
Bacterial Cell Wall Analogue Peptides Control the Oligomeric States and Activity of the Glycopeptide Antibiotic Eremomycin: Solution NMR and Antimicrobial Studies

 , ,
, , 
Abstract
1. Introduction
2. Results
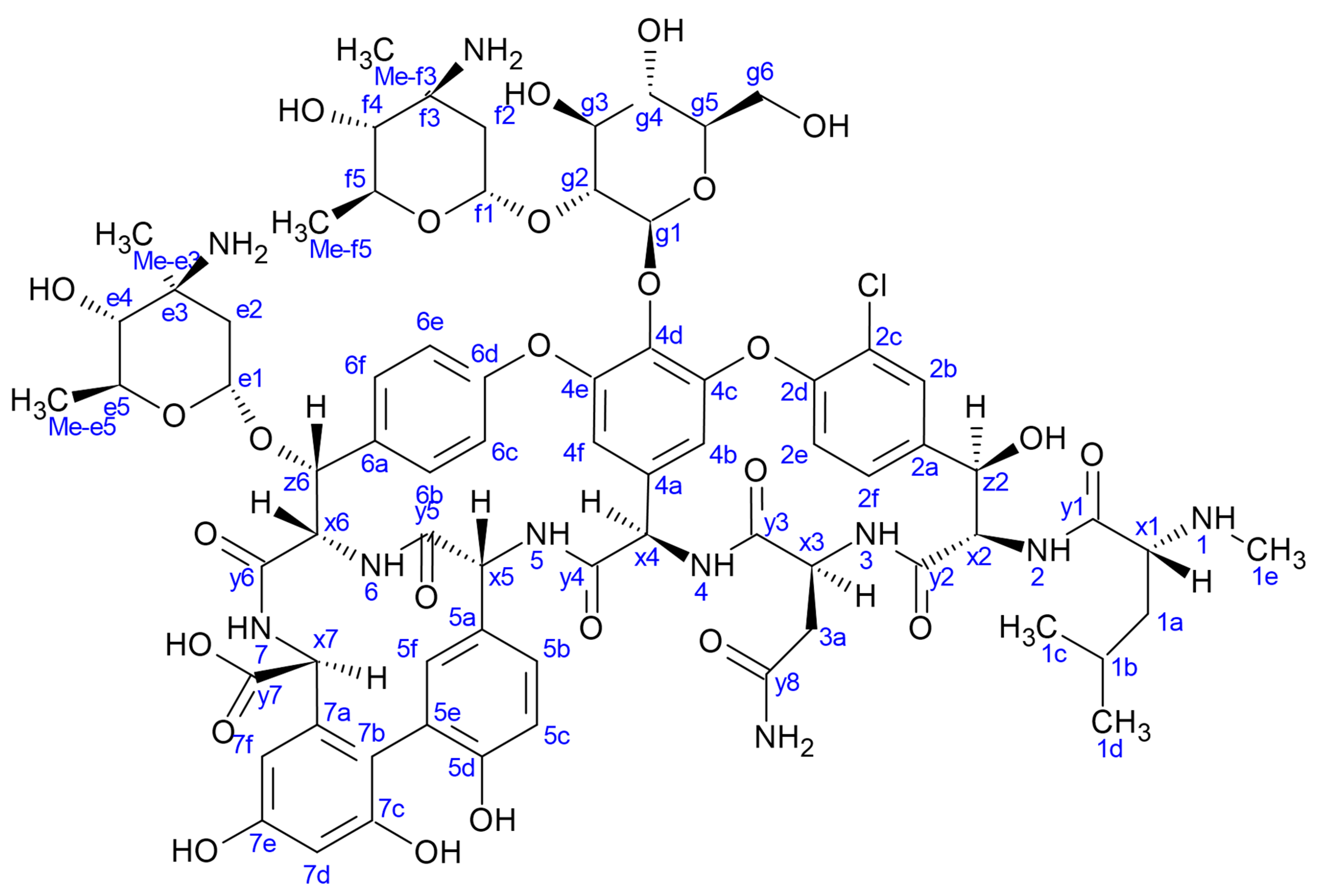
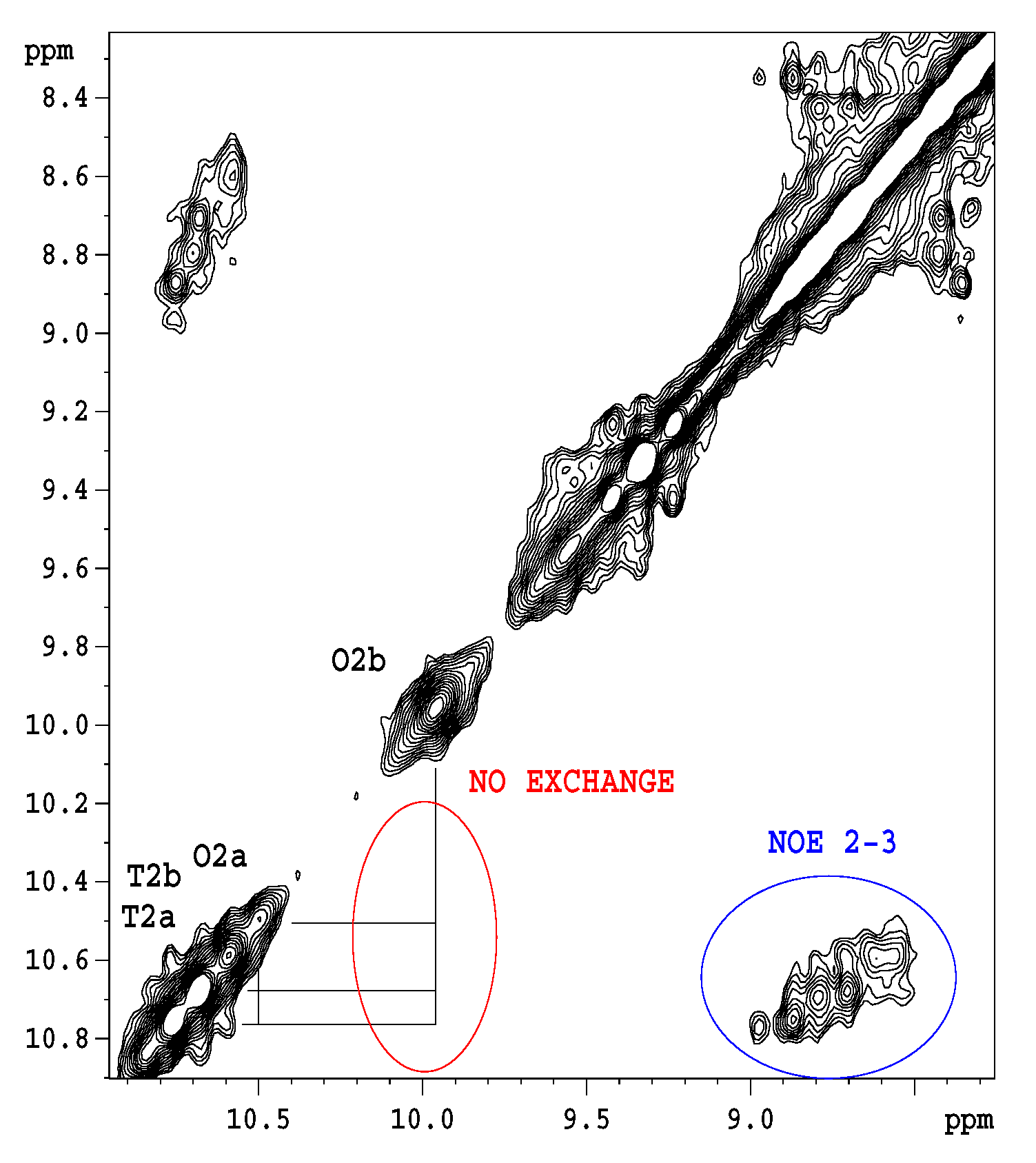
2.1. NMR Relaxation and Diffusion Evidence of Ligand-Dependent Oligomerisation of 15N-Labelled Eremomycin
2.2. Titration of Eremomycin with N-Ac-d-Ala-d-Ala and N-Ac-d-Ala
2.3. Results of Antimicrobial Tests
2.3.1. Checkerboard Method
2.3.2. Disk-Diffusion Method
3. Discussion
4. Materials and Methods
4.1. NMR Spectroscopy
4.2. Antibiotics and Other Reagents
4.3. Bacterial Strains
4.4. Determination of Antibiotic Minimum Inhibitory Concentration
4.5. Interaction of Glycopeptides and the Ligand
4.5.1. Checkerboard Method
4.5.2. Disk-Diffusion Method
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Healy, V.L.; Lessard, I.A.D.; Roper, D.I.; Knox, J.R.; Walsh, C.T. Vancomycin resistance in enterococci: Reprogramming of the D-Ala-D-Ala ligases in bacterial peptidoglycan biosynthesis. Chem. Biol. 2000, 7, R109–R119. [Google Scholar] [CrossRef]
- Reynolds, P.E.; Courvalin, P. Vancomycin resistance in enterococci due to synthesis of precursors terminating in D-Alanyl-D-Serine. Antimicrob. Agents Chemother. 2005, 49, 21–25. [Google Scholar] [CrossRef] [PubMed]
- Zeng, D.; Debabov, D.; Hartsell, T.L.; Cano, R.J.; Adams, S.; Schuyler, J.A.; McMillan, R.; Pace, J.L. Approved Glycopeptide Antibacterial Drugs: Mechanism of Action and Resistance. Cold Spring Harb. Perspect. Med. 2016, 6, a026989. [Google Scholar] [CrossRef] [PubMed]
- Blaskovich, M.A.T.; Hansford, K.A.; Butler, M.S.; Jia, Z.G.; Mark, A.E.; Cooper, M.A. Developments in Glycopeptide Antibiotics. Acs Infect. Dis. 2018, 4, 715–735. [Google Scholar] [CrossRef] [PubMed]
- Loll, P.J.; Axelsen, P.H. The structural biology of molecular recognition by vancomycin. Annu. Rev. Biophys. Biomol. Struct. 2000, 29, 265–289. [Google Scholar] [CrossRef]
