
Flavonoid Composition and Pharmacological Properties of Elaeis guineensis Jacq. Leaf Extracts: A Systematic Review

 ,
, 
 ,
,  and
and 
Abstract
1. Introduction
2. Methods
2.1. Search Strategy
2.2. Article Eligibility Criteria
3. Results
3.1. Secondary Metabolites in the Leaves of E. guineensis
3.2. Classification of Flavonoids Found in the Leave of E. guineensis
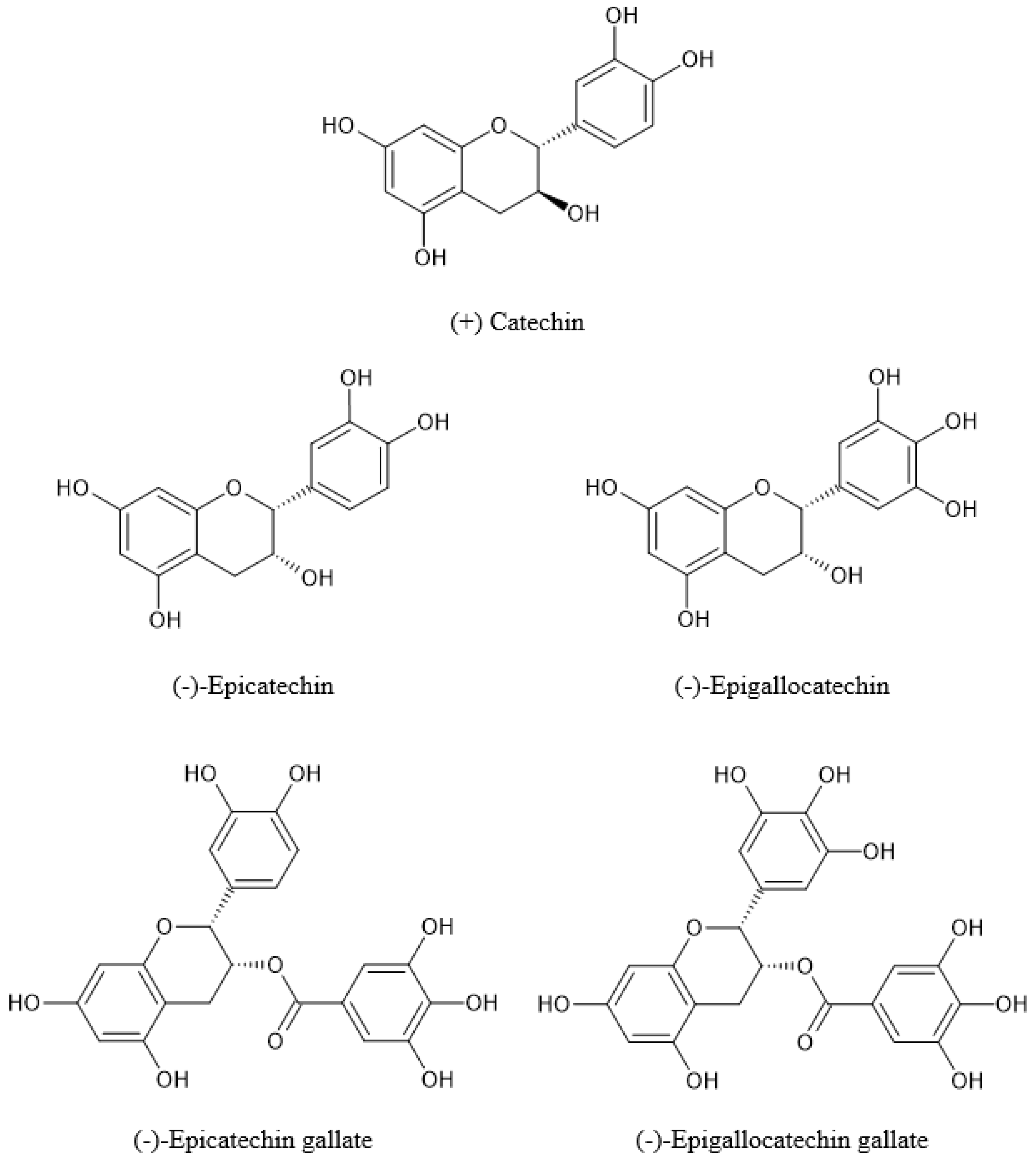
3.2.1. Flavan-3-ols
3.2.2. Flavones
Flavone-C-glycosides
3.3. Total Phenolic Content of E. guineensis Leaf Extract
3.4. Total Flavonoid Content of E. guineensis Leaf Extract
3.5. Pharmacological Activities of E. guineensis Leaf Extracts
3.5.1. Antioxidant Activity
In Vitro Antioxidant Activity
- DPPH Free Radical Scavenging Activity
- Ferric Reducing/Antioxidant Power
- Hydrogen Peroxide Scavenging Activity
- Lipid Peroxidation Activity
- Nitric Oxide Scavenging Activity
- In Vitro LDL Oxidation
- Xanthine Oxidase Inhibitory Activity
- Beta-Carotene–Linoleic Acid Bleaching Assay
- In Vivo Antioxidant Activity
3.5.2. Hypoglycemic Activity
3.5.3. Wound-Healing Activity
3.5.4. Vascular Relaxation
3.5.5. Hypocholesterolemic Activity
3.5.6. Neuroprotective/Neurogenesis
3.5.7. Phytoestrogenic Properties/Osteogenic Activity
3.5.8. Fungicidal/Antimicrobial Activity
3.5.9. Antihypertensive Properties and Cardiovascular Effects
3.5.10. Cytotoxicity/Toxicity Effects
3.6. Strengths and Limitations
3.7. Future Research
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Abbreviations and Acronyms
| AD | Alzheimer’s disease |
| ALT | Alanine aminotransferase |
| AST | Aspartate amino transferase |
| AAE | Ascorbic acid equivalents |
| BHT | Butylated hydroxytoluene |
| BP | Blood pressure |
| CAT | Catalase |
| CE | (+)-Catechin equivalents |
| CFU | Colony-forming unit |
| CNS | Central nervous system |
| DPPH | 2,2-diphenyl-1-picrylhydrazyl |
| EA | Ethyl acetate fraction |
| EC50 | Half maximal effective concentration |
| EC | Epicatechin |
| ECE | (+)-Epicatechin equivalents |
| ECG | Epicatechin gallate |
| EFB | Empty fruit bunches |
| EGC | Epigallocatechin |
| EGCG | Epigallocatechin gallate |
| FAE | Ferulic acid equivalents |
| FRAP | Ferric reducing/antioxidant power |
| FTIR | Fourier transform infrared spectroscopy |
| GAE | Gallic acid equivalents |
| GGT | Gamma-glutamyl transferase |
| GPx | Glutathione peroxidase activity |
| HDL | High density lipoprotein |
| HDL-C | High-density lipoprotein cholesterol |
| HEX | Hexane fraction |
| 1H-NMR | Proton nuclear magnetic resonance |
| HPLC | High-performance liquid chromatography |
| HPSA | Hydrogen peroxide scavenging activity |
