

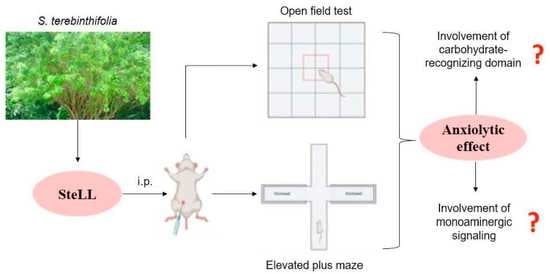
The Anxiolytic Activity of Schinus terebinthifolia Leaf Lectin (SteLL) Is Dependent on Monoaminergic Signaling although Independent of the Carbohydrate-Binding Domain of the Lectin

 ,
,  , and
, and
Abstract
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
1. Introduction
2. Results
2.1. SteLL Revealed an Anxiolytic Effect on the Open Field (OFT) and on the Elevated Plus Maze Test (EPM)
2.2. The Anxiolytic Effect of SteLL Did Not Depend on the Carbohydrate-Recognition Domain (CRD) of the Lectin
2.3. The Anxiolytic Effect of SteLL in the EPM Is Dependent on Monoaminergic Pathways
3. Discussion
4. Materials and Methods
4.1. Animals
4.2. Purification of Lectin
4.3. Drugs and Treatment
4.4. The Effect of the Carbohydrate-Recognizing Domain (CRD) on the Anxiolytic Effect of the Lectin
4.5. Open Field (OF) Test
4.6. Elevated Plus Maze (EPM) Test
4.7. Statistical Analysis
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Anxiety and Depression Association of America (ADAA). Available online: https://adaa.org/ (accessed on 15 August 2022).
- Fedotova, J.; Kubatka, P.; Büsselberg, D.; Shleikin, A.G.; Caprnda, M.; Dragasek, J.; Rodrigo, L.; Pohanka, M.; Gasparova, I.; Nosáľ, V.; et al. Therapeutical strategies for anxiety and anxiety-like disorders using plant-derived natural compounds and plant extracts. Biomed. Pharmacother. 2017, 95, 437–446. [Google Scholar] [CrossRef]
- Aslanargun, P.; Cuvas, O.; Dikmen, B.; Aslan, E.; Yuksel, M.U. Passiflora incarnata Linneaus as an anxiolytic before spinal anesthesia. J. Anesthesia 2012, 26, 39–44. [Google Scholar] [CrossRef]
- Fuladi, S.; Emami, S.A.; Mohammadpour, A.H.; Karimani, A.; Manteghi, A.A.; Sahebkar, A. Assessment of the Efficacy of Withania somnifera Root Extract in Patients with Generalized Anxiety Disorder: A Randomized Double-blind Placebo- Controlled Trial. Curr. Rev. Clin. Exp. Pharmacol. 2021, 16, 191–196. [Google Scholar] [CrossRef]
- Patriota, L.L.S.; Brito, J.S.; Ramos, D.B.M.; Procópio, T.F.; Paiva, P.M.G.; Pontual, E.V.; Melo, C.M.L.; Napoleão, T.H. Plant-derived lectins: A review of their status as alternatives to anticancer chemotherapy. In Horizons in Cancer Research; Watanabe, H.S., Ed.; Nova Science Publishers Inc.: New York, NY, USA, 2019; pp. 171–205. [Google Scholar]
- Lannoo, N.; Van Damme, E.J.M. Lectin domains at the frontiers of plant defense. Front. Plant Sci. 2014, 5, 397. [Google Scholar] [CrossRef]
- Breitenbach Barroso Coelho, L.C.; Marcelino Dos Santos Silva, P.; Felix de Oliveira, W.; de Moura, M.C.; Viana Pontual, E.; Soares Gomes, F.; Guedes Paiva, P.M.; Napoleao, T.H.; Dos Santos Correia, M.T. Lectins as antimicrobial agents. J. Appl. Microbiol. 2018, 125, 1238–1252. [Google Scholar] [CrossRef]
- Russi, M.A.; Vandresen-Filho, S.; Rieger, D.K.; Costa, A.P.; Lopes, M.W.; Cunha, R.M.S.; Teixeira, E.H.; Nascimento, K.S.; Cavada, B.; Tasca, C.I.; et al. ConBr, a Lectin from Canavalia brasiliensis Seeds, Protects Against Quinolinic Acid-Induced Seizures in Mice. Neurochem. Res. 2012, 37, 288–297. [Google Scholar] [CrossRef]
