
Folium Sennae Increased the Bioavailability of Methotrexate through Modulation on MRP 2 and BCRP

Abstract
:1. Introduction
2. Results
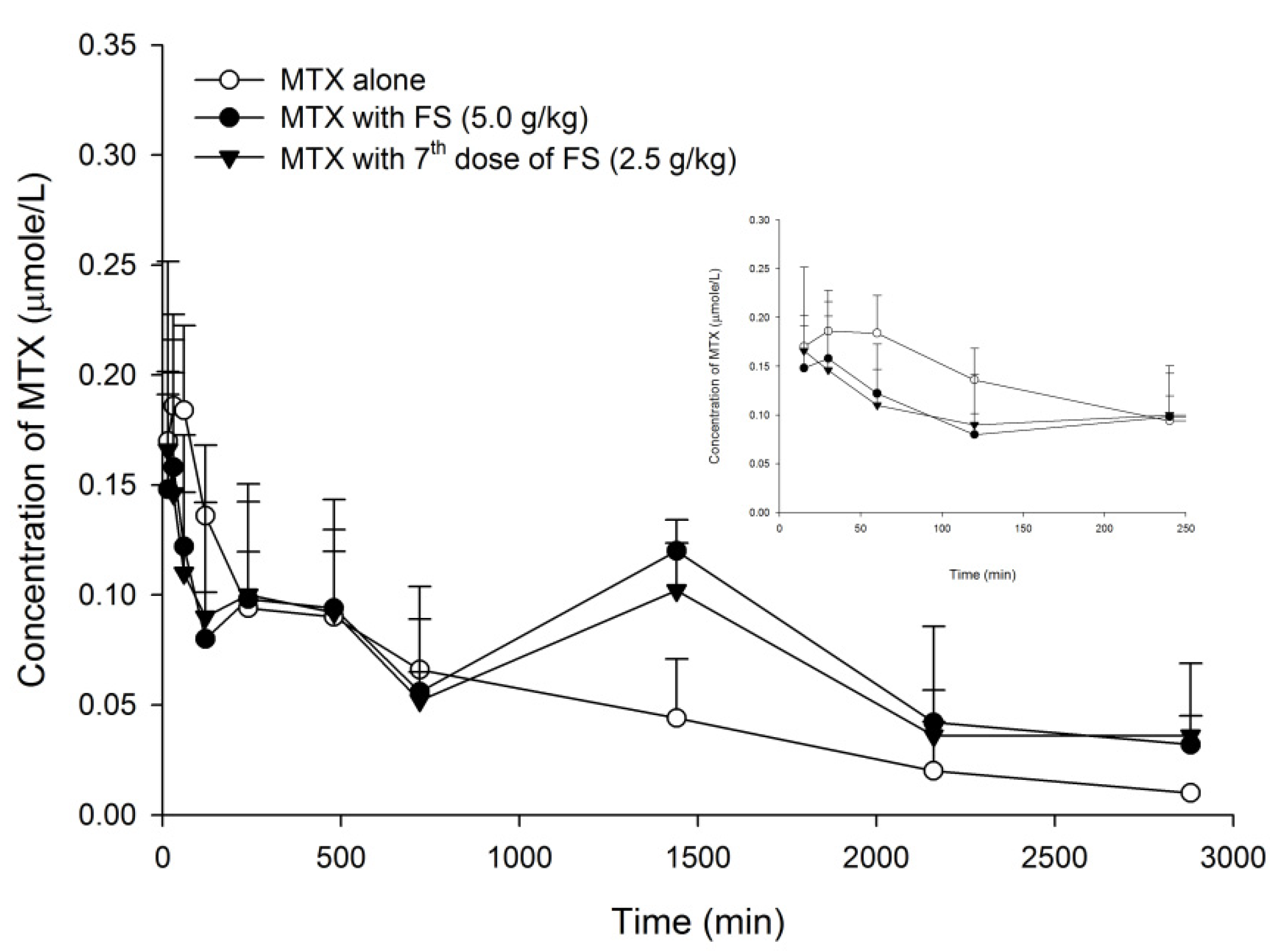
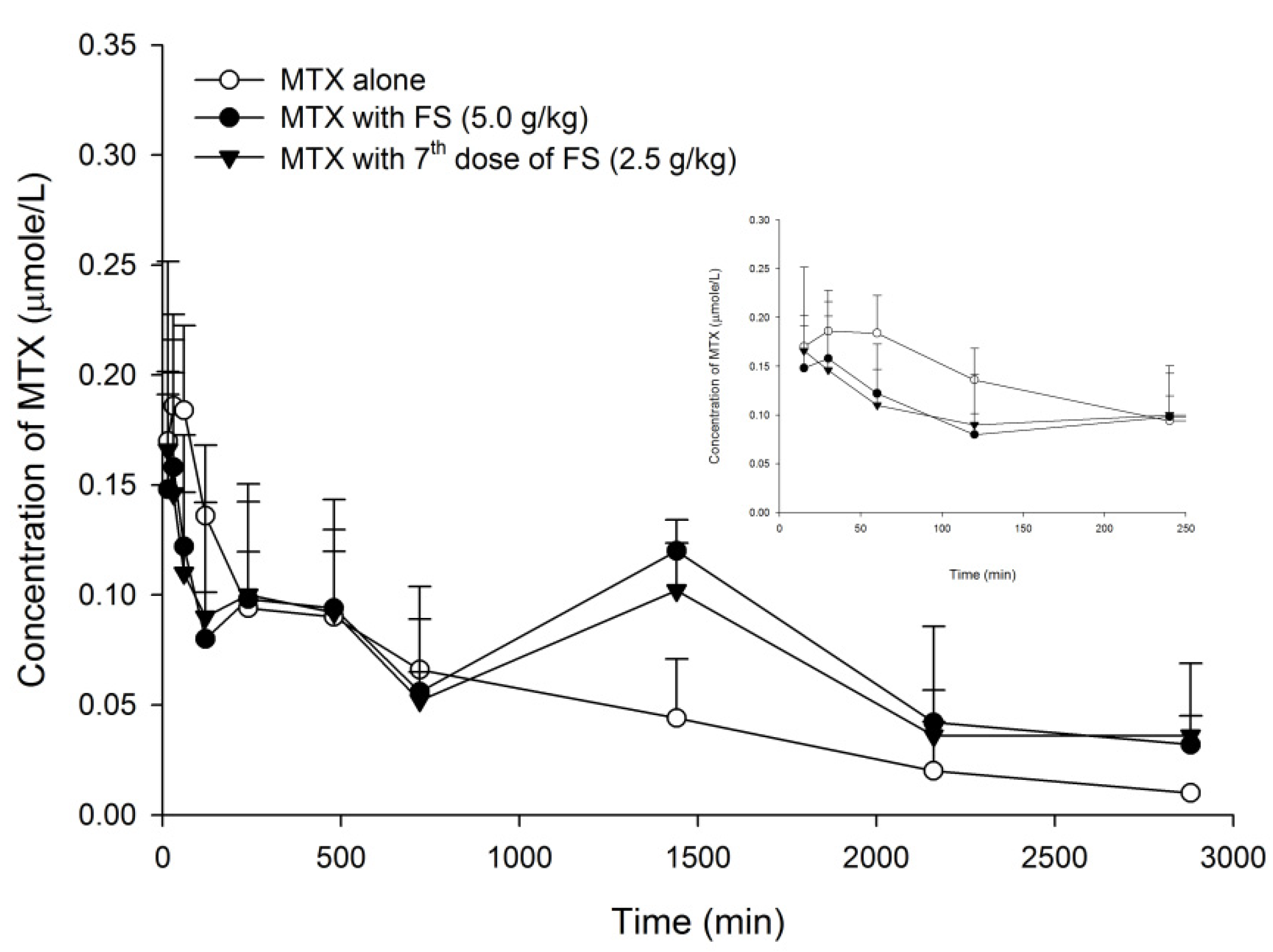
2.1. FS-MTX Pharmacokinetic Interaction and Relevant Mechanisms
2.1.1. Effect of FS on MTX Pharmacokinetics in Rats
2.1.2. Characterization of FSM
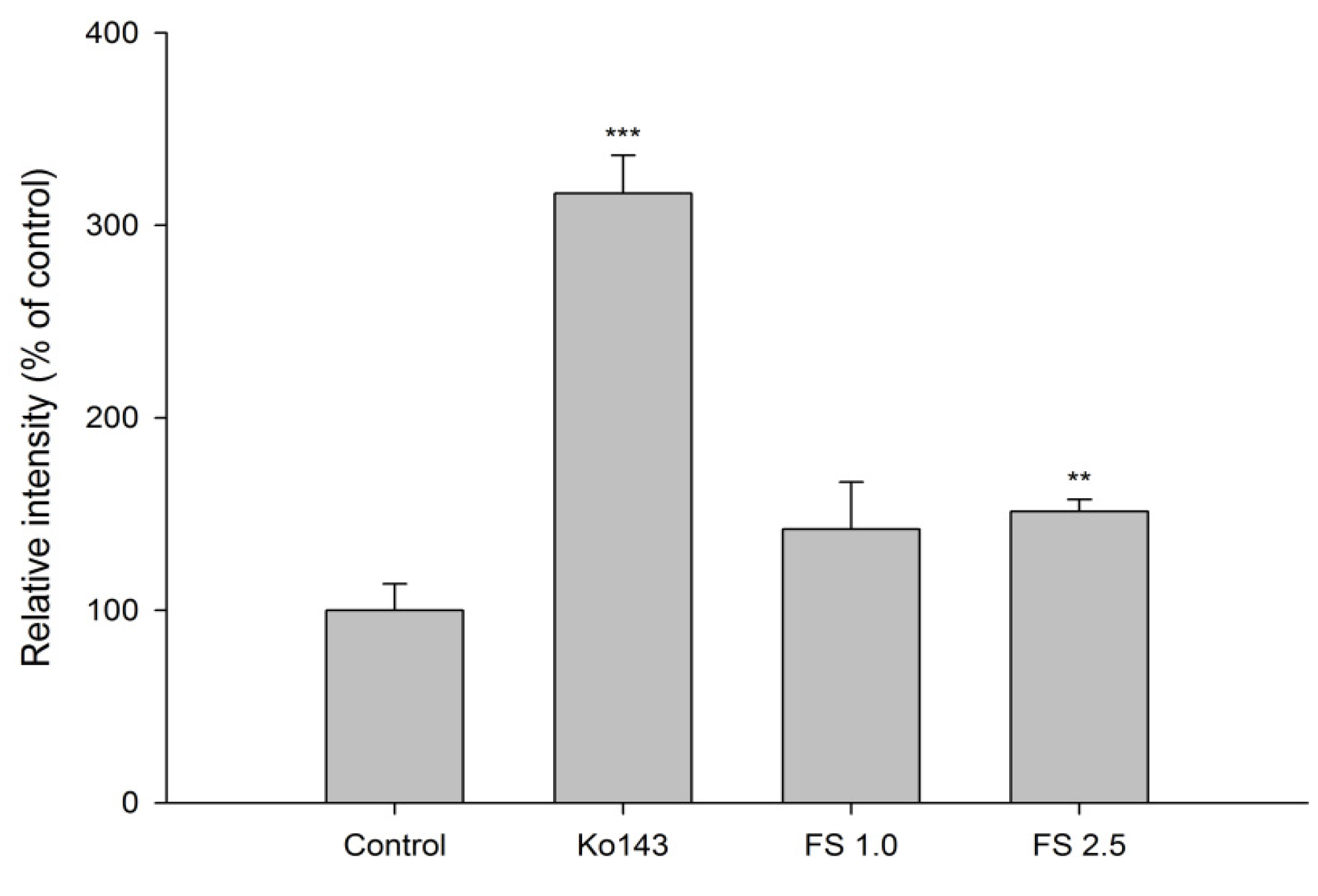
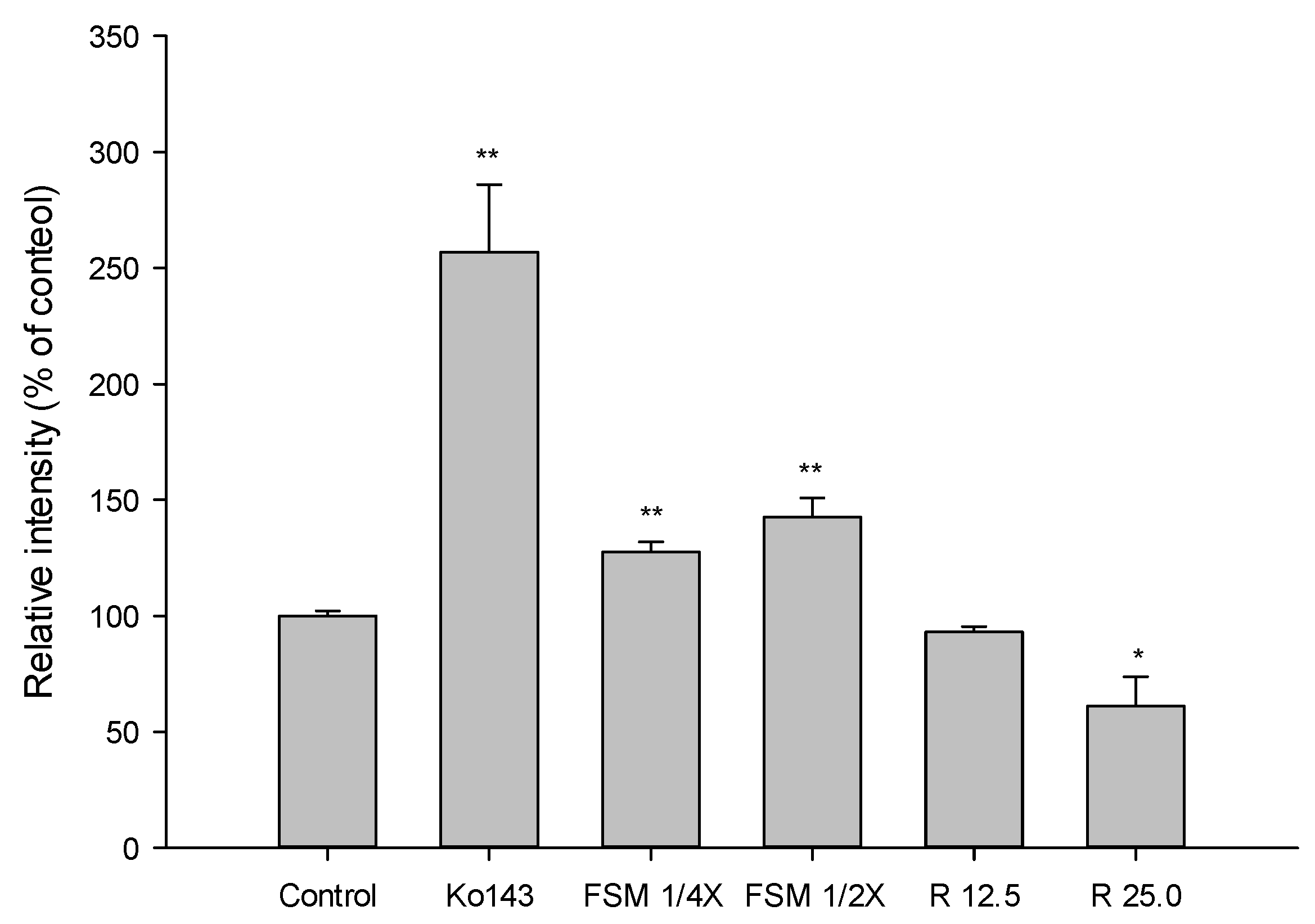
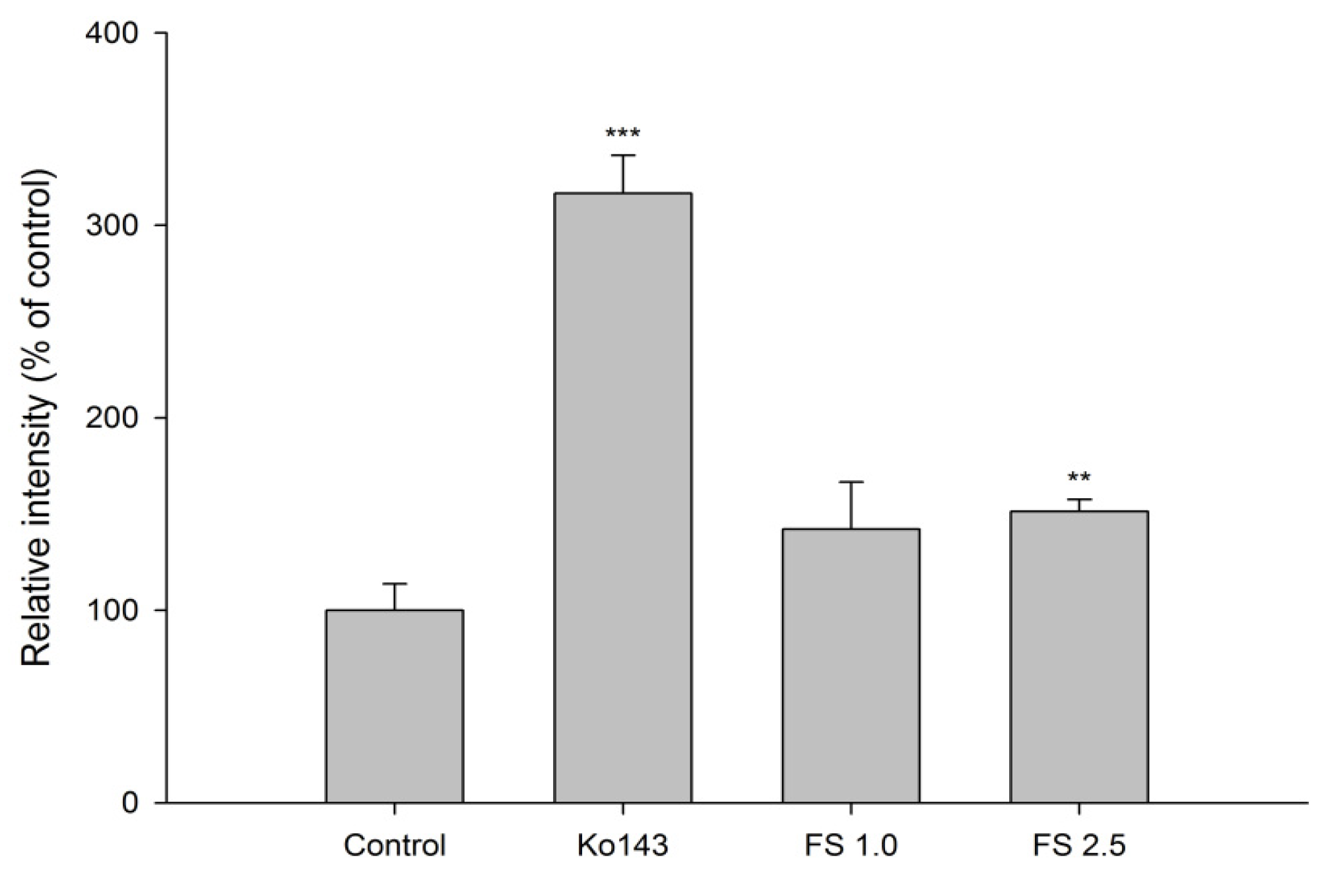
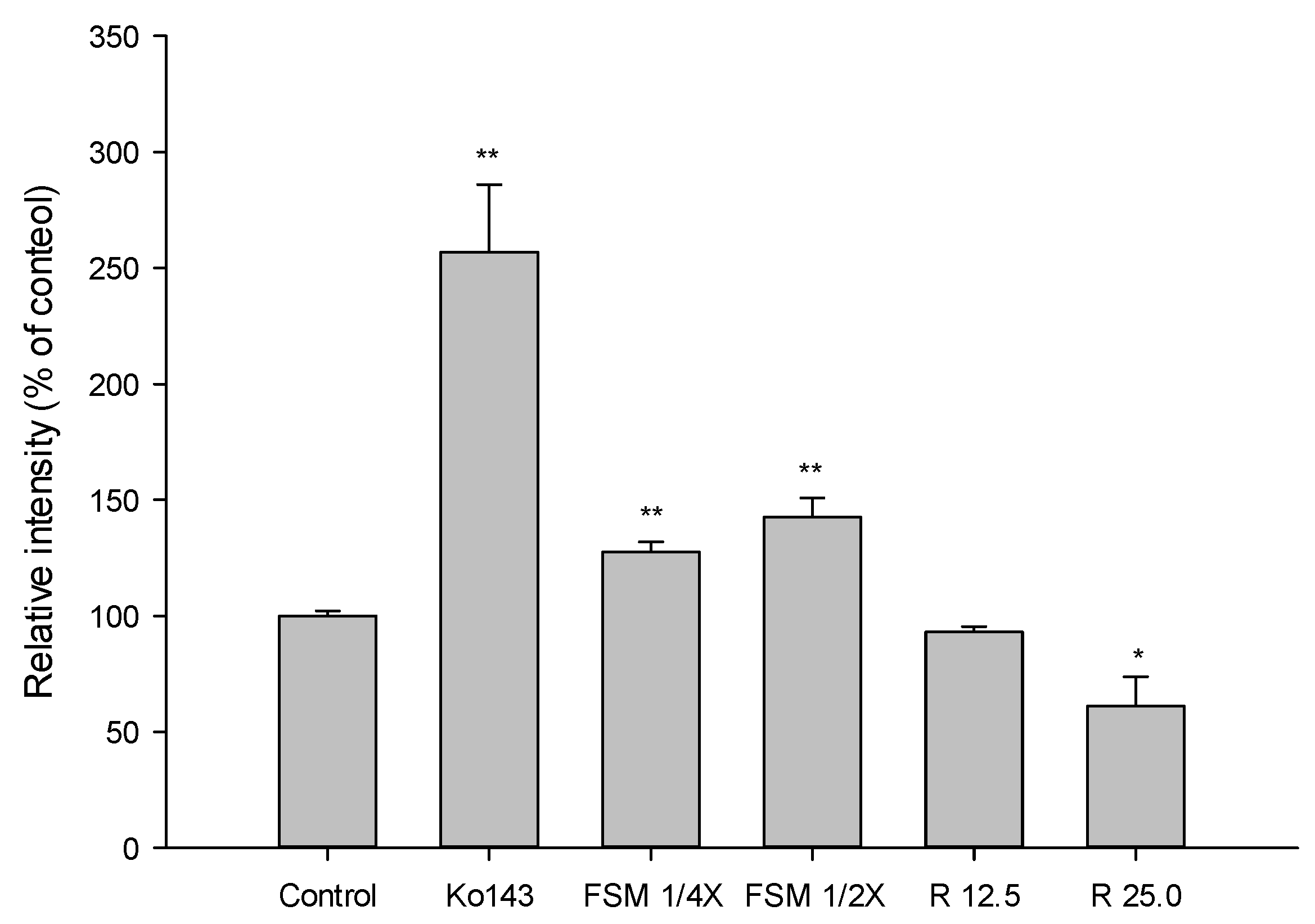
2.1.3. Effects of FS, FSM and Rhein on BCRP Activity
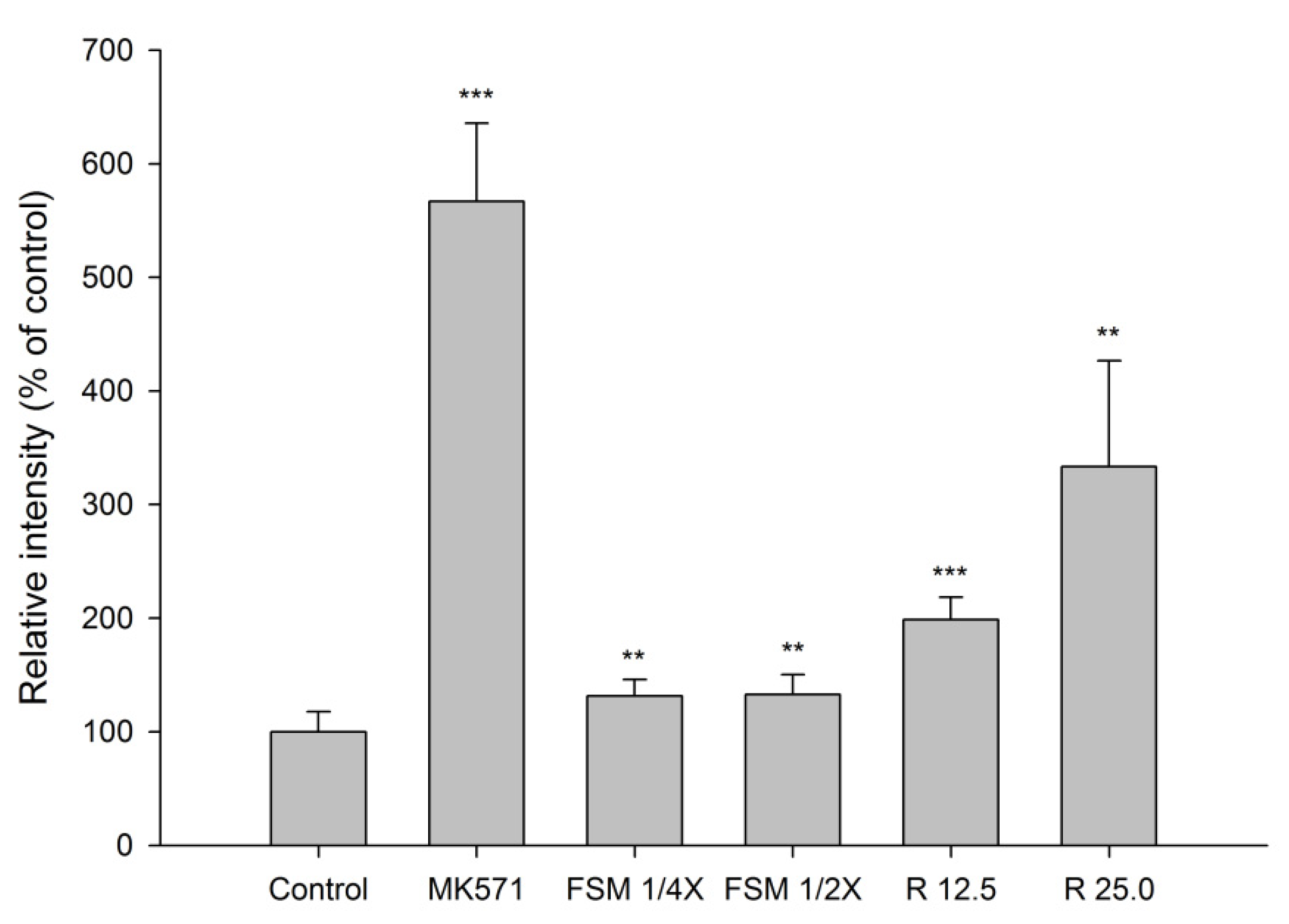