| IC50 | Half-maximal inhibitory concentration |
| INSOL | Insoluble fraction |
| LA | Linoleic acid |
| LC-MS | Liquid chromatography–mass spectrometry |
| LD50 | Median lethal dose |
| LDL | Low-density lipoprotein |
| LDLr | Low-density lipoprotein receptor |
| L-NAME | Nω-nitro-L-arginine methyl ester |
| LPO | Lipid peroxidation |
| MTT | 3-(4,5-Dimethylthiazol-2-yl)-2,5-Diphenyl tetrazolium bromide |
| MDA | Malonyldialdehyde |
| MeOH | Methanolic extract |
| MF | Mesocarp fibre |
| MIC | Minimum inhibitory concentration |
| MMPs | Matrix metalloproteinases |
| NO | Nitric oxide |
| NOLA | Nω-nitro-L-arginine |
| NOS | Nitric oxide synthase |
| NOS | Nitric oxide scavenging |
| N/R | Not reported |
| OPB | Oil palm biomass |
| OPAL | Oil palm leaves alcoholic extract |
| OPL | Oil palm leaves |
| OPLE | Oil palm leaf extract |
| OPT | Oil palm trunk |
| OVX | Ovariectomized |
| PKS | Palm kernel shell |
| POME | Palm oil mill effluent |
| PRISMA | Preferred Reporting Items for Systematic reviews and Meta-Analyses |
| PUFA | Polyunsaturated fatty acid |
| QCE | Quercetin equivalents |
| ROS | Reactive oxygen species |
| SOD | Superoxide dismutase |
| SAR | Structure activity relationship |
| STZ | Streptozotocin |
| TCK | Creatine kinase |
| TBARS | Conjugated diene formation and thiobarbituric acid reactive substance |
| TPC | Total phenolic content |
| TPTZ | Tripyridyltriazine |
| UHPLC-UV/PDA | Ultrahigh performance liquid chromatography-ultraviolet/photodiode array |
| TFC | Total flavonoid content |
| TBARS | Conjugated diene formation and thiobarbituric acid reactive substance |
References
- Awalludin, M.F.; Sulaiman, O.; Hashim, R.; Nadhari, W.N.A.W. An overview of the oil palm industry in Malaysia and its waste utilization through thermochemical conversion, specifically via liquefaction. Renew. Sustain. Energy Rev. 2015, 50, 1469–1484. [Google Scholar] [CrossRef]
- Hui, A.C.; Foon, C.S.; Hock, C.C. Antioxidant activities of Elaeis guineensis leaves. J. Oil Palm Res. 2017, 29, 343–351. [Google Scholar] [CrossRef][Green Version]
- Owoyele, B.; Owolabi, G. Traditional oil palm (Elaeis guineensis jacq.) and its medicinal uses: A review. TANG 2014, 4, 16.1–16.8. [Google Scholar] [CrossRef]
- Soundararajan, V.; Sreenivasan, S. Antioxidant Activity of Elaeis guineensis Leaf Extract: An Alternative Nutraceutical Approach in Impeding Aging. APCBEE Proc. 2012, 2, 153–159. [Google Scholar] [CrossRef]
- Sreenivasan, S.; Rajoo, N.; Rathinam, X.; Lachimanan, Y.L.; Rajoo, A. Wound healing potential of Elaeis guineensis Jacq leaves in an infected albino rat model. Molecules 2010, 15, 3186–3199. [Google Scholar] [CrossRef]