- Jacques, A.V.; Rieger, D.K.; Maestri, M.; Lopes, M.W.; Peres, T.V.; Gonçalves, F.M.; Pedro, D.Z.; Tasca, C.I.; López, M.G.; Egea, J.; et al. Lectin from Canavalia brasiliensis (ConBr) protects hippocampal slices against glutamate neurotoxicity in a manner dependent of PI3K/Akt pathway. Neurochem. Int. 2013, 62, 836–842. [Google Scholar] [CrossRef]
- Santos, A.J.C.A.; Barros, B.R.D.S.; Aguiar, L.M.D.S.; Patriota, L.L.D.S.; Lima, T.D.A.; Zingali, R.B.; Paiva, P.M.G.; Napoleão, T.H.; de Melo, C.M.L.; Pontual, E.V. Schinus terebinthifolia leaf lectin (SteLL) is an immunomodulatory agent by altering cytokine release by mice splenocytes. 3 Biotech 2020, 10, 144. [Google Scholar] [CrossRef]
- Silva, A.R.; de Oliveira, W.F.; da Silva, P.M.; Patriota, L.L.D.S.; Alves, R.R.D.V.; de Oliveira, A.P.S.; Correia, M.T.D.S.; Paiva, P.M.G.; Vainstein, M.H.; Filho, P.E.C.; et al. Quantum dots conjugated to lectins from Schinus terebinthifolia leaves (SteLL) and Punica granatum sarcotesta (PgTeL) as potential fluorescent nanotools for investigating Cryptococcus neoformans. Int. J. Biol. Macromol. 2021, 192, 232–240. [Google Scholar] [CrossRef]
- Gomes, F.S.; Procópio, T.F.; Napoleão, T.H.; Coelho, L.C.B.B.; Paiva, P.M.G. Antimicrobial lectin from Schinus terebinthifolius leaf. J. Appl. Microbiol. 2013, 114, 672–679. [Google Scholar] [CrossRef]
- Lima, I.M.D.S.F.; Zagmignan, A.; Santos, D.M.; Maia, H.S.; Silva, L.D.S.; Cutrim, B.D.S.; Vieira, S.L.; Filho, C.M.B.; de Sousa, E.M.; Napoleão, T.H.; et al. Schinus terebinthifolia leaf lectin (SteLL) has anti-infective action and modulates the response of Staphylococcus aureus-infected macrophages. Sci. Rep. 2019, 9, 18159. [Google Scholar] [CrossRef]
- Ramos, D.B.M.; Araújo, M.T.M.F.; Araújo, T.C.L.; Neto, O.G.S.; Silva, M.G.; Silva, Y.A.; Torres, D.J.L.; Patriota, L.L.S.; de Melo, C.M.L.; de Lorena, V.M.B.; et al. Evaluation of antitumor activity and toxicity of Schinus terebinthifolia leaf extract and lectin (SteLL) in sarcoma 180-bearing mice. J. Ethnopharmacol. 2019, 233, 148–157. [Google Scholar] [CrossRef]
- Santos, A.J.; Araújo, V.F.; França, R.P.; Silva, E.L.; Almeida, W.A.; Procópio, T.F.; Paiva, P.M.; Napoleão, T.H.; Costa, E.V.; Nogueira, R.A.; et al. Schinus terebinthifolia Raddi leaf lectin is an antiangiogenic agent for Coturnix japonica embryos. An. Acad. Bras. Ciências 2022, 94, e20211297. [Google Scholar] [CrossRef]
- Ramos, D.B.M.; Araújo, M.T.M.F.; Araújo, T.C.L.; Silva, Y.A.; Santos, A.C.L.A.; Silva, M.G.; Paiva, P.M.G.; Mendes, R.L.; Napoleão, T.H. Antinociceptive activity of Schinus terebinthifolia leaf lectin (SteLL) in sarcoma 180-bearing mice. J. Ethnopharmacol. 2020, 259, 112952. [Google Scholar] [CrossRef]
- Marinho, A.O.; Brito, J.S.; Costa, J.A.; Silva, A.R.; Silva, S.P.; Amorim, L.C.; Correia, M.T.S.; Paiva, P.M.G.; Oliveira, A.M.; Patriota, L.L.S.; et al. Schinus terebinthifolia leaf lectin has central and peripheral antinociceptive action mediated by its carbohydrate-recognition domain and delta-opioid receptors. J. Ethnopharmacol. 2022, 301, 115817. [Google Scholar] [CrossRef]