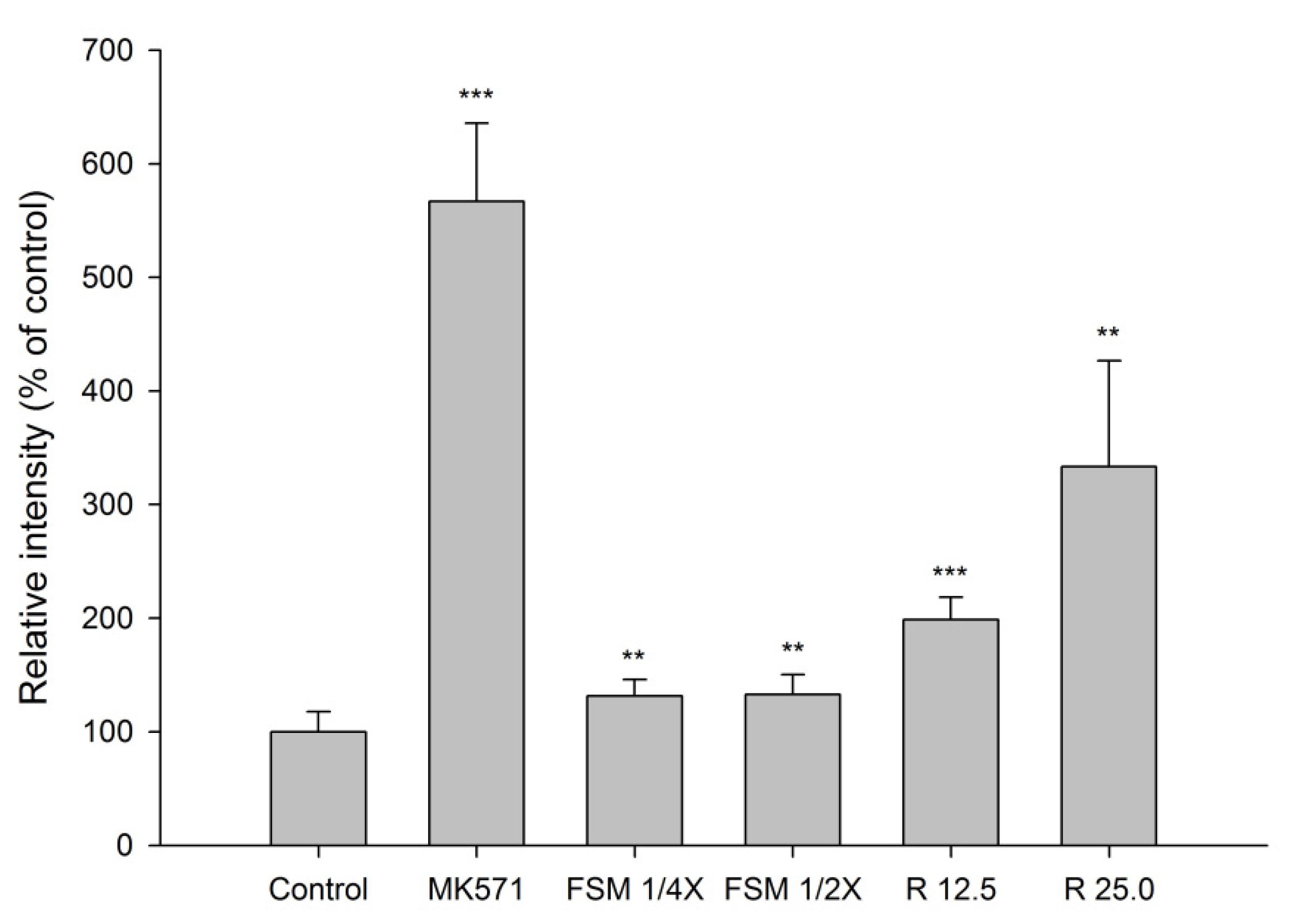
2.1.4. Effect of FSM and Rhein on MRP 2 Activity
3. Discussions
4. Materials and Methods
4.1. Chemicals and Reagents
4.2. Preparation and Characterization of FS Decoction
4.3. Animals and Drug Administration
4.4. Blood Collection and Determination of Serum MTX Concentration
4.5. Cell Line and Culture Conditions
4.6. Cell Viability Assay
4.7. Preparation and Characterization of Serum Metabolites of FS (FSM)
4.8. Effects of FS, FSM and Rhein on BCRP Activity
4.9. Effect of FSM and Rhein on MRP 2 Activity
4.10. Data Analysis
