
In Silico Pharmacokinetic Profiling of the Identified Bioactive Metabolites of Pergularia tomentosa L. Latex Extract and In Vitro Cytotoxic Activity via the Induction of Caspase-Dependent Apoptosis with S-Phase Arrest

 ,
,  , ,
, ,
Abstract
:1. Introduction
2. Results
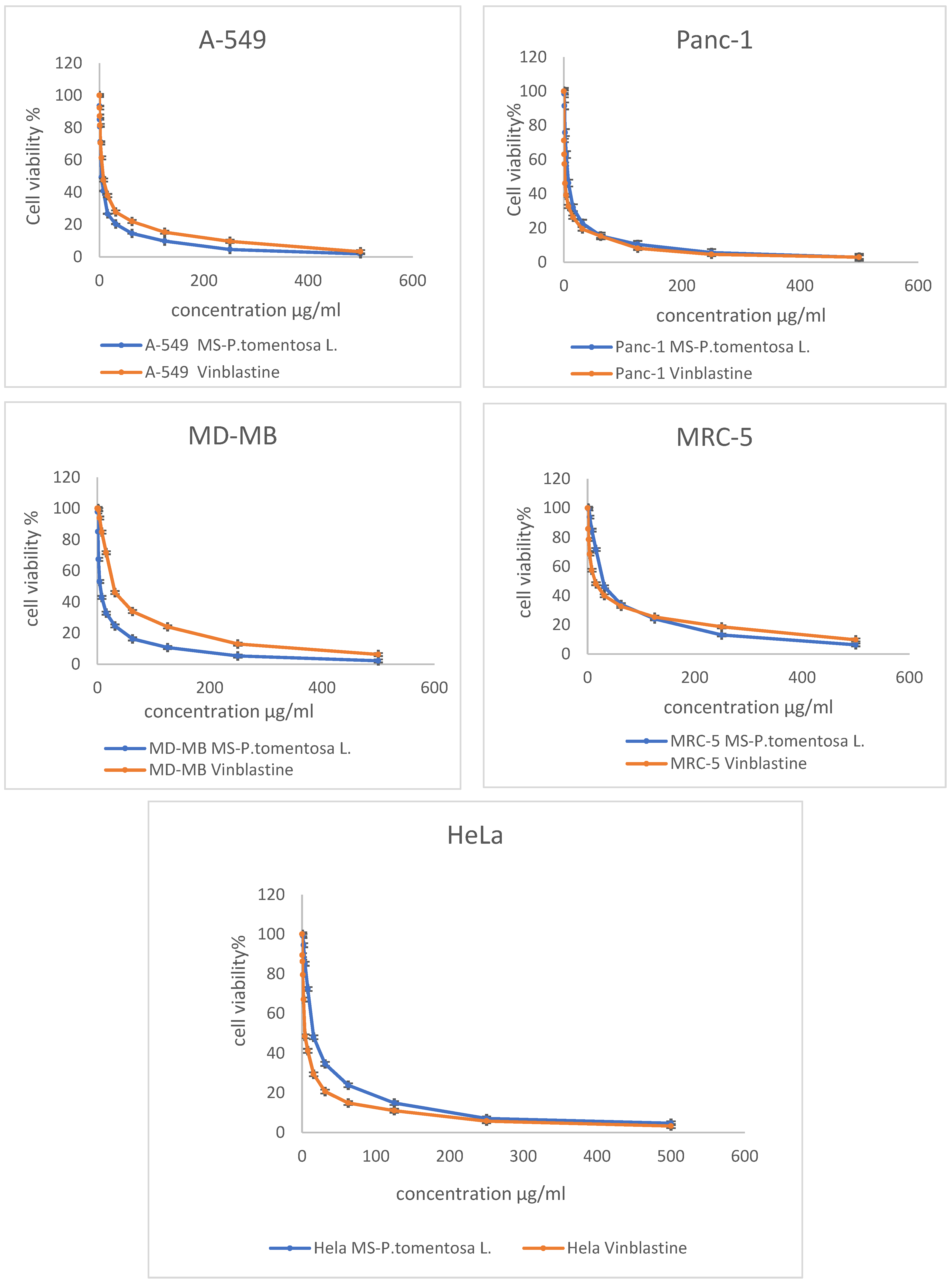
2.1. In Vitro Cytotoxicity Study
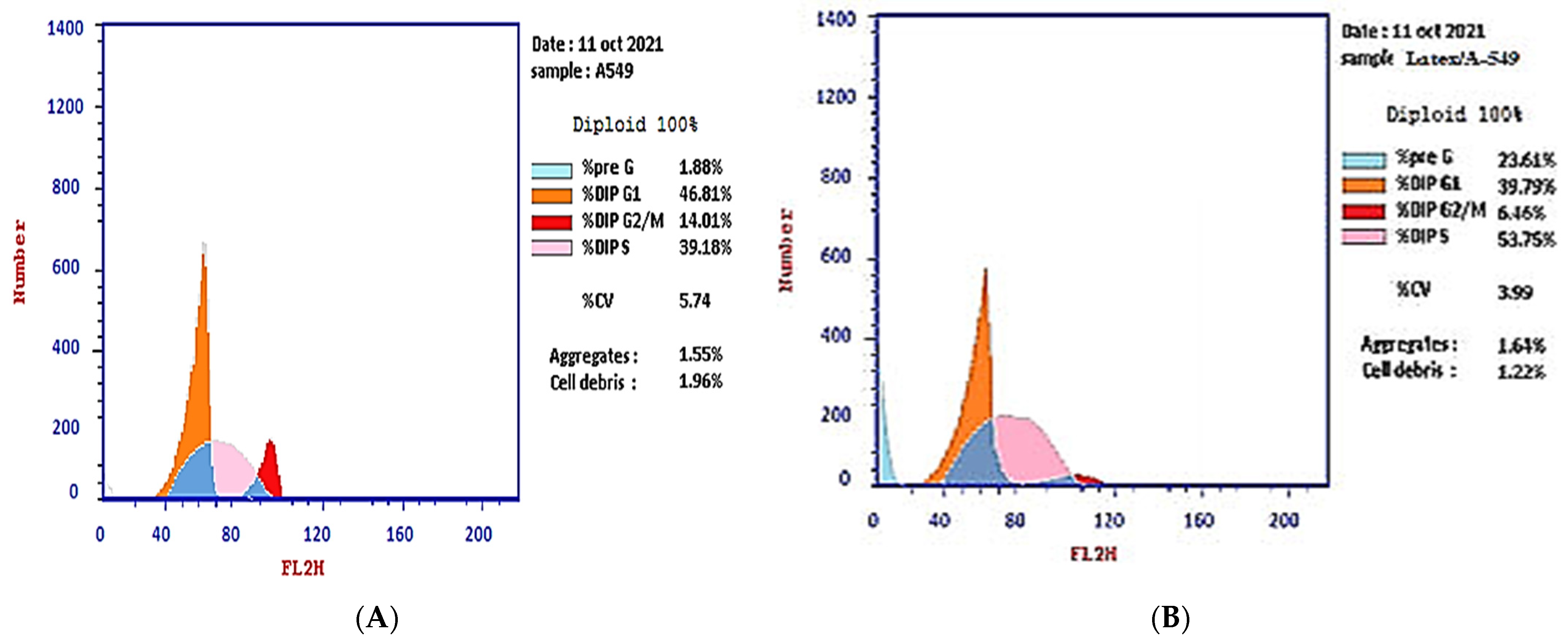
2.2. Cell Cycle Analysis
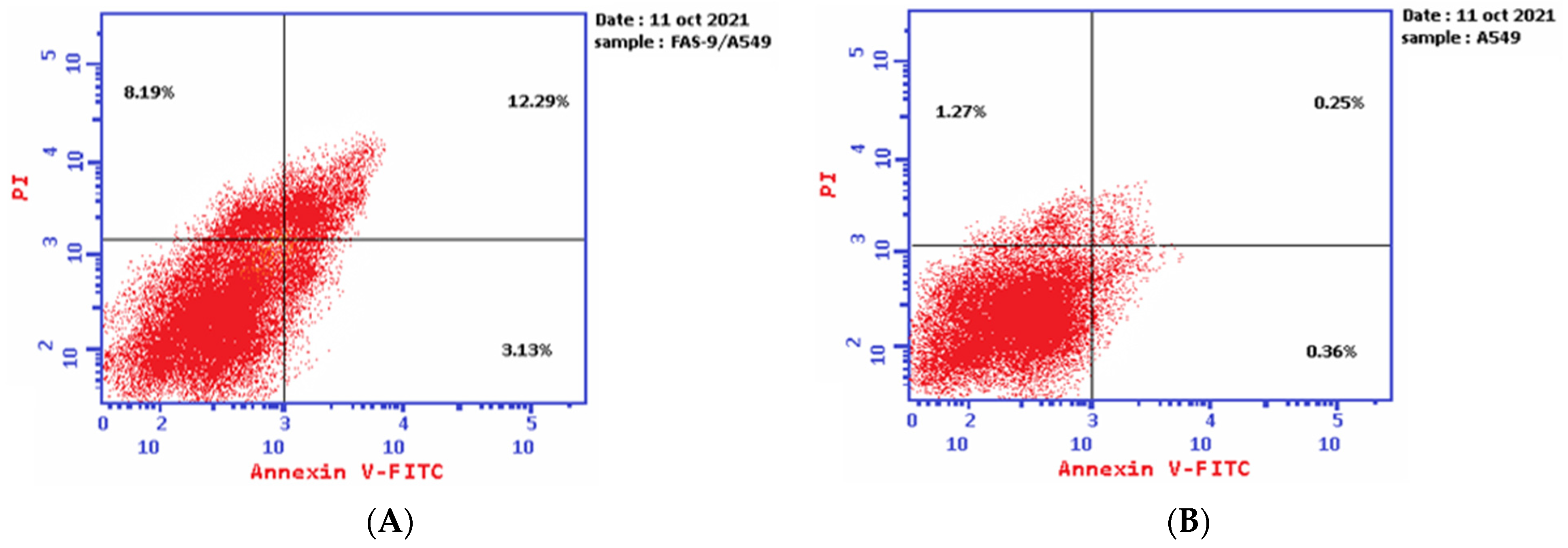
2.3. Annexin V-FITC Assay
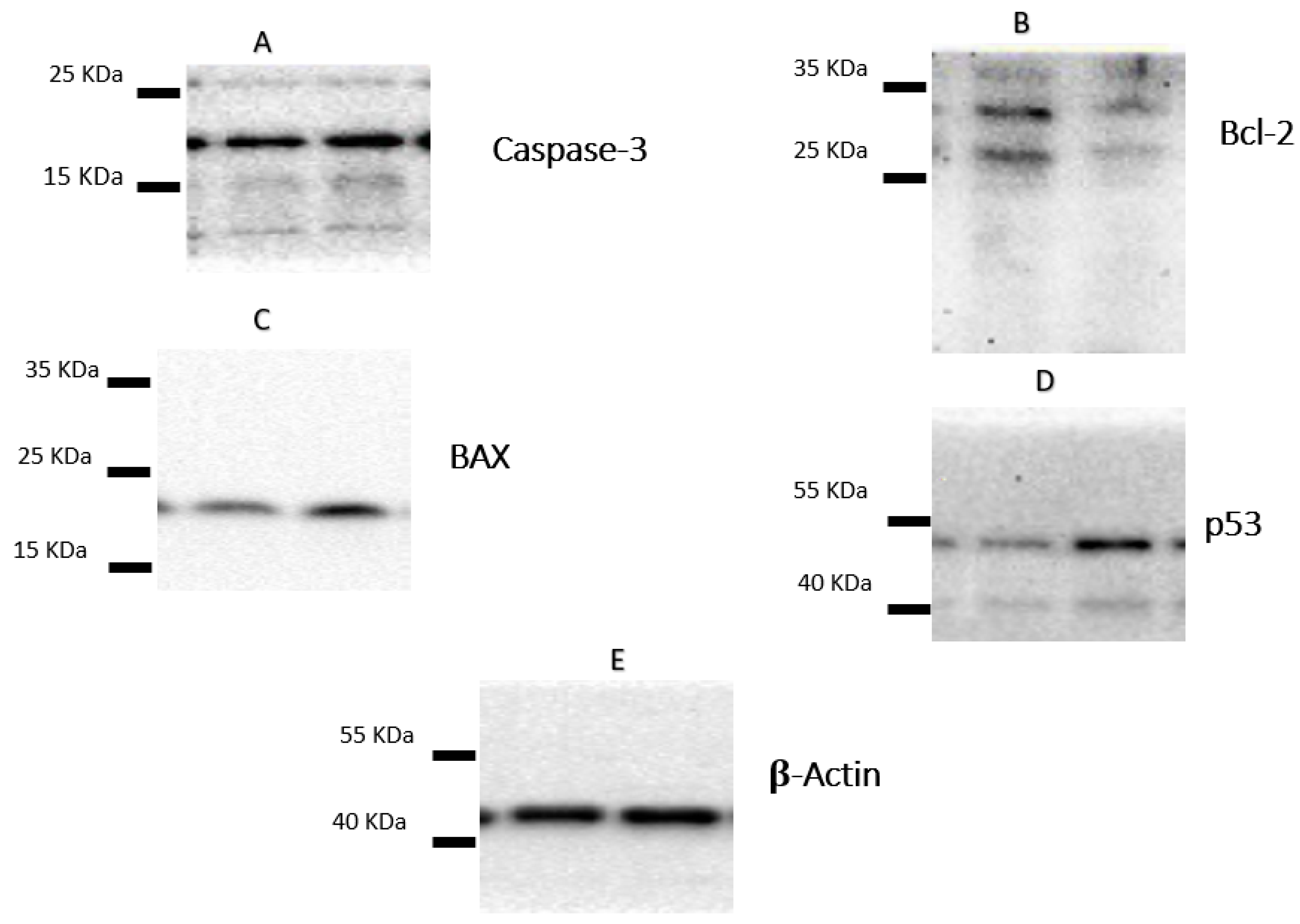
2.4. Apoptotic Markers of Cytotoxicity
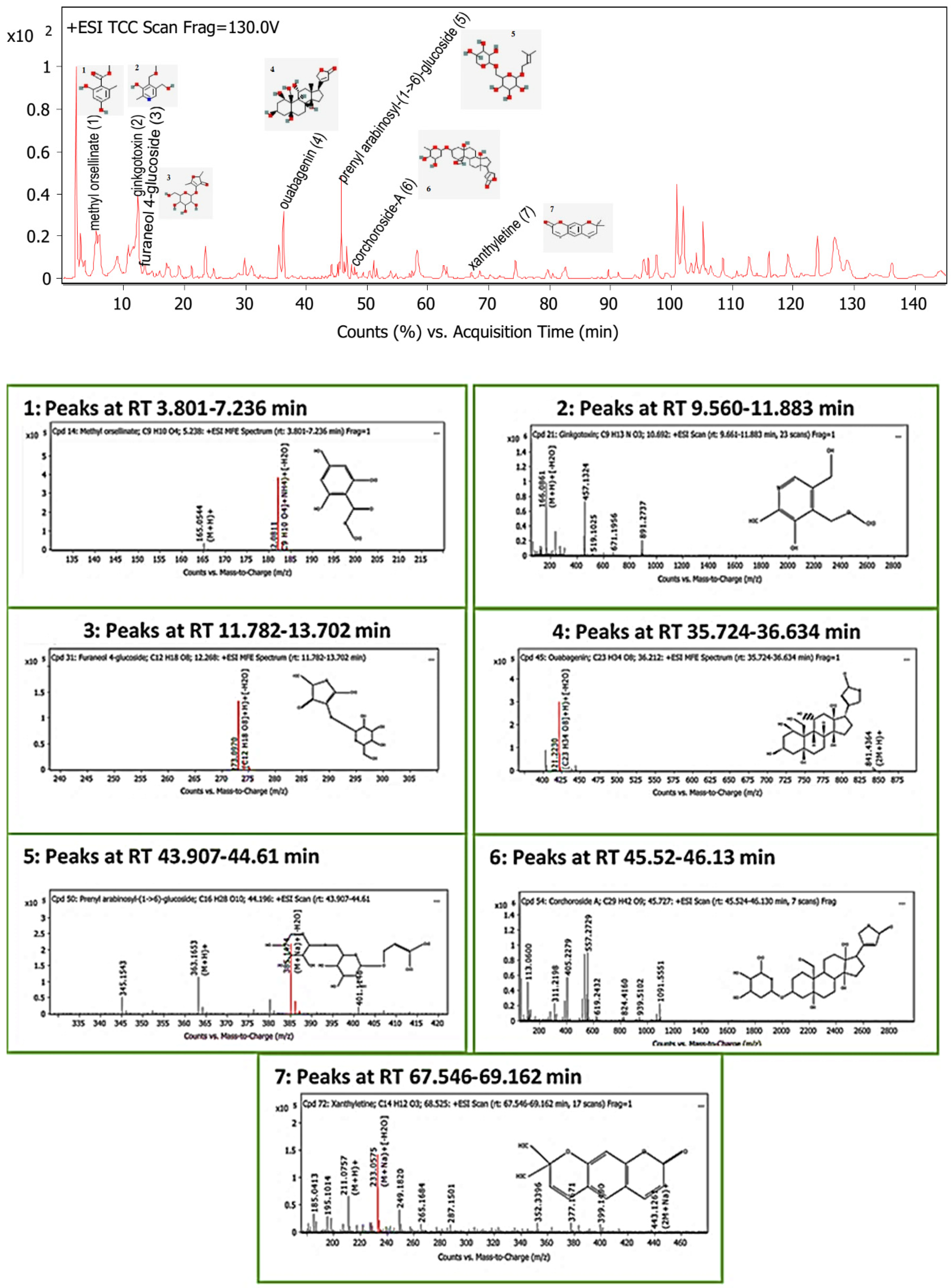
2.5. Screening of Secondary Metabolites
2.6. In Silico Study
2.6.1. Molecular Docking of Screened Metabolites
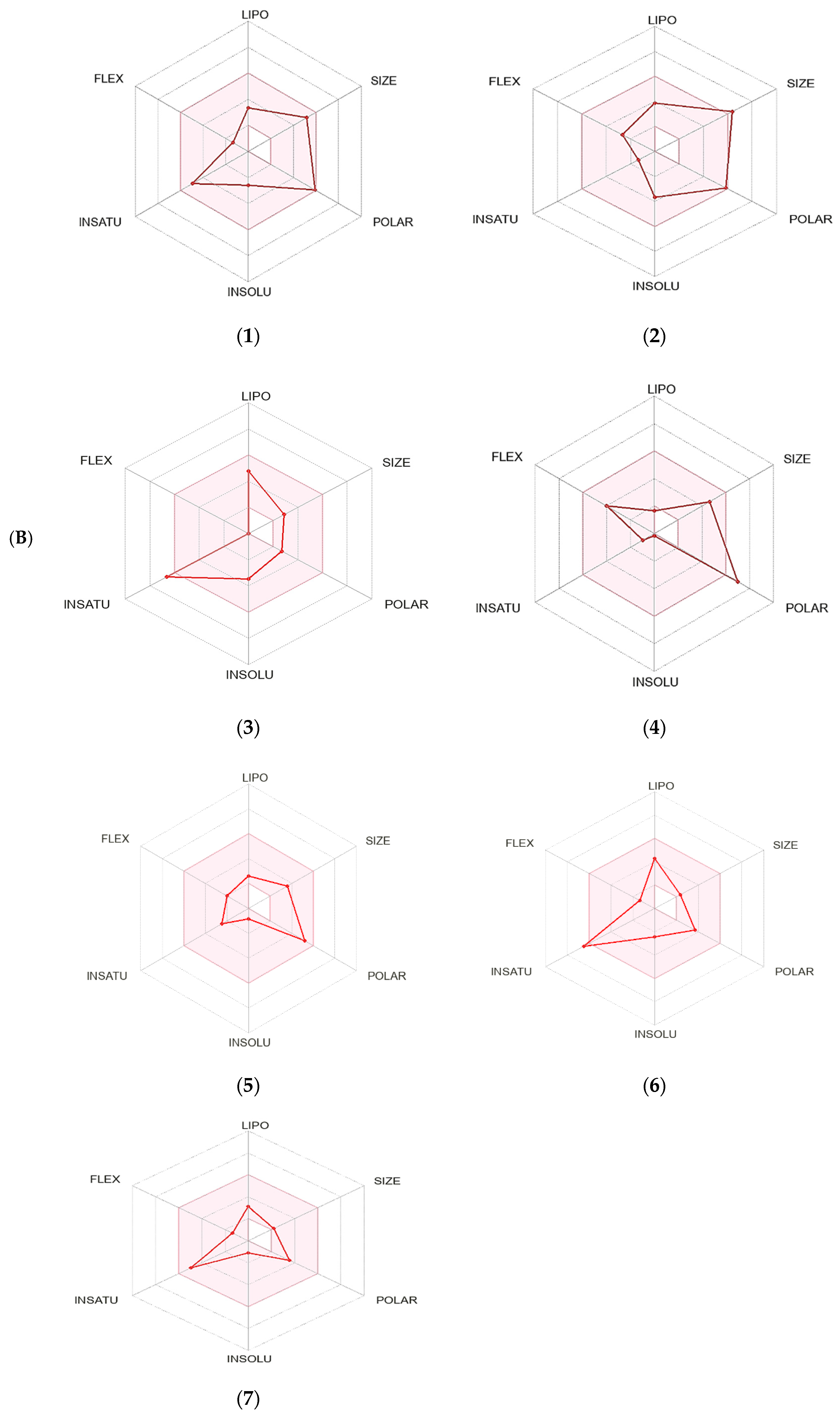
2.6.2. Pharmacokinetics of the Screened Secondary Metabolites
2.6.3. In Silico Toxicity Data
3. Discussion
4. Materials and Methods
4.1. Chemicals
4.2. Plant Material and Extraction
4.3. In Vitro Biological Activity
4.3.1. Cytotoxic Activity
Cell Line Propagation
Evaluation of the MTT Assay for Cytotoxic Activity
4.3.2. Cell Cycle Analysis by Flow Cytometry
4.3.3. Apoptosis Analysis (Annexin V-FITC Assay)
4.3.4. Evaluation of the Apoptotic Markers of Cytotoxicity
4.4. Screening of the Secondary Metabolite by LC-Plant Latex
4.5. In Silico Study
4.5.1. Molecular Docking
4.5.2. In Silico ADME Assessment
4.5.3. In Silico Toxicity Assessment
4.6. Statistical Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Sung, H.; Ferlay, J.; Siegel, R.L.; Laversanne, M.; Soerjomataram, I.; Jemal, A.; Bray, F. Global Cancer Statistics 2020: GLOBOCAN Estimates of Incidence and Mortality Worldwide for 36 Cancers in 185 Countries. CA Cancer J. Clin. 2021, 71, 209–249. [Google Scholar] [CrossRef]
- Basak, D.; Arrighi, S.; Darwiche, Y.; Deb, S. Comparison of Anticancer Drug Toxicities: Paradigm Shift in Adverse Effect Profile. Life 2022, 12, 48. [Google Scholar] [CrossRef]
- Nurgali, K.; Jagoe, R.T.; Abalo, R. Editorial: Adverse Effects of Cancer Chemotherapy: Anything New to Improve Tolerance and Reduce Sequelae? Front. Pharmacol. 2018, 1, 245. [Google Scholar] [CrossRef]
- Arunachalam, S.S.; Shetty, A.P.; Panniyadi, N.; Meena, C.; Kumari, J.; Rani, B.; Das, P.; Kumari, S. Study on Knowledge of Chemotherapy’s Adverse Effects and Their Self-Care Ability to Manage-The Cancer Survivors Impact. Clin. Epidemiol. Glob. Health 2021, 11, 100765. [Google Scholar] [CrossRef]
- Adeleye, O.A.; Femi-Oyewo, M.N.; Bamiro, O.A.; Bakre, L.G.; Alabi, A.; Ashidi, J.S.; Balogun-Agbaje, O.A.; Hassan, O.M.; Fakoya, G. Ethnomedicinal Herbs in African Traditional Medicine with Potential Activity for the Prevention, Treatment, and Management of Coronavirus Disease 2019. Future J. Pharm. Sci. 2021, 7, 72–86. [Google Scholar] [CrossRef]
- Ahmed, I.A. Ethnomedicinal Uses of Some Common Malaysian Medicinal Plants. In Natural Drugs from Plants; El-Shemy, H.A., Ed.; IntechOpen: London, UK, 2022; ISBN 978-1-80356-021-2. [Google Scholar]
- Astutik, S.; Pretzsch, J.; Kimengsi, J.N. Asian Medicinal Plants’ Production and Utilization Potentials: A Review. Sustainability 2019, 11, 5483. [Google Scholar] [CrossRef]
- Hosseini, S.H.; Bibak, H.; Ghara, A.R.; Sahebkar, A.; Shakeri, A. Ethnobotany of the Medicinal Plants Used by the Ethnic Communities of Kerman Province, Southeast Iran. J. Ethnobiol. Ethnomed. 2021, 17, 31. [Google Scholar] [CrossRef]
