
Effects of o,p’-DDE, a Mitotane Metabolite, in an Adrenocortical Carcinoma Cell Line
 , ,
, ,
Abstract
1. Introduction
2. Results
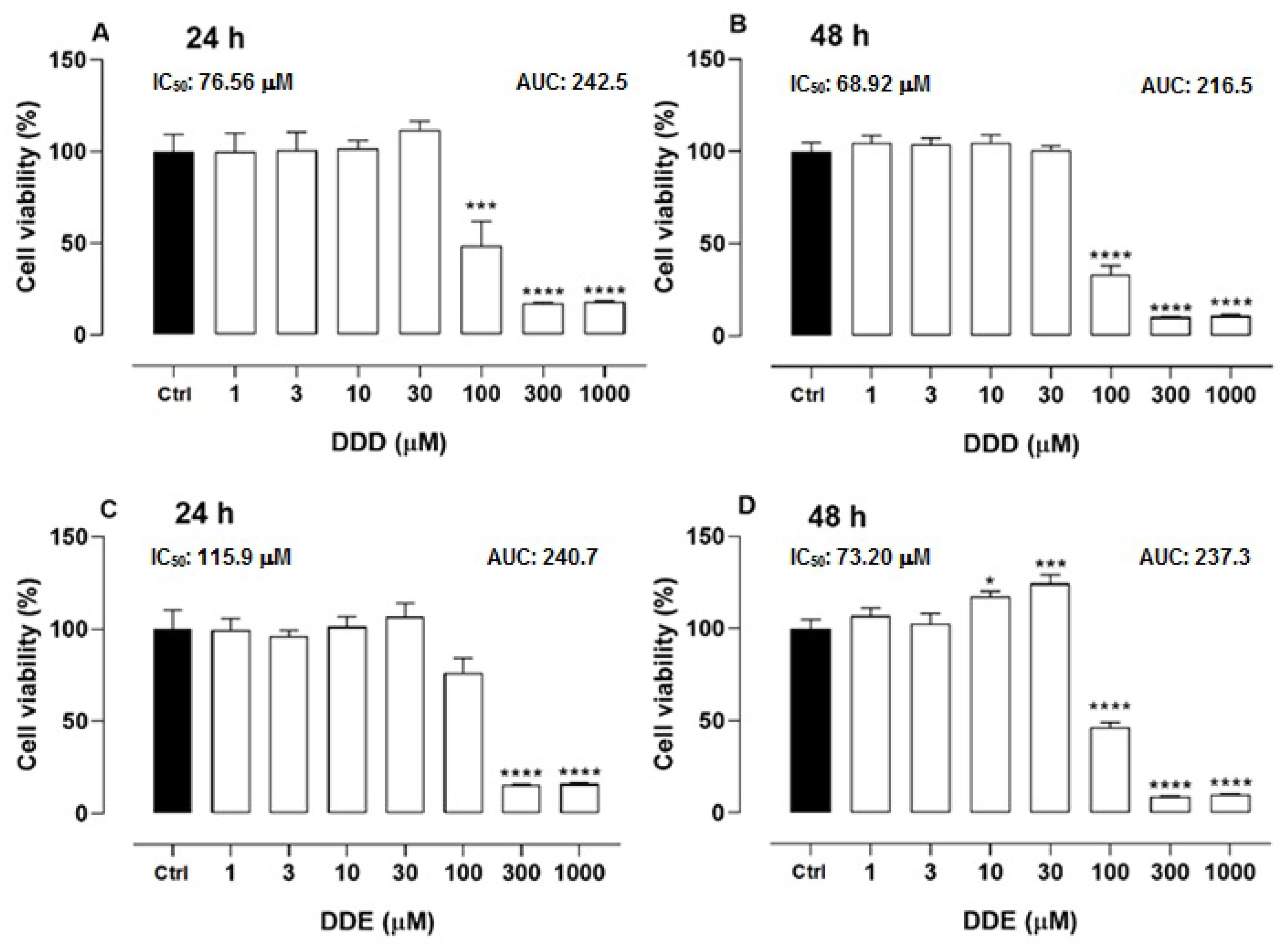
2.1. Cell Viability of o,p’-DDD and o,p’-DDE in H295R Cell Line
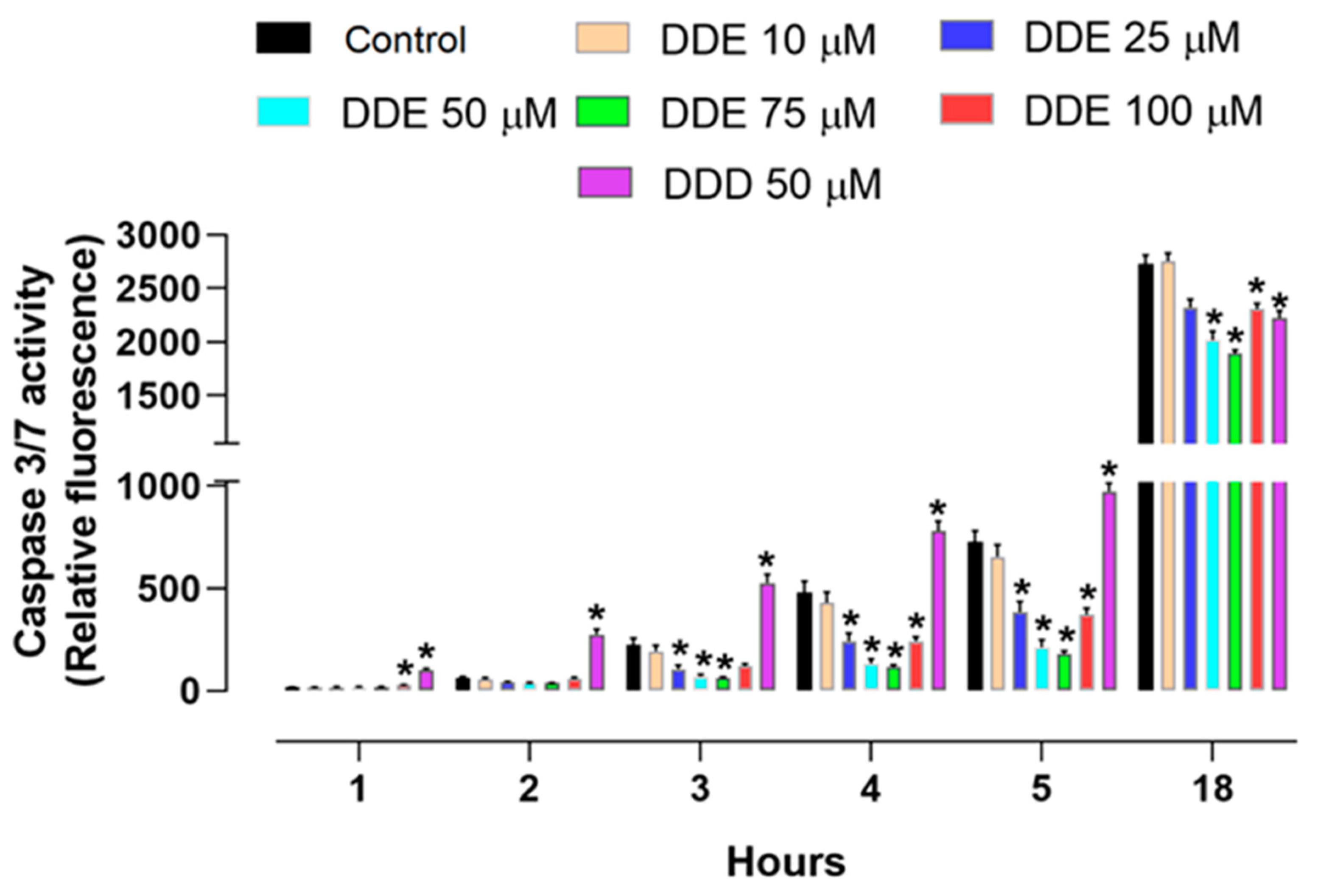
2.2. Effect of o,p’-DDD and o,p’-DDE on Caspase 3/7 Activity in H295R Cell Line
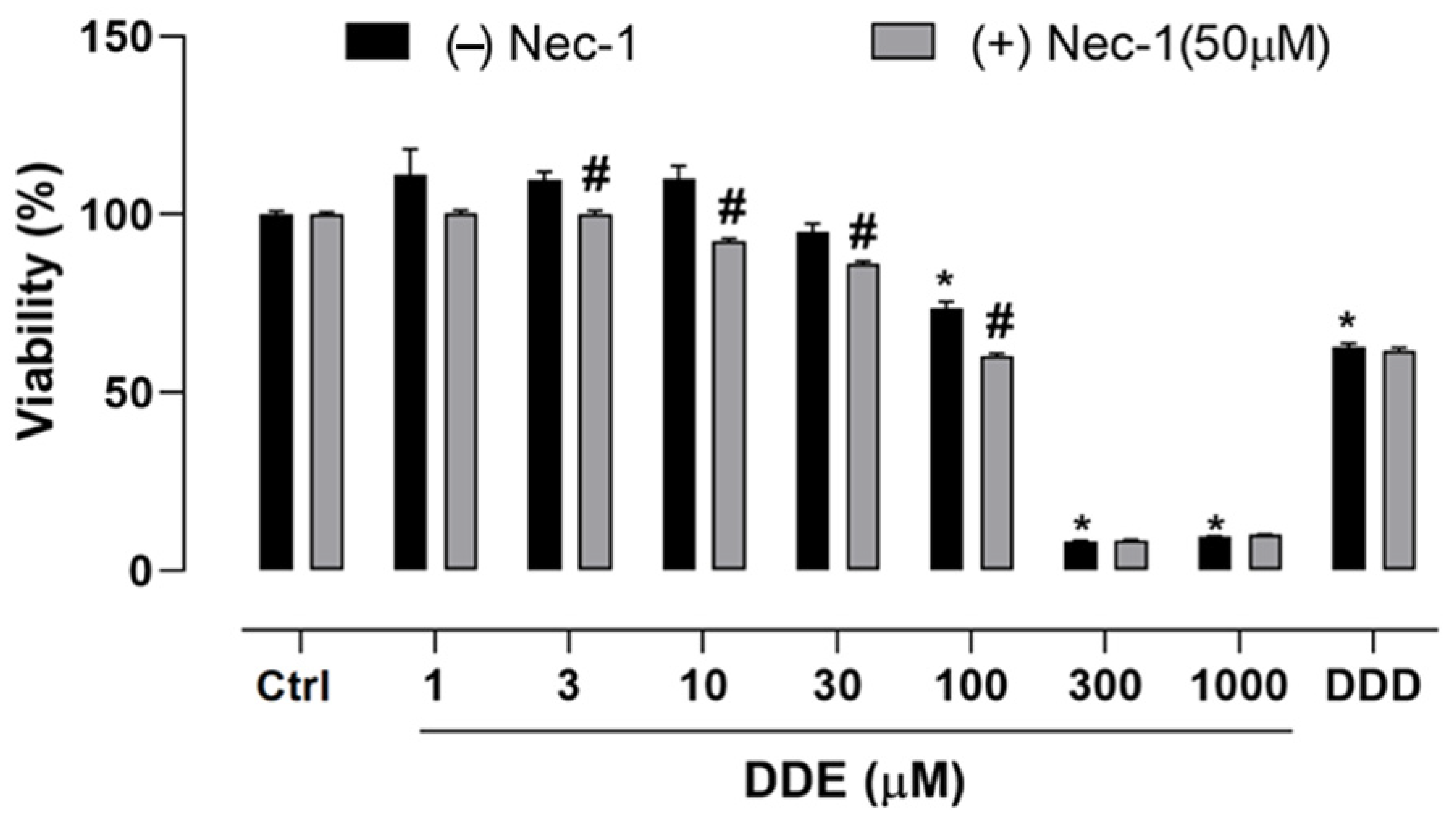
2.3. Effect of o,p’-DDD and o,p’-DDE on Necroptosis in H295R Cell Line
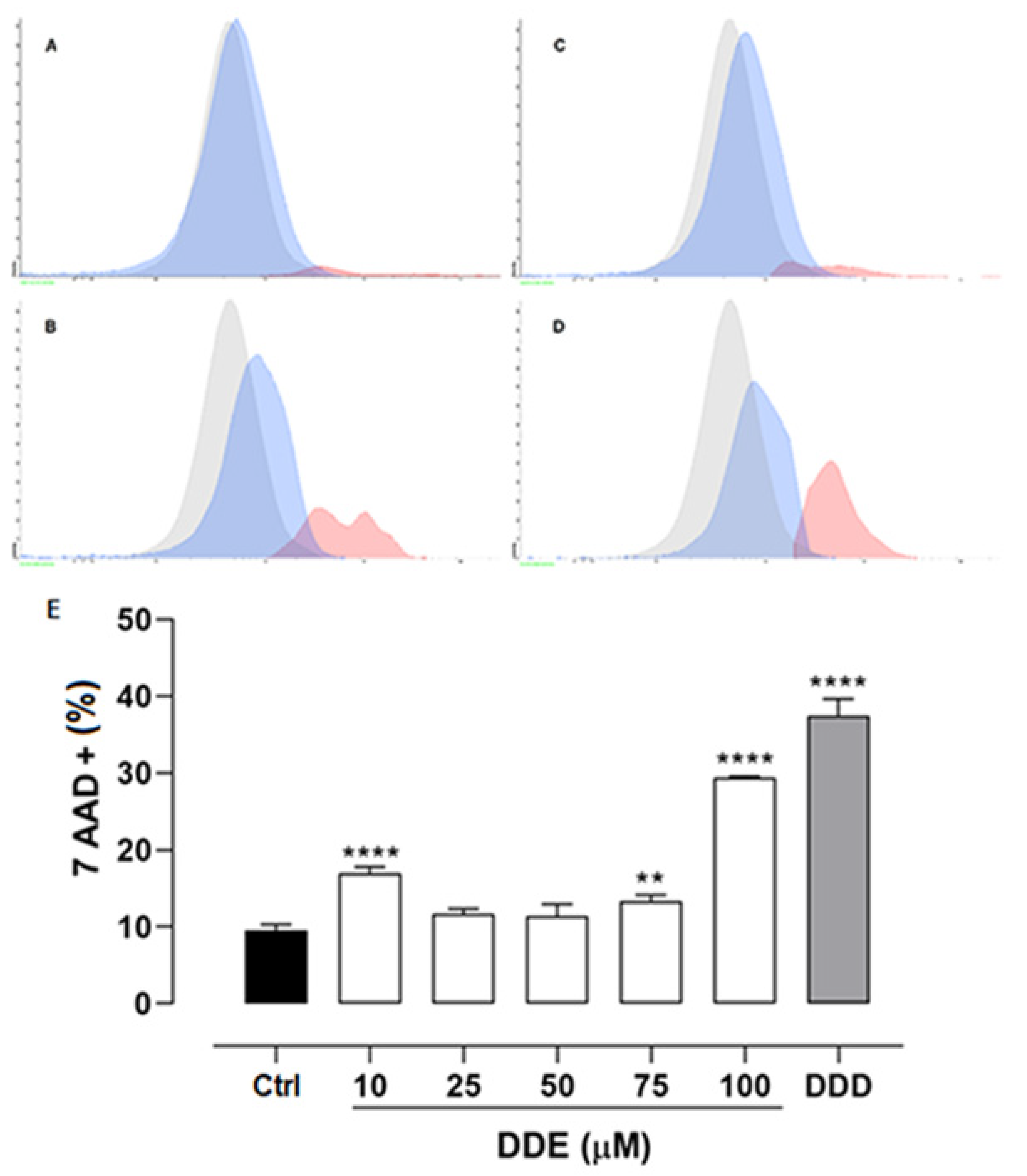
2.4. Effect of o,p’-DDD and o,p’-DDE on Necrosis in H295R Cell Line
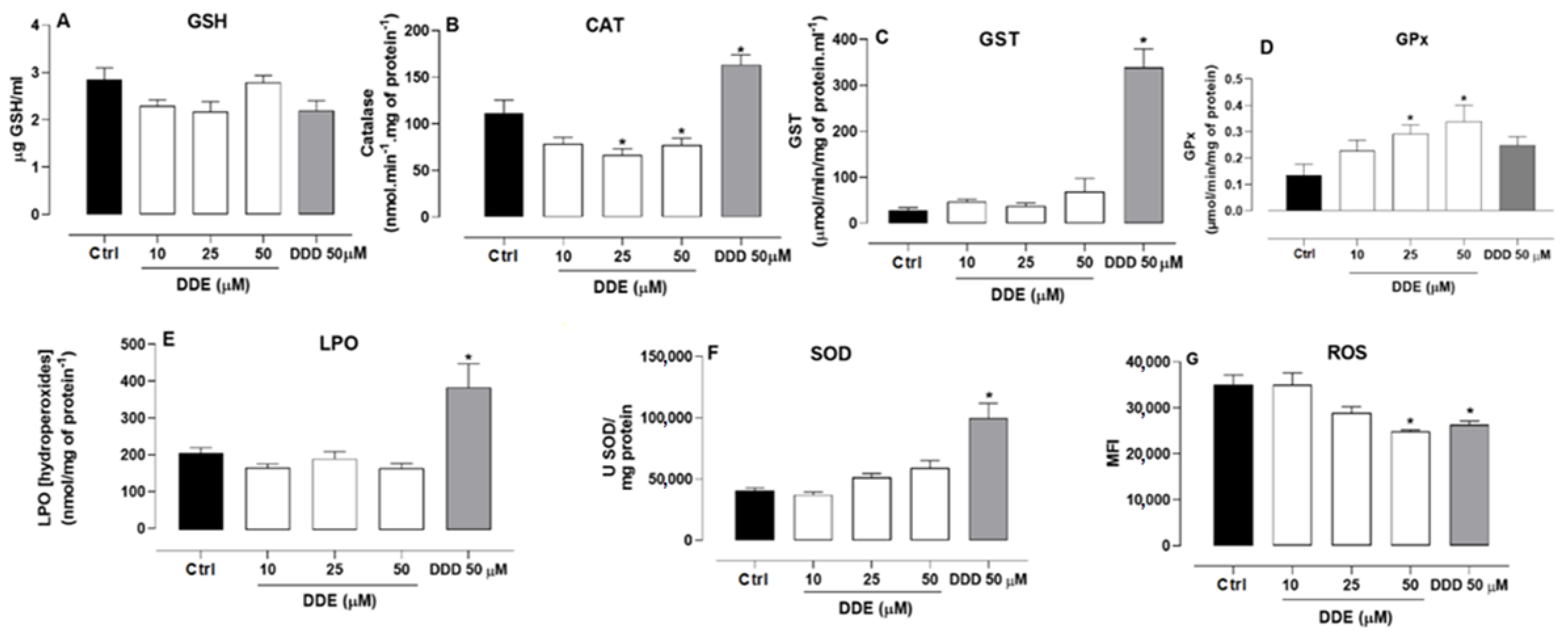
2.5. Effect of o,p’-DDD and o,p’-DDE on Oxidative Parameters in H295R Cell Line
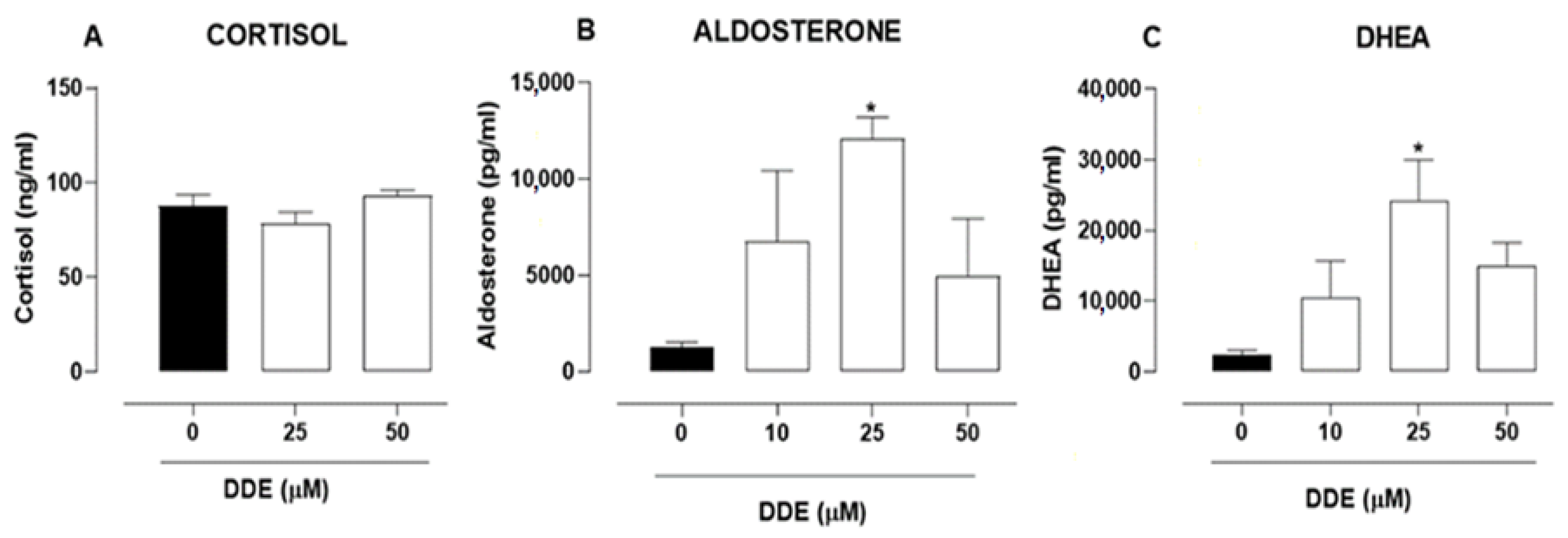
2.6. Effects of o,p’-DDD and o,p’-DDE in Hormone Secretion in H295R Cell Line
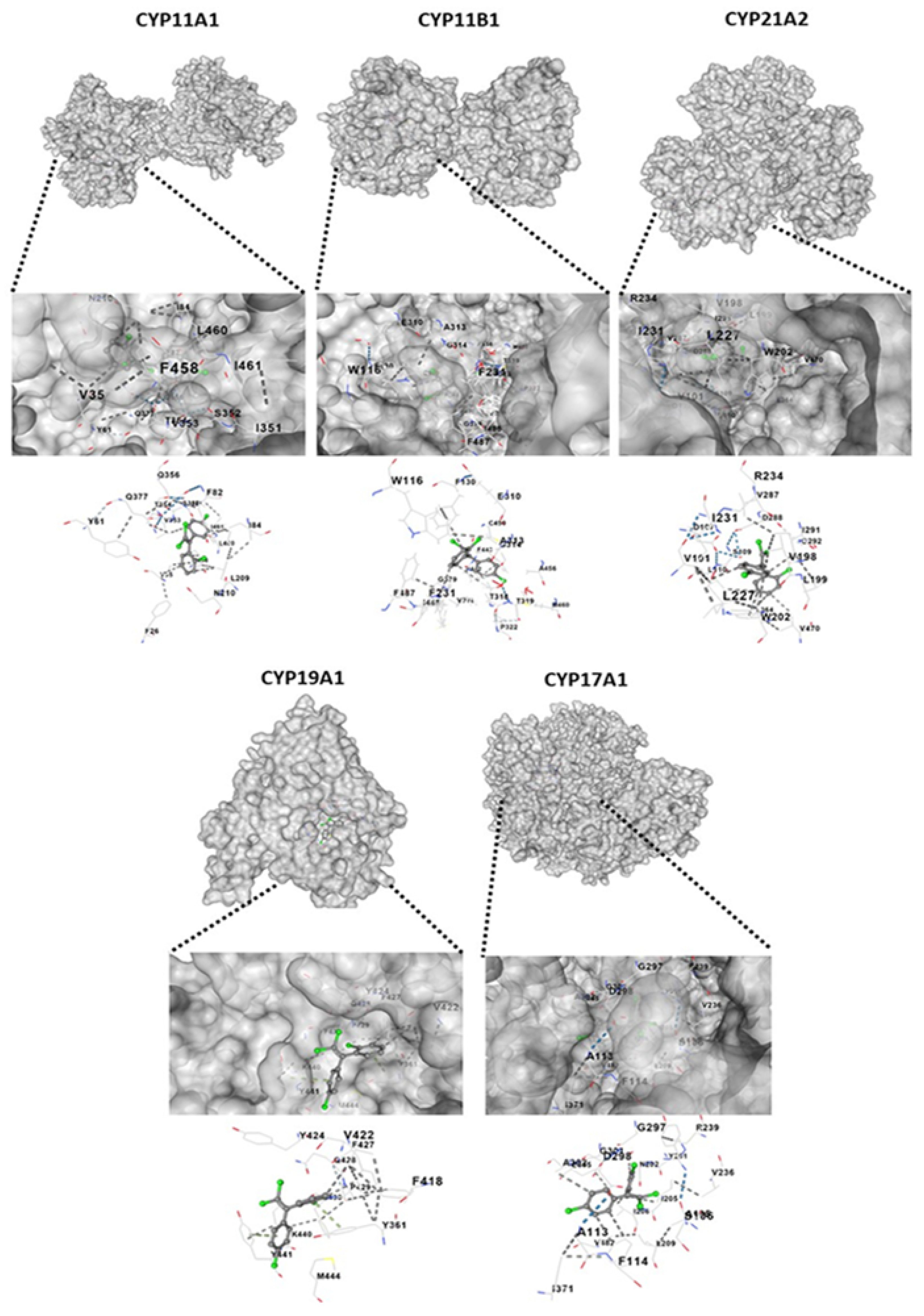
2.7. Molecular Docking of o,p’-DDE with CYP11A1, CYP11B1, CYP21A2, CYP19A1 and CYP17A1
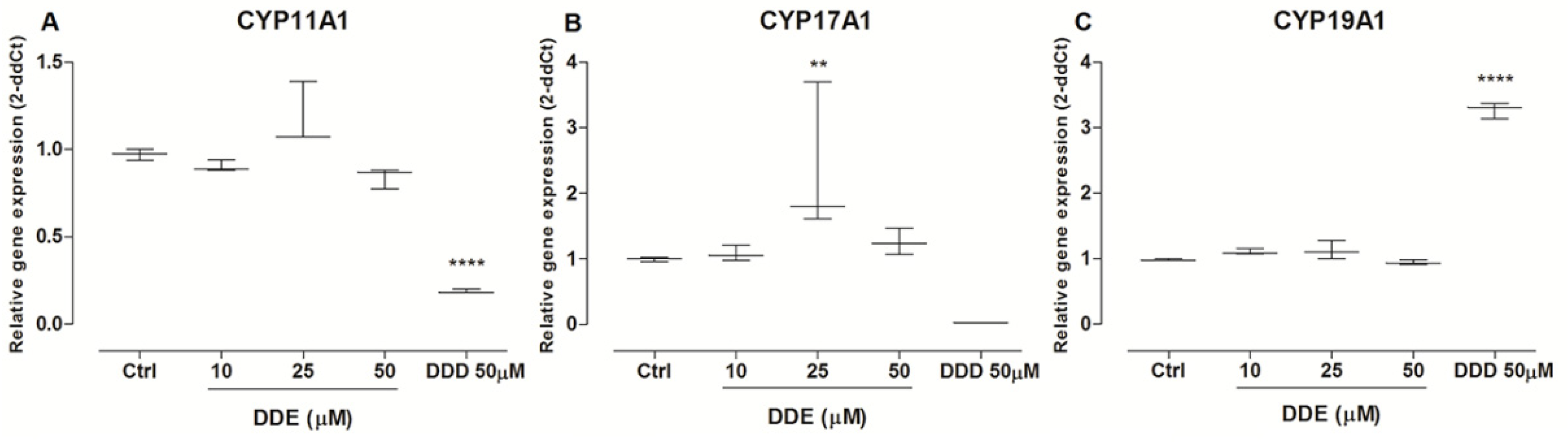
2.8. Effects of o,p’-DDD and o,p’-DDE in CYP’s Expression
3. Discussion
4. Materials and Methods
4.1. Cell Culture
4.2. Cell Viability
4.3. Caspase 3/7 Assessment
4.4. Necroptosis Assessment
4.5. Necrosis Assessment
4.6. Oxidative Parameters
4.6.1. Reduced Glutathione (GSH)
4.6.2. Catalase (CAT)
4.6.3. Glutathione S-Transferase (GST)
4.6.4. Lipoperoxidation Assay (LPO)
4.6.5. Superoxide Dismutase (SOD)
4.6.6. Reactive Oxygen Species (ROS)
4.6.7. Glutathione Peroxidases (GPx)
4.6.8. Protein Quantification
4.7. Dosage of DHEA, Cortisol and Aldosterone
4.8. Molecular Docking
4.9. CYP’s Expression
4.10. Statistical Analysis
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Custódio, G.A.; Parise, G.A.; Filho, N.K.; Komechen, H.; Sabbaga, C.C.; Rosati, R.; Grisa, L.; Parise, I.Z.; Pianovski, M.A.; Fiori, C.M.; et al. Impact of neonatal screening and surveillance for the TP53 R337H mutation on early detection of childhood adrenocortical tumors. J. Clin. Oncol. 2013, 31, 2619–2626. [Google Scholar] [CrossRef]