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Mueller-Lissner, S.A.; Wald, A. Constipation in adults. BMJ Clin. Evid. 2010, 2010, 0413. [Google Scholar] [PubMed]
- Bharucha, A.E.; Wald, A. Chronic constipation. Mayo Clin. Proc. 2019, 94, 2340–2357. [Google Scholar] [CrossRef]
- Fragakis, A.; Zhou, J.; Mannan, H.; Ho, V. Association between drug usage and constipation in the elderly population of Greater Western Sydney Australia. Int. J. Environ. Res. Public Health 2018, 15, 226. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tariq, S.H. Constipation in long-term care. J. Am. Med. Dir. Assoc. 2007, 8, 209–218. [Google Scholar] [CrossRef]
- Alsalimy, N.; Madi, L.; Awaisu, A. Efficacy and safety of laxatives for chronic constipation in long-term care settings: A systematic review. J. Clin. Pharm. Ther. 2018, 43, 595–605. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Every-Palmer, S.; Newton-Howes, G.; Clarke, M.J. Pharmacological treatment for antipsychotic-related constipation. Cochrane Database Syst. Rev. 2017, 1, CD011128. [Google Scholar]
- Collamati, A.; Martone, A.M.; Poscia, A.; Brandi, V.; Celi, M.; Marzetti, E.; Cherubini, A.; Landi, F. Anticholinergic drugs and negative outcomes in the older population: From biological plausibility to clinical evidence. Aging Clin. Exp. Res. 2016, 28, 25–35. [Google Scholar] [CrossRef]
