

Bacterial-Specific Aggregation and Killing of Immunomodulatory Host Defense Peptides



Abstract
:
1. Introduction
2. Results and Discussion
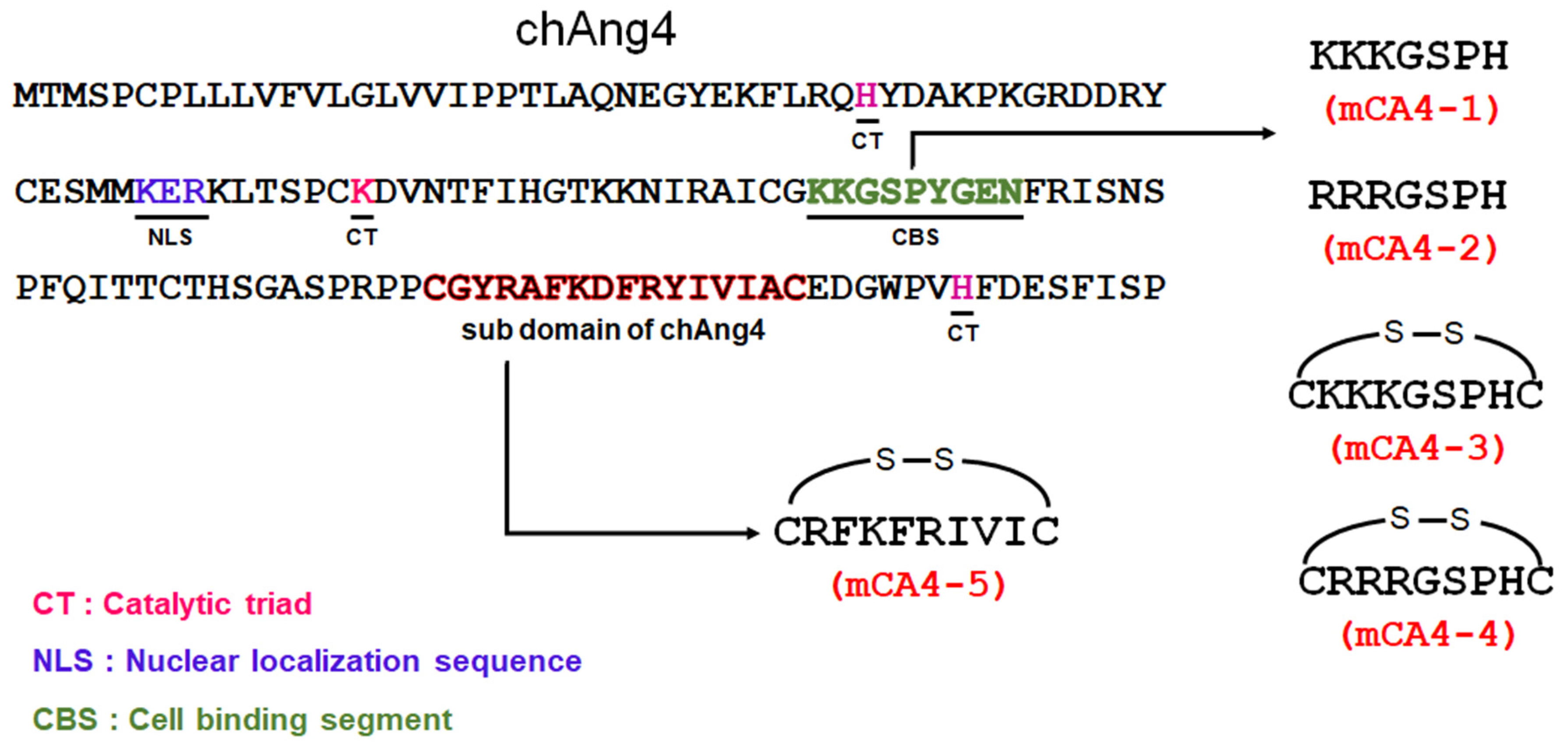
2.1. Peptide Synthesis and Characterization
2.2. Bacterial Toxicity and Uptake
2.3. Membrane Permeabilization
2.4. Role of Disulfide Bridges in Antibacterial Activity
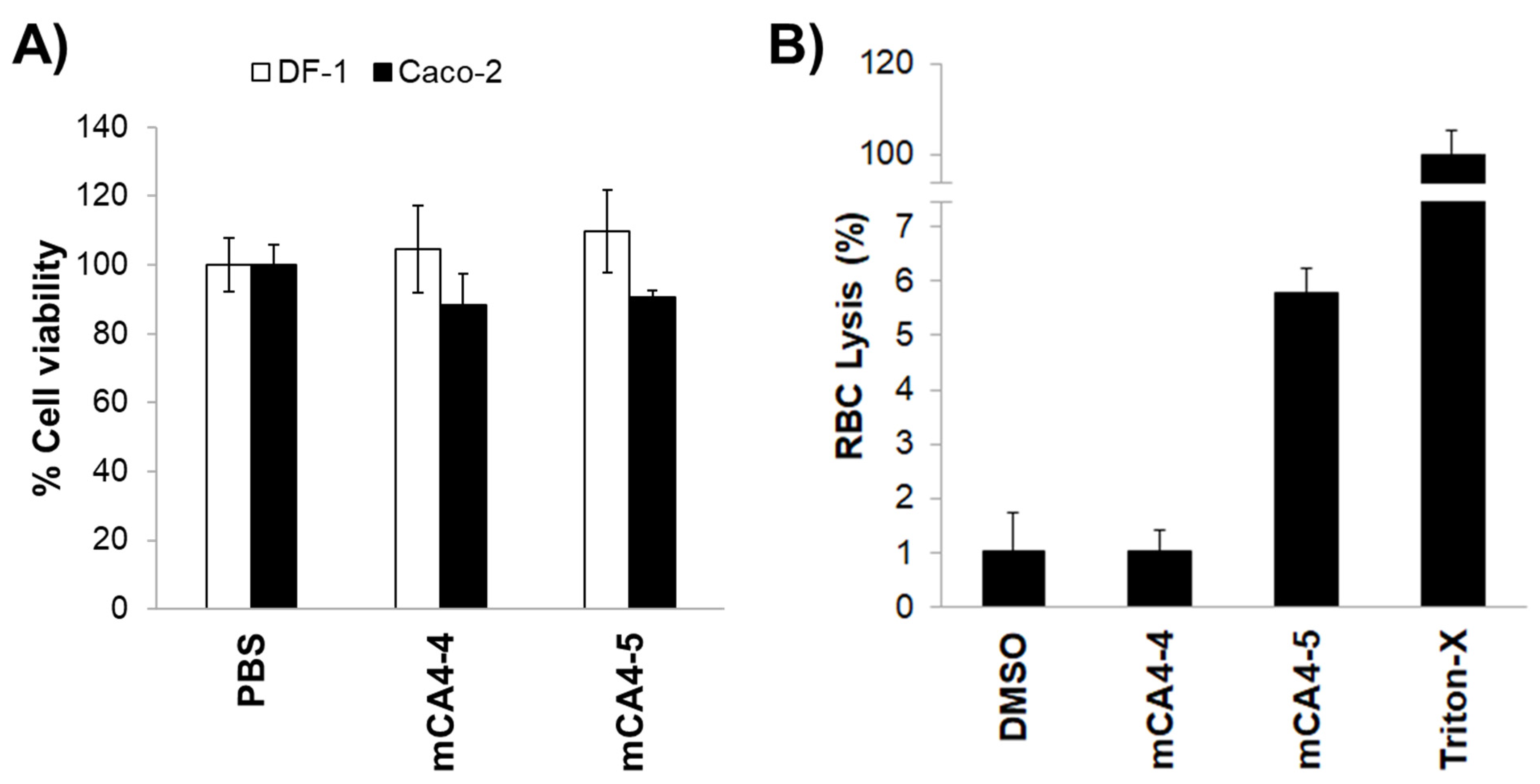
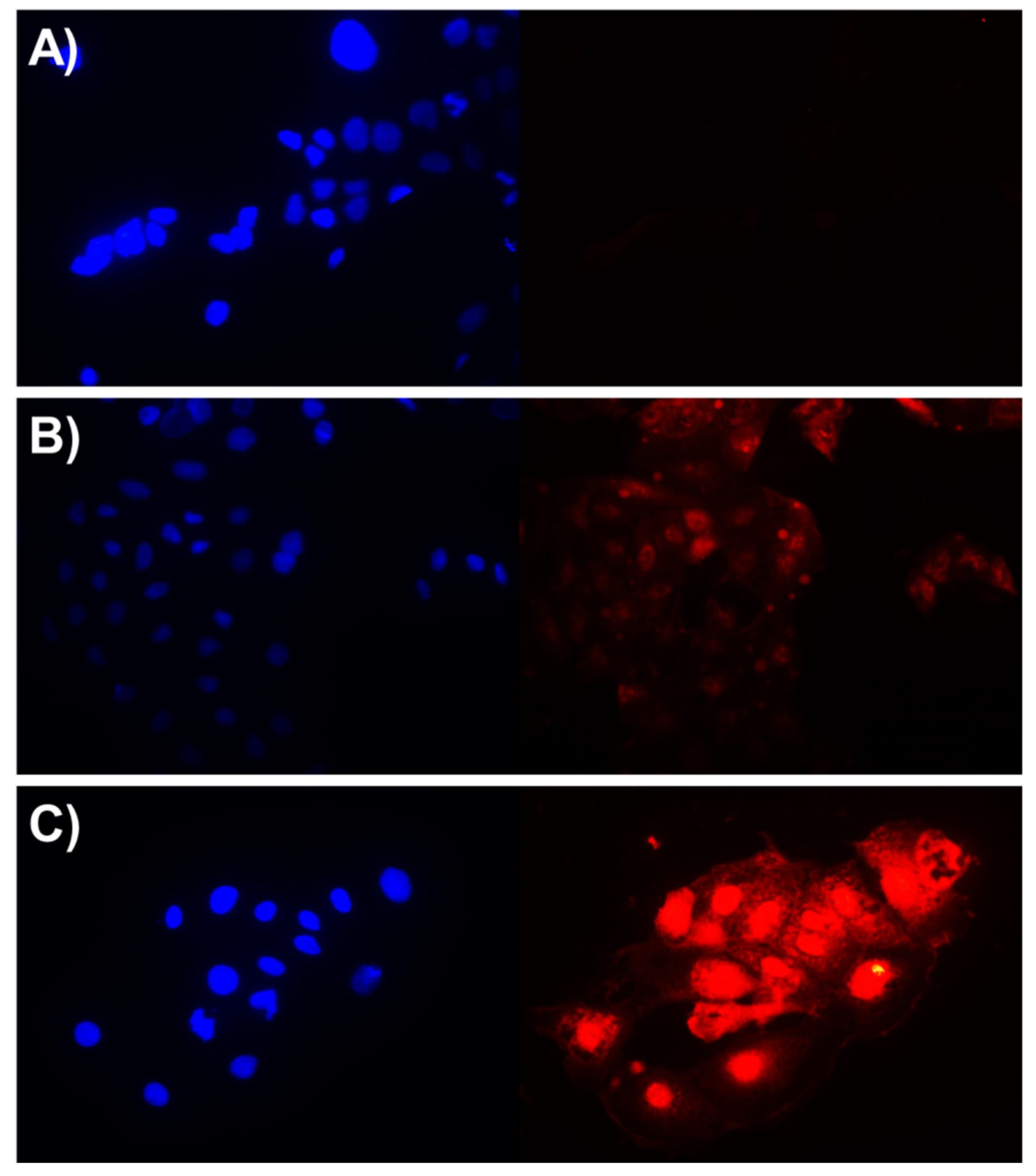
2.5. Cellular Uptake, Cytotoxicity, and Hemolytic Activities
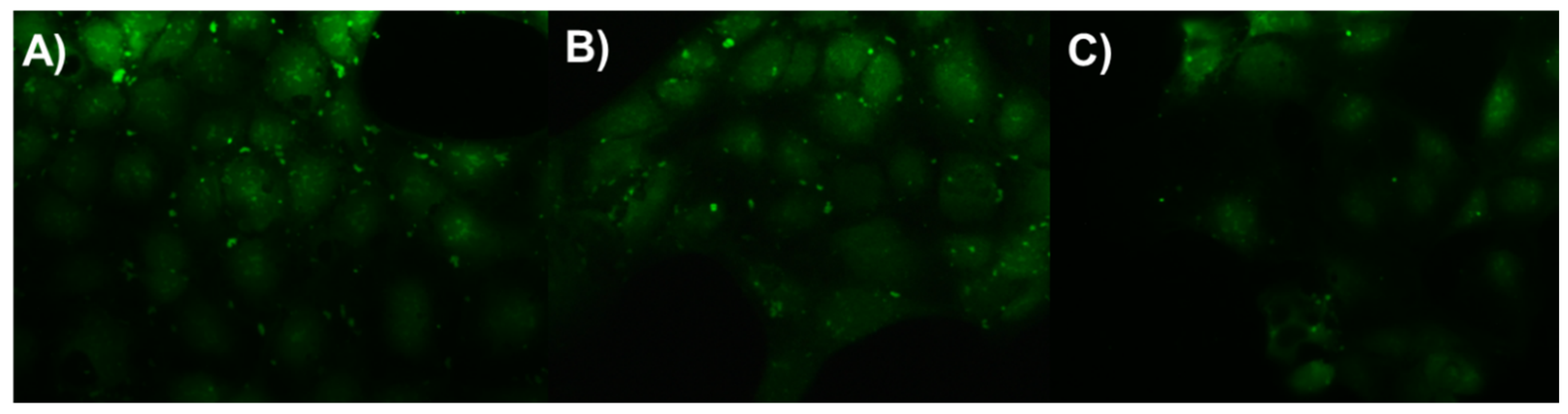
2.6. Intracellular Bacterial Treatment
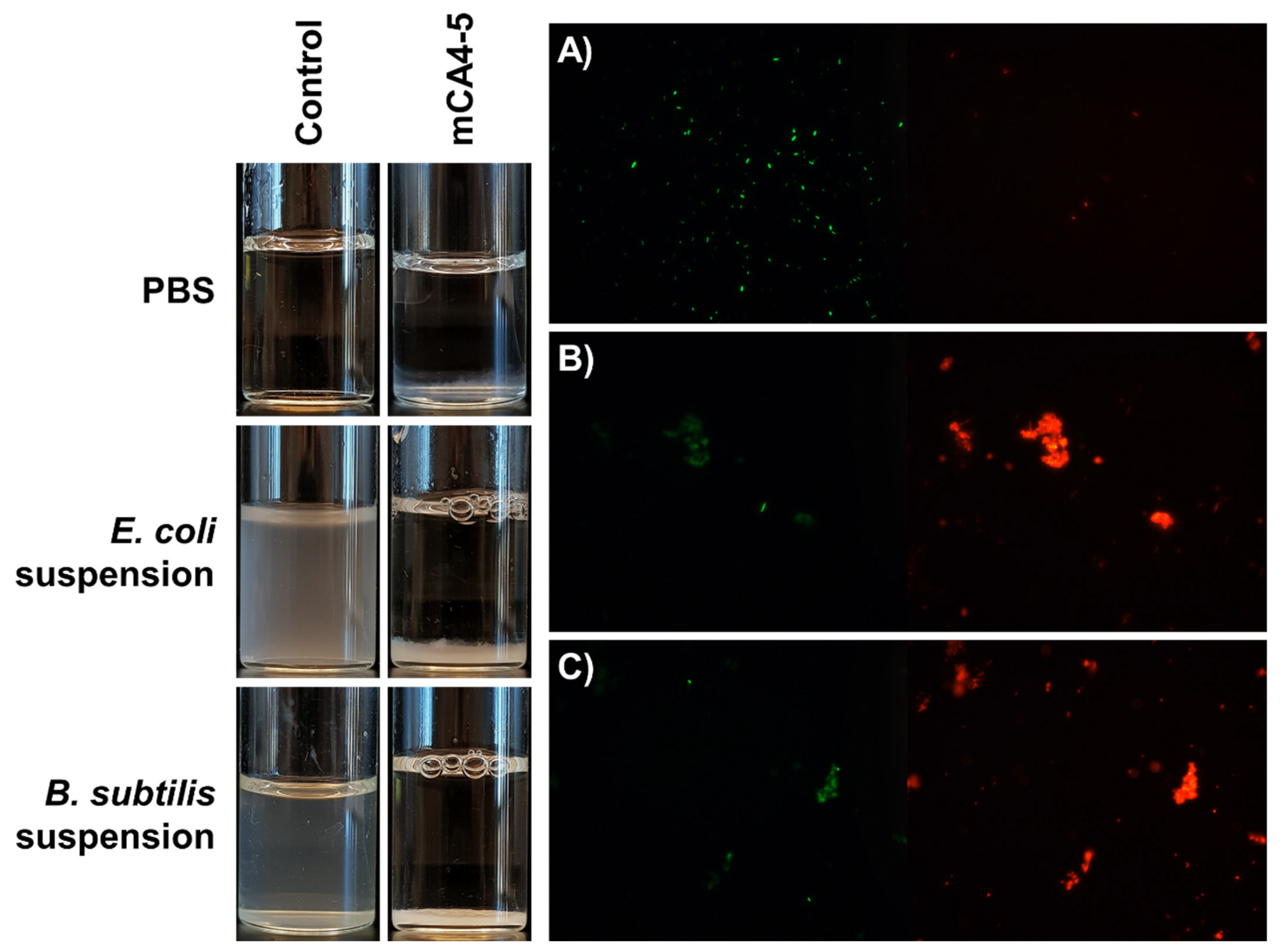
2.7. Bacterial Flocculation Capability
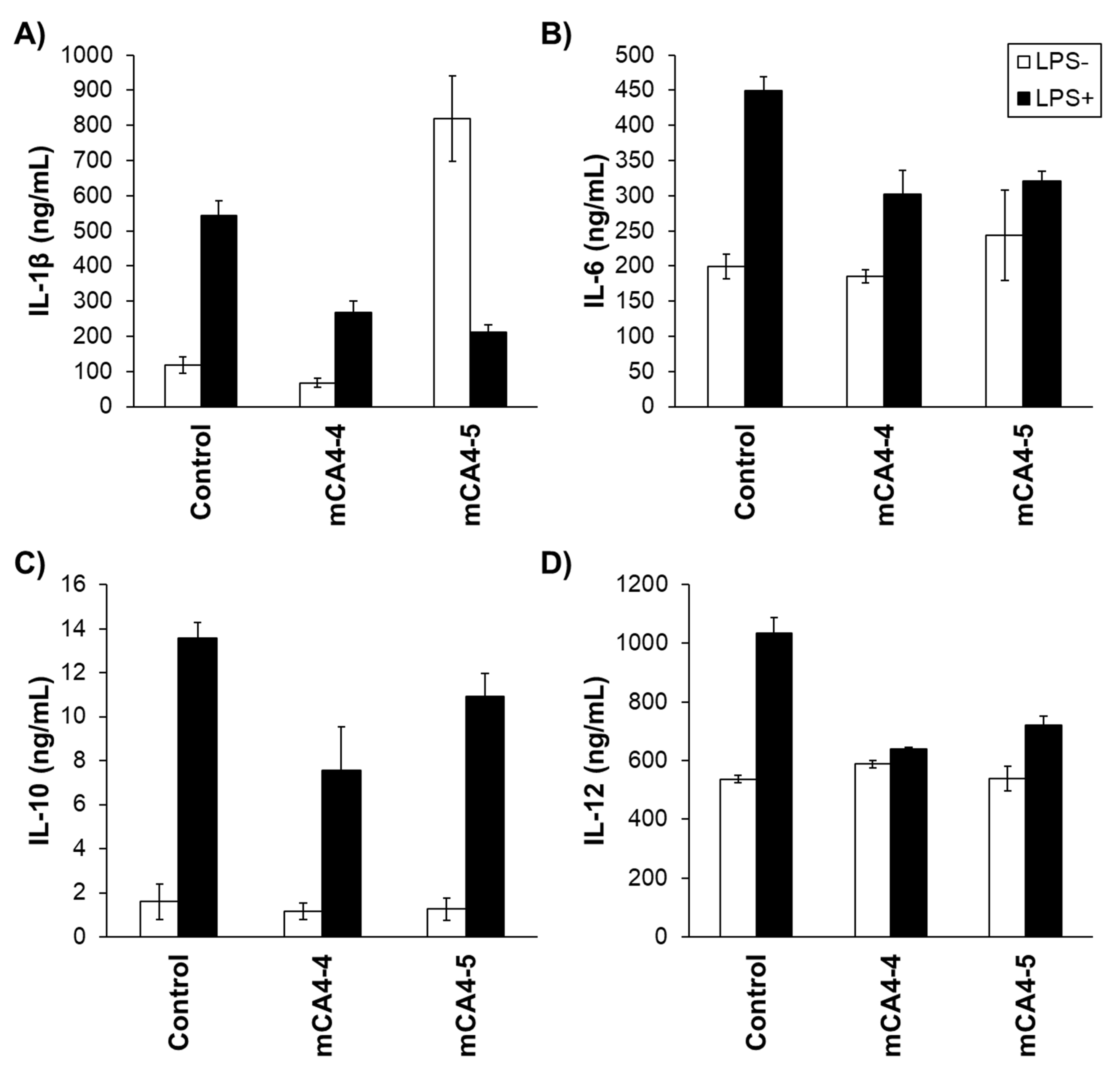
2.8. Neutralization of Bacterial Toxins and Cytokine Release Studies
3. Materials and Methods
3.1. Materials
3.2. Peptide Synthesis and Characterization
3.3. Antibacterial Assay
3.4. Antibacterial Activity of Glutathione-Treated Peptides
3.5. Membrane Permeability Assay
3.6. Flocculation Assay
3.7. Determination of Free Thiol Groups in Peptide Structure
3.8. Bacterial Live/Dead Assay
3.9. Cytotoxicity Assay
3.10. Cellular Uptake Studies
3.11. Bacterial Infection of Caco-2 Cells
3.12. Cytokine Release Studies
3.13. Hemolysis Assay
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Huerta-Cantillo, J.; Navarro-García, F. Properties and design of antimicrobial peptides as potential tools against pathogens and malignant cells. Investig. Discapac. 2016, 5, 96–115. [Google Scholar]
