales, O.; Vazquez-Palacios, G. Antidepressant-like activity of Tagetes lucida Cav. is mediated by 5-HT1A and 5-HT2A receptors. J. Nat. Med. 2015, 69, 463–470. [Google Scholar] [CrossRef]
- Malik, A.; Kushnoor, A.; Saini, V.; Singhal, S.; Kumar, S.; Yadav, Y.C. In vitro antioxidant properties of Scopoletin. J. Chem. Pharm. Res. 2011, 3, 659–665. [Google Scholar]
- Monterrosas-brisson, N.; Herrera-ruiz, M.; Jiménez-ferrer, E.; Bahena-pérez, R.; Avilés-flores, M.; Fuentes-mata, M.; Martínez-duncker, I.; González-cortazar, M.; Avilés-flores, M.; Fuentes-mata, M.; et al. Anti-inflammatory activity of coumarins isolated from Tagetes lucida Cav. Nat. Prod. Res. 2020, 34, 3244–3248. [Google Scholar] [CrossRef]
- Sandra Liliana, P.D.; Manasés, G.C.; Enrique, J.F.; Rubén, R.R.; Cinthya, B.P.; Belen, M.H.G.; Alejandro, Z.; Maribel, H.R. Isolation, chemical characterization, and anti-inflammatory activity of coumarins, flavonoids, and terpenes from Tagetes lucida. Nat. Prod. Res. 2022, 36, 4751–4756. [Google Scholar] [CrossRef] [PubMed]
- Porras-Dávila, S.L.; Jiménez-Ferrer, E.; Román Ramos, R.; González-Cortazar, M.; Almanza-Pérez, J.C.; Herrera-Ruiz, M. Herniarin, Dimethylfraxetin and Extracts from Tagetes lucida, inPsychosis Secondary to Ketamine and Its Interaction with Haloperidol. Plants 2022, 11, 2789. [Google Scholar] [CrossRef]
- Liliana Porras-Dávila, S.; Zamilpa, A.; Jiménez-Ferrer, E.; Jiménez-Aparicio, A.; Alejandra Santillan-Urquiza, M.; Díaz-Patricio, F.; Herrera-Ruiz, M. Anti-Inflammatory and Neuroprotective Effects of Standardized Fractions in Herniarin and Daphnoretin from Distictis buccinatoria. Chem. Biodivers 2023, 20, e202200969. [Google Scholar] [CrossRef]
- Hernández, T.; Canales, M.; Flores, C.; García, A.M.; Durán, A.; Avila, J.G. Antimicrobial activity of Tagetes lucida. Pharm Biol. 2006, 44, 19–22. [Google Scholar] [CrossRef]
- Chaves, O.S.; Teles, Y.C.F.; De Oliveira Monteiro, M.M.; Mendes, L.D.G., Jr.; De Fátima Agra, M.; De Andrade Braga, V.; Silva, T.M.S.; de Fátima Vanderlei de Souza, M. Alkaloids and phenolic compounds from Sida rhombifolia L. (Malvaceae) and vasorelaxant activity of two indoquinoline alkaloids. Molecules 2017, 22, 94. [Google Scholar] [CrossRef]
- Bubols, G.B.; Vianna, D.d.R.; Medina-Remón, A.; von Poser, G.; Lamuela-Raventos, R.M.; Eifler-Lima, V.L.; Garcia, S.C. The Antioxidant Activity of Coumarins and Flavonoids. Mini-Rev. Med. Chem. 2013, 13, 318–334. [Google Scholar] [PubMed]
- Santibáñez, A.; Herrera-Ruiz, M.; González-Cortazar, M.; Nicasio-Torres, P.; Sharma, A.; Jiménez-Ferrer, E. Pharmacokinetics and Tissue Distribution of Coumarins from Tagetes lucida in an LPS-Induced Neuroinflammation Model. Plants 2022, 11, 2805. [Google Scholar] [CrossRef] [PubMed]
