
Multi-Target Actions of Acridones from Atalantia monophylla towards Alzheimer’s Pathogenesis and Their Pharmacokinetic Properties

 , ,
, , 
Abstract
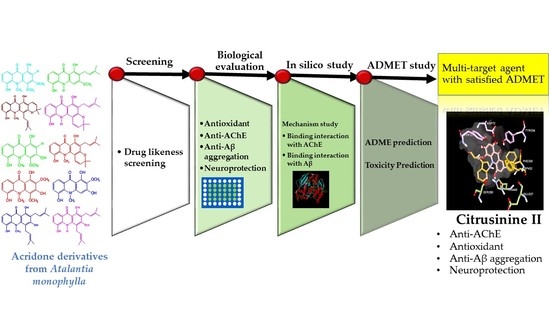
:
1. Introduction
2. Results and Discussion
2.1. Physicochemical Properties of Acridones
2.2. Antioxidant Activity
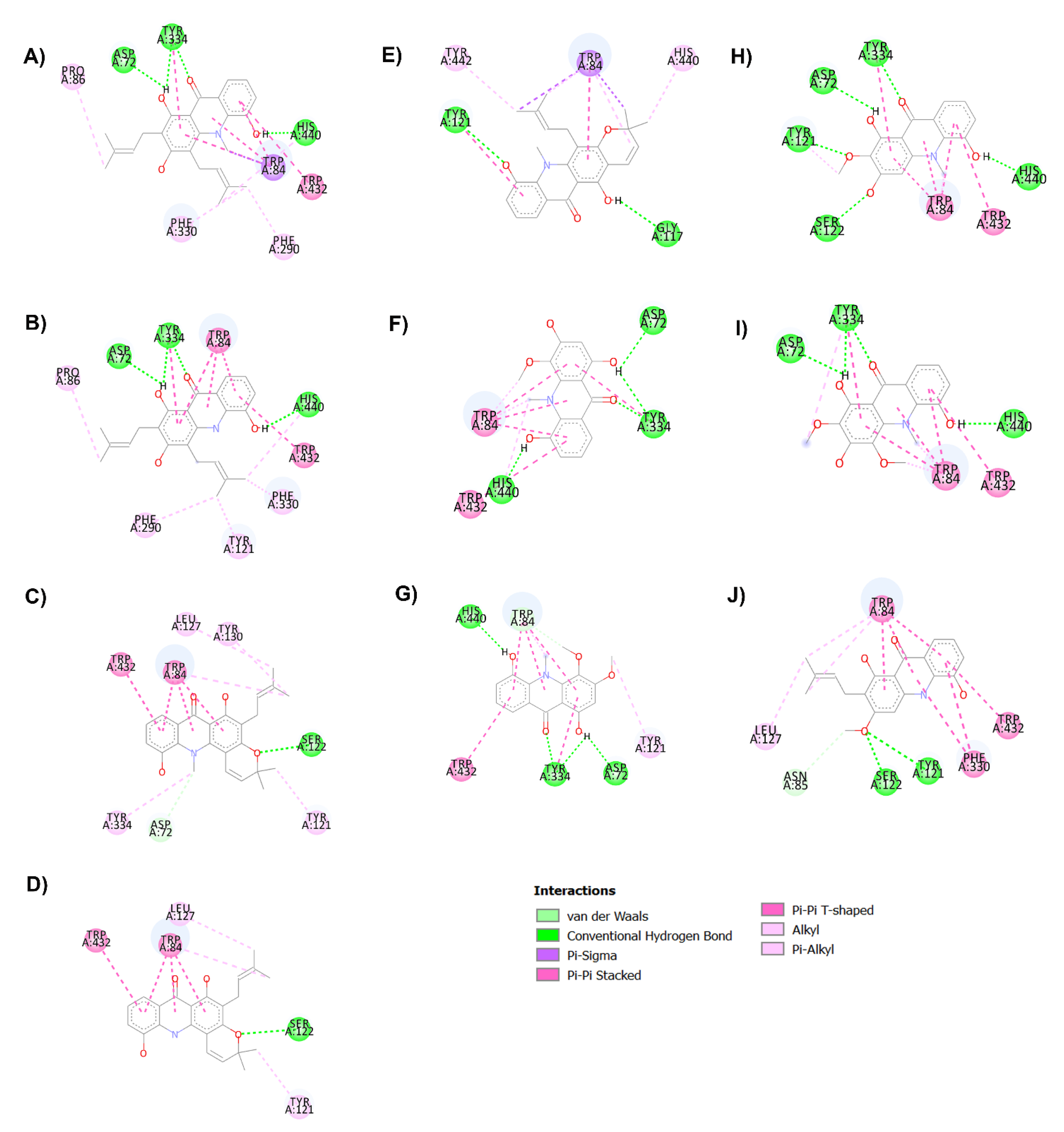
2.3. AChE Inhibition and Molecular Docking Studies
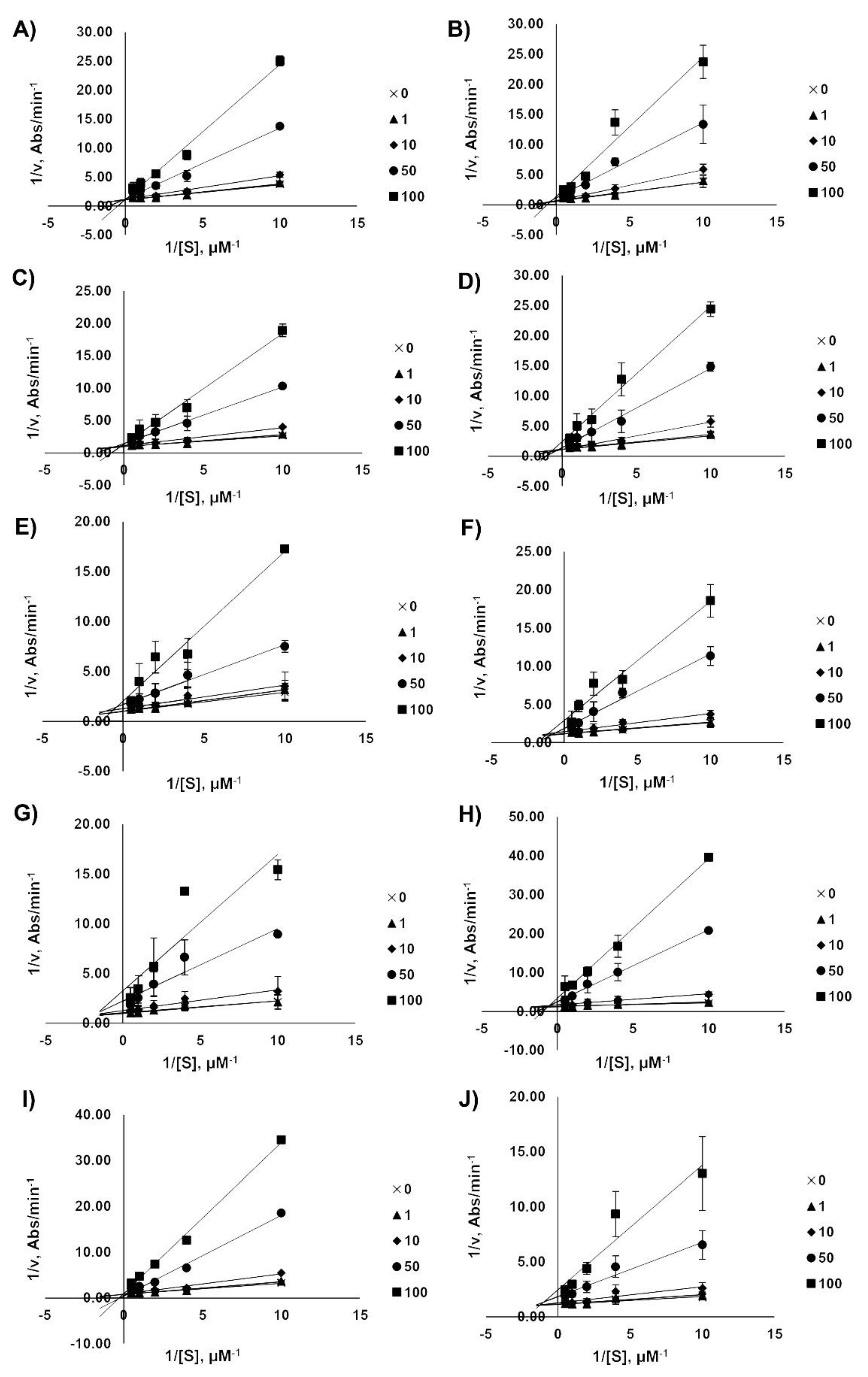
2.4. Kinetic Analysis of AChE Inhibition
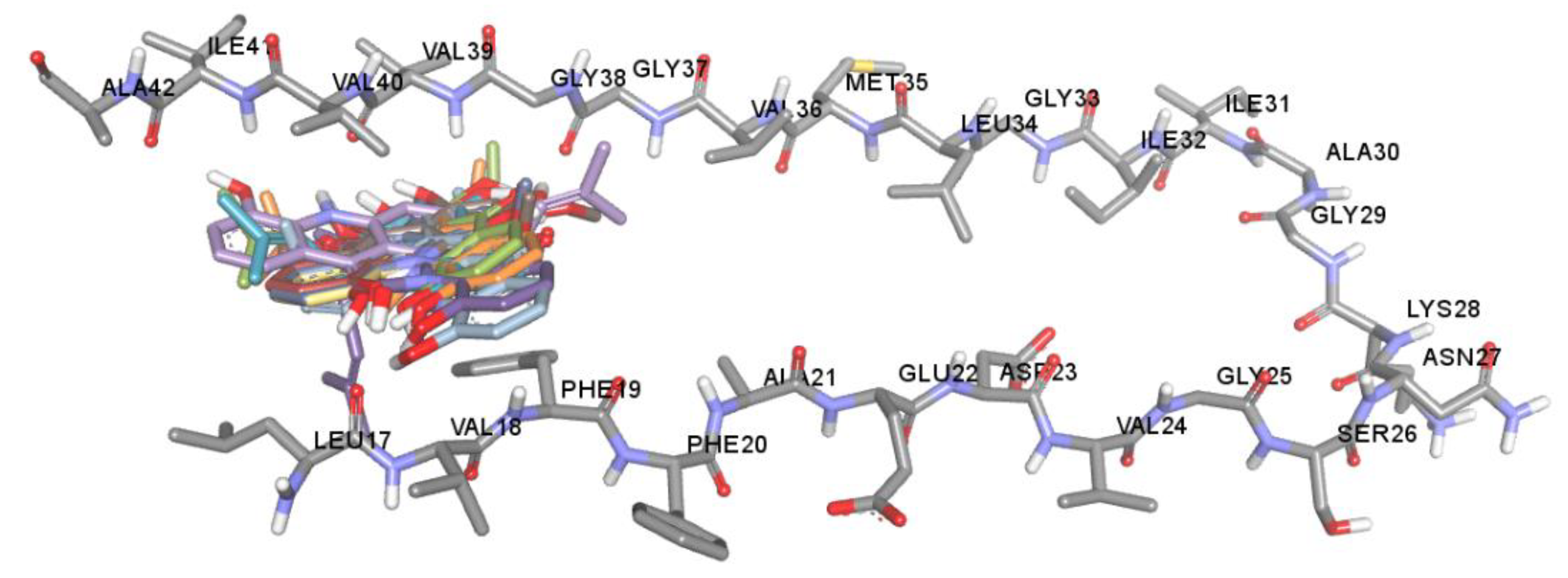
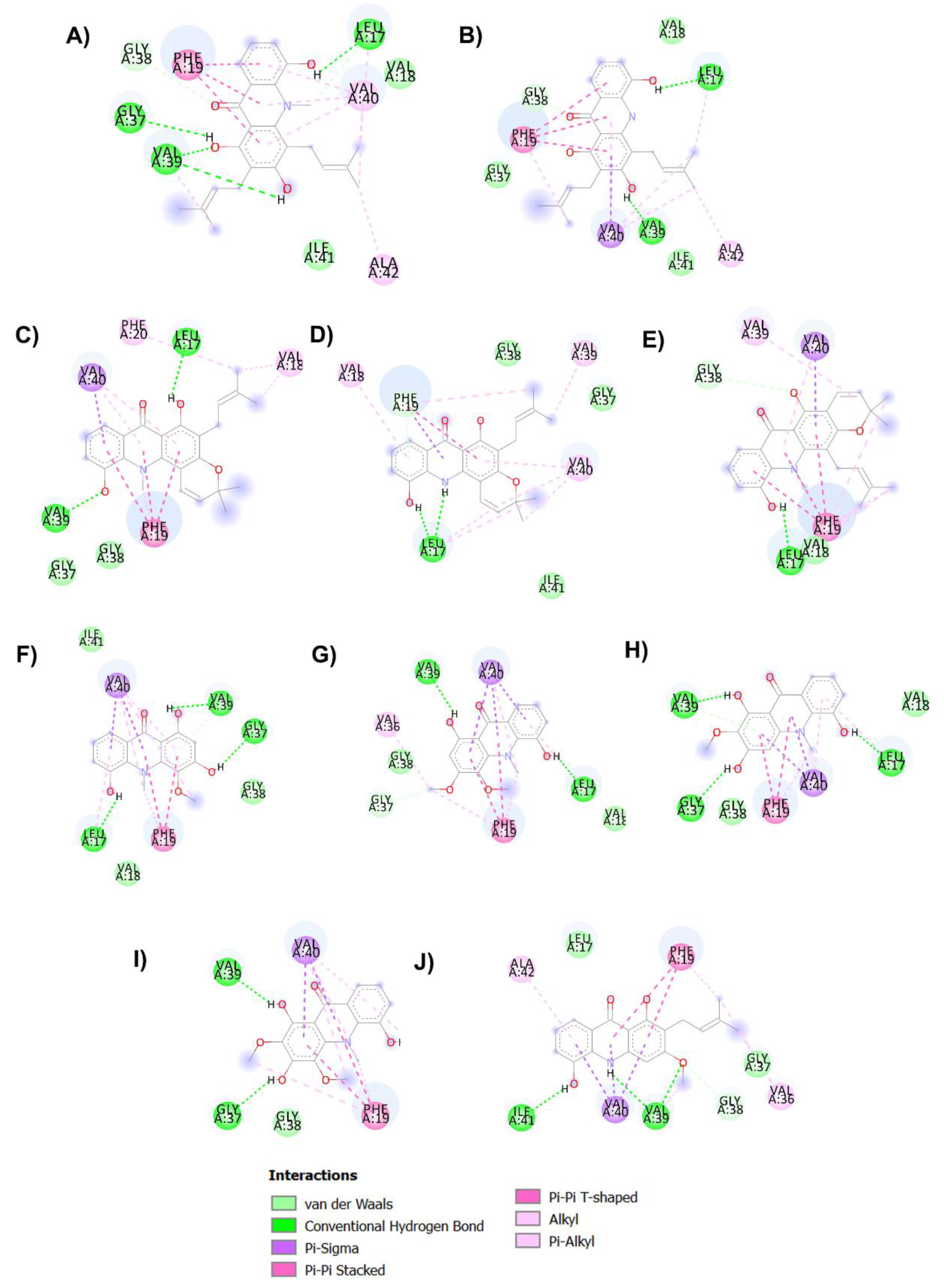
2.5. Anti-Aβ Aggregation Activity and Molecular Docking Studies
2.6. In Silico ADMET Properties of Acridones
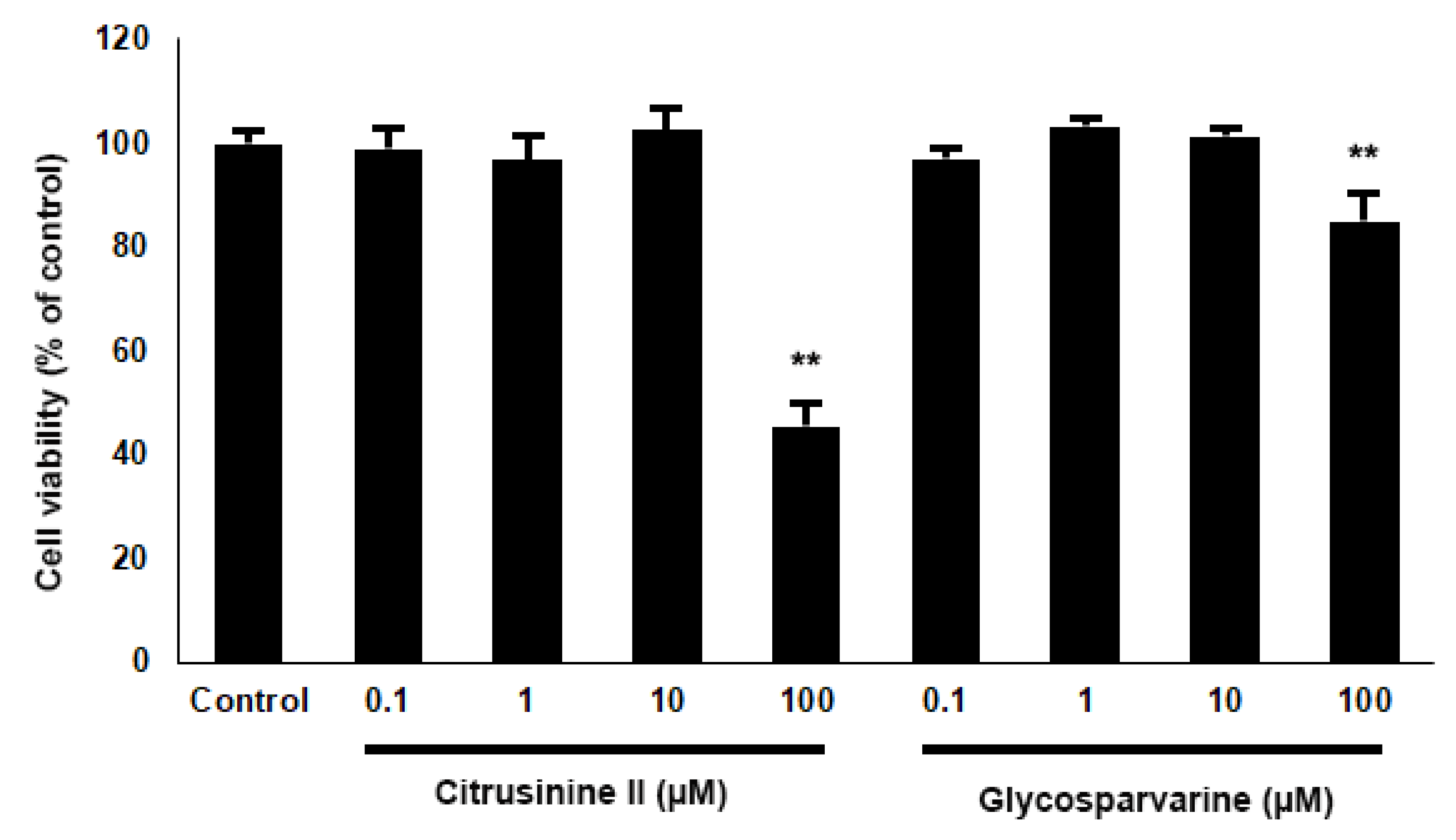
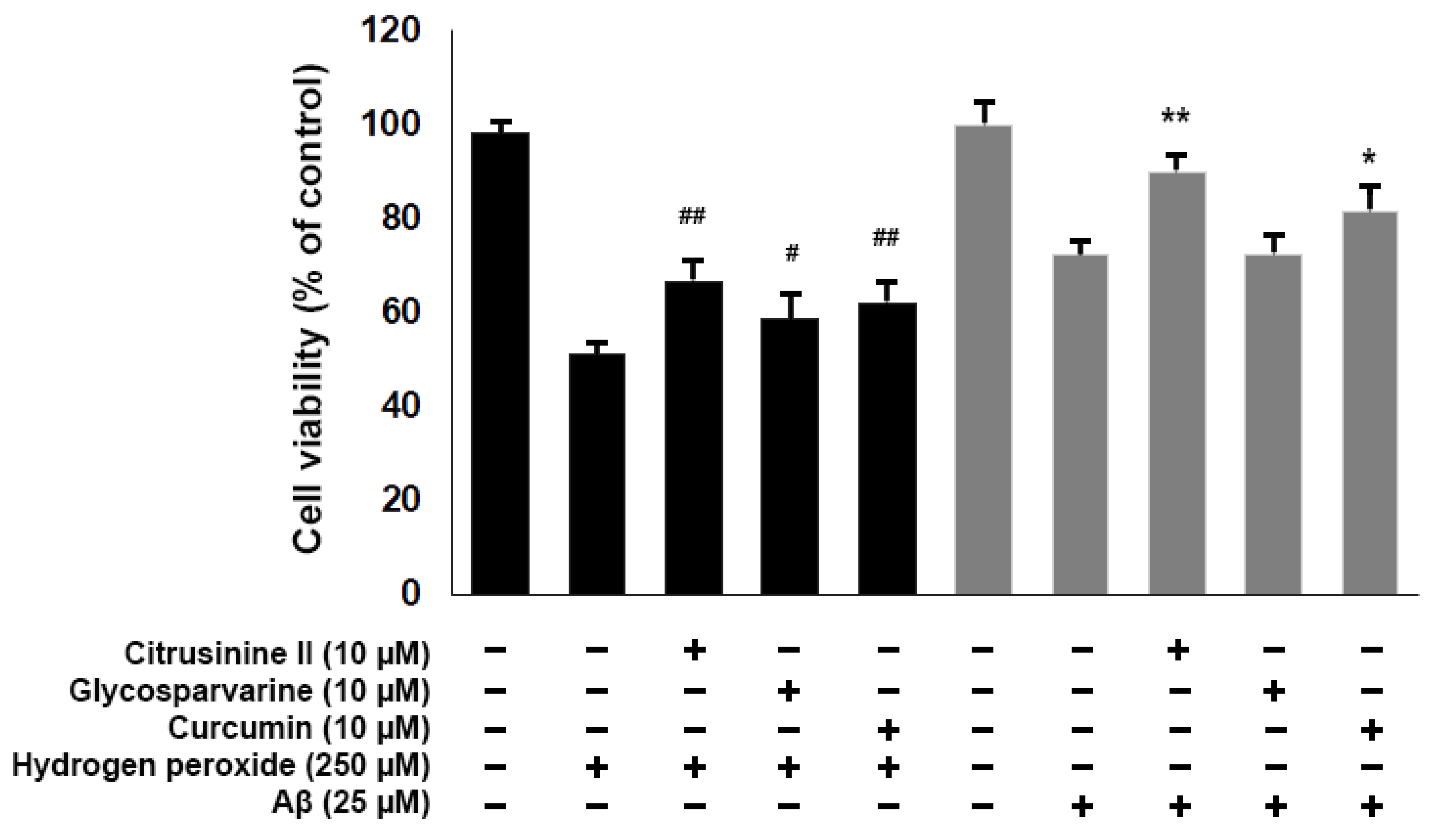
2.7. Neuroprotective Effects against H2O2 and Aβ1-42-Induced Cell Death
3. Materials and Methods
3.1. Acridones and Materials
3.2. In Silico Physicochemical Properties
3.3. In Vitro ABTS•+ Scavenging Activity
3.4. In Vitro AChE Inhibitory Activity
3.5. Kinetic Analysis of AChE Inhibition
3.6. Inhibition of Aβ1-42 Aggregation
3.7. Computational Studies
3.7.1. In Silico Binding Interaction Studies between Acridones and Targets
3.7.2. In Silico Drug ADMET Evaluation
3.8. Neuroprotective Activity against Hydrogen Peroxide (H2O2) and Aβ1-42 Induced Cell Death in Human Neuroblastoma SH-SY5Y Cells
3.9. Statistical Analysis
4. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Desikan, R.S.; Sabuncu, M.R.; Schmansky, N.J.; Reuter, M.; Cabral, H.J.; Hess, C.P.; Weiner, M.W.; Biffi, A.; Anderson, C.D.; Rosand, J.; et al. Selective Disruption of the Cerebral Neocortex in Alzheimer’s Disease. PLoS ONE 2010, 5, e12853. [Google Scholar] [CrossRef]
- Du, X.; Wang, X.; Geng, M. Alzheimer’s disease hypothesis and related therapies. Transl. Neurodegener. 2018, 7, 2. [Google Scholar] [CrossRef] [Green Version]
- Mu, Y.; Gage, F.H. Adult hippocampal neurogenesis and its role in Alzheimer’s disease. Mol. Neurodegener. 2011, 6, 85. [Google Scholar] [CrossRef] [Green Version]
- Tönnies, E.; Trushina, E. Oxidative Stress, Synaptic Dysfunction, and Alzheimer’s Disease. J. Alzheimer’s Dis. 2017, 57, 1105–1121. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Volicer, L. Physiological and pathological functions of beta-amyloid in the brain and alzheimer’s disease: A review. Chin. J. Physiol. 2020, 63, 95–100. [Google Scholar] [CrossRef] [PubMed]