- Waltho, J.P.; Williams, D.H. Aspects of Molecular Recognition - Solvent Exclusion and Dimerization of the Antibiotic Ristocetin When Bound to a Model Bacterial Cell-Wall Precursor. J. Am. Chem. Soc. 1989, 111, 2475–2480. [Google Scholar] [CrossRef]
- Batta, G.; Sztaricskai, F.; Kover, K.E.; Rudel, C.; Berdnikova, T.F. An NMR Study of Eremomycin and its Derivatives Full H-1 and C-13 Assignment, Motional Behavior, Dimerization and Complexation with Ac-D-ALA-D-ALA. J. Antibiot. 1991, 44, 1208–1221. [Google Scholar] [CrossRef][Green Version]
- Gerhard, U.; Mackay, J.P.; Maplestone, R.A.; Williams, D.H. The Role of the Sugar and Chlorine Substituents in the Dimerization of Vancomycin Antibiotics. J. Am. Chem. Soc. 1993, 115, 232–237. [Google Scholar] [CrossRef]
- Groves, P.; Searle, M.S.; Mackay, J.P.; Williams, D.H. The Structure of an Asymmetric Dimer Relevant to the Mode of Action of the Glycopeptide Antibiotics. Structure 1994, 2, 747–754. [Google Scholar] [CrossRef]
- Beauregard, D.A.; Williams, D.H.; Gwynn, M.N.; Knowles, D.J.C. Dimerization and Membrane Anchors in Extracellular Targeting of Vancomycin Group Antibiotics. Antimicrob. Agents Chemother. 1995, 39, 781–785. [Google Scholar] [CrossRef]
- Batta, G.; Cristofaro, M.F.; Sharman, G.J.; Williams, D.H. Demonstration of the difference in binding affinity between the two binding sites of the ristocetin A asymmetric dimer. Chem. Commun. 1996, 1, 101–103. [Google Scholar] [CrossRef]
- Bardsley, B.; Williams, D.H. Measurement of the different affinities of the two halves of glycopeptide dimers for acetate. Chem. Commun. 1997, 11, 1049–1050. [Google Scholar] [CrossRef]
- Shiozawa, H.; Chia, B.C.S.; Davies, N.L.; Zerella, R.; Williams, D.H. Cooperative binding interactions of glycopeptide antibiotics. J. Am. Chem. Soc. 2002, 124, 3914–3919. [Google Scholar] [CrossRef] [PubMed]
- McPhail, D.; Cooper, A. Thermodynamics and kinetics of dissociation of ligand-induced dimers of vancomycin antibiotics. J. Chem. Soc.-Faraday Trans. 1997, 93, 2283–2289. [Google Scholar] [CrossRef]
