
Antiviral Effect of Stenocline ericoides DC. and Stenocline inuloides DC., Two Flavonoid-Rich Endemic Plants from Madagascar, against Dengue and Zika Viruses

 , , ,
, , ,  ,
,
Abstract
1. Introduction
2. Results
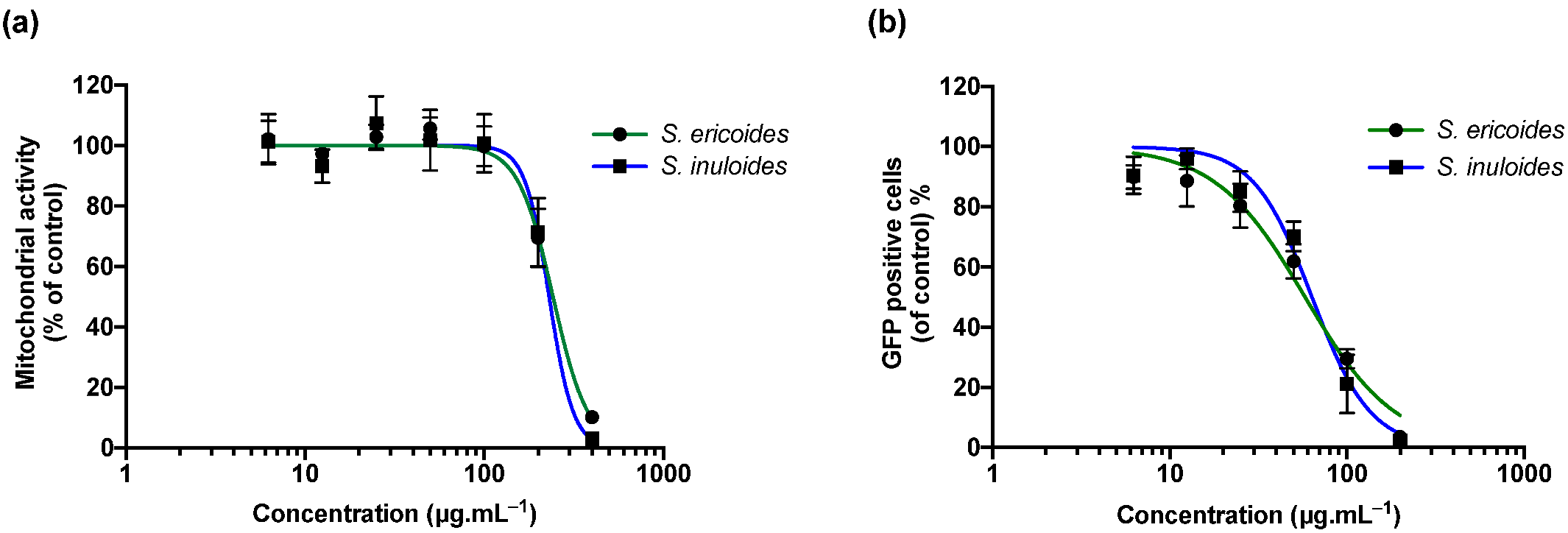
2.1. S. ericoides and S. inuloides Extracts Inhibit ZIKV Infection in A549 Human Cells at Non Cytotoxic Concentrations
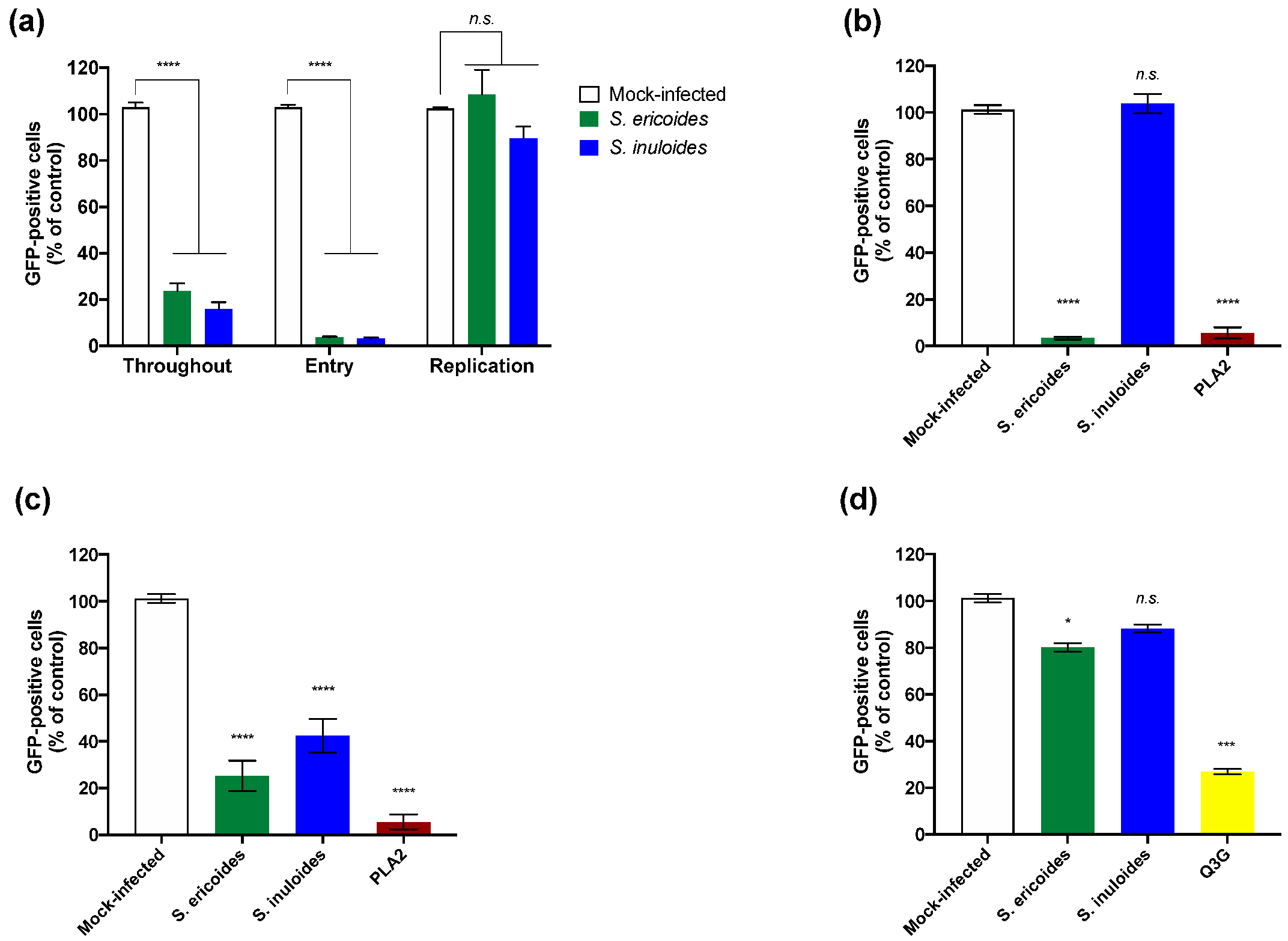
2.2. Inhibition of Viral Infection Mediated by S. ericoides and S. inuloides Extracts Involves the Early Stage of the ZIKV Infectious Cycle
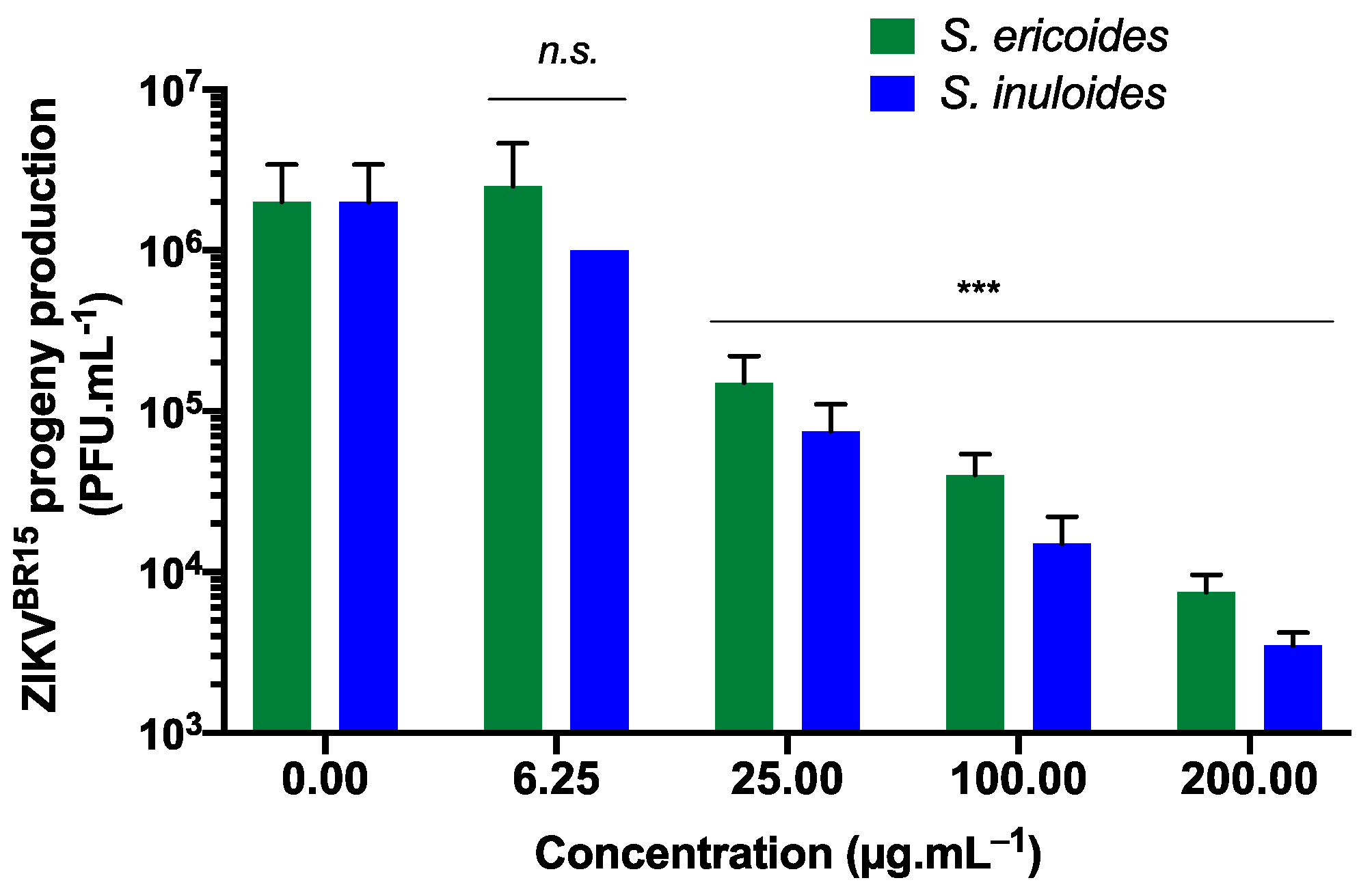
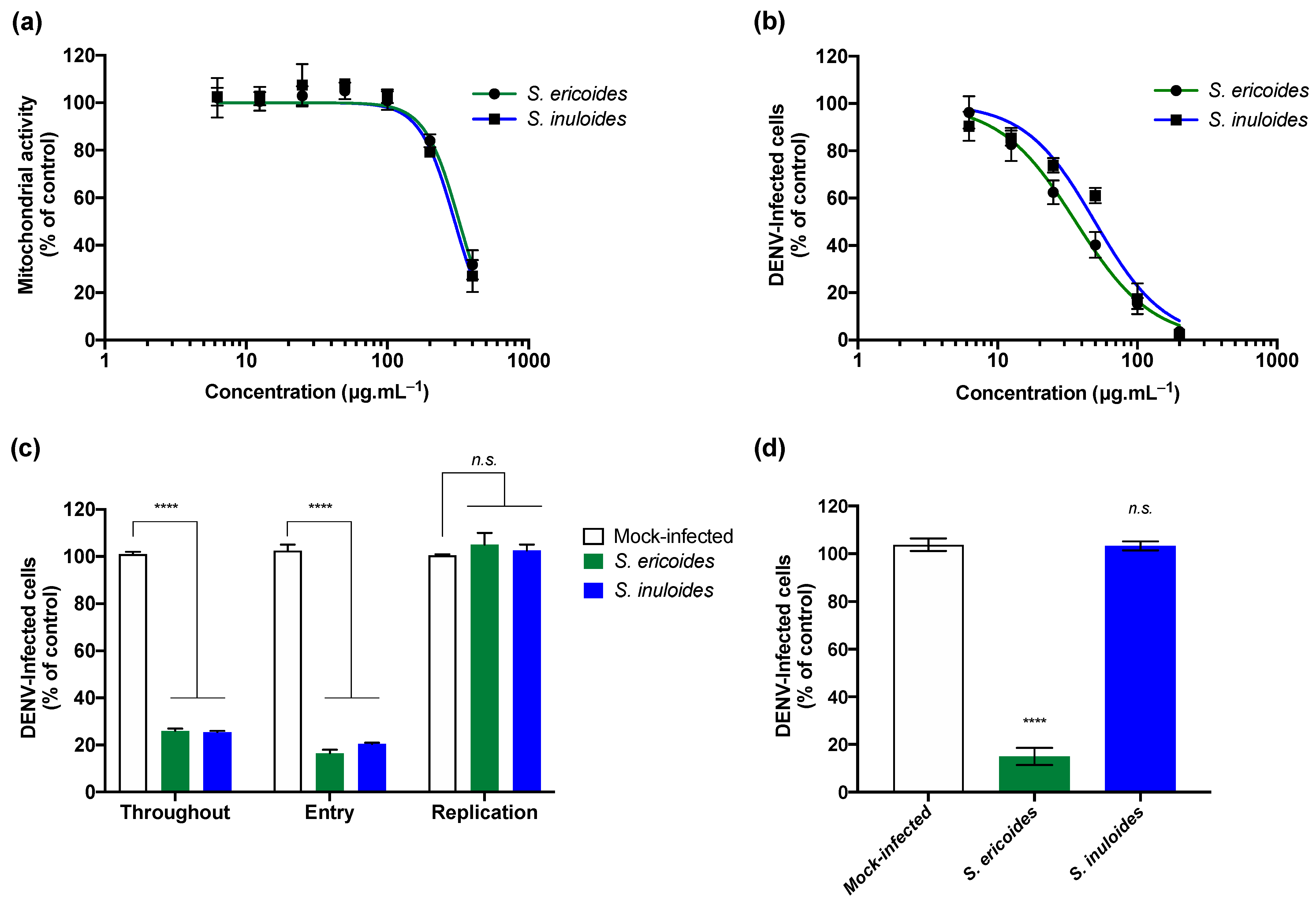
2.3. Stenocline ericoides and Stenocline inuloides Extracts Exert Antiviral Effect against Clinical Isolate of Dengue Virus
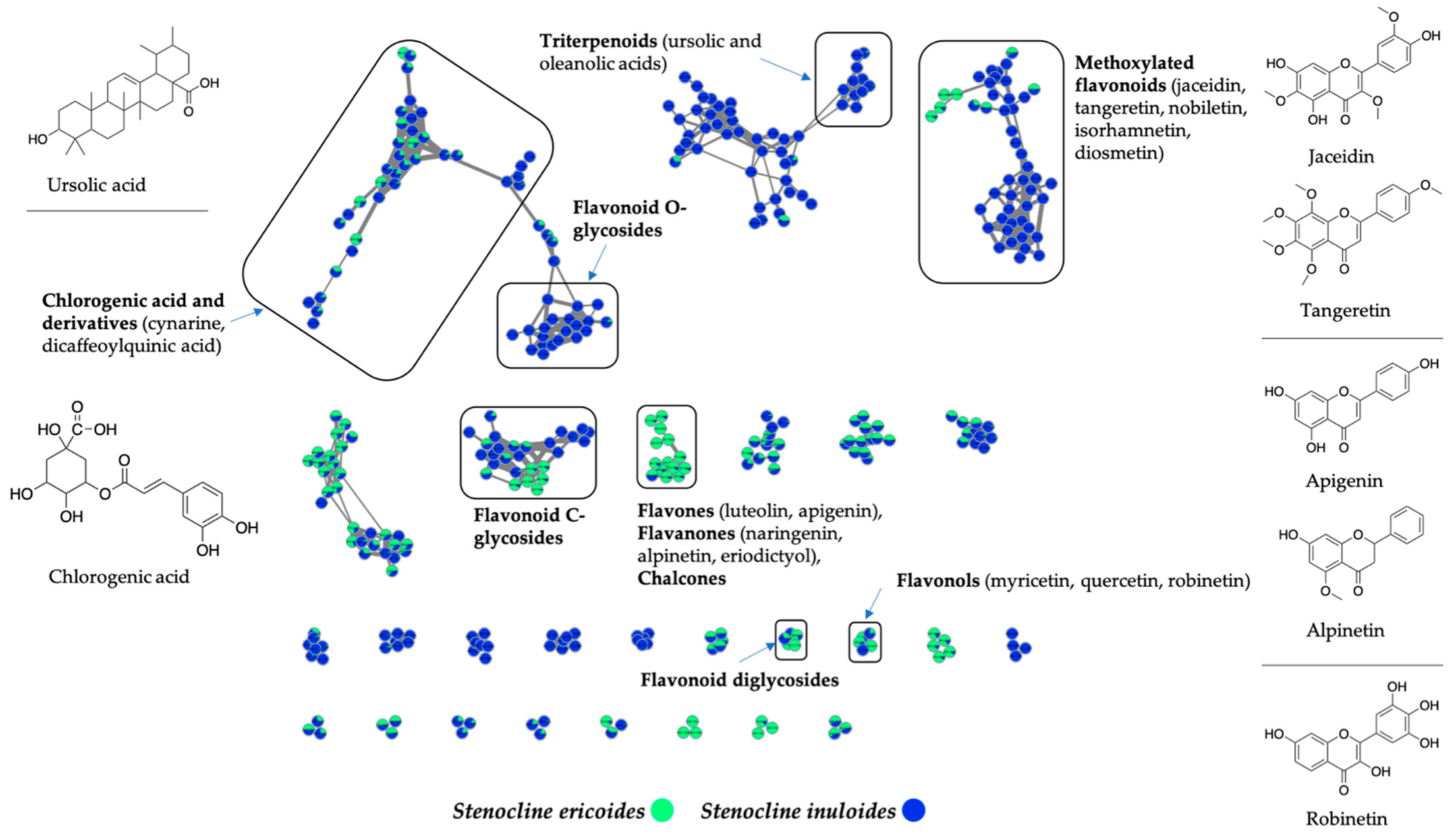
2.4. Stenocline ericoides and Stenocline inuloides Extracts Are Rich in Polyphenols and Flavonoid Compounds
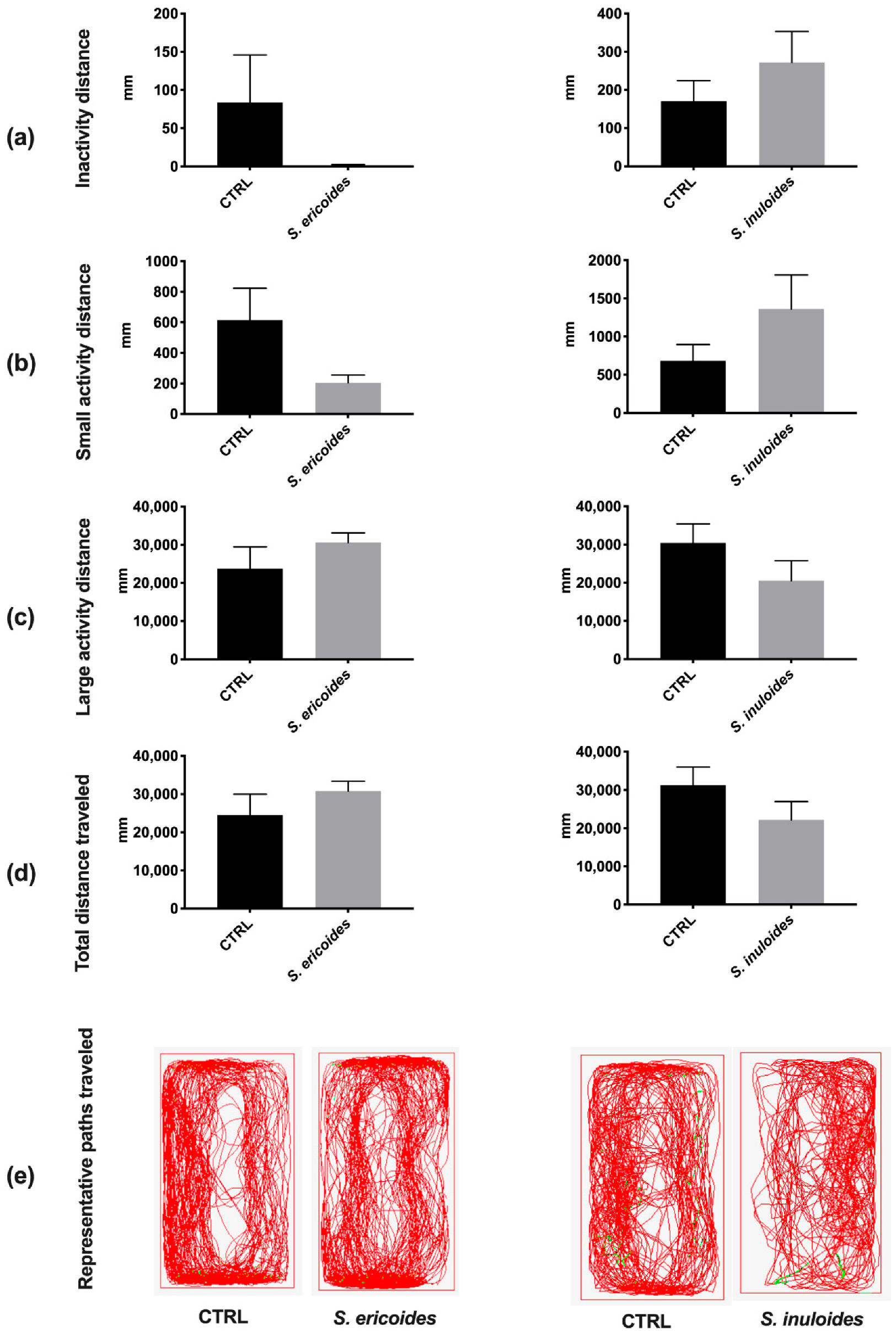
2.5. Stenocline ericoides and Stenocline inuloides Do Not Exert in Vivo Toxicity in Zebrafish
3. Discussion
4. Materials and Methods
4.1. Plant Materials and Samples Preparation
4.2. Cells, Viruses and Reagents
4.3. MTT Assay
4.4. Flow Cytometry Assay
4.5. Virus Inactivation Assay
4.6. Plaque-Forming Assay
4.7. Zebrafish Maintenance, Intraperitoneal Injection and Behavior Monitoring
4.8. Data-Dependent LC-HRMS2 Analysis
4.9. Progenesis QI Preprocessing
4.10. Molecular Networking Analysis
4.11. Data Analysis