- Abdullah, S.; Chong, K.; Ng, S. Phytochemical Constituents From Leaves of Elaies Guineensis and Their Antioxidants and Antimicrobial Activities. Int. J. Pharm. Pharm. Sci. 2013, 5, 137–140. [Google Scholar]
- Ooi, Z.X.; Teoh, Y.P.; Kunasundari, B.; Shuit, S.H. Oil palm frond as a sustainable and promising biomass source in Malaysia: A review. Environ. Prog. Sustain. Energy 2017, 36, 1864–1874. [Google Scholar] [CrossRef]
- Tahir, N.I.; Shaari, K.; Abas, F.; Parveez, G.K.; Ishak, Z.; Ramli, U.S. Characterization of apigenin and luteolin derivatives from oil palm (Elaeis guineensis Jacq.) leaf using LC-ESI-MS/MS. J. Agric. Food Chem. 2012, 60, 11201–11210. [Google Scholar] [CrossRef]
- Ofori-Boateng, C.; Lee, K.T. Sustainable utilization of oil palm wastes for bioactive phytochemicals for the benefit of the oil palm and nutraceutical industries. Phytochem. Rev. 2013, 12, 173–190. [Google Scholar] [CrossRef]
- Onoja, E.; Chandren, S.; Razak, F.; Mahat, N.; Wahab, R. Oil Palm (Elaeis guineensis) Biomass in Malaysia: The Present and Future Prospects. Waste Biomass Valorization 2019, 10. [Google Scholar] [CrossRef]
- Ahmad, N.; Hasan, Z.A.A.; Muhamad, H.; Bilal, S.H.; Yusof, N.Z.; Idris, Z. Determination of total phenol, flavonoid, antioxidant activity of oil palm leaves extracts and their application in transparent soap. J. Oil Palm Res. 2018, 30, 315–325. [Google Scholar] [CrossRef]
- Yusof, N.; Abd Gani, S.S.; Siddiqui, Y.; Mokhtar, N.O.R.; Aldrin, Z. Potential uses of oil palm (Elaeis guineensis) leaf extract in topical application. J. Oil Palm Res. 2016, 28. [Google Scholar] [CrossRef]
- Mohamed, S. Oil Palm Leaf: A New Functional Food Ingredient for Health and Disease Prevention. J. Food Process. Technol. 2014, 5, 1–6. [Google Scholar] [CrossRef]
- Bakhsh, A.; Mustapha, N.M.; Mohamed, S. Catechin-rich oil palm leaf extract enhances bone calcium content of estrogen-deficient rats. Nutrition 2013, 29, 667–672. [Google Scholar] [CrossRef] [PubMed]
- Liberati, A.; Altman, D.G.; Tetzlaff, J.; Mulrow, C.; Gøtzsche, P.C.; Ioannidis, J.P.A.; Clarke, M.; Devereaux, P.J.; Kleijnen, J.; Moher, D. The PRISMA statement for reporting systematic reviews and meta-analyses of studies that evaluate healthcare interventions: Explanation and elaboration. BMJ 2009, 339, b2700. [Google Scholar] [CrossRef] [PubMed]
- Methley, A.M.; Campbell, S.; Chew-Graham, C.; McNally, R.; Cheraghi-Sohi, S. PICO, PICOS and SPIDER: A comparison study of specificity and sensitivity in three search tools for qualitative systematic reviews. BMC Health Serv. Res. 2014, 14, 579. [Google Scholar] [CrossRef] [PubMed]