- Maluf, D.F.; De Oliveira, B.H.; Lalli, E. Therapy of adrenocortical cancer: Present and future. Am. J. Cancer Res. 2011, 1, 222–232. [Google Scholar] [PubMed]
- Nichols, J.; Green, H.D. Effect of DDD treatment on metabolic response of dogs to ACTH injection. Am. J. Physiol. 1954, 176, 374–376. [Google Scholar] [CrossRef] [PubMed]
- Bergenstal, D.M.; Lipsett, M.; Moy, R.; Hertz, R. Regression of Adrenal Cancer and Suppression of Adrenal Function in Man by o,p′-DDD. Biol. Act. Steroids Relat. Cancer 1959, 72, 463–475. [Google Scholar] [CrossRef]
- Waszut, U.; Szyszka, P.; Dworakowska, D. Understanding Mitotane Mode Action. J. Physiol. Pharmacol. 2017, 8, 13–26. [Google Scholar] [CrossRef]
- Mauclère-Denost, S.; Leboulleux, S.; Borget, I.; Paci, A.; Young, J.; Al Ghuzlan, A.; Deandreis, D.; Drouard, L.; Tabarin, A.; Chanson, P.; et al. High-dose mitotane strategy in adrenocortical carcinoma: Prospective analysis of plasma mitotane measurement during the first 3 months of follow-up. Eur. J. Endocrinol. 2012, 166, 261–268. [Google Scholar] [CrossRef][Green Version]
- Zancanella, P.; Pianovski, M.A.D.; Oliveira, B.H.; Ferman, S.; Piovezan, G.C.; Lichtvan, L.L.; Voss, S.Z.; Stinghen, S.T.; Callefe, L.G.; Parise, G.A.; et al. Mitotane Associated with Cisplatin, Etoposide, and Doxorubicin in Advanced Childhood Adrenocortical Carcinoma. J. Pediatr. Hematol. Oncl. 2006, 28, 513–524. [Google Scholar] [CrossRef]
- Faggiano, A.; Leboulleux, S.; Young, J.; Schlumberger, M.; Baudin, E. Rapidly progressing high o,p’DDD doses shorten the time required to reach the therapeutic threshold with an acceptable tolerance: Preliminary results. Clin. Endocrinol. 2006, 64, 110–113. [Google Scholar] [CrossRef] [PubMed]
- Terzolo, M. Low-dose monitored mitotane treatment achieves the therapeutic range with manageable side effects in patients with adrenocortical cancer. J. Clin. Endocrinol. Metab. 2000, 85, 2234–2238. [Google Scholar] [CrossRef] [PubMed]