- Linsdell, H.; Toiron, C.; Bruix, M.; Rivas, G.; Menendez, M. Dimerization of A82846B, vancomycin and ristocetin: Influence on antibiotic complexation with cell wall model peptides. J. Antibiot. 1996, 49, 181–193. [Google Scholar] [CrossRef][Green Version]
- Loll, P.J.; Miller, R.; Weeks, C.M.; Axelsen, P.H. A ligand-mediated dimerization mode for vancomycin. Chem. Biol. 1998, 5, 293–298. [Google Scholar] [CrossRef]
- Schafer, M.; Sheldrick, G.M.; Schneider, T.R.; Vertesy, L. Structure of balhimycin and its complex with solvent molecules. Acta Crystallogr. Sect. D Biol. Crystallogr. 1998, 54, 175–183. [Google Scholar] [CrossRef]
- Loll, P.J.; Derhovanessian, A.; Shapovalov, M.V.; Kaplan, J.; Yang, L.; Axelsen, P.H. Vancomycin Forms Ligand-Mediated Supramolecular Complexes. J. Mol. Biol. 2009, 385, 200–211. [Google Scholar] [CrossRef]
- Jia, Z.G.; O’Mara, M.L.; Zuegg, J.; Cooper, M.A.; Mark, A.E. Vancomycin: Ligand recognition, dimerization and super-complex formation. FEBS J. 2013, 280, 1294–1307. [Google Scholar] [CrossRef]
- Loll, P.J.; Bevivino, A.E.; Korty, B.D.; Axelsen, P.H. Simultaneous recognition of a carboxylate-containing ligand and an intramolecular surrogate ligand in the crystal structure of an asymmetric vancomycin dimer. J. Am. Chem. Soc. 1997, 119, 1516–1522. [Google Scholar] [CrossRef]
- Lehmann, C.; Bunkoczi, G.; Vertesy, L.; Sheldrick, G.M. Structures of glycopeptide antibiotics with peptides that model bacterial cell-wall precursors. J. Mol. Biol. 2002, 318, 723–732. [Google Scholar] [CrossRef]
- Cooper, M.A.; Williams, D.H. Binding of glycopeptide antibiotics to a model of a vancomycin-resistant bacterium. Chem. Biol. 1999, 6, 891–899. [Google Scholar] [CrossRef]
- Cooper, M.A.; Williams, D.H.; Cho, Y.R. Surface plasmon resonance analysis of glycopeptide antibiotic activity at a model membrane surface. Chem. Commun. 1997, 17, 1625–1626. [Google Scholar] [CrossRef]
- Good, V.M.; Gwynn, M.N.; Knowles, D.J.C. MM-45289, A Potent Glycopeptide Antibiotic which Interacts Weakly with Diacetyl-L-Lysyl-D-Alanyl-D-Alanine. J. Antibiot. 1990, 43, 550–555. [Google Scholar] [CrossRef] [PubMed]