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Atanasov, A.G.; Zotchev, S.B.; Dirsch, V.M.; Orhan, I.E.; Banach, M.; Rollinger, J.M.; Barreca, D.; Weckwerth, W.; Bauer, R.; Bayer, E.A.; et al. Natural products in drug discovery: Advances and opportunities. Nat. Rev. Drug Discov. 2021, 20, 200–216. [Google Scholar] [CrossRef] [PubMed]
- Kitazato, K.; Wang, Y.; Kobayashi, N. Viral infectious disease and natural products with antiviral activity. Drug Discov. Ther. 2007, 1, 14–22. [Google Scholar] [PubMed]
- Martinez, J.P.; Sasse, F.; Brönstrup, M.; Diez, J.; Meyerhans, A. Antiviral drug discovery: Broad-spectrum drugs from nature. Nat. Prod. Rep. 2015, 32, 29–48. [Google Scholar] [CrossRef]
- Mattio, L.M.; Catinella, G.; Pinto, A.; Dallavalle, S. Natural and nature-inspired stilbenoids as antiviral agents. Eur. J. Med. Chem. 2020, 202, 112541. [Google Scholar] [CrossRef] [PubMed]
- Goh, V.S.L.; Mok, C.K.; Chu, J.J.H. Antiviral Natural Products for Arbovirus Infections. Molecules 2020, 25, 2796. [Google Scholar] [CrossRef]
- Haddad, J.G.; Gadea, G.; Desprès, P.; El Kalamouni, C. Chapter 38—Medicinal plants as promising source of natural antiviral substances against Zika virus. In Zika Virus Impact, Diagnosis, Control, and Models; Martin, C.R., Hollins Martin, C.J., Preedy, V.R., Rajendram, R., Eds.; Academic Press: Cambridge, MA, USA, 2021; pp. 397–407. [Google Scholar]
- Bos, S.; Gadea, G.; Despres, P. Dengue: A growing threat requiring vaccine development for disease prevention. Pathog. Glob Health 2018, 112, 294–305. [Google Scholar] [CrossRef]