- Louisiana State University. Systematic Reviews: Inclusion and Exclusion Criteria. Available online: https://guides.lib.lsu.edu/c.php?g=872965&p=7866853 (accessed on 28 February 2021).
- Irine, R.; Noordin, M.M.; Radzali, M.; Azizah, H.; Hapizah, N.; Mahinda, Y.A.; Suhaila, M. Antioxidant and hypocholesterolemic effects of Elaeis guineensis frond extract on hypercholesterolemic rabbits. ASEAN Food J. 2003, 12, 137–147. [Google Scholar]
- Mohamed, S.; Lee Ming, T.; Jaffri, J.M. Cognitive enhancement and neuroprotection by catechin-rich oil palm leaf extract supplement. J. Sci. Food Agric. 2013, 93, 819–827. [Google Scholar] [CrossRef]
- Han, N.M.; May, C.Y. Determination of antioxidants in oil palm leaves (Elaeis guineensis). Am. J. Appl. Sci. 2010, 7, 1243–1247. [Google Scholar] [CrossRef]
- Che, Z.M.S.; Lee, S.Y.; Shaari, K.; Sarian, M.N.; Fakurazi, S.; Sarian, M.N. In Vitro Wound Healing Potential of Flavonoid C-Glycosides from Oil Palm (Elaeis guineensis Jacq.) Leaves on 3T3 Fibroblast Cells. Antioxidants 2020, 9, 326. [Google Scholar]
- Che, Z.M.S.; Lee, S.Y.; Shaari, K.; Nasir, N.M.; Shaari, K.; Fakurazi, S. Metabolite Characterization and Correlations with Antioxidant and Wound Healing Properties of Oil Palm (Elaeis guineensis Jacq.) Leaflets via (1)H-NMR-Based Metabolomics Approach. Molecules 2020, 25, 5636. [Google Scholar]
- Ibraheem, Z.; Zubaid, M.; Abdullah, N.; Rathore, H.; Johns, E. Toxicity, phytochemical content and antioxidant activity assessment studies for a standardized ethanolic fraction of palm oil leaf extract. Pharmacogn. Commun. 2012, 2, 21–30. [Google Scholar] [CrossRef]
- Guhathakurta, S.; Anandhan, S.; Singha, N.K.; Chattopadhyay, R.N.; Bhowmick, A.K. Waste natural gum as a multifunctional additive in rubber. J. Appl. Polym. Sci. 2006, 102, 4897–4907. [Google Scholar] [CrossRef]
- Kopustinskiene, D.M.; Jakstas, V.; Savickas, A.; Bernatoniene, J. Flavonoids as Anticancer Agents. Nutrients 2020, 12, 457. [Google Scholar] [CrossRef] [PubMed]
- Ghorbani, A.; Rashidi, R.; Shafiee-Nick, R. Flavonoids for preserving pancreatic beta cell survival and function: A mechanistic review. Biomed. Pharmacother. 2019, 111, 947–957. [Google Scholar] [CrossRef] [PubMed]
- Al-Ishaq, R.K.; Abotaleb, M.; Kubatka, P.; Kajo, K.; Büsselberg, D. Flavonoids and Their Anti-Diabetic Effects: Cellular Mechanisms and Effects to Improve Blood Sugar Levels. Biomolecules 2019, 9, 430. [Google Scholar] [CrossRef] [PubMed]
- Visioli, F.; Panaite, S.-A.; Tomé-Carneiro, J. Wine’s Phenolic Compounds and Health: A Pythagorean View. Molecules 2020, 25, 4105. [Google Scholar] [CrossRef] [PubMed]