- Beauregard, D.A.; Maguire, A.J.; Williams, D.H.; Reynolds, P.E. Semiquantitation of cooperativity in binding of vancomycin-group antibiotics to vancomycin-susceptible and -resistant organisms. Antimicrob. Agents Chemother. 1997, 41, 2418–2423. [Google Scholar] [CrossRef][Green Version]
- Mackay, J.P.; Gerhard, U.; Beauregard, D.A.; Westwell, M.S.; Searle, M.S.; Williams, D.H. Glycopeptide Antibiotic-Activity and the Possible Role of Dimerization—A Model for Biological Signaling. J. Am. Chem. Soc. 1994, 116, 4581–4590. [Google Scholar] [CrossRef]
- Batta, G.; Sztaricskai, F.; Makarova, M.O.; Gladkikh, E.G.; Pogozheva, V.V.; Berdnikova, T.F. Backbone dynamics and amide proton exchange at the two sides of the eremomycin dimer by N-15 NMR. Chem. Commun. 2001, 5, 501–502. [Google Scholar] [CrossRef]
- Allen, N.E.; Nicas, T.I. Mechanism of action of oritavancin and related glycopeptide antibiotics. FEMS Microbiol. Rev. 2003, 26, 511–532. [Google Scholar] [CrossRef]
- Farrow, N.A.; Muhandiram, R.; Singer, A.U.; Pascal, S.M.; Kay, C.M.; Gish, G.; Shoelson, S.E.; Pawson, T.; Formankay, J.D.; Kay, L.E. Backbone Dynamics of a Free and a Phosphopeptide -Complexed SRC Homology-2 Domain Studied by N-15 NMR Relaxation. Biochemistry 1994, 33, 5984–6003. [Google Scholar] [CrossRef]
- Lipari, G.; Szabo, A. Model-Free Approach to the Interpretation of Nuclear Magnetic-Resonance Relaxation in Macromolecules. 1. Theory and Range of Validity. J. Am. Chem. Soc. 1982, 104, 4546–4559. [Google Scholar] [CrossRef]
- Lipari, G.; Szabo, A. Model-Free Approach to the Interpretation of Nuclear Magnetic-Resonance Relaxation in Macromolecules. 2. Analysis of Experimental Results. J. Am. Chem. Soc. 1982, 104, 4559–4570. [Google Scholar] [CrossRef]
- Batta, G.; Kover, K.E.; Szekely, Z.; Sztaricskai, F. Glycopeptide Binding-Site Spied through Transferred Heteronuclear NOE - 1-C-13 Ac-D-Ala-D-Ala Bonded to Vancomycin and Ristocetin-A. J. Am. Chem. Soc. 1992, 114, 2757–2758. [Google Scholar] [CrossRef]