- Tang, Y.; Liu, Z.; Zou, Z.; Liang, J.; Lu, Y.; Zhu, Y. Benefits of Adjuvant Mitotane after Resection of Adrenocortical Carcinoma: A Systematic Review and Meta-Analysis. BioMed Res. Int. 2018, 2018, 9362108. [Google Scholar] [CrossRef]
- Daffara, F.; De Francia, S.; Reimondo, G.; Zaggia, B.; Aroasio, E.; Porpiglia, F.; Volante, M.; Termine, A.; Di Carlo, F.; Dogliotti, L.; et al. Prospective evaluation of mitotane toxicity in adrenocortical cancer patients treated adjuvantly. Endocr. Relat. Cancer 2008, 15, 1043–1053. [Google Scholar] [CrossRef] [PubMed]
- Corso, C.R.; Acco, A.; Bach, C.; Bonatto, S.J.R.; de Figueiredo, B.C.; de Souza, L.M. Pharmacological profile and effects of mitotane in adrenocortical carcinoma. Br. J. Clin. Pharmacol. 2021, 87, 2698–2710. [Google Scholar] [CrossRef]
- Martz, F.; Straw, J.A. Metabolism and covalent binding of 1-(o-chlorophenyl)-1-(p-chlorophenyl)-2,2-dichloroethane (o,p’-DDD). Correlation between adrenocorticolytic activity and metabolic activation by adrenocortical mitochondria. Drug Metab. Dispos. 1980, 8, 127–130. [Google Scholar]
- Hermsen, I.G.; Fassnacht, M.; Terzolo, M.; Houterman, S.; den Hartigh, J.; Leboulleux, S.; Daffara, F.; Berruti, A.; Chadarevian, R.; Schlumberger, M.; et al. Plasma concentrations of o,p′DDD, o,p′DDA, and o,p′DDE as predictors of tumor response to mitotane in adrenocortical carcinoma: Results of a retrospective ENS@T multicenter study. J. Clin. Endocrinol. Metab. 2011, 96, 1844–1851. [Google Scholar] [CrossRef][Green Version]
- Germano, A. CYP11B1 has no role in mitotane action and metabolism in adrenocortical carcinoma cells. PLoS ONE. 2018, 13, e0196931. [Google Scholar] [CrossRef]
- Hescot, S. The Lack of Antitumor Effects of o,p′DDA Excludes Its Role as an Active Metabolite of Mitotane for Adrenocortical Carcinoma Treatment. Horm. Cancer 2014, 5, 312–323. [Google Scholar] [CrossRef] [PubMed]