- Talib, W.H.; Alsalahat, I.; Daoud, S.; Abutayeh, R.F.; Mahmod, A.I. Plant-Derived Natural Products in Cancer Research: Extraction, Mechanism of Action, and Drug Formulation. Molecules 2020, 25, 5319. [Google Scholar] [CrossRef]
- Lahmar, I.; Belghith, H.; Ben Abdallah, F.; Belghith, K. Nutritional Composition and Phytochemical, Antioxidative, and Antifungal Activities of Pergularia tomentosa L. Biomed. Res. Int. 2017, 2017, 6903817. [Google Scholar] [CrossRef]
- Ads, E.N.; Abouzied, A.S.; Alshammari, M.K. Evaluation of Cytotoxic Effects of Methanolic Extract of Pergularia tomentosa L Growing Wild in KSA. Asian Pac. J. Cancer Prev. 2021, 22, 67–72. [Google Scholar] [CrossRef]
- Hamed, A.I.; Plaza, A.; Balestrieri, M.L.; Mahalel, U.A.; Springuel, I.V.; Oleszek, W.; Pizza, C.; Piacente, S. Cardenolide Glycosides from Pergularia tomentosa and Their Proapoptotic Activity in Kaposi’s Sarcoma Cells. J. Nat. Prod. 2006, 69, 1319–1322. [Google Scholar] [CrossRef]
- Alghanem, S.M.; El-amier, Y.A. Phytochemical and Biological Evaluation of Pergularia tomentosa L. (Solanaceae) Naturally Growing in Arid Ecosystem. Int. J. Plant Sci. Ecol. 2017, 3, 7–15. [Google Scholar]
- Hosseini, M.; Ayyari, M.; Meyfour, A.; Piacente, S.; Cerulli, A.; Crawford, A.; Pahlavan, S. Cardenolide-Rich Fraction of Pergularia tomentosa as a Novel Antiangiogenic Agent Mainly Targeting Endothelial Cell Migration. DARU J. Pharm. Sci. 2020, 28, 533–543. [Google Scholar] [CrossRef]
- Haddaji, F.; Papetti, A.; Noumi, E.; Colombo, R.; Deshpande, S.; Aouadi, K.; Adnan, M.; Kadri, A.; Selmi, B.; Snoussi, M. Bioactivities and in Silico Study of Pergularia tomentosa L. Phytochemicals as Potent Antimicrobial Agents Targeting Type IIA Topoisomerase, TyrRS, and Sap1 Virulence Proteins. Environ. Sci. Pollut. Res. Int. 2021, 28, 25349–25367. [Google Scholar] [CrossRef]
- Piacente, S.; Masullo, M.; De Nève, N.; Dewelle, J.; Hamed, A.; Kiss, R.; Mijatovic, T. Cardenolides from Pergularia tomentosa Display Cytotoxic Activity Resulting from Their Potent Inhibition of Na+/K+-ATPase. J. Nat. Prod. 2009, 72, 1087–1091. [Google Scholar] [CrossRef]
- Thomas, R.; Weihua, Z. Rethink of EGFR in Cancer with Its Kinase Independent Function on Board. Front. Oncol. 2019, 9, 800. [Google Scholar] [CrossRef]
- Daina, A.; Michielin, O.; Zoete, V. SwissADME: A Free Web Tool to Evaluate Pharmacokinetics, Drug-Likeness and Medicinal Chemistry Friendliness of Small Molecules. Sci. Rep. 2017, 7, 42717. [Google Scholar] [CrossRef]
- Jain, A.K.; Singh, D.; Dubey, K.; Maurya, R.; Mittal, S.; Pandey, A.K. Models and Methods for In Vitro Toxicity. In Vitro Toxicol. 2018, 2018, 45–65. [Google Scholar] [CrossRef]
- Lee, H.M.; Yu, M.S.; Kazmi, S.R.; Oh, S.Y.; Rhee, K.H.; Bae, M.A.; Lee, B.H.; Shin, D.S.; Oh, K.S.; Ceong, H.; et al. Computational Determination of HERG-Related Cardiotoxicity of Drug Candidates. BMC Bioinform. 2019, 20, 67–73. [Google Scholar] [CrossRef]
- Nohmi, T. Thresholds of Genotoxic and Non-Genotoxic Carcinogens. Toxicol. Res. 2018, 34, 281. [Google Scholar] [CrossRef]
- Pucci, B.; Kasten, M.; Giordano, A. Cell Cycle and Apoptosis. Neoplasia 2000, 2, 291. [Google Scholar] [CrossRef] [Green Version]
- Malumbres, M.; Barbacid, M. Cell Cycle, CDKs and Cancer: A Changing Paradigm. Nat. Rev. Cancer 2009, 9, 153–166. [Google Scholar] [CrossRef]