- Dalmagro, A.P.; Camargo, A.; Zeni, A.L.B. Morus nigra and its major phenolic, syringic acid, have antidepressant-like and neuroprotective effects in mice. Metab. Brain Dis. 2017, 32, 1963–1973. [Google Scholar] [CrossRef]
- Herbst, L.; Gaigher, T.; Siqueira, A.; Joca, S.; Sampaio, K.; Beijamini, V. New evidence for refinement of anesthetic choice in procedures preceding the forced swimming test and the elevated plus-maze. Behav. Brain Res. 2019, 368, 111897. [Google Scholar] [CrossRef]
- Planchez, B.; Surget, A.; Belzung, C. Animal models of major depression: Drawbacks and challenges. J. Neural Transm. 2019, 126, 1383–1408. [Google Scholar] [CrossRef]
- Liu, Y.; Zhao, J.; Guo, W. Emotional Roles of Mono-Aminergic Neurotransmitters in Major Depressive Disorder and Anxiety Disorders. Front. Psychol. 2018, 9, 2201. [Google Scholar] [CrossRef]
- Bandelow, B.; Baldwin, D.; Abelli, M.; Bolea-Alamanac, B.; Bourin, M.; Chamberlain, S.R.; Cinosi, E.; Davies, S.; Domschke, K.; Fineberg, N.; et al. Biological markers for anxiety disorders, OCD and PTSD: A consensus statement. Part II: Neurochemistry, neurophysiology and neurocognition. World J. Biol. Psychiatry 2017, 18, 162–214. [Google Scholar] [CrossRef]
- Montoya, A.; Bruins, R.; Katzman, M.; Blier, P. The noradrenergic paradox: Implications in the management of depression and anxiety. Neuropsychiatr. Dis. Treat. 2016, 12, 541–557. [Google Scholar] [CrossRef]
- Duan, C.; Chen, Y.; Shen, A.; Xu, J.; Zhao, Y.; Cai, R.; Liu, Y.; Zhou, L.; Lei, Y.; Hamre, K.; et al. Genetic expression analysis of E2F-associated phosphoprotein in stress responses in the mouse. Gene 2016, 581, 130–138. [Google Scholar] [CrossRef]
- Nestler, E.J.; Barrot, M.; DiLeone, R.J.; Eisch, A.J.; Gold, S.J.; Monteggia, L.M. Neurobiology of depression. Neuron 2002, 34, 13–25. [Google Scholar] [CrossRef]
- Wang, Q.; Timberlake, M.A.; Prall, K.; Dwivedi, Y. The recent progress in animal models of depression. Prog. Neuro-Psychopharmacology Biol. Psychiatry 2017, 77, 99–109. [Google Scholar] [CrossRef]
- Snyder, C.N.; Brown, A.R.; Buffalari, D. Similar tests of anxiety-like behavior yield different results: Comparison of the open field and free exploratory rodent procedures. Physiol. Behav. 2021, 230, 113246. [Google Scholar] [CrossRef]
- Bourin, M. Animal models for screening anxiolytic-like drugs: A perspective. Dialogues Clin. Neurosci. 2015, 17, 295–303. [Google Scholar] [CrossRef]
- Bourin, M.; Petit-Demoulière, B.; Nic Dhonnchadha, B.; Hascöet, M. Animal models of anxiety in mice. Fundam. Clin. Pharmacol. 2007, 21, 567–574. [Google Scholar] [CrossRef]
- Biedermann, S.V.; Biedermann, D.G.; Wenzlaff, F.; Kurjak, T.; Nouri, S.; Auer, M.K.; Wiedemann, K.; Briken, P.; Haaker, J.; Lonsdorf, T.B.; et al. An elevated plus-maze in mixed reality for studying human anxiety-related behavior. BMC Biol. 2017, 15, 125. [Google Scholar] [CrossRef]