- Hafsia, S.; Haramboure, M.; Wilkinson, D.A.; Baldet, T.; Yemadje-Menudier, L.; Vincent, M.; Tran, A.; Atyame, C.; Mavingui, P. Overview of dengue outbreaks in the southwestern Indian Ocean and analysis of factors involved in the shift toward endemicity in Reunion Island: A systematic review. PLoS Negl. Trop. Dis. 2022, 16, e0010547. [Google Scholar] [CrossRef]
- Ferraris, P.; Yssel, H.; Misse, D. Zika virus infection: An update. Microbes. Infect. 2019, 21, 353–360. [Google Scholar] [CrossRef]
- Lee, C.Y.; Ng, L.F.P. Zika virus: From an obscurity to a priority. Microbes. Infect. 2018, 20, 635–645. [Google Scholar] [CrossRef]
- Cao-Lormeau, V.-M.; Blake, A.; Mons, S.; Lastère, S.; Roche, C.; Vanhomwegen, J.; Dub, T.; Baudouin, L.; Teissier, A.; Larre, P.; et al. Guillain-Barré Syndrome outbreak associated with Zika virus infection in French Polynesia: A case-control study. Lancet 2016, 387, 1531–1539. [Google Scholar] [CrossRef]
- Cauchemez, S.; Besnard, M.; Bompard, P.; Dub, T.; Guillemette-Artur, P.; Eyrolle-Guignot, D.; Salje, H.; Van Kerkhove, M.D.; Abadie, V.; Garel, C.; et al. Association between Zika virus and microcephaly in French Polynesia, 2013–2015: A retrospective study. Lancet 2016, 387, 2125–2132. [Google Scholar] [CrossRef] [PubMed]
- Duggal, N.K.; Ritter, J.M.; Pestorius, S.E.; Zaki, S.R.; Davis, B.S.; Chang, G.J.; Bowen, R.A.; Brault, A.C. Frequent Zika Virus Sexual Transmission and Prolonged Viral RNA Shedding in an Immunodeficient Mouse Model. Cell Rep. 2017, 18, 1751–1760. [Google Scholar] [CrossRef] [PubMed]
- Perera-Lecoin, M.; Meertens, L.; Carnec, X.; Amara, A. Flavivirus Entry Receptors: An Update. Viruses 2013, 6, 69–88. [Google Scholar] [CrossRef]