- Pisani, C.; Ramella, M.; Boldorini, R.; Loi, G.; Billia, M.; Boccafoschi, F.; Volpe, A.; Krengli, M. Apoptotic and Predictive Factors by Bax, Caspases 3/9, Bcl-2, P53 and Ki-67 in Prostate Cancer after 12 Gy Single-Dose. Sci. Rep. 2020, 10, 7050. [Google Scholar] [CrossRef]
- Chen, J. The Cell-Cycle Arrest and Apoptotic Functions of P53 in Tumor Initiation and Progression. Cold Spring Harb. Perspect. Med. 2016, 6, a026104. [Google Scholar] [CrossRef]
- Huang, S.; Benavente, S.; Armstrong, E.A.; Li, C.; Wheeler, D.L.; Harari, P.M. P53 Modulates Acquired Resistance to EGFR Inhibitors and Radiation. Cancer Res. 2011, 71, 7071–7079. [Google Scholar] [CrossRef]
- Schuler, M.; Green, D.R. Mechanisms of P53-Dependent Apoptosis. Biochem. Soc. Trans. 2001, 29, 684. [Google Scholar] [CrossRef]
- Howells, C.C.; Baumann, W.T.; Samuels, D.C.; Finkielstein, C.V. The Bcl-2-Associated Death Promoter (BAD) Lowers the Threshold at Which the Bcl-2-Interacting Domain Death Agonist (BID) Triggers Mitochondria Disintegration. J. Theor. Biol. 2011, 271, 114–123. [Google Scholar] [CrossRef]
- Adrain, C.; Creagh, E.M.; Martin, S.J. Apoptosis-Associated Release of Smac/DIABLO from Mitochondria Requires Active Caspases and Is Blocked by Bcl-2. EMBO J. 2001, 20, 6627–6636. [Google Scholar] [CrossRef]
- Vince, J.E.; De Nardo, D.; Gao, W.; Vince, A.J.; Hall, C.; McArthur, K.; Simpson, D.; Vijayaraj, S.; Lindqvist, L.M.; Bouillet, P.; et al. The Mitochondrial Apoptotic Effectors BAX/BAK Activate Caspase-3 and -7 to Trigger NLRP3 Inflammasome and Caspase-8 Driven IL-1β Activation. Cell Rep. 2018, 25, 2339–2353.e4. [Google Scholar] [CrossRef]
- Swanton, E.; Savory, P.; Cosulich, S.; Clarke, P.; Woodman, P. Bcl-2 Regulates a Caspase-3/Caspase-2 Apoptotic Cascade in Cytosolic Extracts. Oncogene 1999, 18, 1781–1787. [Google Scholar] [CrossRef]
- Ingólfsdóttir, K.; Gudmundsdóttir, G.F.; Ögmundsdóttir, H.M.; Paulus, K.; Haraldsdóttir, S.; Kristinsson, H.; Bauer, R. Effects of Tenuiorin and Methyl Orsellinate from the Lichen Peltigera Leucophlebia on 5-/15-Lipoxygenases and Proliferation of Malignant Cell Lines in Vitro. Phytomedicine 2002, 9, 654–658. [Google Scholar] [CrossRef]
- Nugraha, A.S.; Laksono, T.A.; Firli, L.N.; Sukrisno Putri, C.P.Z.; Pratoko, D.K.; Zulfikar, Z.; Untari, L.F.; Wongso, H.; Lambert, J.M.; Dillon, C.T.; et al. Anti-Cancer Evaluation of Depsides Isolated from Indonesian Folious Lichens: Physcia Millegrana, Parmelia Dilatata and Parmelia Aurulenta. Biomolecules 2020, 10, 1420. [Google Scholar] [CrossRef]
- Reddy, S.T.; Mendonza, J.J.; Makani, V.K.K.; Bhadra, M.P.; Uppuluri, V.M. Synthesis of Some Novel Methyl β-Orsellinate Based 3, 5-Disubstituted Isoxazoles and Their Anti-Proliferative Activity: Identification of Potent Leads Active against MCF-7 Breast Cancer Cell. Bioorg. Chem. 2020, 105, 104374. [Google Scholar] [CrossRef]
- Sung, W.S.; Jung, H.J.; Lee, I.S.; Kim, H.S.; Lee, D.G. Antimicrobial Effect of Furaneol against Human Pathogenic Bacteria and Fungi. J. Microbiol. Biotechnol. 2006, 16, 349–354. [Google Scholar]
- Valente, R.C.; Capella, L.S.; Oliveira, M.M.M.; Nunes-Lima, L.T.; Cruz, F.C.M.; Palmieri, R.R.; Lopes, A.G.; Capella, M.A.M. Diverse Actions of Ouabain and Its Aglycone Ouabagenin in Renal Cells. Cell Biol. Toxicol. 2010, 26, 201–213. [Google Scholar] [CrossRef]