- Sharman, G.J.; Searle, M.S.; Benhamu, B.; Groves, P.; Williams, D.H. Burial of Hydrocarbon Causes Cooperative Enhancement of Electrostatic Binding. Angew. Chem. Int. Ed. Eng. 1995, 34, 1483–1485. [Google Scholar] [CrossRef]
- Phillips-Jones, M.K.; Lithgo, R.; Dinu, V.; Gillis, R.B.; Harding, J.E.; Adams, G.G.; Harding, S.E. Full hydrodynamic reversibility of the weak dimerization of vancomycin and elucidation of its interaction with VanS monomers at clinical concentration. Sci. Rep. 2017, 7, 12697. [Google Scholar] [CrossRef] [PubMed]
- Orosz, L.; Batta, G.; Keki, S.; Nagy, M.; Deak, G.; Zsuga, M. Self-association of bis-(alpha,beta-D-glucopyranosyl)-polyisobutylene. Carbohydr. Res. 2007, 342, 1323–1328. [Google Scholar] [CrossRef] [PubMed]
- Nicolaou, K.C.; Boddy, C.N.C.; Brase, S.; Winssinger, N. Chemistry, biology, and medicine of the glycopeptide antibiotics. Angew. Chem.-Int. Ed. 1999, 38, 2096–2152. [Google Scholar] [CrossRef]
- Iadanza, M.G.; Jackson, M.P.; Hewitt, E.W.; Ranson, N.A.; Radford, S.E. A new era for understanding amyloid structures and disease. Nat. Rev. Mol. Cell Biol. 2018, 19, 755–773. [Google Scholar] [CrossRef] [PubMed]
- Szűcs, Z.; Naesens, L.; Stevaert, A.; Ostorházi, E.; Batta, G.; Herczegh, P.; Borbás, A. Reprogramming of the Antibacterial Drug Vancomycin Results in Potent Antiviral Agents Devoid of Antibacterial Activity. Pharmaceuticals 2020, 13, 139. [Google Scholar] [CrossRef]














{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Microorganisms | Glycopeptide | Concentration of Ligand (μM/mL) | ||||||||
|---|---|---|---|---|---|---|---|---|---|---|
| 0 | 8 | 16 | 32 | 64 | 128 | 256 | 512 | 1024 | ||
| Glycopeptide MIC (μM/mL) | ||||||||||
| S. aureus 20450 | vancomycin | 1 | 2 | 2 | 2 | 2 | 2 | 2 | 4 | 8 |
| eremomycin | 0.12 | 0.06 | 0.06 | 0.06 | 0.06 | 0.125 | 0.125 | 0.125 | 0.5 | |
| E. faecalis 9 | vancomycin | 16 | 16 | 16 | 16 | 16 | 16 | 32 | 32 | 64 |
| eremomycin | 0.25 | 0.12 | 0.25 | 0.25 | 0.25 | 0.25 | 0.25 | 0.5 | 1 | |
| S. aureus 209P | vancomycin | 1 | 1 | 1 | 1 | 1 | 1 | 2 | 2 | 4 |