- Mansour, S.C.; Pena, O.M.; Hancock, R.E.W. Host defense peptides: Front-line immunomodulators. Trends Immunol. 2014, 35, 443–450. [Google Scholar] [CrossRef]
- Cole, J.N.; Nizet, V. Bacterial evasion of host antimicrobial peptide defenses. Microbiol. Spectr. 2016, 4, 1. [Google Scholar] [CrossRef]
- Haney, E.F.; Wu, B.; Lee, K.; Hilchie, A.L.; Hancock, R.E.W. Aggregation and its influence on the immunomodulatory activity of synthetic innate defense regulator peptides. Cell Chem. Biol. 2017, 24, 969–980. [Google Scholar] [CrossRef]
- Vaezi, Z.; Bortolotti, A.; Luca, V.; Perilli, G.; Mangoni, M.L.; Khosravi-Far, R.; Bobone, S.; Stella, L. Aggregation determines the selectivity of membrane-active anticancer and antimicrobial peptides: The case of killerFLIP. Biochim. Biophys. Acta (BBA)-Biomembr. 2020, 1862, 183107. [Google Scholar] [CrossRef] [PubMed]
- Wang, C.; Shao, C.; Fang, Y.; Wang, J.; Dong, N.; Shan, A. Binding loop of sunflower trypsin inhibitor 1 serves as a design motif for proteolysis-resistant antimicrobial peptides. Acta Biomater. 2021, 124, 254–269. [Google Scholar] [CrossRef] [PubMed]
- Lee, D.G.; Hahm, K.S.; Shin, S.Y. Structure and fungicidal activity of a synthetic antimicrobial peptide, P18, and its truncated peptides. Biotechnol. Lett. 2004, 26, 337–341. [Google Scholar] [CrossRef] [PubMed]