- Kopanitsa, M.V.; Lehtimäki, K.K.; Forsman, M.; Suhonen, A.; Koponen, J.; Piiponniemi, T.O.; Kärkkäinen, A.M.; Pavlidi, P.; Shatillo, A.; Sweeney, P.J.; et al. Cognitive disturbances in the cuprizone model of multiple sclerosis. Genes Brain Behav. 2021, 20, e12663. [Google Scholar] [CrossRef]
- Mathiasen, J.R.; Moser, V.C. The Irwin Test and Functional Observational Battery (FOB) for Assessing the Effects of Compounds on Behavior, Physiology, and Safety Pharmacology in Rodents. Curr. Protoc. Pharmacol. 2018, 83, e43. [Google Scholar] [CrossRef] [PubMed]
- McPhetres, J.; Zickfeld, J.H. The physiological study of emotional piloerection: A systematic review and guide for future research. Int. J. Psychophysiol. 2022, 179, 6–20. [Google Scholar] [CrossRef] [PubMed]
- Sternberg, Z. Sympathetic Nervous System Dysfunction in Multiple Sclerosis, Linking Neurodegeneration to a Reduced Response to Therapy. Curr. Pharm. Des. 2012, 18, 1635–1644. [Google Scholar] [CrossRef]
- Andrade, J.C.; Monteiro, Á.B.; Andrade, H.H.N.; Gonzaga, T.K.S.N.; Silva, P.R.; Alves, D.N.; Castro, R.D.; Maia, M.S.; Scotti, M.T.; Sousa, D.P.; et al. Involvement of GABA A Receptors in the Anxiolytic-Like Effect of Hydroxycitronellal. BioMed Res. Int. 2021, 2021, 9929805. [Google Scholar] [CrossRef]
- Kraeuter, A.K.; Guest, P.C.; Sarnyai, Z. The Open Field Test for Measuring Locomotor Activity and Anxiety-like Behavior. Methods Mol. Biol. 2019, 1916, 99–103. [Google Scholar] [CrossRef]
- Liu, H.; Huang, X.; Xu, J.; Mao, H.; Li, Y.; Ren, K.; Ma, G.; Xue, Q.; Tao, H.; Wu, S.; et al. Dissection of the relationship between anxiety and stereotyped self-grooming using the Shank3B mutant autistic model, acute stress model and chronic pain model. Neurobiol. Stress 2021, 15, 100417. [Google Scholar] [CrossRef]
- Wang, X.; Chang, L.; Wan, X.; Tan, Y.; Qu, Y.; Shan, J.; Yang, Y.; Ma, L.; Hashimoto, K. (R)-ketamine ameliorates demyelination and facilitates remyelination in cuprizone-treated mice: A role of gut–microbiota–brain axis. Neurobiol. Dis. 2022, 165, 105635. [Google Scholar] [CrossRef]
- Franco-Pons, N.; Torrente, M.; Colomina, M.T.; Vilella, E. Behavioral deficits in the cuprizone-induced murine model of demyelination/remyelination. Toxicol. Lett. 2007, 169, 205–213. [Google Scholar] [CrossRef]
- Guo, H.; Cao, H.; Cui, X.; Zheng, W.; Wang, S.; Yu, J.; Chen, Z. Silymarin’s inhibition and treatment effects for Alzheimer’s disease. Molecules 2019, 24, 1748. [Google Scholar] [CrossRef] [PubMed]
- Kren, V.; Walterová, D. Silybin and silymarin--new effects and applications. Biomed. Pap. Med. Fac. Univ. Palacky. Olomouc. Czech. Repub. 2005, 149, 29–41. [Google Scholar] [CrossRef]
- Berghoff, S.A.; Düking, T.; Spieth, L.; Winchenbach, J.; Stumpf, S.K.; Gerndt, N.; Kusch, K.; Ruhwedel, T.; Möbius, W.; Saher, G. Blood-brain barrier hyperpermeability precedes demyelination in the cuprizone model. Acta Neuropathol. Commun. 2017, 5, 94. [Google Scholar] [CrossRef] [PubMed]