- Bakhtiari, M.; Panahi, Y.; Ameli, J.; Darvishi, B. Protective effects of flavonoids against Alzheimer’s disease-related neural dysfunctions. Biomed. Pharm. 2017, 93, 218–229. [Google Scholar] [CrossRef] [PubMed]
- Yang, B.; Liu, H.; Yang, J.; Gupta, V.K.; Jiang, Y. New insights on bioactivities and biosynthesis of flavonoid glycosides. Trends Food Sci. Technol. 2018, 79, 116–124. [Google Scholar] [CrossRef]
- Jaffri, J.M.; Mohamed, S.; Rohimi, N.; Ahmad, I.N.; Noordin, M.M.; Manap, Y.A. Antihypertensive and cardiovascular effects of catechin-rich oil palm (Elaeis guineensis) leaf extract in nitric oxide-deficient rats. J. Med. Food 2011, 14, 775–783. [Google Scholar] [CrossRef]
- Isemura, M. Catechin in Human Health and Disease. Molecules 2019, 24, 528. [Google Scholar] [CrossRef]
- Yu, Y.; Deng, Y.; Lu, B.M.; Liu, Y.X.; Li, J.; Bao, J.K. Green tea catechins: A fresh flavor to anticancer therapy. Apoptosis 2014, 19, 1–18. [Google Scholar] [CrossRef]
- Aziz, N.; Kim, M.Y.; Cho, J.Y. Anti-inflammatory effects of luteolin: A review of in vitro, in vivo, and in silico studies. J. Ethnopharmacol. 2018, 225, 342–358. [Google Scholar] [CrossRef]
- Abdulai, I.L.; Kwofie, S.K.; Gbewonyo, W.S.; Boison, D.; Puplampu, J.B.; Adinortey, M.B. Multitargeted Effects of Vitexin and Isovitexin on Diabetes Mellitus and Its Complications. Sci. World J. 2021, 2021, 6641128. [Google Scholar] [CrossRef] [PubMed]
- Rauter, A.P.; Lopes, R.G.; Martins, A. C-Glycosylflavonoids: Identification, Bioactivity and Synthesis. Nat. Prod. Commun. 2007, 2, 1934578X0700201125. [Google Scholar] [CrossRef]
- Waterhouse, A.L. Determination of Total Phenolics. Curr. Protoc. Food Anal. Chem. 2002, 6, I1.1.1–I1.1.8. [Google Scholar] [CrossRef]
- Kabtni, S.; Sdouga, D.; Bettaib Rebey, I.; Save, M.; Trifi-Farah, N.; Fauconnier, M.-L.; Marghali, S. Influence of climate variation on phenolic composition and antioxidant capacity of Medicago minima populations. Sci. Rep. 2020, 10, 8293. [Google Scholar] [CrossRef] [PubMed]
- Ghafoor, K.; Mohamed Ahmed, I.; Doğu, S.; Uslu, N.; Fadimu, G.; Al Juhaimi, F.; Babiker, E.; Özcan, M. The Effect of Heating Temperature on Total Phenolic Content, Antioxidant Activity, and Phenolic Compounds of Plum and Mahaleb Fruits. Int. J. Food Eng. 2019, 15. [Google Scholar] [CrossRef]
- Chandra, S.; Khan, S.; Avula, B.; Lata, H.; Yang, M.H.; ElSohly, M.A.; Khan, I.A. Assessment of Total Phenolic and Flavonoid Content, Antioxidant Properties, and Yield of Aeroponically and Conventionally Grown Leafy Vegetables and Fruit Crops: A Comparative Study. Evid. -Based Complementary Altern. Med. 2014, 2014, 253875. [Google Scholar] [CrossRef]