- Murphy, M.P.; Levine, H. Alzheimer’s disease and the amyloid-β peptide. J. Alzheimer’s Dis. 2010, 19, 311–323. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chen, G.F.; Xu, T.H.; Yan, Y.; Zhou, Y.R.; Jiang, Y.; Melcher, K.; Xu, H.E. Amyloid beta: Structure, biology and structure-based therapeutic development. Acta Pharmacol. Sin. 2017, 38, 1205–1235. [Google Scholar] [CrossRef] [PubMed]
- Eanes, E.D.; Glenner, G.G. X-ray diffraction studies on amyloid filaments. J. Histochem. Cytochem. 1968, 16, 673–677. [Google Scholar] [CrossRef]
- Kirschner, D.A.; Abraham, C.; Selkoe, D.J. X-ray diffraction from intraneuronal paired helical filaments and extraneuronal amyloid fibers in Alzheimer disease indicates cross-β conformation. Proc. Natl. Acad. Sci. USA 1986, 83, 503–507. [Google Scholar] [CrossRef] [Green Version]
- Caughey, B.; Lansbury, P.T. Protofibrils, pores, fibrils, and neurodegeneration: Separating the responsible protein aggregates from the innocent bystanders. Annu. Rev. Neurosci. 2003, 26, 267–298. [Google Scholar] [CrossRef] [PubMed]
- Soto, P.; Griffin, M.A.; Shea, J.E. New insights into the mechanism of Alzheimer amyloid-β fibrillogenesis inhibition by N-methylated peptides. Biophys. J. 2007, 93, 3015–3025. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Luhrs, T.; Ritter, C.; Adrian, M.; Riek-Loher, D.; Bohrmann, B.; Dobeli, H.; Schubert, D.; Riek, R. 3D structure of Alzheimer’s amyloid- (1-42) fibrils. Proc. Natl. Acad. Sci. USA 2005, 102, 17342–17347. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Petkova, A.T.; Buntkowsky, G.; Dyda, F.; Leapman, R.D.; Yau, W.M.; Tycko, R. Solid state NMR reveals a pH-dependent antiparallel β-sheet registry in fibrils formed by a β-amyloid peptide. J. Mol. Biol. 2004, 335, 247–260. [Google Scholar] [CrossRef]
- Petkova, A.T.; Yau, W.M.; Tycko, R. Experimental constraints on quaternary structure in Alzheimer’s β-amyloid fibrils. Biochemistry 2006, 45, 498–512. [Google Scholar] [CrossRef] [Green Version]
- Tycko, R. Solid-State NMR as a Probe of Amyloid Structure. Protein Pept. Lett. 2006, 13, 229–234. [Google Scholar] [CrossRef]
- Tycko, R. Solid-state NMR as a probe of amyloid fibril structure. Curr. Opin. Chem. Biol. 2000, 4, 500–506. [Google Scholar] [CrossRef]
- Tycko, R. Molecular structure of amyloid fibrils: Insights from solid-state NMR. Q. Rev. Biophys. 2006, 39, 1–55. [Google Scholar] [CrossRef] [PubMed]
- Kheterpal, I.; Chen, M.; Cook, K.D.; Wetzel, R. Structural Differences in Aβ Amyloid Protofibrils and Fibrils Mapped by Hydrogen Exchange—Mass Spectrometry with On-line Proteolytic Fragmentation. J. Mol. Biol. 2006, 361, 785–795. [Google Scholar] [CrossRef]