- Bibi Sadeer, N.; Haddad, J.G.; Oday Ezzat, M.; Desprès, P.; Abdallah, H.H.; Zengin, G.; Uysal, A.; El Kalamouni, C.; Gallo, M.; Montesano, D.; et al. Bruguiera gymnorhiza (L.) Lam. at the Forefront of Pharma to Confront Zika Virus and Microbial Infections-An In Vitro and In Silico Perspective. Molecules 2021, 26, 5768. [Google Scholar] [CrossRef] [PubMed]
- Alagarasu, K.; Patil, P.; Kaushik, M.; Chowdhury, D.; Joshi, R.K.; Hegde, H.V.; Kakade, M.B.; Hoti, S.L.; Cherian, S.; Parashar, D. In Vitro Antiviral Activity of Potential Medicinal Plant Extracts Against Dengue and Chikungunya Viruses. Front. Cell Infect. Microbiol. 2022, 12, 866452. [Google Scholar] [CrossRef]
- Clain, E.; Haddad, J.G.; Koishi, A.C.; Sinigaglia, L.; Rachidi, W.; Desprès, P.; Duarte Dos Santos, C.N.; Guiraud, P.; Jouvenet, N.; El Kalamouni, C. The Polyphenol-Rich Extract from Psiloxylon mauritianum, an Endemic Medicinal Plant from Reunion Island, Inhibits the Early Stages of Dengue and Zika Virus Infection. Int. J. Mol. Sci. 2019, 20, 1860. [Google Scholar] [CrossRef]
- Clain, E.; Sinigaglia, L.; Koishi, A.C.; Gorgette, O.; Gadea, G.; Viranaicken, W.; Krejbich-Trotot, P.; Mavingui, P.; Desprès, P.; Nunes Duarte Dos Santos, C.; et al. Extract from Aphloia theiformis, an edible indigenous plant from Reunion Island, impairs Zika virus attachment to the host cell surface. Sci. Rep. 2018, 8, 10856. [Google Scholar] [CrossRef]
- Haddad, J.G.; Koishi, A.C.; Gaudry, A.; Nunes Duarte Dos Santos, C.; Viranaicken, W.; Desprès, P.; El Kalamouni, C. Doratoxylon apetalum, an Indigenous Medicinal Plant from Mascarene Islands, Is a Potent Inhibitor of Zika and Dengue Virus Infection in Human Cells. Int. J. Mol. Sci. 2019, 20, 2382. [Google Scholar] [CrossRef]