- Khatri, H.R.; Bhattarai, B.; Kaplan, W.; Li, Z.; Curtis Long, M.J.; Aye, Y.; Nagorny, P. Modular Total Synthesis and Cell-Based Anticancer Activity Evaluation of Ouabagenin and Other Cardiotonic Steroids with Varying Degrees of Oxygenation. J. Am. Chem. Soc. 2019, 141, 4849. [Google Scholar] [CrossRef]
- Tamura, S.; Okada, M.; Kato, S.; Shinoda, Y.; Shioda, N.; Fukunaga, K.; Ui-Tei, K.; Ueda, M. Ouabagenin Is a Naturally Occurring LXR Ligand without Causing Hepatic Steatosis as a Side Effect. Sci. Rep. 2018, 8, 2305. [Google Scholar] [CrossRef]
- Al-Snafi, A.E. The Contents and Pharmacological Importance of Corchorus Capsularis-A Review. IOSR J. Pharm. 2016, 6, 58–63. [Google Scholar]
- Tahsin, T.; Wansi, J.D.; Al-Groshi, A.; Evans, A.; Nahar, L.; Martin, C.; Sarker, S.D. Cytotoxic Properties of the Stem Bark of Citrus Reticulata Blanco (Rutaceae). Phytother. Res. 2017, 31, 1215–1219. [Google Scholar] [CrossRef]
- Seshacharyulu, P.; Ponnusamy, M.P.; Haridas, D.; Jain, M.; Ganti, A.K.; Batra, S.K. Targeting the EGFR Signaling Pathway in Cancer Therapy. Expert Opin. Ther. Targets 2012, 16, 15. [Google Scholar] [CrossRef]
- Ling, Y.H.; Li, T.; Yuan, Z.; Haigentz, M.; Weber, T.K.; Perez-Soler, R. Erlotinib, an Effective Epidermal Growth Factor Receptor Tyrosine Kinase Inhibitor, Induces P27KIP1 up-Regulation and Nuclear Translocation in Association with Cell Growth Inhibition and G1/S Phase Arrest in Human Non-Small-Cell Lung Cancer Cell Lines. Mol. Pharmacol. 2007, 72, 248–258. [Google Scholar] [CrossRef] [PubMed]
- Umar, A.B.; Uzairu, A.; Shallangwa, G.A.; Uba, S. In Silico Evaluation of Some 4-(Quinolin-2-Yl)Pyrimidin-2-Amine Derivatives as Potent V600E-BRAF Inhibitors with Pharmacokinetics ADMET and Drug-Likeness Predictions. Future J. Pharm. Sci. 2020, 6, 61. [Google Scholar] [CrossRef]
- El-Wahab, H.A.A.A.; Ali, A.M.; Abdel-Rahman, H.M.; Qayed, W.S. Synthesis, Biological Evaluation, and Molecular Modeling Studies of Acetophenones-Tethered 1,2,4-Triazoles and Their Oximes as Epidermal Growth Factor Receptor Inhibitors. Chem. Biol. Drug Des. 2021. [Google Scholar] [CrossRef]
- Pathania, S.; Kumar Singh, P. Analyzing FDA-Approved Drugs for Compliance of Pharmacokinetic Principles: Should There Be a Critical Screening Parameter in Drug Designing Protocols? Expert Opin. Drug Metab. Toxicol. 2021, 17, 351–354. [Google Scholar] [CrossRef]
- Sadeghi-Aliabadi, H.; Minaiyan, M.; Dabestan, A. Cytotoxic Evaluation of Doxorubicin in Combination with Simvastatin against Human Cancer Cells. Res. Pharm. Sci. 2010, 5, 127. [Google Scholar]
- Sak, K.; Lust, H.; Kase, M.; Jaal, J. Cytotoxic Action of Methylquercetins in Human Lung Adenocarcinoma Cells. Oncol. Lett. 2018, 15, 1973. [Google Scholar] [CrossRef]
- Guzmán, E.A.; Johnson, J.D.; Carrier, M.K.; Meyer, C.I.; Pitts, T.P.; Gunasekera, S.P.; Wright, A.E. Selective Cytotoxic Activity of the Marine Derived Batzelline Compounds against Pancreatic Cancer Cell Lines. Anticancer Drugs 2009, 20, 149. [Google Scholar] [CrossRef]
- Razak, N.A.; Abu, N.; Ho, W.Y.; Zamberi, N.R.; Tan, S.W.; Alitheen, N.B.; Long, K.; Yeap, S.K. Cytotoxicity of Eupatorin in MCF-7 and MDA-MB-231 Human Breast Cancer Cells via Cell Cycle Arrest, Anti-Angiogenesis and Induction of Apoptosis. Sci. Rep. 2019, 9, 1514. [Google Scholar] [CrossRef]