- Iwansyah, A.; Manh, T.; Andriana, Y.; Hessan, M.; Kormin, F.; Dang, X.; Cuong, D.; Xuan Hoan, N.; Thai Ha, H.; Dang, T.; et al. Effects of Various Drying Methods on Selected Physical and Antioxidant Properties of Extracts from Moringa oliefera Leaf Waste. Sustainability 2020, 12, 8586. [Google Scholar] [CrossRef]
- Yin, Q.; Mu, H.; Zeng, M.; Gao, D.; Qin, F.; Chen, J.; He, Z. Effects of heating on the total phenolic content, antioxidant activities and main functional components of simulated Chinese herb candy during boiling process. J. Food Meas. Charact. 2019, 13, 476–486. [Google Scholar] [CrossRef]
- Maghsoudlou, Y.; Asghari Ghajari, M.; Tavasoli, S. Effects of heat treatment on the phenolic compounds and antioxidant capacity of quince fruit and its tisane’s sensory properties. J. Food Sci. Technol. 2019, 56, 2365–2372. [Google Scholar] [CrossRef] [PubMed]
- Abeywardena, M.; Runnie, I.; Nizar, M.; Momamed, S.; Head, R. Polyphenol-enriched extract of oil palm fronds (Elaeis guineensis) promotes vascular relaxation via endothelium-dependent mechanisms. Asia Pac. J. Clin. Nutr. 2002, 11, S467–S472. [Google Scholar] [CrossRef]
- Salleh, M.N.; Runnie, I.; Roach, P.D.; Mohamed, S.; Abeywardena, M.Y. Inhibition of Low-Density Lipoprotein Oxidation and Up-Regulation of Low-Density Lipoprotein Receptor in HepG2 Cells by Tropical Plant Extracts. J. Agric. Food Chem. 2002, 50, 3693–3697. [Google Scholar] [CrossRef] [PubMed]
- Rosalina Tan, R.T.; Mohamed, S.; Samaneh, G.F.; Noordin, M.M.; Goh, Y.M.; Manap, M.Y.A. Polyphenol rich oil palm leaves extract reduce hyperglycaemia and lipid oxidation in STZ-rats. Int. Food Res. J. 2011, 18, 179–187. [Google Scholar]
- Sasidharan, S.; Logeswaran, S.; Latha, L.Y. Wound healing activity of Elaeis guineensis leaf extract ointment. Int. J. Mol. Sci. 2012, 13, 336–347. [Google Scholar] [CrossRef] [PubMed]
- Sikarwar, M.S.; Patil, M.B. Antidiabetic activity of Pongamia pinnata leaf extracts in alloxan-induced diabetic rats. Int. J. Ayurveda Res. 2010, 1, 199–204. [Google Scholar] [CrossRef]
- Schwartz, B.G.; Economides, C.; Mayeda, G.S.; Burstein, S.; Kloner, R.A. The endothelial cell in health and disease: Its function, dysfunction, measurement and therapy. Int. J. Impot. Res. 2010, 22, 77–90. [Google Scholar] [CrossRef]
- Ajay, M.; Gilani, A.U.; Mustafa, M.R. Effects of flavonoids on vascular smooth muscle of the isolated rat thoracic aorta. Life Sci. 2003, 74, 603–612. [Google Scholar] [CrossRef]