- Hampel, H.; Mesulam, M.M.; Cuello, A.C.; Farlow, M.R.; Giacobini, E.; Grossberg, G.T.; Khachaturian, A.S.; Vergallo, A.; Cavedo, E.; Snyder, P.J.; et al. The cholinergic system in the pathophysiology and treatment of Alzheimer’s disease. Brain 2018, 141, 1917–1933. [Google Scholar] [CrossRef]
- Castro, A.; Martinez, A. Peripheral and Dual Binding Site Acetylcholinesterase Inhibitors: Implications in treatment of Alzheimer’s Disease. Mini Rev. Med. Chem. 2001, 1, 267–272. [Google Scholar] [CrossRef] [PubMed]
- Tougu, V. Acetylcholinesterase: Mechanism of Catalysis and Inhibition. Curr. Med. Chem. Nerv. Syst. Agents 2001, 1, 155–170. [Google Scholar] [CrossRef]
- Inestrosa, N.C.; Alvarez, A.; Pérez, C.A.; Moreno, R.D.; Vicente, M.; Linker, C.; Casanueva, O.I.; Soto, C.; Garrido, J. Acetylcholinesterase accelerates assembly of amyloid-beta-peptides into Alzheimer’s fibrils: Possible role of the peripheral site of the enzyme. Neuron 1996, 16, 881–891. [Google Scholar] [CrossRef] [Green Version]
- Alvarez, A.; Opazo, C.; Alarcón, R.; Garrido, J.; Inestrosa, N.C. Acetylcholinesterase promotes the aggregation of amyloid-β-peptide fragments by forming a complex with the growing fibrils. J. Mol. Biol. 1997, 272, 348–361. [Google Scholar] [CrossRef] [PubMed]
- Carlier, P.R.; Chow, E.S.H.; Han, Y.; Liu, J.; El Yazal, J.; Pang, Y.P. Heterodimeric tacrine-based acetylcholinesterase inhibitors: Investigating ligand-peripheral site interactions. J. Med. Chem. 1999, 42, 4225–4231. [Google Scholar] [CrossRef]
- Bartolini, M.; Bertucci, C.; Cavrini, V.; Andrisano, V. β-Amyloid aggregation induced by human acetylcholinesterase: Inhibition studies. Biochem. Pharmacol. 2003, 65, 407–416. [Google Scholar] [CrossRef]
- Piazzi, L.; Rampa, A.; Bisi, A.; Gobbi, S.; Belluti, F.; Cavalli, A.; Bartolini, M.; Andrisano, V.; Valenti, P.; Recanatini, M. 3-(4-{[benzyl(methyl)amino]methyl}-phenyl)-6,7-dimethoxy-2H-2-chromenone (AP2238) inhibits both acetylcholinesterase and acetylcholinesterase-induced β-amyloid aggregation: A dual function lead for Alzheimer’s disease therapy. J. Med. Chem. 2003, 46, 2279–2282. [Google Scholar] [CrossRef]
- Christen, Y. Oxidative stress and Alzheimer disease. Am. J. Clin. Nutr. 2000, 71, 621S–629S. [Google Scholar] [CrossRef]
- Praticò, D.; Uryu, K.; Leight, S.; Trojanoswki, J.Q.; Lee, V.M.Y. Increased lipid peroxidation precedes amyloid plaque formation in an animal model of alzheimer amyloidosis. J. Neurosci. 2001, 21, 4183–4187. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Federico, A.; Cardaioli, E.; Da Pozzo, P.; Formichi, P.; Gallus, G.N.; Radi, E. Mitochondria, oxidative stress and neurodegeneration. J. Neurol. Sci. 2012, 322, 254–262. [Google Scholar] [CrossRef]