- Gaudry, A.; Bos, S.; Viranaicken, W.; Roche, M.; Krejbich-Trotot, P.; Gadea, G.; Desprès, P.; El-Kalamouni, C. The Flavonoid Isoquercitrin Precludes Initiation of Zika Virus Infection in Human Cells. Int. J. Mol. Sci. 2018, 19, 1093. [Google Scholar] [CrossRef]
- Haddad, J.G.; Picard, M.; Bénard, S.; Desvignes, C.; Desprès, P.; Diotel, N.; El Kalamouni, C. Ayapana triplinervis Essential Oil and Its Main Component Thymohydroquinone Dimethyl Ether Inhibit Zika Virus at Doses Devoid of Toxicity in Zebrafish. Molecules 2019, 24, 3447. [Google Scholar] [CrossRef]
- Helmstetter, A.J.; Cable, S.; Rakotonasolo, F.; Rabarijaona, R.; Rakotoarinivo, M.; Eiserhardt, W.L.; Baker, W.J.; Papadopulos, A.S.T. The demographic history of Madagascan micro-endemics: Have rare species always been rare? Proc. Biol. Sci. 2021, 288, 20210957. [Google Scholar] [CrossRef]
- Riondato, I.; Donno, D.; Roman, A.; Razafintsalama, V.E.; Petit, T.; Mellano, M.G.; Torti, V.; De Biaggi, M.; Rakotoniaina, E.N.; Giacoma, C.; et al. First ethnobotanical inventory and phytochemical analysis of plant species used by indigenous people living in the Maromizaha forest, Madagascar. J. Ethnopharmacol. 2019, 232, 73–89. [Google Scholar] [CrossRef] [PubMed]
- Hou, Y.; Harinantenaina, L. New and bioactive natural products isolated from madagascar plants and marine organisms. Curr. Med. Chem. 2010, 17, 1191–1219. [Google Scholar] [CrossRef] [PubMed]