- Lv, Y.; Wang, J.; Gao, H.; Wang, Z.; Dong, N.; Ma, Q.; Shan, A. Antimicrobial properties and membrane-active mechanism of a potential α-helical antimicrobial derived from cathelicidin PMAP-36. PLoS ONE 2014, 9, e86364. [Google Scholar] [CrossRef]
- Lee, H.; Hwang, J.; Lee, J.; Kim, J.I.; Lee, D.G. Scolopendin 2, a cationic antimicrobial peptide from centipede, and its membrane-active mechanism. Biochim. Biophys. Acta 2015, 1848, 634–642. [Google Scholar] [CrossRef]
- Gopal, R.; Kim, Y.G.; Lee, J.H.; Lee, S.K.; Chae, J.D.; Son, B.K.; Seo, C.H.; Park, Y. Synergistic effects and antibiofilm properties of chimeric peptides against multidrug-resistant Acinetobacter baumannii strains. Antimicrob. Agents Chemother. 2014, 58, 1622–1629. [Google Scholar] [CrossRef]
- Lundy, F.T.; Nelson, J.; Lockhart, D.; Greer, B.; Harriott, P.; Marley, J.J. Antimicrobial activity of truncated α-defensin (human neutrophil peptide (HNP)-1) analogues without disulphide bridges. Mol. Immunol. 2008, 45, 190–193. [Google Scholar] [CrossRef]
- Lin, M.C.; Hui, C.F.; Chen, J.Y.; Wu, J.L. Truncated antimicrobial peptides from marine organisms retain anticancer activity and antibacterial activity against multidrug-resistant Staphylococcus aureus. Peptides 2013, 44, 139–148. [Google Scholar] [CrossRef] [PubMed]
- Kwon, J.Y.; Kim, M.K.; Mereuta, L.; Seo, C.H.; Luchian, T.; Park, Y. Mechanism of action of antimicrobial peptide P5 truncations against Pseudomonas aeruginosa and Staphylococcus aureus. AMB Express 2019, 9, 122. [Google Scholar] [CrossRef] [PubMed]