- Yan, M.H.; Wang, X.; Zhu, X. Mitochondrial defects and oxidative stress in Alzheimer disease and Parkinson disease. Free Radic. Biol. Med. 2013, 62, 90–101. [Google Scholar] [CrossRef] [Green Version]
- Cassidy, L.; Fernandez, F.; Johnson, J.B.; Naiker, M.; Owoola, A.G.; Broszczak, D.A. Oxidative stress in alzheimer’s disease: A review on emergent natural polyphenolic therapeutics. Complement. Ther. Med. 2020, 49, 1–11. [Google Scholar] [CrossRef]
- Allan Butterfield, D.; Castegna, A.; Lauderback, C.M.; Drake, J. Evidence that amyloid beta-peptide-induced lipid peroxidation and its sequelae in Alzheimer’s disease brain contribute to neuronal death. Neurobiol. Aging 2002, 23, 655–664. [Google Scholar] [CrossRef]
- Butterfield, D.A.; Boyd-Kimball, D. Redox proteomics and amyloid β-peptide: Insights into Alzheimer disease. J. Neurochem. 2019, 151, 459–487. [Google Scholar] [CrossRef] [Green Version]
- Butterfield, D.A. Amyloid β-peptide (1-42)-induced oxidative stress and neurotoxicity: Implications for neurodegeneration in Alzheimer’s disease brain. A review. Free Radic. Res. 2002, 36, 1307–1313. [Google Scholar] [CrossRef]
- Grundman, M.; Delaney, P. Antioxidant strategies for Alzheimer’s disease. Proc. Nutr. Soc. 2002, 61, 191–202. [Google Scholar] [CrossRef] [Green Version]
- Staehelin, H.B. Micronutrients and Alzheimer’s disease. Proc. Nutr. Soc. 2005, 64, 565–570. [Google Scholar] [CrossRef] [Green Version]
- Davis, K.L.; Powchick, P. Tacrine. Lancet 1995, 345, 625–630. [Google Scholar] [CrossRef]
- Crismon, M.L. Tacrine: First drug approved for alzheimer’s disease. Ann. Pharmacother. 1994, 28, 744–751. [Google Scholar] [CrossRef] [PubMed]
- Qizilbash, N.; Birks, J.; Lopez, A.J.; Lewington, S.; Szeto, S. Tacrine for Alzheimer’s disease (Withdrawn paper. 1999, art. no. CD000202). Cochrane Database Syst. Rev. 2007, 18, 1–4. [Google Scholar]
- Cholewiñski, G.; Dzierzbicka, K.; Kolodziejczyk, A.M. Natural and synthetic acridines/acridones as antitumor agents: Their biological activities and methods of synthesis. Pharmacol. Rep. 2011, 63, 305–336. [Google Scholar] [CrossRef]
- Wouatsa, V.N.; Misra, L.; Kumar, S.; Prakash, O.; Khan, F.; Tchoumbougnang, F.; Venkatesh, R.K. Aromatase and glycosyl transferase inhibiting acridone alkaloids from fruits of Cameroonian Zanthoxylum species. Chem. Cent. J. 2013, 7, 125. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Han, Y.; Luo, A.; Kamau, P.; Takomthong, P.; Hu, J.; Boonyarat, C.; Luo, L.; Lai, R. A plant-derived TRPV3 inhibitor suppresses pain and itch. Br. J. Pharmacol. 2021, 178, 1669–1683. [Google Scholar] [CrossRef]