- Kroiss, M.; Plonné, D.; Kendl, S.; Schirmer, D.; Ronchi, C.L.; Schirbel, A.; Zink, M.; Lapa, C.; Klinker, H.; Fassnacht, M.; et al. Association of mitotane with chylomicrons and serum lipoproteins: Practical implications for treatment of adrenocortical carcinoma. Eur. J. Endocrinol. 2016, 174, 343–353. [Google Scholar] [CrossRef] [PubMed]
- Garg, M.B.; Sakoff, J.A.; Ackland, S.P. A simple HPLC method for plasma level monitoring of mitotane and its two main metabolites in adrenocortical cancer patients. J. Chromatogr. B Anal. Technol. Biomed. Life Sci. 2011, 879, 2201–2205. [Google Scholar] [CrossRef]
- Germano, A.; Rapa, I.; Volante, M.; De Francia, S.; Migliore, C.; Berruti, A.; Papotti, M.; Terzolo, M. RRM1 modulates mitotane activity in adrenal cancer cells interfering with its metabolization. Mol. Cell. Endocrinol. 2015, 401, 105–110. [Google Scholar] [CrossRef]
- Liu, Y.; Grimm, M.; Dai, W.-T.; Hou, M.-C.; Xiao, Z.-X.; Cao, Y. CB-Dock: A web server for cavity detection-guided protein-ligand blind docking. Acta Pharmacol. Sin. 2020, 41, 138–144. [Google Scholar] [CrossRef]
- Tao, Y.G.; Huang, X.-F.; Wang, J.-Y.; Kang, M.-R.; Wang, L.-J.; Xian, S.-X. Exploring Molecular Mechanism of Huangqi in Treating Heart Failure Using Network Pharmacology, Evidence-Based Complement. Altern. Med. 2020, 2020, 6473745. [Google Scholar] [CrossRef]
- Lindhe, Ö.; Skogseid, B. Mitotane effects in a H295R xenograft model of adjuvant treatment of adrenocortical cancer. Horm. Metab. Res. 2010, 42, 725–730. [Google Scholar] [CrossRef]
- Hescot, S.; Amazit, L.; Lhomme, M.; Travers, S.; Dubow, A.; Battini, S.; Boulate, G.; Namer, I.J.; Lombes, A.; Kontush, A.; et al. Identifying Mitotane-induced Mitochondria-associated membranes dysfunctions: Metabolomic and Lipidomic approaches. Oncotarget 2017, 8, 109924–109940. [Google Scholar] [CrossRef] [PubMed]
- Lehmann, T.P.; Wrzesiński, T.; Jagodziński, P.P. The effect of mitotane on viability, steroidogenesis and gene expression in NCI-H295R adrenocortical cells. Mol. Med. Rep. 2013, 7, 893–900. [Google Scholar] [CrossRef]