- Takii, T.; Yamamoto, Y.; Chiba, T.; Abe, C.; Belisle, J.T.; Brennan, P.J.; Onozaki, K. Simple Fibroblast-Based Assay for Screening of New Antimicrobial Drugs against Mycobacterium Tuberculosis. Antimicrob. Agents Chemother. 2002, 46, 2533. [Google Scholar] [CrossRef]
- Mosmann, T. Rapid Colorimetric Assay for Cellular Growth and Survival: Application to Proliferation and Cytotoxicity Assays. J. Immunol. Methods 1983, 65, 55–63. [Google Scholar] [CrossRef]
- Eldehna, W.M.; Abo-Ashour, M.F.; Nocentini, A.; Gratteri, P.; Eissa, I.H.; Fares, M.; Ismael, O.E.; Ghabbour, H.A.; Elaasser, M.M.; Abdel-Aziz, H.A.; et al. Novel 4/3-((4-Oxo-5-(2-Oxoindolin-3-Ylidene)Thiazolidin-2-Ylidene)Amino) Benzenesulfonamides: Synthesis, Carbonic Anhydrase Inhibitory Activity, Anticancer Activity and Molecular Modelling Studies. Eur. J. Med. Chem. 2017, 139, 250–262. [Google Scholar] [CrossRef]
- Eldehna, W.M.; Abo-Ashour, M.F.; Ibrahim, H.S.; Al-Ansary, G.H.; Ghabbour, H.A.; Elaasser, M.M.; Ahmed, H.Y.A.; Safwat, N.A. Novel [(3-Indolylmethylene)Hydrazono]Indolin-2-Ones as Apoptotic Anti-Proliferative Agents: Design, Synthesis and in Vitro Biological Evaluation. J. Enzyme Inhib. Med. Chem. 2018, 33, 686–700. [Google Scholar] [CrossRef]
- Aborehab, N.M.; Elnagar, M.R.; Waly, N.E. Gallic Acid Potentiates the Apoptotic Effect of Paclitaxel and Carboplatin via Overexpression of Bax and P53 on the MCF-7 Human Breast Cancer Cell Line. J. Biochem. Mol. Toxicol. 2021, 35, e22638. [Google Scholar] [CrossRef]
- Burnette, W.N. “Western Blotting”: Electrophoretic Transfer of Proteins from Sodium Dodecyl Sulfate--Polyacrylamide Gels to Unmodified Nitrocellulose and Radiographic Detection with Antibody and Radioiodinated Protein A. Anal. Biochem. 1981, 112, 195–203. [Google Scholar] [CrossRef]
- Liu, M.H.; Tong, X.; Wang, J.X.; Zou, W.; Cao, H.; Su, W.W. Rapid Separation and Identification of Multiple Constituents in Traditional Chinese Medicine Formula Shenqi Fuzheng Injection by Ultra-Fast Liquid Chromatography Combined with Quadrupole-Time-of-Flight Mass Spectrometry. J. Pharm. Biomed. Anal. 2013, 74, 141–155. [Google Scholar] [CrossRef]
- Morris, G.M.; Ruth, H.; Lindstrom, W.; Sanner, M.F.; Belew, R.K.; Goodsell, D.S.; Olson, A.J. AutoDock4 and AutoDockTools4: Automated Docking with Selective Receptor Flexibility. J. Comput. Chem. 2009, 30, 2785–2791. [Google Scholar] [CrossRef]
- Cosconati, S.; Forli, S.; Perryman, A.L.; Harris, R.; Goodsell, D.S.; Olson, A.J. Virtual Screening with AutoDock: Theory and Practice. Expert Opin. Drug Discov. 2010, 5, 597–607. [Google Scholar] [CrossRef]
- Forli, S.; Olson, A.J. A Force Field with Discrete Displaceable Waters and Desolvation Entropy for Hydrated Ligand Docking. J. Med. Chem. 2012, 55, 623–638. [Google Scholar] [CrossRef]
- Irwin, J.J.; Shoichet, B.K. ZINC-A Free Database of Commercially Available Compounds for Virtual Screening. J. Chem. Inf. Model. 2005, 45, 177. [Google Scholar] [CrossRef]
- O’Boyle, N.M.; Banck, M.; James, C.A.; Morley, C.; Vandermeersch, T.; Hutchison, G.R. Open Babel: An Open Chemical Toolbox. J. Cheminform. 2011, 3, 33. [Google Scholar] [CrossRef]
- Pettersen, E.F.; Goddard, T.D.; Huang, C.C.; Couch, G.S.; Greenblatt, D.M.; Meng, E.C.; Ferrin, T.E. UCSF Chimera-A Visualization System for Exploratory Research and Analysis. J. Comput. Chem. 2004, 25, 1605–1612. [Google Scholar] [CrossRef]