- Yamamoto, N.; Furukawa, H.; Ito, Y.; Yoshida, S.; Maeno, K.; Nishiyama, Y. Anti-herpesvirus activity of citrusinine-I, a new acridone alkaloid, and related compounds. Antiviral Res. 1989, 12, 21–36. [Google Scholar] [CrossRef]
- Parveen, M.; Aslam, A.; Nami, S.A.A.; Malla, A.M.; Alam, M.; Lee, D.-U.; Rehman, S.; Silva, P.S.P.; Silva, M.R. Potent acetylcholinesterase inhibitors: Synthesis, biological assay and docking study of nitro acridone derivatives. J. Photochem. Photobiol. B Biol. 2016, 161, 304–311. [Google Scholar] [CrossRef]
- Sombatsri, A.; Thummanant, Y.; Sribuhom, T.; Boonmak, J.; Youngme, S.; Phusrisom, S.; Kukongviriyapan, V.; Yenjai, C. New limonophyllines A-C from the stem of Atalantia monophylla and cytotoxicity against cholangiocarcinoma and HepG2 cell lines. Arch. Pharm. Res. 2018, 41, 431–437. [Google Scholar] [CrossRef] [PubMed]
- Lipinski, C.A.; Lombardo, F.; Dominy, B.W.; Feeney, P.J. Experimental and computational approaches to estimate solubility and permeability in drug discovery and development settings. Adv. Drug Deliv. Rev. 2001, 46, 3–26. [Google Scholar] [CrossRef]
- Veber, D.F.; Johnson, S.R.; Cheng, H.Y.; Smith, B.R.; Ward, K.W.; Kopple, K.D. Molecular properties that influence the oral bioavailability of drug candidates. J. Med. Chem. 2002, 45, 2615–2623. [Google Scholar] [CrossRef] [PubMed]
- Vajragupta, O.; Toasaksiri, S.; Boonyarat, C.; Wongkrajang, Y.; Peungvicha, P.; Watanabe, H.; Boonchoong, P. Chroman amide and nicotinyl amide derivatives: Inhibition of lipid peroxidation and protection against head trauma. Free Radic. Res. 2000, 32, 145–155. [Google Scholar] [CrossRef]
- Blokland, A.; Geraerts, E.; Been, M. A detailed analysis of rats’ spatial memory in a probe trial of a Morris task. Behav. Brain Res. 2004, 154, 71–75. [Google Scholar] [CrossRef]
- Hasselmo, M.E. The role of acetylcholine in learning and memory. Curr. Opin. Neurobiol. 2006, 16, 710–715. [Google Scholar] [CrossRef] [Green Version]
- Ahmed, M.; Davis, J.; Aucoin, D.; Sato, T.; Ahuja, S.; Aimoto, S.; Elliott, J.I.; Van Nostrand, W.E.; Smith, S.O. Structural conversion of neurotoxic amyloid-Β 1-42 oligomers to fibrils. Nat. Struct. Mol. Biol. 2010, 17, 561–567. [Google Scholar] [CrossRef] [Green Version]
- Antzutkin, O.N.; Leapman, R.D.; Balbach, J.J.; Tycko, R. Supramolecular structural constraints on Alzheimer’s β-amyloid fibrils from electron microscopy and solid-state nuclear magnetic resonance. Biochemistry 2002, 41, 15436–15450. [Google Scholar] [CrossRef] [PubMed]