- Poli, G.; Guasti, D.; Rapizzi, E.; Fucci, R.; Canu, L.; Bandinelli, A.; Cini, N.; Bani, D.; Mannelli, M.; Luconi, M.; et al. Morphofunctional effects of mitotane on mitochondria in human adrenocortical cancer cells. Endocr. Relat. Cancer 2013, 20, 537–550. [Google Scholar] [CrossRef] [PubMed]
- Sbiera, S.; Leich, E.; Liebisch, G.; Sbiera, I.; Schirbel, A.; Wiemer, L.; Matysik, S.; Eckhardt, C.; Gardill, F.; Gehl, A.; et al. Mitotane inhibits sterol-o-Acyl transferase 1 triggering lipid-mediated endoplasmic reticulum stress and apoptosis in adrenocortical carcinoma cells. Endocrinology 2015, 156, 3895–3908. [Google Scholar] [CrossRef] [PubMed]
- D’Arcy, M.S. Cell death: A review of the major forms of apoptosis, necrosis and autophagy. Cell Biol. Int. 2019, 43, 582–592. [Google Scholar] [CrossRef]
- Alegre-Cortés, E.; Muriel-González, A.; Canales-Cortés, S.; Uribe-Carretero, E.; Martínez-Chacón, G.; Aiastui, A.; López De Munain, A.; Niso-Santano, M.; Gonzalez-Polo, R.A.; Fuentes, J.M.; et al. Toxicity of Necrostatin-1 in Parkinson’s Disease Models. Antioxidants 2020, 9, 524. [Google Scholar] [CrossRef]
- Duan, X.; Liu, X.; Liu, N.; Huang, Y.; Jin, Z.; Zhang, S.; Ming, Z.; Chen, H. Inhibition of keratinocyte necroptosis mediated by RIPK1/RIPK3/MLKL provides a protective effect against psoriatic inflammation. Cell Death Dis. 2020, 11, 134. [Google Scholar] [CrossRef]
- Weigand, I. Active steroid hormone synthesis renders adrenocortical cells highly susceptible to type II ferroptosis induction. Cell Death Dis. 2020, 11, 192. [Google Scholar] [CrossRef]
- Yang, W.S.; Ramaratnam, R.; Welsch, M.E.; Shimada, K.; Skouta, R.; Viswanathan, V.S.; Cheah, J.H.; Clemons, P.A.; Shamji, A.F.; Clish, C.B.; et al. Regulation of Ferroptotic Cancer Cell Death by GPX4. Cell 2014, 156, 317–331. [Google Scholar] [CrossRef]
- Belavgeni, A.; Bornstein, S.R.; von Mässenhausen, A.; Tonnus, W.; Stumpf, J.; Meyer, C.; Othmar, E.; Latk, M.; Kanczkowski, W.; Kroiss, M.; et al. Exquisite sensitivity of adrenocortical carcinomas to induction of ferroptosis. Proc. Natl. Acad. Sci. USA 2019, 116, 22269–22274. [Google Scholar] [CrossRef]