- Cirillo, C.; Capasso, R. Constipation and botanical medicines: An overview. Phytother. Res. 2015, 29, 1488–1493. [Google Scholar] [CrossRef]
- Leung, L.; Riutta, T.; Kotecha, J.; Rosser, W. Chronic constipation: An evidence-based review. J. Am. Board Fam. Med. 2011, 24, 436–451. [Google Scholar] [CrossRef]
- Garcia-Alvarez, A.; Mila-Villarroel, R.; Ribas-Barba, L.; Egan, B.; Badea, M.; Maggi, F.M.; Salmenhaara, M.; Restani, P.; Serra-Majem, L. Usage of Plant Food Supplements (PFS) for weight control in six European countries: Results from the PlantLIBRA PFS Consumer Survey 2011–2012. BMC Complement. Altern. Med. 2016, 16, 254. [Google Scholar] [CrossRef] [Green Version]
- Ulbricht, C.; Conquer, J.; Costa, D.; Hamilton, W.; Higdon, E.R.; Isaac, R.; Rusie, E.; Rychlik, I.; Serrano, J.M.; Tanguay-Colucci, S.; et al. An evidence-based systematic review of senna (Cassia senna) by the Natural Standard Research Collaboration. J. Diet. Suppl. 2011, 8, 189–238. [Google Scholar] [CrossRef]
- Franz, G. The senna drug and its chemistry. Pharmacology 1993, 47 (Suppl. 1), 2–6. [Google Scholar] [CrossRef] [PubMed]