- Kanthawong, S.; Bolscher, J.G.M.; Veerman, E.C.I.; Marle, J.; de Soet, H.J.J.; Nazmi, K.; Wongratanacheewin, S.; Taweechaisupapong, S. Antimicrobial and antibiofilm activity of LL-37 and its truncated variants against Burkholderia pseudomallei. Int. J. Antimicrob. Agents 2012, 39, 39–44. [Google Scholar] [CrossRef]
- Chen, G.; Miao, Y.; Ma, C.; Zhou, M.; Shi, Z.; Chen, X.; Burrows, J.F.; Xi, X.; Chen, T.; Wang, L. Brevinin-2GHk from Sylvirana guentheri and the design of truncated analogs exhibiting the enhancement of antimicrobial activity. Antibiotics 2020, 9, 85. [Google Scholar] [CrossRef] [PubMed]
- Fan, Y.; Li, X.D.; He, P.P.; Hu, X.X.; Zhang, K.; Fan, J.Q.; Yang, P.P.; Zheng, H.Y.; Tian, W.; Chen, Z.M.; et al. A biomimetic peptide recognizes and traps bacteria in vivo as human defensin-6. Sci. Adv. 2020, 6, eaaz4767. [Google Scholar] [CrossRef] [PubMed]
- Zhang, J.; Liu, S.; Li, H.; Tian, X.; Li, X. Tryptophan-based self-assembling peptides with bacterial flocculation and antimicrobial properties. Langmuir 2020, 36, 11316–11323. [Google Scholar] [CrossRef] [PubMed]
- Rodríguez-Lecompte, J.C.; Yitbarek, A.; Brady, J.; Sharif, S.; Cavanagh, M.D.; Crow, G.; Guenter, W.; House, J.D.; Camelo-Jaimes, G. The effect of microbial-nutrient interaction on the immune system of young chicks after early probiotic and organic acid administration. J. Anim. Sci. 2012, 90, 2246–2254. [Google Scholar] [CrossRef]