- Araújo, J.R.C.; Campos, A.R.; Damasceno, M.B.M.V.; Santos, S.A.A.R.; Ferreira, M.K.A.; Moreira, R.A.M.; Monteiro-Moreira, A.C.O. Neuropharmacological Characterization of Dioclea altissima Seed Lectin (DAL) in Mice: Evidence of Anxiolytic-like Effect Mediated by Serotonergic, GABAergic Receptors and NO Pathway. Curr. Pharm. Des. 2020, 26, 3895–3904. [Google Scholar] [CrossRef]
- Ward, E.M.; Kizer, M.E.; Imperiali, B. Strategies and Tactics for the Development of Selective Glycan-Binding Proteins. ACS Chem. Biol. 2021, 16, 1795–1813. [Google Scholar] [CrossRef]
- Nascimento, K.S.; Silva, M.T.L.; Oliveira, M.V.; Lossio, C.F.; Pinto-Junior, V.R.; Osterne, V.J.S.; Cavada, B.S. Dalbergieae lectins: A review of lectins from species of a primitive Papilionoideae (leguminous) tribe. Int. J. Biol. Macromol. 2020, 144, 509–526. [Google Scholar] [CrossRef] [PubMed]
- Araújo, J.R.C.; Coelho, C.B.; Campos, A.R.; Moreira, R.D.A.; Monteiro-Moreira, A.C.D.O. Animal Galectins and Plant Lectins as Tools for Studies in Neurosciences. Curr. Neuropharmacol. 2020, 18, 202–215. [Google Scholar] [CrossRef] [PubMed]
- Rosa, M.M.; Ferreira, M.A.; Lima, C.A.O.; Mendonça, A.C.S.; Silva, Y.M.; Sharjeel, M.; Rego, M.J.B.M.; Pereira, M.C.; Pitta, M.G.R. Alzheimer’s disease: Is there a role for galectins? Eur. J. Pharmacol. 2021, 909, 174437. [Google Scholar] [CrossRef] [PubMed]
- Tao, C.C.; Cheng, K.M.; Ma, Y.L.; Hsu, W.L.; Chen, Y.C.; Fuh, J.L.; Lee, W.J.; Chao, C.C.; Lee, E.H. Galectin-3 promotes Aβ oligomerization and Aβ toxicity in a mouse model of Alzheimer’s disease. Cell Death Differ. 2020, 27, 192–209. [Google Scholar] [CrossRef]
- Yazar, T.; Yazar, H.O.; Cihan, M. Evaluation of serum galectin-3 levels at Alzheimer patients by stages: A preliminary report. Acta Neurol. Belg. 2021, 121, 949–954. [Google Scholar] [CrossRef]
- Chen, H.-L.; Liao, F.; Lin, T.-N.; Liu, F.-T. Galectins and Neuroinflammation. Adv. Neurobiol. 2014, 9, 517–542. [Google Scholar] [CrossRef]
- Bezerra, Y.B.S.; Oliveira, C.R.F.; Dornelles, L.P.; Guedes, C.C.S.; Oliveira, A.P.S.; Coelho, L.C.B.B.; Oliveira, C.H.C.M.; Santos, G.A.; Napoleão, T.H.; Lima, T.A.; et al. Pesticidal activity of Cratyllia mollis seed lectin preparation (cramoll 1,2,3) against the termite Nasutitermes corniger and mite Tetranychus bastosi. Crop Prot. 2022, 162, 106093. [Google Scholar] [CrossRef]
- National Research Council (US) Committee for the Update of the Guide for the Care and Use of Laboratory Animals. Guide for the Care and Use of Laboratory Animals, 8th ed.; National Academies Press (US): Washington, DC, USA, 2011. [Google Scholar]
- Lowry, O.H.; Rosebrough, N.J.; Farr, A.L.; Randall, R.J. Protein measurement with the Folin phenol reagent. J. Biol. Chem. 1951, 193, 265–275. [Google Scholar] [CrossRef]