- Abbott, N.J.; Patabendige, A.A.K.; Dolman, D.E.M.; Yusof, S.R.; Begley, D.J. Structure and function of the blood-brain barrier. Neurobiol. Dis. 2010, 37, 13–25. [Google Scholar] [CrossRef] [PubMed]
- Radu, M.; Chernoff, J. An in vivo assay to test blood vessel permeability. J. Vis. Exp. 2013, 73, e50062. [Google Scholar] [CrossRef]
- Goldim, M.P.d.S.; Della Giustina, A.; Petronilho, F. Using Evans Blue Dye to Determine Blood-Brain Barrier Integrity in Rodents. Curr. Protoc. Immunol. 2019, 126, e83. [Google Scholar] [CrossRef]
- Krasselt, M.; Baerwald, C. Efficacy and safety of modified-release prednisone in patients with rheumatoid arthritis. Drug Des. Devel. Ther. 2016, 10, 1047–1058. [Google Scholar] [CrossRef]
- Wan, C.; Wei, Y.; Ma, J.; Geng, X. Protective effects of scoparone against ischemia-reperfusion-induced myocardial injury. Mol. Med. Rep. 2018, 18, 1752–1760. [Google Scholar] [CrossRef]
- Suzuki, K. Giant hepatic mitochondria: Production in mice fed with cuprizone. Science 1969, 163, 81–82. [Google Scholar] [CrossRef]
- Kesterson, J.W.; Carlton, W.W. Monoamine oxidase inhibition and the activity of other oxidative enzymes in the brains of mice fed cuprizone. Toxicol. Appl. Pharmacol. 1971, 20, 386–395. [Google Scholar] [CrossRef] [PubMed]
- Kaddatz, H.; Joost, S.; Nedelcu, J.; Chrzanowski, U.; Schmitz, C.; Gingele, S.; Gudi, V.; Stangel, M.; Zhan, J.; Santrau, E.; et al. Cuprizone-induced demyelination triggers a CD8-pronounced T cell recruitment. Glia 2021, 69, 925–942. [Google Scholar] [CrossRef] [PubMed]
- Slavin, A.; Ewing, C.; Liu, J.; Ichikawa, M.; Slavin, J.; Bernard, C.C.A. Induction of a multiple sclerosis-like disease in mice with an immunodominant epitope of myelin oligodendrocyte glycoprotein. Autoimmunity 1998, 28, 109–120. [Google Scholar] [CrossRef] [PubMed]
- Pasquini, L.A.; Calatayud, C.A.; Bertone Uña, A.L.; Millet, V.; Pasquini, J.M.; Soto, E.F. The neurotoxic effect of cuprizone on oligodendrocytes depends on the presence of pro-inflammatory cytokines secreted by microglia. Neurochem. Res. 2007, 32, 279–292. [Google Scholar] [CrossRef]
- Miller, S.D.; Karpus, W.J.; Davidson, T.S. Experimental Autoimmune Encephalomyelitis in the Mouse. Curr. Protoc. Immunol. 2010, 88, 15.1.1–15.1.20. [Google Scholar] [CrossRef]
- Honeycutt, S.E.; O’Brien, L.L. Injection of Evans blue dye to fluorescently label and image intact vasculature. Biotechniques 2020, 70, 181–185. [Google Scholar] [CrossRef]



{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Extract | IC | HN | DF | SC | PE |
|---|---|---|---|---|---|
| (µg/mg) | (µg/mg) | (µg/mg) | (µg/mg) | (µg/mg) | |
| TlHx | 40.1 | 79.9 | 19.9 | 15,528.4 | 0.1 |
| TlAcOEt | 18.1 | 381.9 | 75.3 | 230,692.7 | 3.05 |
| Week | Healtly | VEH | SLM | PDN | TlHx | TlAcOEt | DF | PE | IC | HN | SC |
|---|---|---|---|---|---|---|---|---|---|---|---|