- Dong, J.; Wang, N.N.; Yao, Z.J.; Zhang, L.; Cheng, Y.; Ouyang, D.; Lu, A.P.; Cao, D.S. ADMETlab: A Platform for Systematic ADMET Evaluation Based on a Comprehensively Collected ADMET Database. J. Cheminform. 2018, 10, 29. [Google Scholar] [CrossRef]


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Sample Code | Concentration (µg/mL) | %Pre G1 | %G1 | %S | %G2/M |
|---|---|---|---|---|---|
| Plant Latex P. tomentosa L. (Treated cells) | 3.89 | 23.61 b,*** | 39.79 b,* | 53.75 b,** | 6.46 b,*** |
| A-549 cells (control) | 0 | 1.88 | 46.81 | 39.18 | 14.01 |
| Sample | Tested Conc. (µg/mL) | Early Apoptosis | Late Apoptosis | Necrosis |
|---|---|---|---|---|
| Plant- Latex P. tomentosa L. (Treated cells) | 3.89 | 3.13 b,** | 12.29 b,*** | 8.19 b,** |
| A-549 cells (control) | 0 | 0.36 | 0.25 | 1.27 |
| Samples | Protein Expression (Normalized to β-Actin) * | ||||
|---|---|---|---|---|---|
| BAX | Bcl-2 | BAX/Bcl-2 Ratio | Caspases-3 | p53 | |
| Control (A-549 cells, nontreated) | 1.00 | 1.00 | 1.00 | 1.00 | 1.00 |
| Plant latex of P. tomentosa L. (3.89 µg/mL) | 2.13 b,* | 0.41 b,** | 5.19 b,*** | 1.69 b,* | 3.94 b,** |
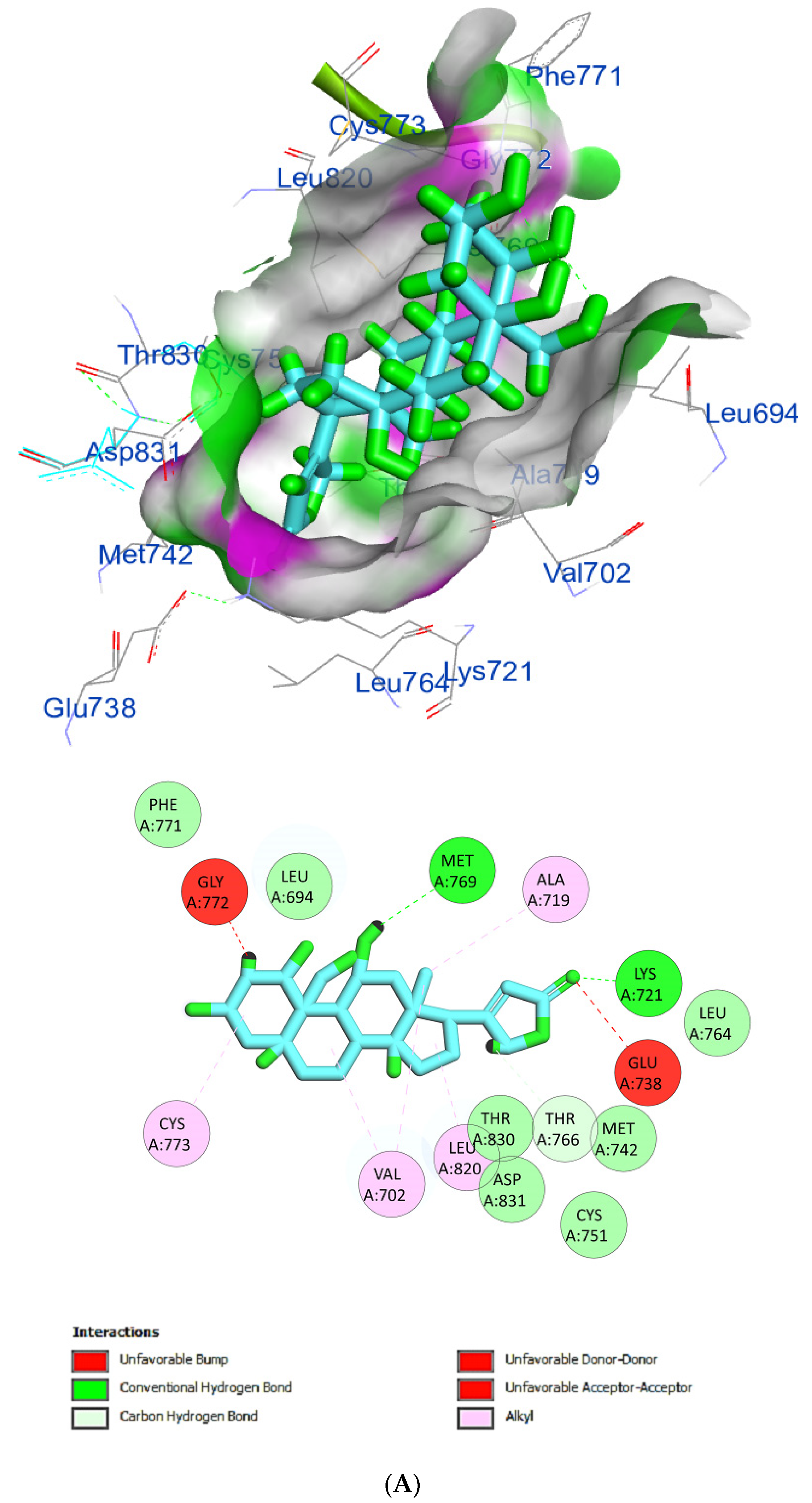
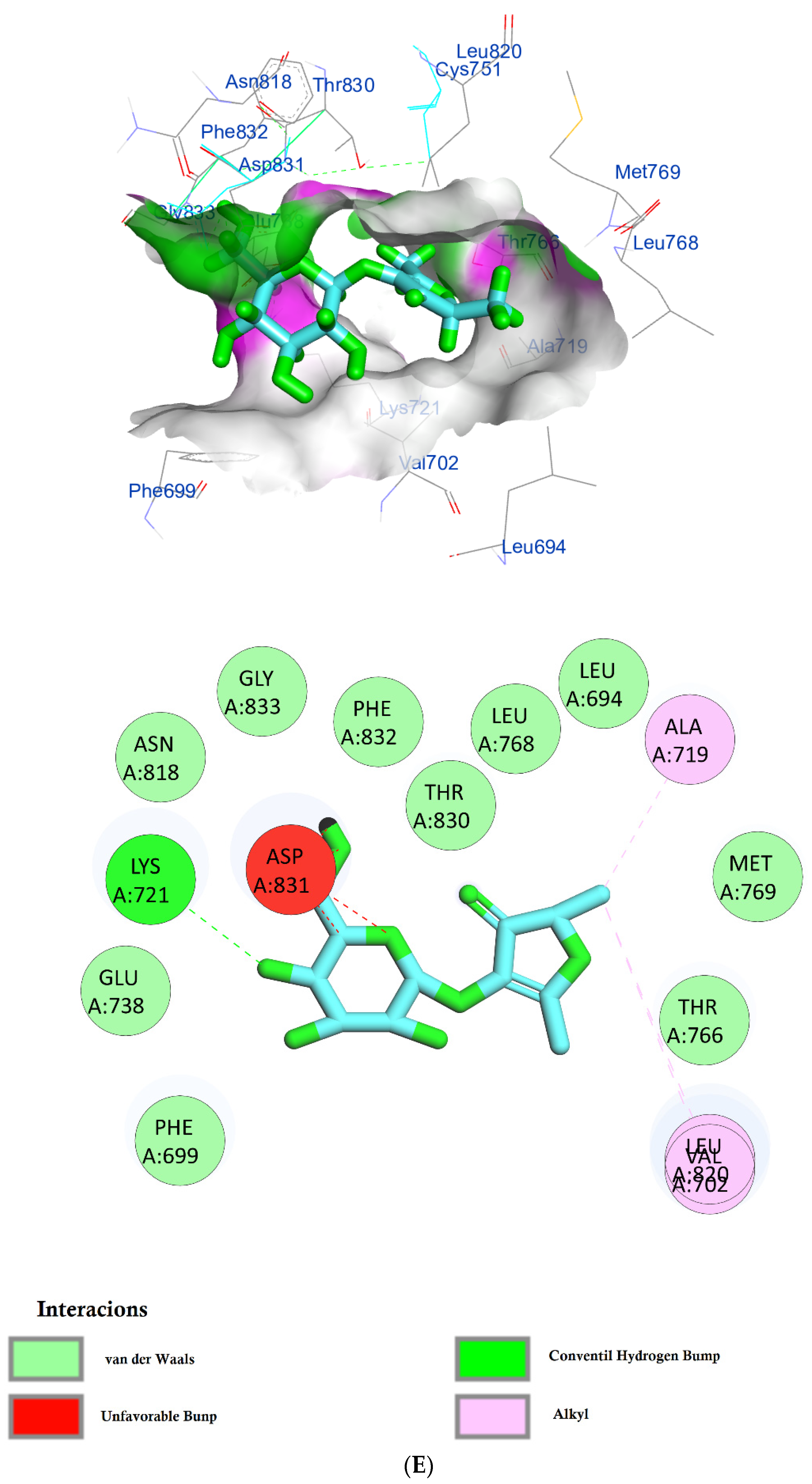
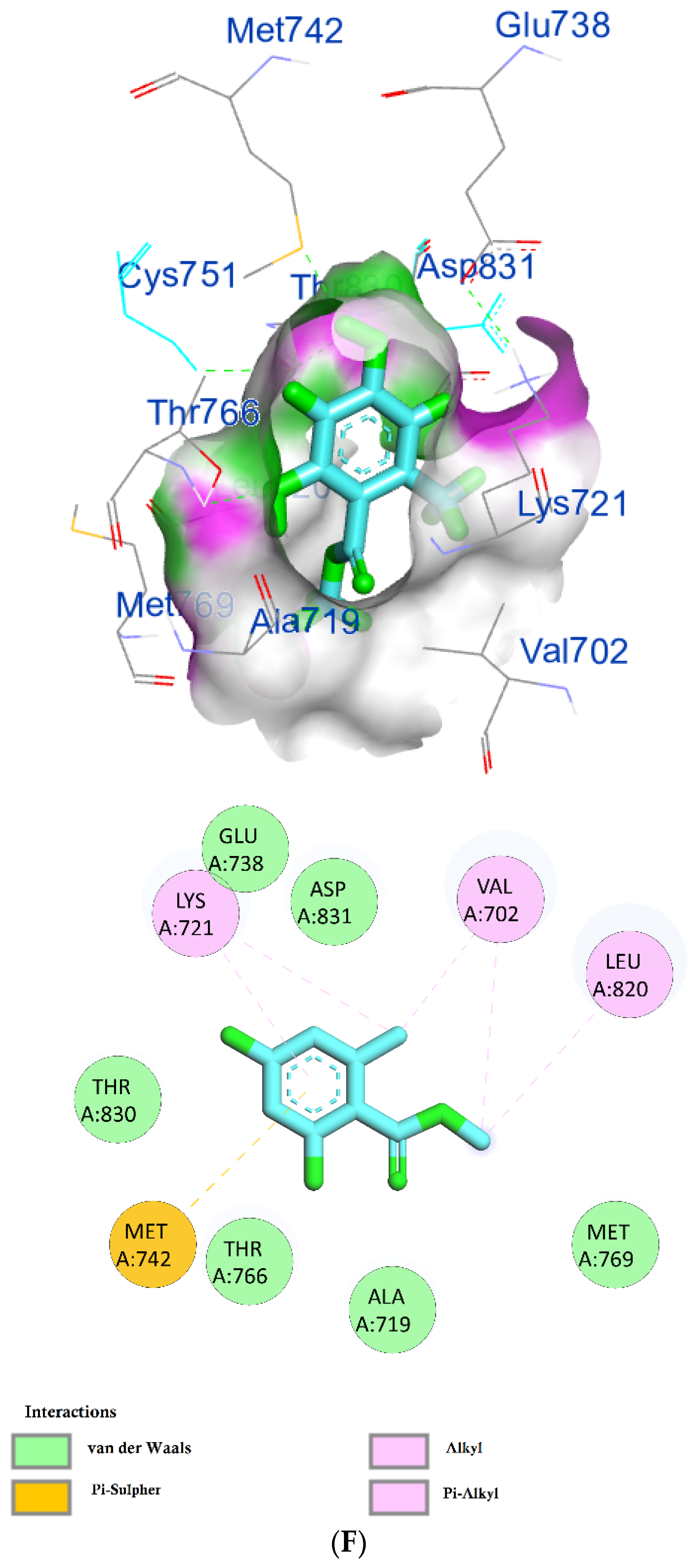
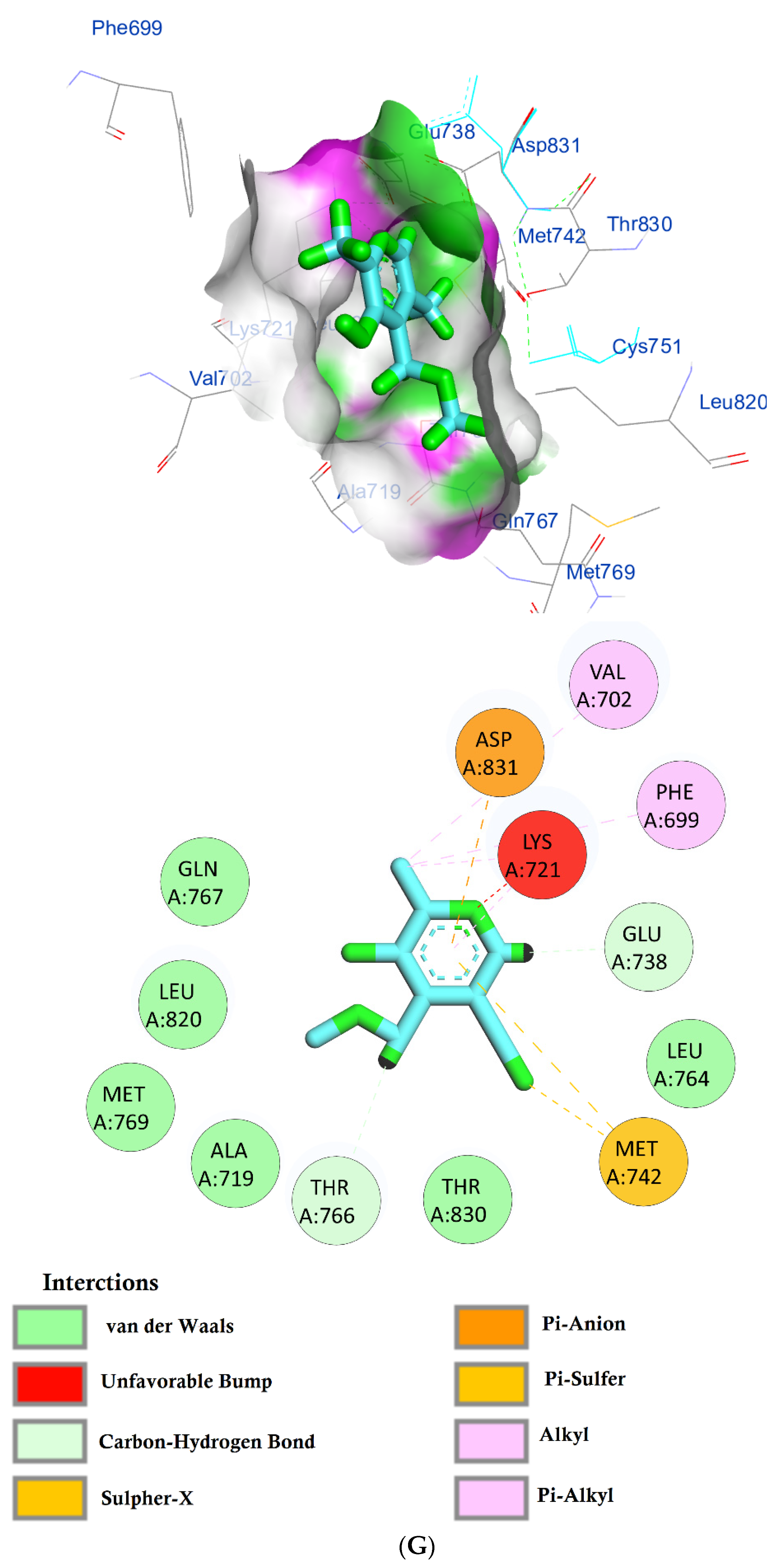
| Phytochemical Compounds | Glide Docking Score ΔG Kcal/mol | Receptor–Ligand Interaction | Distance Å |
|---|---|---|---|
| Ouabagenin | −8.6 | (CYS773)—(Ligand) C–H interaction | 2.76 |
| (VAL702)—(Ligand) Alkyl interaction | 3.88 | ||
| (LEU820)—(Ligand)Alkyl interaction | 2.51 | ||
| (THR766)—(Ligand) C–H interaction | 2.01 | ||
| (GLU736)—(Ligand) Hydrophobic interaction | 2.63 | ||
| (GLU772)—(Ligand) Hydrophobic interaction | 1.46 | ||
| (ALA719)—(Ligand) Hydrophobic interaction | 3.27 | ||
| (MET769)—(Ligand) H-bond interaction | 2.48 | ||
| (LYS721)—(Ligand) H-bond interaction and carboxylate salt bridge interactions | 2.60 4.13 | ||
| Corchoroside A | −8.1 | (CYS773)—(Ligand) H-bond interaction | 2.44 |
| (LYS704)—(Ligand) H-bond interaction | 2.09 | ||
| (GLY772)—(Ligand) H-bond interaction | 3.51 | ||
| (PRO770)—(Ligand) H-bond interaction | 3.06 | ||
| (LEU694)—(Ligand) H-bond interaction | 3.78 | ||
| (ASP831)—(Ligand) H-bond interaction | 3.90 | ||
| (ALA719)—(Ligand) Alkyl interaction and pi–alkyl hydrophobic interaction | 3.55 | ||
| (PHE771)—(Ligand) Alkyl interaction and pi–alkyl hydrophobic interaction | 4.20 | ||
| (LEU820)—(Ligand) Alkyl interaction and pi–alkyl hydrophobic interaction | 3.42 | ||
| (VAL702)—(Ligand) Alkyl interaction and pi–alkyl hydrophobic interaction | 3.85 | ||
| (MET769)—(Ligand) C–H bond interaction | 2.77 | ||
| (LYS721)—(Ligand) Carboxylate salt bridge interaction | 3.45 | ||
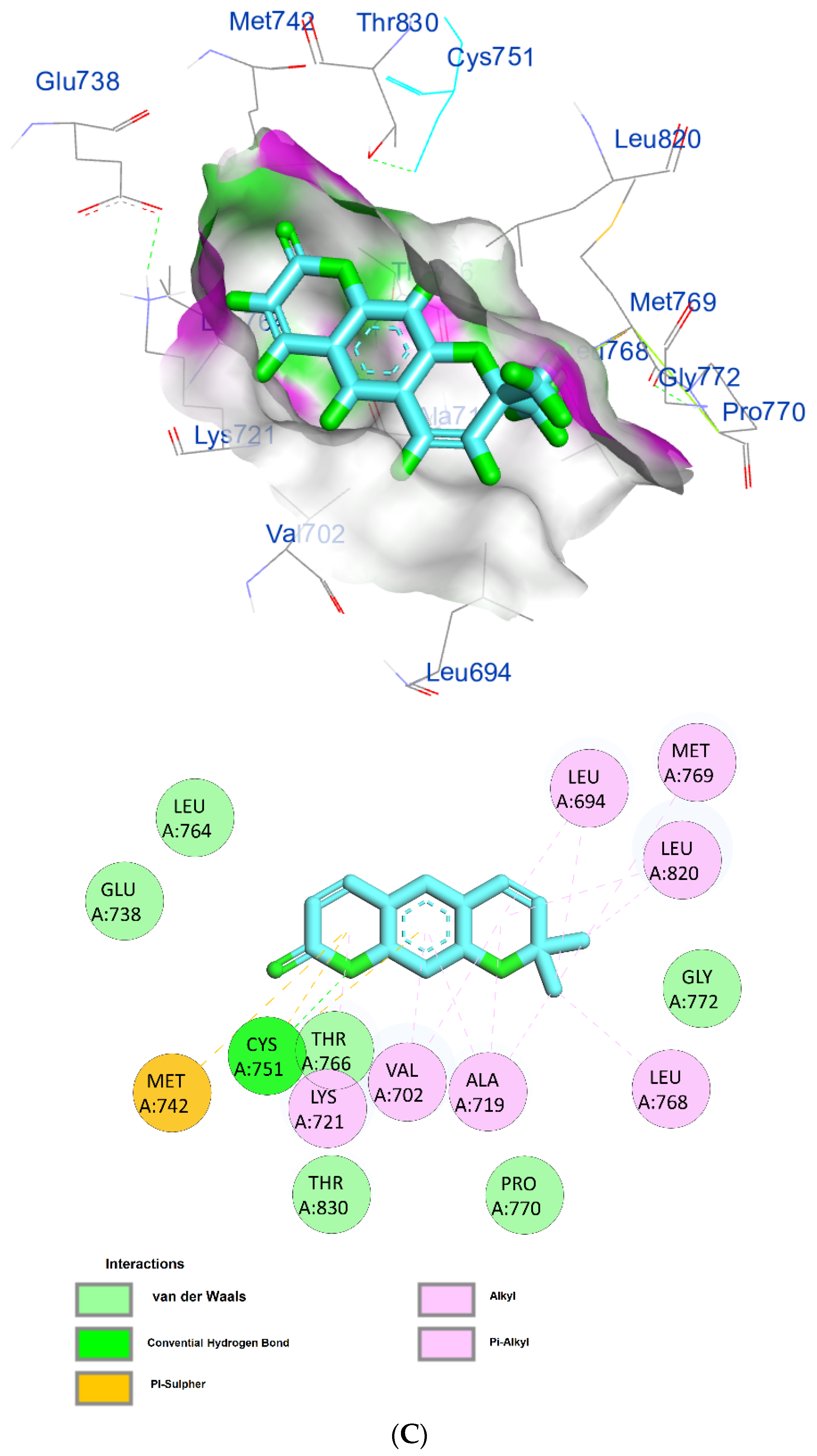
| Xanthyletine | −7.8 | (LEU694)—(Ligand) Alkyl and pi–alkyl hydrophobic interaction | 5.42 5.37 |
| (MET769)—(Ligand) Alkyl and pi–alkyl hydrophobic interaction | 5.36 | ||
| (LEU820)—(Ligand) Alkyl and pi–alkyl hydrophobic interaction | 3.89 5.28 | ||
| (LEU768)—(Ligand) Alkyl and pi–alkyl hydrophobic interaction | 4.19 | ||
| (ALA719)—(Ligand) Alkyl and pi–alkyl hydrophobic interaction | 3.95 3.76 4.04 | ||
| (VAL702)—(Ligand) Alkyl and pi–alkyl hydrophobic interaction | 4.54 4.61 | ||