- Stockwell, B.R.; Jiang, X. The Chemistry and Biology of Ferroptosis. Cell Chem. Biol. 2020, 176, 100–106. [Google Scholar] [CrossRef]
- Slimen, I.B.; Najar, T.; Ghram, A.; Dabbebi, H.; Ben Mrad, M.; Abdrabbah, M. Reactive oxygen species, heat stress and oxidative-induced mitochondrial damage. A review. Int. J. Hyperth. 2014, 30, 513–523. [Google Scholar] [CrossRef] [PubMed]
- Curtin, J.F.; Donovan, M.; Cotter, T.G. Regulation and measurement of oxidative stress in apoptosis. J. Immunol. Methods 2002, 265, 49–72. [Google Scholar] [CrossRef] [PubMed]
- Sinha, K.; Das, J.; Pal, P.B.; Sil, P.C. Oxidative stress: The mitochondria-dependent and mitochondria-independent pathways of apoptosis. Arch. Toxicol. 2013, 87, 1157–1180. [Google Scholar] [CrossRef]
- Rossini, E.; Giacopuzzi, E.; Gangemi, F.; Tamburello, M.; Cosentini, D.; Abate, A.; Laganà, M.; Berruti, A.; Grisanti, S.; Sigala, S. Estrogen-like effect of mitotane explained by its agonist activity on estrogen receptor-α. Biomedicines 2021, 9, 681. [Google Scholar] [CrossRef]
- Zsippai, A.; Szabó, D.R.; Tömböl, Z.; Szabó, P.M.; Éder, K.; Pállinger, E.; Gaillard, R.C.; Patócs, A.; Tóth, S.; Falus, A.; et al. Effects of mitotane on gene expression in the adrenocortical cell line NCI-H295R: A microarray study. Pharmacogenomics 2012, 13, 1351–1361. [Google Scholar] [CrossRef] [PubMed]
- Asp, V. In Vitro Studies of Adrenocorticolytic DDT Metabolites, with Special Focus on 3-Methylsulfonyl-DDE. Ph.D. Thesis, Uppsala University, Uppsala, Swedish, 2010. [Google Scholar]
- Asp, V.; Ullerås, E.; Lindström, V.; Bergström, U.; Oskarsson, A.; Brandt, I. Biphasic hormonal responses to the adrenocorticolytic DDT metabolite 3-methylsulfonyl-DDE in human cells. Toxicol. Appl. Pharmacol. 2010, 242, 281–289. [Google Scholar] [CrossRef] [PubMed]