- Peng, X.; Zhou, R.; Wang, B.; Yu, X.; Yang, X.; Liu, K.; Mi, M. Effect of green tea consumption on blood pressure: A meta-analysis of 13 randomized controlled trials. Sci. Rep. 2014, 4, 6251. [Google Scholar] [CrossRef]
- Sainz, J.; Wangensteen, R.; Rodríguez Gómez, I.; Moreno, J.M.; Chamorro, V.; Osuna, A.; Bueno, P.; Vargas, F. Antioxidant enzymes and effects of tempol on the development of hypertension induced by nitric oxide inhibition. Am. J. Hypertens. 2005, 18, 871–877. [Google Scholar] [CrossRef] [PubMed]
- Pechánová, O.; Bernátová, I.; Babál, P.; Martínez, M.C.; Kyselá, S.; Stvrtina, S.; Andriantsitohaina, R. Red wine polyphenols prevent cardiovascular alterations in L-NAME-induced hypertension. J. Hypertens. 2004, 22, 1551–1559. [Google Scholar] [CrossRef]
- Procházková, D.; Boušová, I.; Wilhelmová, N. Antioxidant and prooxidant properties of flavonoids. Fitoterapia 2011, 82, 513–523. [Google Scholar] [CrossRef]
- Ecobichon, D.J. The Basis of Toxicity Testing; CRC Press: Boca Raton, FL, USA, 1992. [Google Scholar]
- Morrison, A.; Polisena, J.; Husereau, D.; Moulton, K.; Clark, M.; Fiander, M.; Mierzwinski-Urban, M.; Clifford, T.; Hutton, B.; Rabb, D. The effect of English-language restriction on systematic review-based meta-analyses: A systematic review of empirical studies. Int. J. Technol. Assess. Health Care 2012, 28, 138–144. [Google Scholar] [CrossRef] [PubMed]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Keywords | |
|---|---|
| Oil Palm Leaves | “Elaeis guineensis” OR “Oil Palm Leaves” OR “Oil Palm Leaf” OR “Oil Palm Frond” OR “Oil Palm Fronds” |
| Bioactive Compounds | “Phytochemicals” OR “Phytochemical” OR “Phytonutrients” OR “Phytonutrient” OR “Flavonoids” OR “Flavonoid” OR “Chalcones” OR “Chalcone” OR “Flavones” OR “Flavone” OR “Isoflavones” OR “Isoflavone” OR “Flavanols” OR “Flavanol” OR “Flavanones” OR “Flavanone” OR “Flavonols” OR “Flavonol” OR “Anthocyanidins” OR “Anthocyanidin” OR “Phenolics” OR “Phenolic” OR “Phenolic acids” OR “Phenolic acid” OR “Coumarins” OR “Coumarin” OR “Lignins” OR “Lignin” |
| Reference | Methods for Detecting and Identifying Flavonoids | Types of Flavonoids | TPC (mg GAE g−1 Sample or Extract) | TFC |
|---|---|---|---|---|
| [2] | N/R | N/R | INSOL fraction: 118.44 ± 0.0 9BHT (positive control): 116.22 ± 0.04 MeOH extract: 63.67 ± 0.14 WATER extract: 61.13 ± 0.28 EA extract: 44.03 ± 0.03 HEX extract: 16.96 ± 0.13 | N/R |
| [4] | FTIR Spectroscopy | Flavonoids | Moderately high phenolic content of the aqueous methanolic extract: 0.33 | N/R |
| [18] | HPLC | Luteolin | MeOH extract: 24.3 ± 1.7 Green text extract: 22.5 ± 1.7 | N/R |