- Marina, G.B.; Kirkitadze, D.; Lomakin, A.; Vollers, S.S.; Benedek, G.B.; Teplow, D.B. Amyloid β-protein (Aβ) assembly: Aβ40 and Aβ42 oligomerize through distinct pathways. Proc. Natl. Acad. Sci. USA 2003, 100, 330–335. [Google Scholar]
- Urbanc, B.; Cruz, L.; Yun, S.; Buldyrev, S.V.; Bitan, G.; Teplow, D.B.; Stanley, H.E. In silico study of amyloid β-protein folding and oligomerization. Proc. Natl. Acad. Sci. USA 2004, 101, 17345–17350. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pires, D.E.V.; Blundell, T.L.; Ascher, D.B. pkCSM: Predicting small-molecule pharmacokinetic and toxicity properties using graph-based signatures. J. Med. Chem. 2015, 58, 4066–4072. [Google Scholar] [CrossRef]
- Ohashi, R.; Watanabe, R.; Esaki, T.; Taniguchi, T.; Torimoto-Katori, N.; Watanabe, T.; Ogasawara, Y.; Takahashi, T.; Tsukimoto, M.; Mizuguchi, K. Development of Simplified in Vitro P-Glycoprotein Substrate Assay and in Silico Prediction Models to Evaluate Transport Potential of P-Glycoprotein. Mol. Pharm. 2019, 16, 1851–1863. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Furge, L.L.; Guengerich, F.P. Cytochrome P450 enzymes in drug metabolism and chemical toxicology: An introduction. Biochem. Mol. Biol. Educ. 2006, 34, 66–74. [Google Scholar] [CrossRef]
- Tumiatti, V.; Minarini, A.; Bolognesi, M.L.; Milelli, A.; Rosini, M.; Melchiorre, C. Tacrine Derivatives and Alzheimers Disease. Curr. Med. Chem. 2010, 17, 1825–1838. [Google Scholar] [CrossRef]
- Daina, A.; Michielin, O.; Zoete, V. SwissADME: A free web tool to evaluate pharmacokinetics, drug-likeness and medicinal chemistry friendliness of small molecules. Sci. Rep. 2017, 7, 1–13. [Google Scholar]
- Chheng, C.; Waiwut, P.; Plekratoke, K.; Chulikhit, Y.; Daodee, S.; Monthakantirat, O.; Pitiporn, S.; Musigavong, N.; Kwankhao, P.; Boonyarat, C. Multitarget activities of kleeb bua daeng, a Thai traditional herbal formula, against alzheimer’s disease. Pharmaceuticals 2020, 13, 79. [Google Scholar] [CrossRef]
- Ellman, G.L.; Courtney, K.D.; Andres, V.; Featherstone, R.M. A new and rapid colorimetric determination of acetylcholinesterase activity. Biochem. Pharmacol. 1961, 7, 88–95. [Google Scholar] [CrossRef]
- LeVine, H. Quantification of beta-sheet amyloid fibril structures with thioflavin T. Methods Enzymol. 1999, 309, 274–284. [Google Scholar] [PubMed]