| eremomycin | 0.06 | 0.03 | 0.03 | 0.03 | 0.03 | 0.03 | 0.03 | 0.06 | 0.125 | |
| N-Ac-d-Ala-d-Ala (μg/mL) | 0 | 10 | 100 | ||||||
|---|---|---|---|---|---|---|---|---|---|
| Glycopeptide Concentration (μg/mL) | 5 | 10 | 20 | 5 | 10 | 20 | 5 | 10 | 20 |
| Vancomycin | diffusion zones (mm) | ||||||||
| 14.5 | 16.3 | 18.1 | 11.6 | 13 | 14.5 | 8 | 10.2 | 12 | |
| relative potency | |||||||||
| - | −25% | −25% | −25% | −80% | −60% | −20% | |||
| Eremomycin | diffusion zones (mm) | ||||||||
| 15.8 | 17.6 | 19.5 | 17.7 | 19.4 | 21.5 | 18.2 | 21.1 | 21.5 | |
| relative potency | |||||||||
| - | +12% | +10% | +10% | +15% | +20% | +10% | |||
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Izsépi, L.; Erdei, R.; Tevyashova, A.N.; Grammatikova, N.E.; Shchekotikhin, A.E.; Herczegh, P.; Batta, G. Bacterial Cell Wall Analogue Peptides Control the Oligomeric States and Activity of the Glycopeptide Antibiotic Eremomycin: Solution NMR and Antimicrobial Studies. Pharmaceuticals 2021, 14, 83. https://doi.org/10.3390/ph14020083
Izsépi L, Erdei R, Tevyashova AN, Grammatikova NE, Shchekotikhin AE, Herczegh P, Batta G. Bacterial Cell Wall Analogue Peptides Control the Oligomeric States and Activity of the Glycopeptide Antibiotic Eremomycin: Solution NMR and Antimicrobial Studies. Pharmaceuticals. 2021; 14(2):83. https://doi.org/10.3390/ph14020083
Chicago/Turabian StyleIzsépi, László, Réka Erdei, Anna N. Tevyashova, Natalia E. Grammatikova, Andrey E. Shchekotikhin, Pál Herczegh, and Gyula Batta. 2021. "Bacterial Cell Wall Analogue Peptides Control the Oligomeric States and Activity of the Glycopeptide Antibiotic Eremomycin: Solution NMR and Antimicrobial Studies" Pharmaceuticals 14, no. 2: 83. https://doi.org/10.3390/ph14020083
APA StyleIzsépi, L., Erdei, R., Tevyashova, A. N., Grammatikova, N. E., Shchekotikhin, A. E., Herczegh, P., & Batta, G. (2021). Bacterial Cell Wall Analogue Peptides Control the Oligomeric States and Activity of the Glycopeptide Antibiotic Eremomycin: Solution NMR and Antimicrobial Studies. Pharmaceuticals, 14(2), 83. https://doi.org/10.3390/ph14020083