- Procópio, T.F.; Patriota, L.L.S.; Moura, M.C.; Silva, P.M.; Oliveira, A.P.S.; Carvalho, L.V.N.; Lima, T.A.; Soares, T.; Silva, T.D.; Coelho, L.C.B.B.; et al. CasuL: A new lectin isolated from Calliandra surinamensis leaf pinnulae with cytotoxicity to cancer cells, antimicrobial activity and antibiofilm effect. Int. J. Biol. Macromol. 2017, 98, 419–429. [Google Scholar] [CrossRef]
- Araújo, J.R.C.; Júnior, J.M.A.M.; Damasceno, M.B.M.V.; Santos, S.A.A.R.; Vieira-Neto, A.E.; Lobo, M.D.P.; Campos, A.R.; Moreira, R.A.; Monteiro-Moreira, A.C.O. Neuropharmacological characterization of frutalin in mice: Evidence of an antidepressant-like effect mediated by the NMDA receptor/NO/cGMP pathway. Int. J. Biol. Macromol. 2018, 112, 548–554. [Google Scholar] [CrossRef]
- Omidi-Ardali, H.; Badi, A.G.; Saghaei, E.; Amini-Khoei, H. Nitric oxide mediates the antidepressant-like effect of modafinil in mouse forced swimming and tail suspension tests. J. Basic Clin. Physiol. Pharmacol. 2020, 32, 25–31. [Google Scholar] [CrossRef] [PubMed]
- Harsha, S.N.; Anilakumar, K.R. Anxiolytic property of hydro-alcohol extract of Lactuca sativa and its effect on behavioral activities of mice. J. Biomed. Res. 2013, 27, 37–42. [Google Scholar] [CrossRef] [PubMed]
- Moreno-Santos, B.; Coelho, C.M.; Ferreira, W.C.; Crestani, C.C. Angiotensinergic receptors in the medial amygdaloid nucleus differently modulate behavioral responses in the elevated plus-maze and forced swimming test in rats. Behav. Brain Res. 2021, 397, 112947. [Google Scholar] [CrossRef] [PubMed]






Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Lima, B.R.F.d.; Patriota, L.L.d.S.; Marinho, A.d.O.; Costa, J.A.d.; Napoleão, T.H.; Rosa, M.M.d.; Paiva, P.M.G. The Anxiolytic Activity of Schinus terebinthifolia Leaf Lectin (SteLL) Is Dependent on Monoaminergic Signaling although Independent of the Carbohydrate-Binding Domain of the Lectin. Pharmaceuticals 2022, 15, 1364. https://doi.org/10.3390/ph15111364
Lima BRFd, Patriota LLdS, Marinho AdO, Costa JAd, Napoleão TH, Rosa MMd, Paiva PMG. The Anxiolytic Activity of Schinus terebinthifolia Leaf Lectin (SteLL) Is Dependent on Monoaminergic Signaling although Independent of the Carbohydrate-Binding Domain of the Lectin. Pharmaceuticals. 2022; 15(11):1364. https://doi.org/10.3390/ph15111364
Chicago/Turabian StyleLima, Bárbara Raíssa Ferreira de, Leydianne Leite de Siqueira Patriota, Amanda de Oliveira Marinho, Jainaldo Alves da Costa, Thiago Henrique Napoleão, Michelle Melgarejo da Rosa, and Patrícia Maria Guedes Paiva. 2022. "The Anxiolytic Activity of Schinus terebinthifolia Leaf Lectin (SteLL) Is Dependent on Monoaminergic Signaling although Independent of the Carbohydrate-Binding Domain of the Lectin" Pharmaceuticals 15, no. 11: 1364. https://doi.org/10.3390/ph15111364
APA StyleLima, B. R. F. d., Patriota, L. L. d. S., Marinho, A. d. O., Costa, J. A. d., Napoleão, T. H., Rosa, M. M. d., & Paiva, P. M. G. (2022). The Anxiolytic Activity of Schinus terebinthifolia Leaf Lectin (SteLL) Is Dependent on Monoaminergic Signaling although Independent of the Carbohydrate-Binding Domain of the Lectin. Pharmaceuticals, 15(11), 1364. https://doi.org/10.3390/ph15111364