- Khamphukdee, C.; Monthakantirat, O.; Chulikhit, Y.; Buttachon, S.; Lee, M.; Silva, A.M.S.; Sekeroglu, N.; Kijjoa, A. Chemical Constituents and Antidepressant-Like Effects in Ovariectomized Mice of the Ethanol Extract of Alternanthera philoxeroides. Molecules 2018, 23, 2202. [Google Scholar] [CrossRef] [Green Version]
- Takomthong, P.; Waiwut, P.; Yenjai, C.; Sripanidkulchai, B.; Reubroycharoen, P.; Lai, R.; Kamau, P.; Boonyarat, C. Structure-activity analysis and molecular docking studies of coumarins from toddalia asiatica as multifunctional agents for alzheimer’s disease. Biomedicines 2020, 8, 107. [Google Scholar] [CrossRef] [PubMed]
- Thiratmatrakul, S.; Yenjai, C.; Waiwut, P.; Vajragupta, O.; Reubroycharoen, P.; Tohda, M.; Boonyarat, C. Synthesis, biological evaluation and molecular modeling study of novel tacrine-carbazole hybrids as potential multifunctional agents for the treatment of Alzheimer’s disease. Eur. J. Med. Chem. 2014, 75, 21–30. [Google Scholar] [CrossRef]









{kind=link}
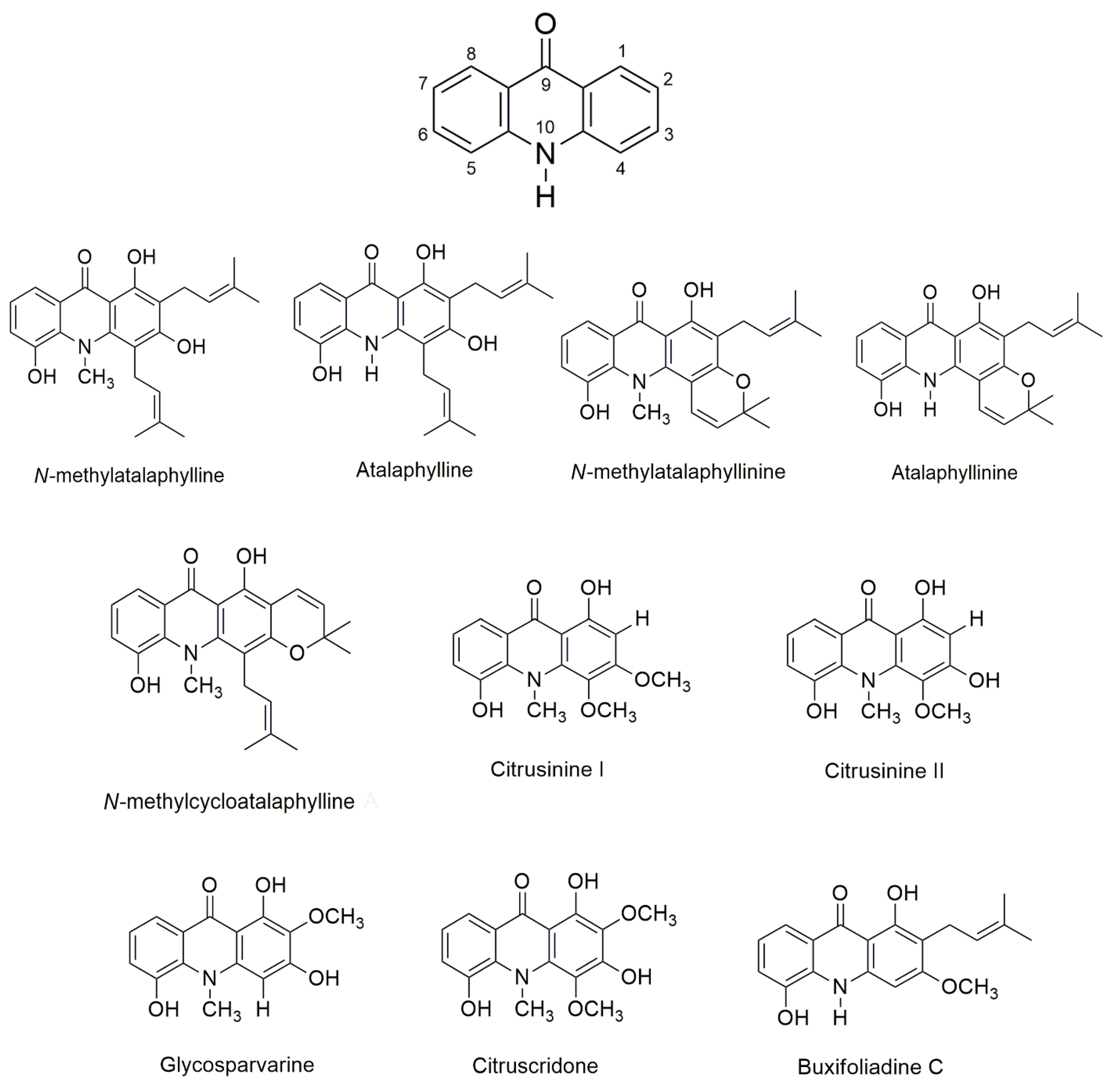{kind=link}
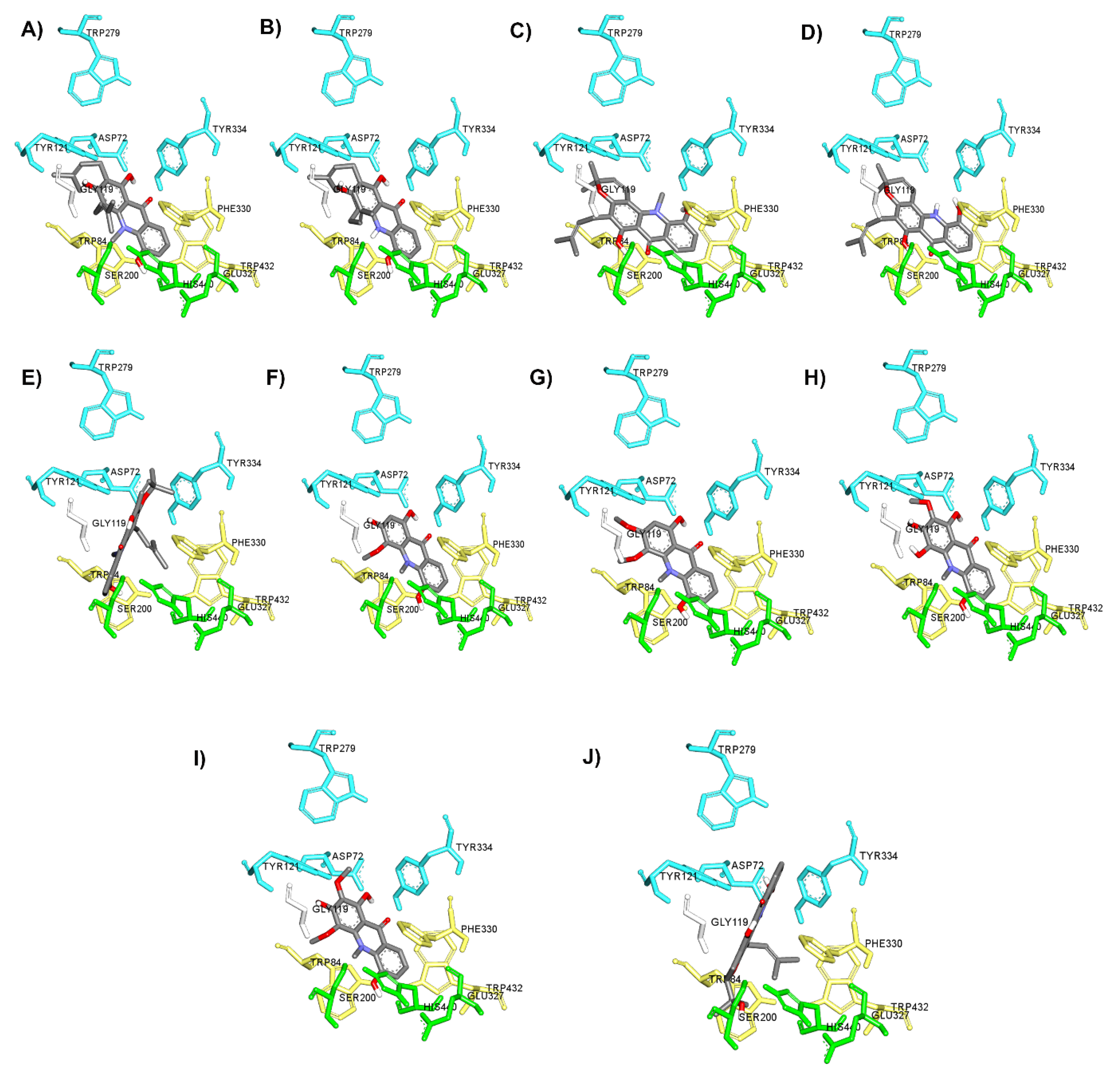{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Compounds | Physicochemical Properties | |||||
|---|---|---|---|---|---|---|
| MW (g/mol) | TPSA a (Å2) | Num. Rotatable Bonds | Num. H-Bond Acceptors | Num. H-Bond Donors | Log Po/w b | |
| N-methylatalaphylline | 393.48 | 82.69 | 4 | 4 | 3 | 4.55 |
| Atalaphylline | 379.45 | 93.55 | 4 | 4 | 4 | 4.61 |
| N-methylatalaphyllinine | 391.46 | 71.69 | 2 | 4 | 2 | 4.31 |
| Atalaphyllinine | 379.45 | 82.55 | 2 | 4 | 3 | 4.51 |
| N-methylcycloatalaphylline A | 391.46 | 71.69 | 2 | 4 | 2 | 4.37 |
| Citrusinine II | 287.27 | 91.92 | 1 | 5 | 3 | 1.68 |
| Citrusinine I | 301.29 | 80.92 | 2 | 5 | 2 | 2.04 |
| Glycosparvarine | 287.37 | 91.92 | 1 | 5 | 3 | 1.66 |
| Citruscridone | 317.29 | 101.15 | 2 | 6 | 3 | 1.72 |
| Buxifoliadine C | 325.36 | 82.55 | 3 | 4 | 3 | 4.55 |
| Compounds | ABTS Scavenging Activity | Anti-AChE | Anti-Aβ Aggregation |
|---|---|---|---|
| N-methylatalaphylline | 50.73 ± 1.02 c,d | 49.08 ± 2.75 d,e | 7.32 ± 0.09 |
| Atalaphylline | 46.24 ± 2.31 c | 58.33 ± 2.99 f,g,h | 6.58 ± 1.30 |
| N-methylatalaphyllinine | 57.53 ± 3.52 d,e | 54.04 ± 2.03 e,f,g | 6.14 ± 3.85 |
| Atalaphyllinine | 52.96 ± 1.83 c,d,e | 60.78 ± 4.62 g,h | 4.79 ± 0.67 |
| N-methylcycloatalaphylline A | 79.58 ± 3.26 f | 66.62 ± 2.51 h | 5.84 ± 0.99 |
| Citrusinine II | 19.98 ± 3.89 a | 42.56 ± 4.81 c,d | 6.17 ± 2.29 |
| Citrusinine I | 33.77 ± 0.29 b | 35.37 ± 1.61 c | 7.05 ± 1.45 |
| Glycosparvarine | 22.88 ± 2.24 a | 18.61 ± 2.77 b | 6.58 ± 2.03 |
| Citruscridone | 31.69 ± 0.56 b | 46.96 ± 2.40 d,e | 8.61 ± 0.75 |