| [19] | HPLC | (−)-Epigallocatechin, (+)-catechin, (−)-epicatechin, (−)-epigallocatechin gallate, (−)-epicatechin gallate, and their glucosides | N/R | N/R |
| [20] | N/R | N/R | Extract prepared from dried leaves: 10.2 Extract prepared from wet leaves: 5.1 | N/R |
| [11] | N/R | N/R | OPAL M1: 1.160 ± 0.001 OPAL M2: 0.982 ± 0.001 OPAL M3 0.304 ± 0.044 OPAL M4: 0.440 ± 0.006 Classification refers to Table S1. | OPAL M1: 1.16 ± 0.001 c OPAL M2: 0.55 ± 0.002 c OPAL M3 0.30 ± 0.002 c OPAL M4: 0.33 ± 0.001 c |
| [21] | UHPLC-UV/PDA LCMS/MS | Major compounds: orientin, isoorientin, vitexin, isovitexin Minor compounds: luteolin-6-8-di-C-hexose, apigenin-6,8-di-C-hexose, luteolin-6-8-di-C-hexose, apigenin-6-C-pentose-8-C-hexose, apigenin-6-C-hexose-8-C-pentose, luteolin-6-C-hexose-8-C-deoxyhexose, and apigenin-6-C-hexose-8-C-deoxyhexose. | N/R | N/R |
| [22] | NMR Spectroscopy | (+)-Catechin, vitexin, isovitexin, orientin, isoorientin, and (−)-epicatechin | Aqueous methanol extract: 393.61 ± 36.11 Absolute methanol extract: 213.08 ± 41.61 Ethyl acetate-methanol extract: 174.19 ± 32.40 Ethyl acetate extract: 121.71 ± 32.78 | Aqueous methanol extract: 129.72 ± 8.70 b Absolute methanol extract: 135.40 ± 9.76 b Ethyl acetate-methanol extract: 121.48 ± 6.67 b Ethyl acetate extract: 5.94 ± 5.38 b |
| [23] | Phytochemical qualitative analysis | Flavonoids | Ethanol extract: 52.4 | Ethanol extract: 15.4 a |
| [5] | Phytochemical qualitative analysis | Flavonoids | N/R | N/R |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Tow, W.-K.; Goh, A.P.-T.; Sundralingam, U.; Palanisamy, U.D.; Sivasothy, Y. Flavonoid Composition and Pharmacological Properties of Elaeis guineensis Jacq. Leaf Extracts: A Systematic Review. Pharmaceuticals 2021, 14, 961. https://doi.org/10.3390/ph14100961
Tow W-K, Goh AP-T, Sundralingam U, Palanisamy UD, Sivasothy Y. Flavonoid Composition and Pharmacological Properties of Elaeis guineensis Jacq. Leaf Extracts: A Systematic Review. Pharmaceuticals. 2021; 14(10):961. https://doi.org/10.3390/ph14100961
Chicago/Turabian StyleTow, Wai-Kit, Asly Poh-Tze Goh, Usha Sundralingam, Uma Devi Palanisamy, and Yasodha Sivasothy. 2021. "Flavonoid Composition and Pharmacological Properties of Elaeis guineensis Jacq. Leaf Extracts: A Systematic Review" Pharmaceuticals 14, no. 10: 961. https://doi.org/10.3390/ph14100961
APA StyleTow, W.-K., Goh, A. P.-T., Sundralingam, U., Palanisamy, U. D., & Sivasothy, Y. (2021). Flavonoid Composition and Pharmacological Properties of Elaeis guineensis Jacq. Leaf Extracts: A Systematic Review. Pharmaceuticals, 14(10), 961. https://doi.org/10.3390/ph14100961