- Wang, T.; Rainey, W.E. Human Adrenocortical Carcinoma Cell Lines. Bone 2012, 351, 58–65. [Google Scholar] [CrossRef]
- Mosmann, T. Rapid Colorimetric Assay for Cellular Growth and Survival: Application to Proliferation and Cytotoxicity Assays. J. Oflmmunol. Methods 1983, 65, 55–63. [Google Scholar] [CrossRef] [PubMed]
- Rajamohan, S.B.; Raghuraman, G.; Prabhakar, N.R.; Kumar, G.K. NADPH oxidase-derived H2O2 contributes to angiotensin II-induced aldosterone synthesis in human and rat adrenal cortical cells. Antioxid. Redox Signal. 2012, 17, 445–459. [Google Scholar] [CrossRef] [PubMed]
- Hescot, S.; Slama, A.; Lombès, A.; Paci, A.; Remy, H.; Leboulleux, S.; Chadarevian, R.; Trabado, S.; Amazit, L.; Young, J.; et al. Mitotane alter mitochondrial respiratory chain activity by inducing cytochrome c oxidase defect in human adrenocortical cells. Endocr. Relat. Cancer 2013, 20, 371–381. [Google Scholar] [CrossRef] [PubMed]
- Sedlak, J.; Lindsay, R.H. Estimation of Total, Protein-Bound, and Nonprotein Sulfhydryl Groups in Tissue with Ellman’s Reagent. Anal. Biochem. 1968, 25, 192–205. [Google Scholar] [CrossRef] [PubMed]
- Aebi, H. Catalase in vitro. In Methods in Enzymology; Academic Press: Cambridge, MA, USA, 1984; Volume 105, pp. 121–126. [Google Scholar]
- Habig, W.H.; Pabst, M.J.; Jakoby, W.B. Glutathione S transferases. The first enzymatic step in mercapturic acid formation. J. Biol. Chem. 1974, 249, 7130–7139. [Google Scholar] [CrossRef]
- Jiang, Z.; Woollard, A.C.S.; Wolff, S.P. Lipid Hydroperoxide Measurement by Oxidation of Fe2+ in the Presence of Xylenol Orange. Comparison with the TBA Assay and an Iodometric Method. Lipids 1991, 26, 853–856. [Google Scholar] [CrossRef]
- Gao, R.; Yuan, Z.; Zhao, Z.; Gao, X. Mechanism of pyrogallol autoxidation and determination of superoxide dismutase enzyme activity. Bioelectrochemistry Bioenerg. 1998, 45, 41–45. [Google Scholar] [CrossRef]
- Eruslanov, E.; Kusmartsev, S. Identification of ROS Using Oxidized DCFDA and Flow-Cytometry. In Advanced Protocols in Oxidative Stress II; Humana Press: Totowa, NJ, USA, 2009; Volume 594, pp. 1–477. [Google Scholar] [CrossRef]