- Losada-Medina, D.; Yitbarek, A.; Nazeer, N.; Uribe-Diaz, S.; Ahmed, M.; Rodriguez-Lecompte, J.C. Identification, tissue characterization, and innate immune role of Angiogenin-4 expression in young broiler chickens. Poult. Sci. 2020, 99, 2992–3000. [Google Scholar] [CrossRef]
- Brewer, D.; Lajoie, G. Structure-based design of potent histatin analogues. Biochemistry 2002, 41, 5526–5536. [Google Scholar] [CrossRef]
- Neubauer, D.; Jaskiewicz, M.; Sikorska, E.; Bartoszewska, S.; Bauer, M.; Kapusta, M.; Narajczyk, M.; Kamysz, W. Effect of disulfide cyclization of ultrashort cationic lipopeptides on antimicrobial activity and cytotoxicity. Int. J. Mol. Sci. 2020, 21, 7208. [Google Scholar] [CrossRef]
- Ebenhan, T.; Gheysens, O.; Kruger, H.G.; Zeevaart, J.R.; Sathekge, M.M. Antimicrobial peptides: Their role as infection-selective tracers for molecular imaging. BioMed Res. Int. 2014, 2014, 867381. [Google Scholar] [CrossRef]
- Lázár, V.; Martins, A.; Spohn, R.; Daruka, L.; Grézal, G.; Fekete, G.; Számel, M.; Jangir, P.K.; Kintses, B.; Csörgő, B.; et al. Antibiotic-resistant bacteria show widespread collateral sensitivity to antimicrobial peptides. Nat. Microbiol. 2018, 3, 718–731. [Google Scholar] [CrossRef]
- Dosler, S.; Karaaslan, E. Inhibition and destruction of Pseudomonas aeruginosa biofilms by antibiotics and antimicrobial peptides. Peptides 2014, 62, 32–37. [Google Scholar] [CrossRef] [PubMed]