- Cao, S.; Kingston, D.G. Biodiversity conservation and drug discovery: Can they be combined? The Suriname and Madagascar experiences. Pharm. Biol. 2009, 47, 809–823. [Google Scholar] [CrossRef] [PubMed][Green Version]
- Barrett, M.A.; Brown, J.L.; Morikawa, M.K.; Labat, J.N.; Yoder, A.D. Conservation. CITES designation for endangered rosewood in Madagascar. Science 2010, 328, 1109–1110. [Google Scholar] [CrossRef]
- Consiglio, T.; Schatz, G.E.; McPherson, G.; Lowry, P.P., 2nd; Rabenantoandro, J.; Rogers, Z.S.; Rabevohitra, R.; Rabehevitra, D. Deforestation and plant diversity of Madagascar’s littoral forests. Conserv. Biol. 2006, 20, 1799–1803. [Google Scholar] [CrossRef]
- Guillaumet, J.-L. CHAPTER 2-The Vegetation: An Extraordinary Diversity. In Key Environments: Madagascar; Jolly, A., OberlÉ, P., Albignac, R., Eds.; Pergamon: Amsterdam, The Netherlands, 1984; pp. 27–54. [Google Scholar]
- Frumence, E.; Roche, M.; Krejbich-Trotot, P.; El-Kalamouni, C.; Nativel, B.; Rondeau, P.; Missé, D.; Gadea, G.; Viranaicken, W.; Desprès, P. The South Pacific epidemic strain of Zika virus replicates efficiently in human epithelial A549 cells leading to IFN-β production and apoptosis induction. Virology 2016, 493, 217–226. [Google Scholar] [CrossRef] [PubMed]
- Gadea, G.; Bos, S.; Krejbich-Trotot, P.; Clain, E.; Viranaicken, W.; El-Kalamouni, C.; Mavingui, P.; Desprès, P. A robust method for the rapid generation of recombinant Zika virus expressing the GFP reporter gene. Virology 2016, 497, 157–162. [Google Scholar] [CrossRef]