| (LYS721)—(Ligand) Alkyl and pi–alkyl hydrophobic interaction | 4.31 | ||
| (MET742)—(Ligand) Alkyl and pi–alkyl hydrophobic interaction | 5.40 | ||
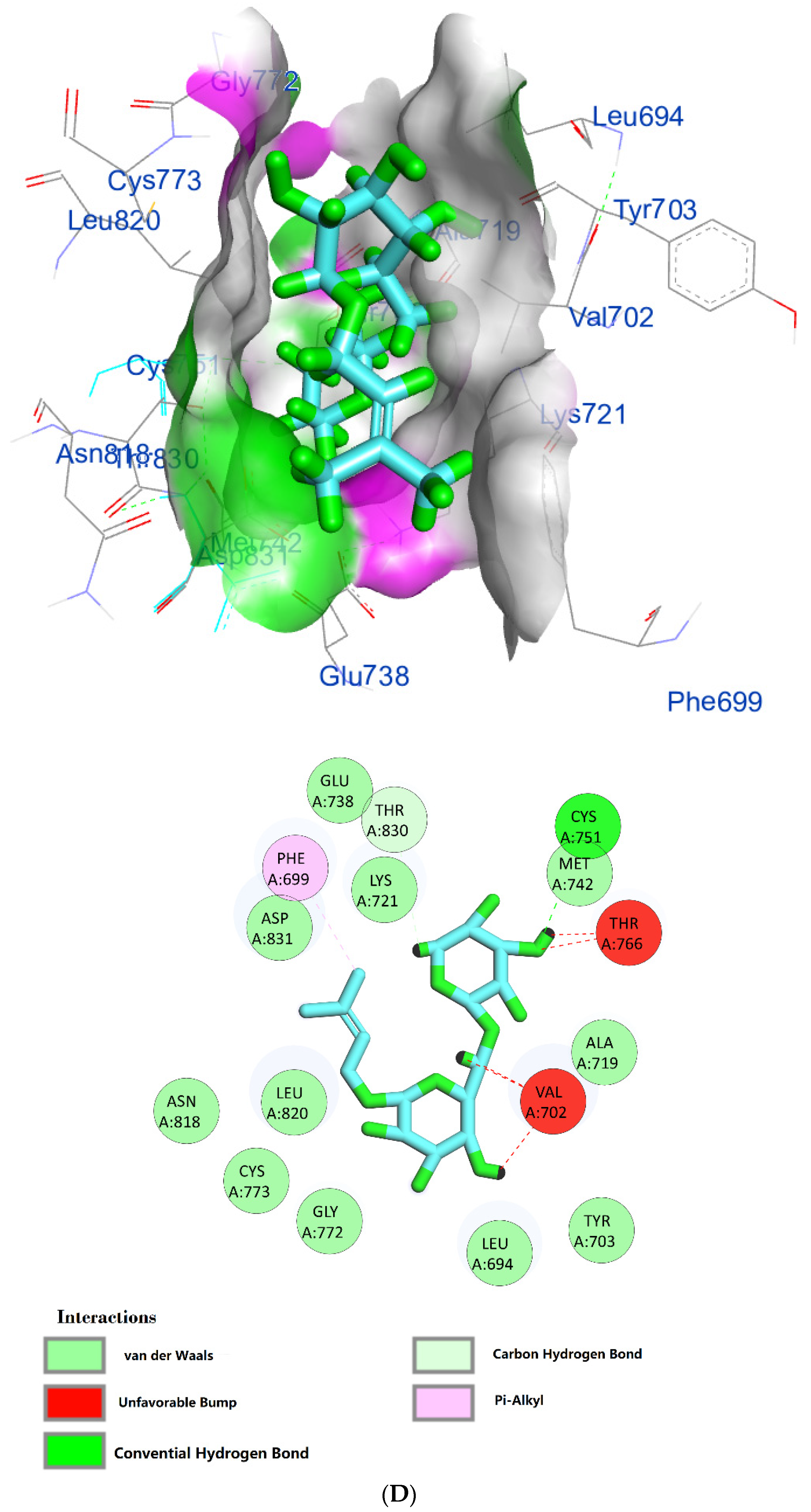
| Prenyl arabinosyl-(1->6)-glucoside | −7.3 | (CYS751)—(Ligand) H-bond interaction | 3.17 |
| (VAL702)—(Ligand) Hydrophobic interaction | 1.42 1.62 | ||
| (THR766)—(Ligand) Hydrophobic interaction | 2.07 2.15 | ||
| (PHE699)—(Ligand) Pi–alkyl hydrophobic interaction | 3.02 | ||
| (THR830)—(Ligand) C–H bond interaction | 3.03 | ||
| Furaneol 4-glucoside | −7.1 | (LYS721)—(Ligand) Conventional hydrogen bond and carboxylate salt bridge interactions | 2.36 4.55 |
| (ASP831)—(Ligand) 4 H-bonds and hydrophobic interactions | 3.10, 1.90, 1.75, 2.57 and 2.17 | ||
| (ALA719)—(Ligand) Hydrophobic interaction | 3.12 | ||
| (VAL702)—(Ligand) Alkyl hydrophobic interaction | 5.30 | ||
| Erlotinib | −6.8 | (GLU767) & (THR830)—(Ligand) Carbon hydrogen bond interaction | 3.15 4.07 |
| (MET769)—(Ligand) Conventional hydrogen bond interaction | |||
| (ALA719)—(Ligand) Alkyl and pi–alkyl hydrophobic interaction | 2.70 | ||
| (LEU764)—(Ligand) Alkyl and pi–alkyl hydrophobic interaction | 3.74 5.35 | ||
| (LYS721)—(Ligand) Alkyl and pi–alkyl hydrophobic interaction | 4.21 | ||
| (LEU694)—(Ligand) Pi–sigma interaction | 4.14 4.81 | ||
| (LEU920)—(Ligand) Pi–sigma interaction | 3.68 | ||
| (MET742)—(Ligand) Pi–sulfur interaction | 3.86 5.31 | ||
| Methyl orsellinate | −5.9 | (LYS721)—(Ligand) Alkyl and pi–alkyl interactions | 5.50 |
| (VAL702)—(Ligand) Alkyl and pi–alkyl interactions | 4.60 4.41 | ||
| (LEU820)—(Ligand) Pi–alkyl interaction | 4.10 4.87 | ||
| (ASP831)—(Ligand) Pi–anion interaction | 4.57 | ||
| Ginkgotoxin | −5.4 | (MET742)—(Ligand) Pi–sulfur and pi–anion interaction | 4.02 |
| (GLU738)—(Ligand) Pi–sulfur interaction | 6.00 3.25 | ||
| (PHE699)—(Ligand) Pi–alkyl interaction | 1.97 | ||
| (VAL702)—(Ligand) Alkyl interaction | 4.90 | ||
| (LYS721)—(Ligand) Unfavorable bump and pi–alkyl and alkyl interactions | 4.84 | ||
| (THR766)—(Ligand) Carbon–hydrogen bond interaction | 2.15 1.13 4.11 |
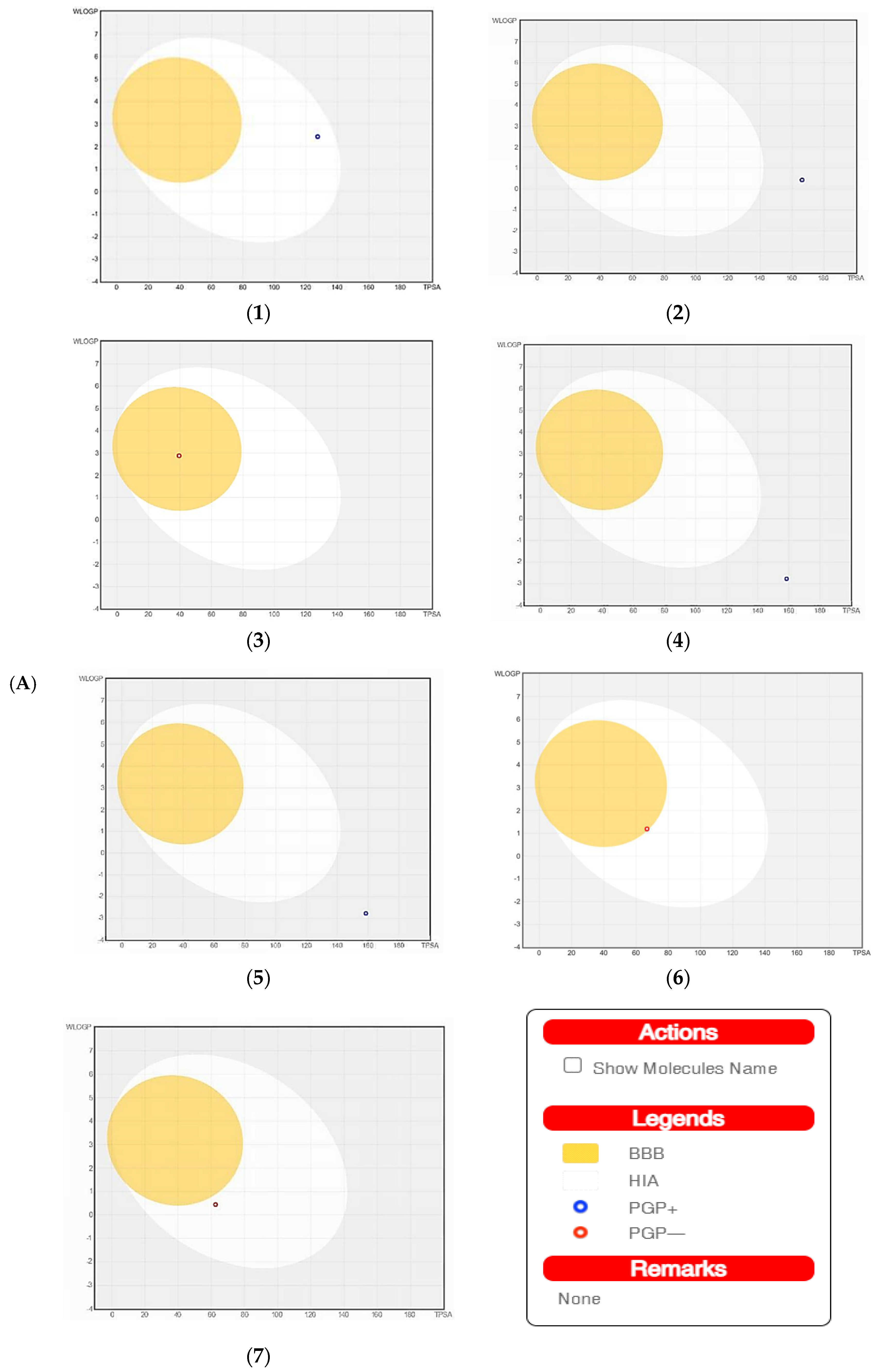
| Pharmacokinetics | Ouabagenin | Corchoroside A | Xanthyletine | Prenyl Arabinosyl-(1->6)-Glucoside | Furaneol 4-Glucoside | Methyl Orsellinate | Ginkgotoxin |
|---|---|---|---|---|---|---|---|
| GI absorption | High | High | High | Low | High | High | High |
| BBB permeant | No | No | Yes | No | No | Yes | No |
| P-gp substrate | Yes | Yes | No | Yes | Yes | No | No |
| CYP1A2 inhibitor | No | No | Yes | No | No | No | No |
| CYP2C19 inhibitor | No | No | Yes | No | No | No | No |
| CYP2C9 inhibitor | Yes | No | No | No | No | No | No |
| CYP2D6 inhibitor | No | No | No | No | No | No | No |
| CYP3A4 inhibitor | No | No | No | No | No | No | No |
| Log Kp (cm/s) | −8.62 | −8.64 | −5.68 | −10.49 | −8.72 | −6.4 | −7.35 |
| Drug-likeness | |||||||
| Lipinski | yes | yes | yes | Yes | yes | yes | yes |
| Ghose | yes | No | yes | No | No | Yes | Yes |
| Veber | yes | Yes | yes | No | Yes | Yes | Yes |
| Egan | yes | Yes | yes | No | Yes | Yes | Yes |
| Muegge | yes | Yes | yes | No | Yes | No | No |
| Bioavailability score | 0.55 | 0.55 | 0.55 | 0.55 | 0.55 | 0.55 | 0.55 |
| Medicinal chemistry | |||||||
| PAINS | 0 | 0 | 0 | 0 | 0 | 1 | 0 |