- Wiradharma, N.; Khan, M.; Yong, L.K.; Hauser, C.A.E.; Seow, S.V.; Zhang, S.; Yang, Y.Y. The effect of thiol functional group incorporation into cationic helical peptides on antimicrobial activities and spectra. Biomaterials 2011, 32, 9100–9108. [Google Scholar] [CrossRef] [PubMed]
- Nekhotiaeva, N.; Elmquist, A.; Rajarao, G.K.; Hallbrink, M.; Langel, Ü.; Good, L. Cell entry and antimicrobial properties of eukaryotic cell-penetrating peptides. FASEB J. 2004, 18, 394–396. [Google Scholar] [CrossRef] [PubMed]
- Xu, L.; Shao, C.; Li, G.; Shan, A.; Chou, S.; Wang, J.; Ma, Q.; Dong, N. Conversion of broad-spectrum antimicrobial peptides into species-specifc antimicrobials capable of precisely targeting pathogenic bacteria. Sci. Rep. 2020, 10, 944. [Google Scholar] [CrossRef]
- Walker, C.R.; Hautefort, I.; Dalton, J.E.; Overweg, K.; Egan, C.E.; Bongaerts, R.J.; Newton, D.J.; Cruickshank, S.M.; Andrew, E.M.; Carding, S.R. Intestinal intraepithelial lymphocyte-enterocyte crosstalk regulates production of bactericidal Angiogenin 4 by Paneth cells upon microbial challenge. PLoS ONE 2013, 8, e84553. [Google Scholar] [CrossRef] [PubMed]
- Lee, S.; Jun, H.; Lee, H.; Chung, C.; Choi, B. Antibacterial and lipopolysaccharide (LPS)-neutralising activity of human cationic antimicrobial peptides against periodontopathogens. Int. J. Antimicrob. Agents 2010, 35, 138–145. [Google Scholar] [CrossRef]