- Bos, S.; Viranaicken, W.; Turpin, J.; El-Kalamouni, C.; Roche, M.; Krejbich-Trotot, P.; Desprès, P.; Gadea, G. The structural proteins of epidemic and historical strains of Zika virus differ in their ability to initiate viral infection in human host cells. Virology 2018, 516, 265–273. [Google Scholar] [CrossRef]
- Chen, M.; Aoki-Utsubo, C.; Kameoka, M.; Deng, L.; Terada, Y.; Kamitani, W.; Sato, K.; Koyanagi, Y.; Hijikata, M.; Shindo, K.; et al. Broad-spectrum antiviral agents: Secreted phospholipase A(2) targets viral envelope lipid bilayers derived from the endoplasmic reticulum membrane. Sci. Rep. 2017, 7, 15931. [Google Scholar] [CrossRef]
- Pascalis, H.; Turpin, J.; Roche, M.; Krejbich, P.; Gadea, G.; Nten, C.A.; Desprès, P.; Mavingui, P. The epidemic of Dengue virus type-2 Cosmopolitan genotype on Reunion Island relates to its active circulation in the Southwestern Indian Ocean neighboring islands. Heliyon 2019, 5, e01455. [Google Scholar] [CrossRef] [PubMed]
- Wang, M.; Carver, J.J.; Phelan, V.V.; Sanchez, L.M.; Garg, N.; Peng, Y.; Nguyen, D.D.; Watrous, J.; Kapono, C.A.; Luzzatto-Knaan, T.; et al. Sharing and community curation of mass spectrometry data with Global Natural Products Social Molecular Networking. Nat. Biotechnol. 2016, 34, 828–837. [Google Scholar] [CrossRef] [PubMed]
- Olivon, F.; Elie, N.; Grelier, G.; Roussi, F.; Litaudon, M.; Touboul, D. MetGem Software for the Generation of Molecular Networks Based on the t-SNE Algorithm. Anal. Chem. 2018, 90, 13900–13908. [Google Scholar] [CrossRef] [PubMed]
- Global Natural Products Social Molecular Networking. Available online: https://gnps.ucsd.edu/ProteoSAFe/libraries.jsp (accessed on 20 June 2022).