- Chance, B.; Sies, H.; Boveris, A. Hydroperoxide metabolism in mammalian organs. Physiol. Rev. 1979, 59, 527–605. [Google Scholar] [CrossRef]
- Bradford, M.M. A rapid and sensitive method for the quantitation of microgram quantities of protein utilizing the principle of protein-dye binding. Anal. Biochem. 1976, 72, 248–254. [Google Scholar] [CrossRef]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Receptor | PDB ID | Vina Score | Cavity Size | Center | Size | ||||
|---|---|---|---|---|---|---|---|---|---|
| x | y | z | x | y | z | ||||
| CYP11A1 | 3N9Y | −7.9 | 3903 | 14 | −5 | 17 | 19 | 31 | 19 |
| CYP11B1 | 6M7X | −7.5 | 2562 | 51 | −47 | −8 | 25 | 27 | 34 |
| CYP21A2 | 4Y8W | −8 | 2231 | −13 | 12 | 28 | 25 | 19 | 19 |
| CYP19A1 | 5JL6 | −7 | 2120 | 84 | 51 | 49 | 19 | 19 | 30 |
| CYP17A1 | 3RUK | −7.5 | 19,788 | 26 | 16 | 43 | 35 | 35 | 35 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Bach, C.; Corso, C.R.; Veiga, A.d.A.; Paraizo, M.M.; de Souza, L.M. Effects of o,p’-DDE, a Mitotane Metabolite, in an Adrenocortical Carcinoma Cell Line. Pharmaceuticals 2022, 15, 1486. https://doi.org/10.3390/ph15121486
Bach C, Corso CR, Veiga AdA, Paraizo MM, de Souza LM. Effects of o,p’-DDE, a Mitotane Metabolite, in an Adrenocortical Carcinoma Cell Line. Pharmaceuticals. 2022; 15(12):1486. https://doi.org/10.3390/ph15121486
Chicago/Turabian StyleBach, Camila, Claudia Rita Corso, Alan de Almeida Veiga, Mariana Martins Paraizo, and Lauro Mera de Souza. 2022. "Effects of o,p’-DDE, a Mitotane Metabolite, in an Adrenocortical Carcinoma Cell Line" Pharmaceuticals 15, no. 12: 1486. https://doi.org/10.3390/ph15121486
APA StyleBach, C., Corso, C. R., Veiga, A. d. A., Paraizo, M. M., & de Souza, L. M. (2022). Effects of o,p’-DDE, a Mitotane Metabolite, in an Adrenocortical Carcinoma Cell Line. Pharmaceuticals, 15(12), 1486. https://doi.org/10.3390/ph15121486