| Buxifoliadine C | 21.79 ± 2.71 a | 50.81 ± 3.74 d,e,f | 8.81 ± 3.62 |
| Tacrine | nd | 0.28 ± 0.04 a | nd |
| Trolox | 58.92 ± 2.38 e | nd | nd |
| Curcumin | nd | nd | 4.98 ± 0.72 |
| Name | Absorption | Distribution | Metabolism | Excretion | Toxicity | ||
|---|---|---|---|---|---|---|---|
| Intestinal Absorption (Human) (%Absorbed) | BBB Permeability (Log BB) | CYP2C9 Inhibitor | CYP3A4 Inhibitor | Total Clearance (Log mL/min/kg) | Renal OCT2 Substrate | Hepatotoxicity | |
| N-methylatalaphylline | 96.252 | −0.864 | Yes | No | 0.401 | No | Yes |
| Atalaphylline | 85.288 | −0.887 | Yes | No | 0.325 | No | Yes |
| N-methylatalaphyllinine | 94.544 | 0.169 | Yes | Yes | 0.293 | No | Yes |
| Atalaphyllinine | 91.158 | −0.734 | Yes | Yes | 0.155 | No | Yes |
| N-methylcycloatalaphylline A | 94.951 | 0.244 | Yes | Yes | 0.256 | No | Yes |
| Citrusinine II | 89.485 | −0.996 | No | No | 0.309 | No | No |
| Citrusinine I | 99.555 | −0.197 | Yes | No | 0.423 | No | Yes |
| Glycosparvarine | 79.129 | −0.948 | No | No | 0.316 | No | No |
| Citruscridone | 68.876 | −1.082 | Yes | No | 0.198 | No | No |
| Buxifoliadine C | 93.124 | −0.789 | Yes | No | 0.402 | No | Yes |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Takomthong, P.; Waiwut, P.; Yenjai, C.; Sombatsri, A.; Reubroycharoen, P.; Lei, L.; Lai, R.; Chaiwiwatrakul, S.; Boonyarat, C. Multi-Target Actions of Acridones from Atalantia monophylla towards Alzheimer’s Pathogenesis and Their Pharmacokinetic Properties. Pharmaceuticals 2021, 14, 888. https://doi.org/10.3390/ph14090888
Takomthong P, Waiwut P, Yenjai C, Sombatsri A, Reubroycharoen P, Lei L, Lai R, Chaiwiwatrakul S, Boonyarat C. Multi-Target Actions of Acridones from Atalantia monophylla towards Alzheimer’s Pathogenesis and Their Pharmacokinetic Properties. Pharmaceuticals. 2021; 14(9):888. https://doi.org/10.3390/ph14090888
Chicago/Turabian StyleTakomthong, Pitchayakarn, Pornthip Waiwut, Chavi Yenjai, Aonnicha Sombatsri, Prasert Reubroycharoen, Luo Lei, Ren Lai, Suchada Chaiwiwatrakul, and Chantana Boonyarat. 2021. "Multi-Target Actions of Acridones from Atalantia monophylla towards Alzheimer’s Pathogenesis and Their Pharmacokinetic Properties" Pharmaceuticals 14, no. 9: 888. https://doi.org/10.3390/ph14090888
APA StyleTakomthong, P., Waiwut, P., Yenjai, C., Sombatsri, A., Reubroycharoen, P., Lei, L., Lai, R., Chaiwiwatrakul, S., & Boonyarat, C. (2021). Multi-Target Actions of Acridones from Atalantia monophylla towards Alzheimer’s Pathogenesis and Their Pharmacokinetic Properties. Pharmaceuticals, 14(9), 888. https://doi.org/10.3390/ph14090888