- Hill, A.J.; Teraoka, H.; Heideman, W.; Peterson, R.E. Zebrafish as a model vertebrate for investigating chemical toxicity. Toxicol. Sci. 2005, 86, 6–19. [Google Scholar] [CrossRef]
- Howe, K.; Clark, M.D.; Torroja, C.F.; Torrance, J.; Berthelot, C.; Muffato, M.; Collins, J.E.; Humphray, S.; McLaren, K.; Matthews, L.; et al. The zebrafish reference genome sequence and its relationship to the human genome. Nature 2013, 496, 498–503. [Google Scholar] [CrossRef]
- Tamkutė, L.; Haddad, J.G.; Diotel, N.; Desprès, P.; Venskutonis, P.R.; El Kalamouni, C. Cranberry Pomace Extract Exerts Antiviral Activity against Zika and Dengue Virus at Safe Doses for Adult Zebrafish. Viruses 2022, 14, 1101. [Google Scholar] [CrossRef]
- Langendries, L.; Jacobs, S.; Abdelnabi, R.; Verwimp, S.; Kaptein, S.; Baatsen, P.; Van Mellaert, L.; Delang, L. Perturbation of Alphavirus and Flavivirus Infectivity by Components of the Bacterial Cell Wall. J. Virol. 2022, 96, e0006022. [Google Scholar] [CrossRef]
- Sharma, N.; Murali, A.; Singh, S.K.; Giri, R. Epigallocatechin gallate, an active green tea compound inhibits the Zika virus entry into host cells via binding the envelope protein. Int. J. Biol. Macromol. 2017, 104, 1046–1054. [Google Scholar] [CrossRef]
- Vázquez-Calvo, Á.; Jiménez De Oya, N.; Martín-Acebes, M.A.; Garcia-Moruno, E.; Saiz, J.-C. Antiviral Properties of the Natural Polyphenols Delphinidin and Epigallocatechin Gallate against the Flaviviruses West Nile Virus, Zika Virus, and Dengue Virus. Front. Microbiol. 2017, 8, 1314. [Google Scholar] [CrossRef]
- Balasubramanian, A.; Pilankatta, R.; Teramoto, T.; Sajith, A.M.; Nwulia, E.; Kulkarni, A.; Padmanabhan, R. Inhibition of dengue virus by curcuminoids. Antivir. Res. 2019, 162, 71–78. [Google Scholar] [CrossRef]
- Byler, K.G.; Ogungbe, I.V.; Setzer, W.N. In-silico screening for anti-Zika virus phytochemicals. J. Mol. Graph. Model. 2016, 69, 78–91. [Google Scholar] [CrossRef] [PubMed]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Number of Fish Alive | |||||||
|---|---|---|---|---|---|---|---|
| Number of Injected Fish | 1 dpi | 2 dpi | 3 dpi | 4 dpi | 5 dpi | Survival Rate at 5 dpi (%) | |
| 1× PBS (vehicle) | 7 | 7 | 7 | 7 | 7 | 7 | 100 |
| S. ericoides | 5 | 5 | 5 | 5 | 5 | 5 | 100 |
| S. inuloides | 7 | 7 | 7 | 7 | 7 | 7 | 100 |
| Feeding behavior | normal | normal | normal | normal | normal | ||
| Locomotor behavior | normal | normal | normal | normal | normal | ||
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ramiharimanana, F.D.; Haddad, J.G.; Andrianavalonirina, M.A.; Apel, C.; Olivon, F.; Diotel, N.; Desprès, P.; Ramanandraibe, V.V.; El Kalamouni, C. Antiviral Effect of Stenocline ericoides DC. and Stenocline inuloides DC., Two Flavonoid-Rich Endemic Plants from Madagascar, against Dengue and Zika Viruses. Pharmaceuticals 2022, 15, 1500. https://doi.org/10.3390/ph15121500
Ramiharimanana FD, Haddad JG, Andrianavalonirina MA, Apel C, Olivon F, Diotel N, Desprès P, Ramanandraibe VV, El Kalamouni C. Antiviral Effect of Stenocline ericoides DC. and Stenocline inuloides DC., Two Flavonoid-Rich Endemic Plants from Madagascar, against Dengue and Zika Viruses. Pharmaceuticals. 2022; 15(12):1500. https://doi.org/10.3390/ph15121500
Chicago/Turabian StyleRamiharimanana, Fenia D., Juliano G. Haddad, Maminiaina A. Andrianavalonirina, Cécile Apel, Florent Olivon, Nicolas Diotel, Philippe Desprès, Voahangy Vestalys Ramanandraibe, and Chaker El Kalamouni. 2022. "Antiviral Effect of Stenocline ericoides DC. and Stenocline inuloides DC., Two Flavonoid-Rich Endemic Plants from Madagascar, against Dengue and Zika Viruses" Pharmaceuticals 15, no. 12: 1500. https://doi.org/10.3390/ph15121500
APA StyleRamiharimanana, F. D., Haddad, J. G., Andrianavalonirina, M. A., Apel, C., Olivon, F., Diotel, N., Desprès, P., Ramanandraibe, V. V., & El Kalamouni, C. (2022). Antiviral Effect of Stenocline ericoides DC. and Stenocline inuloides DC., Two Flavonoid-Rich Endemic Plants from Madagascar, against Dengue and Zika Viruses. Pharmaceuticals, 15(12), 1500. https://doi.org/10.3390/ph15121500