- Schuerholz, T.; Doemming, S.; Hornef, M.; Martin, L.; Simon, T.; Heinbockel, L.; Brandenburg, K.; Marx, G. The anti-inflammatory effect of the synthetic antimicrobial peptide 19-2.5 in a murine sepsis model: A prospective randomized study. Crit. Care 2013, 17, 1–9. [Google Scholar] [CrossRef]
- Popovic, S.; Urbán, E.; Lukic, M.; Conlon, J.M. Peptides with antimicrobial and anti-inflammatory activities that have therapeutic potential for treatment of acne vulgaris. Peptides 2012, 34, 275–282. [Google Scholar] [CrossRef] [PubMed]
- Choi, D.H.; Lee, D.; Jo, B.S.; Park, K.; Lee, K.E.; Choi, J.K.; Park, Y.J.; Lee, J.; Park, Y.S. A synthetic cell-penetrating heparin-binding peptide derived from BMP4 with anti-inflammatory and chondrogenic functions for the treatment of arthritis. Int. J. Mol. Sci. 2020, 21, 4251. [Google Scholar] [CrossRef] [PubMed]
- Liu, Y.; Xia, X.; Xu, L.; Wang, Y. Design of hybrid β-hairpin peptides with enhanced cell specificity and potent anti-inflammatory activity. Biomaterials 2013, 34, 237–250. [Google Scholar] [CrossRef] [PubMed]

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Sample | Peptide Sequences | Theoretical MW (g/mol) | Experimental MW (g/mol) | Net Charge | % ACN Elution |
|---|---|---|---|---|---|
| mCA4-1 | HPSGKKK | 779.5 | 779.5 | +3 | - |
| mCA4-2 | HPSGRRR | 863.5 | 863.5 | +3 | - |
| mCA4-3 | CHPSGKKKC | 985.5 | 985.5 | +3 | - |