| Brenk | 1 | 2 | 1 | 1 | 0 | 1 | 0 |
| Lead-likeness | No | No | No | No | Yes | No | No |
| Synthetic accessibility | 5.64 | 7.24 | 3.24 | 5.77 | 5.23 | 1.55 | 1.78 |
| Toxicity Model | Ouabagenin | Corchoroside A | Xanthyletine | Prenyl arabinosyl-(1->6)-Glucoside | Furaneol 4-Glucoside | Methyl Orsellinate | Ginkgotoxin |
|---|---|---|---|---|---|---|---|
| AMES toxicity | No | No | No | No | No | Yes | No |
| Max. tolerated dose (human) (log mg/kg/day) | 0.276 | −1.229 | −0.001 | 1.32 | 1.414 | 0.983 | 1.059 |
| hERG I inhibitor | No | No | No | No | No | No | No |
| hERG II inhibitor | No | No | No | No | No | No | No |
| Hepatotoxicity | No | No | No | No | No | No | No |
| Skin sensitization | No | No | No | No | No | No | No |
| T. Pyriformis toxicity (log ug/L) | 0.285 | 0.285 | 0.871 | 0.29 | 0.285 | 0.585 | −0.258 |
| Minnow toxicity (log mM) | 4.323 | 3.312 | 0.997 | 7.58 | 5.153 | 2.16 | 2.65 |
| Oral rat acute toxicity (LD50) (mol/kg) | 2.802 | 2.854 | 2.307 | 1.95 | 1.983 | 1.731 | 1.963 |
| Oral rat chronic toxicity (LOAEL) (log mg/kg_bw/day) | 2.928 | 2.251 | 1.866 | 3.16 | 3.518 | 2.137 | 2.148 |
| Toxicophore rules | |||||||
| Genotoxic carcinogenicity rule | NO | NO | NO | NO | NO | NO | NO |
| Non-genotoxic carcinogenicity rule | NO | NO | yes | yes | NO | NO | NO |
| Time | Solvent A (Water 0.1% Formic Acid) | Solvent B (Acetonitrile 0.1% Formic Acid) |
|---|---|---|
| 0 | 98 | 2 |
| 2 | 98 | 2 |
| 15 | 90 | 10 |
| 35 | 80 | 20 |
| 60 | 50 | 50 |
| 80 | 30 | 70 |
| 100 | 0 | 100 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Abouzied, A.S.; Abd-Rabo, M.M.; Huwaimel, B.; Almahmoud, S.A.; Almarshdi, A.A.; Alharbi, F.M.; Alenzi, S.S.; Albsher, B.N.; Alafnan, A. In Silico Pharmacokinetic Profiling of the Identified Bioactive Metabolites of Pergularia tomentosa L. Latex Extract and In Vitro Cytotoxic Activity via the Induction of Caspase-Dependent Apoptosis with S-Phase Arrest. Pharmaceuticals 2022, 15, 1132. https://doi.org/10.3390/ph15091132
Abouzied AS, Abd-Rabo MM, Huwaimel B, Almahmoud SA, Almarshdi AA, Alharbi FM, Alenzi SS, Albsher BN, Alafnan A. In Silico Pharmacokinetic Profiling of the Identified Bioactive Metabolites of Pergularia tomentosa L. Latex Extract and In Vitro Cytotoxic Activity via the Induction of Caspase-Dependent Apoptosis with S-Phase Arrest. Pharmaceuticals. 2022; 15(9):1132. https://doi.org/10.3390/ph15091132
Chicago/Turabian StyleAbouzied, Amr S., Marwa M. Abd-Rabo, Bader Huwaimel, Suliman A. Almahmoud, Afnan Abdulkareem Almarshdi, Fai Mutaz Alharbi, Sulafa Salem Alenzi, Bayan Naef Albsher, and Ahmed Alafnan. 2022. "In Silico Pharmacokinetic Profiling of the Identified Bioactive Metabolites of Pergularia tomentosa L. Latex Extract and In Vitro Cytotoxic Activity via the Induction of Caspase-Dependent Apoptosis with S-Phase Arrest" Pharmaceuticals 15, no. 9: 1132. https://doi.org/10.3390/ph15091132
APA StyleAbouzied, A. S., Abd-Rabo, M. M., Huwaimel, B., Almahmoud, S. A., Almarshdi, A. A., Alharbi, F. M., Alenzi, S. S., Albsher, B. N., & Alafnan, A. (2022). In Silico Pharmacokinetic Profiling of the Identified Bioactive Metabolites of Pergularia tomentosa L. Latex Extract and In Vitro Cytotoxic Activity via the Induction of Caspase-Dependent Apoptosis with S-Phase Arrest. Pharmaceuticals, 15(9), 1132. https://doi.org/10.3390/ph15091132