| Cuprizone | |||||||||||
| 1 | N/O | N/O | N/O | N/O | N/O | N/O | N/O | N/O | N/O | N/O | N/O |
| 2 | N/O | 20 * | 13 * | 15 * | 27 * | 20 * | 7 * | N/O | 13 * | 20 * | 27 * |
| 3 | N/O | 53 * | 40 * | 37 * | 27 * | 47 * | 27 * | 40 * | 33 * | 33 * | 20 * |
| 4 | N/O | 93 * | 73 * | 40 * | 27 * | 33 * | 13 * | 33 * | 40 * | 27 * | 13 * |
| 5 | N/O | 93 * | 60 * | 26 * | 20 * | 20 * | 20 * | 7 * | 13 * | 20 * | 6 * |
| Week | Healtly | VEH | SLM | PDN | TlHx | TlAcOEt | DF | PE | IC | HN | SC |
|---|---|---|---|---|---|---|---|---|---|---|---|
| Cuprizone | |||||||||||
| 1 | N/O | 27 * | 33 * | 20 * | 20 * | 13 * | 27 * | 13 * | 40 * | 33 * | 20 * |
| 2 | N/O | 33 * | 33 * | 37 * | 27 * | 27 * | 40 * | 60 * | 47 * | 47 * | 40 * |
| 3 | N/O | 87 * | 53 * | 40 * | 27 * | 40 * | 33 * | 53 * | 33 * | 40 * | 33 * |
| 4 | N/O | 87 * | 33 * | 28 * | 20 * | 33 * | 20 * | 40 * | 27 * | 27 * | 20 * |
| 5 | N/O | 100 * | 20 * | 20 * | 13 * | N/O | N/O | N/O | 13 * | 13 * | 13 * |
| Undisturbed Observations | Animal Handling | After Administration | |||
|---|---|---|---|---|---|
| a | Writhing | f | Abdominal tone | n | Crowding |
| b | Jumping | g | Limb tone | o | Ataxia |
| c | Tremor | h | Pupil size | p | Increased locomotion |
| d | Convulsion | i | Visual placement | q | No exploration |
| e | Raised hair | j | Lacrimation | ||
| k | Salivation | ||||
| m | Diarrhea | ||||
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Castro-Martínez, G.; Herrera-Ruiz, M.; González-Cortázar, M.; Porras-Dávila, S.L.; Almanza Pérez, J.C.; Jimenez-Ferrer, E. Effects of Five Coumarins and Standardized Extracts from Tagetes lucida Cav. on Motor Impairment and Neuroinflammation Induced with Cuprizone. Pharmaceuticals 2023, 16, 1391. https://doi.org/10.3390/ph16101391
Castro-Martínez G, Herrera-Ruiz M, González-Cortázar M, Porras-Dávila SL, Almanza Pérez JC, Jimenez-Ferrer E. Effects of Five Coumarins and Standardized Extracts from Tagetes lucida Cav. on Motor Impairment and Neuroinflammation Induced with Cuprizone. Pharmaceuticals. 2023; 16(10):1391. https://doi.org/10.3390/ph16101391
Chicago/Turabian StyleCastro-Martínez, Gabriela, Maribel Herrera-Ruiz, Manases González-Cortázar, Sandra Liliana Porras-Dávila, Julio Cesar Almanza Pérez, and Enrique Jimenez-Ferrer. 2023. "Effects of Five Coumarins and Standardized Extracts from Tagetes lucida Cav. on Motor Impairment and Neuroinflammation Induced with Cuprizone" Pharmaceuticals 16, no. 10: 1391. https://doi.org/10.3390/ph16101391
APA StyleCastro-Martínez, G., Herrera-Ruiz, M., González-Cortázar, M., Porras-Dávila, S. L., Almanza Pérez, J. C., & Jimenez-Ferrer, E. (2023). Effects of Five Coumarins and Standardized Extracts from Tagetes lucida Cav. on Motor Impairment and Neuroinflammation Induced with Cuprizone. Pharmaceuticals, 16(10), 1391. https://doi.org/10.3390/ph16101391