| mCA4-4 | CHPSGRRRC | 1069.5 | 1069.5 | +3 | - |
| mCA4-5 | CIVIRFKFRC | 1282.7 | 1281.7 | +3 | 23% |
| Sample Name | Peptide Sequence | IC50 (µM) | MIC (µM) | ||||||
|---|---|---|---|---|---|---|---|---|---|
| E. coli | S. aureus | S. enterica | L. monocytogenes | E. coli | S. aureus | S. enterica | L. monocytogenes | ||
| Ampicillin | – | 15.2 | 3.6 | 7 | 11.2 | 26.9 | 6.7 | 13.5 | 32.3 |
| mCA4-4 | CRRRGSPHC | 68.8 | 52 | 28.2 | >100 | >100 | 100 | 50 | >100 |
| mCA4-5 | CRFKFRIVIC | 15.1 | 4 | 10.2 | 5.2 | 40 | 15 | 30 | 10 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Nazeer, N.; Rodriguez-Lecompte, J.C.; Ahmed, M. Bacterial-Specific Aggregation and Killing of Immunomodulatory Host Defense Peptides. Pharmaceuticals 2021, 14, 839. https://doi.org/10.3390/ph14090839
Nazeer N, Rodriguez-Lecompte JC, Ahmed M. Bacterial-Specific Aggregation and Killing of Immunomodulatory Host Defense Peptides. Pharmaceuticals. 2021; 14(9):839. https://doi.org/10.3390/ph14090839
Chicago/Turabian StyleNazeer, Nauman, Juan Carlos Rodriguez-Lecompte, and Marya Ahmed. 2021. "Bacterial-Specific Aggregation and Killing of Immunomodulatory Host Defense Peptides" Pharmaceuticals 14, no. 9: 839. https://doi.org/10.3390/ph14090839
APA StyleNazeer, N., Rodriguez-Lecompte, J. C., & Ahmed, M. (2021). Bacterial-Specific Aggregation and Killing of Immunomodulatory Host Defense Peptides. Pharmaceuticals, 14(9), 839. https://doi.org/10.3390/ph14090839