- Bharti, S.K.; Krishnan, S.; Kumar, A. Antidiabetic phytoconstituents and their mode of action on metabolic pathways. Ther. Adv. Endocrinol. Metab. 2018, 9, 81–100. [Google Scholar] [CrossRef] [PubMed]
- Choi, R.J.; Ngoc, T.M.; Bae, K.; Cho, H.J.; Kim, D.D.; Chun, J.; Khan, S.; Kim, Y.S. Anti-inflammatory properties of anthraquinones and their relationship with the regulation of P-glycoprotein function and expression. Eur. J. Pharm. Sci. 2013, 48, 272–281. [Google Scholar] [CrossRef]
- Huang, Q.; Lu, G.; Shen, H.M.; Chung, M.C.; Ong, C.N. Anti-cancer properties of anthraquinones from rhubarb. Med. Res. Rev. 2007, 27, 609–630. [Google Scholar] [CrossRef]
- Sun, H.; Luo, G.; Chen, D.; Xiang, Z. A Comprehensive and system review for the pharmacological mechanism of action of rhein, an active anthraquinone ingredient. Front. Pharmacol. 2016, 7, 247. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- DeGorter, M.K.; Xia, C.Q.; Yang, J.J.; Kim, R.B. Drug transporters in drug efficacy and toxicity. Annu. Rev. Pharmacol. Toxicol. 2012, 52, 249–273. [Google Scholar] [CrossRef]
- Zha, W. Transporter-mediated natural product-drug interactions for the treatment of cardiovascular diseases. J. Food Drug Anal. 2018, 26, S32–S44. [Google Scholar] [CrossRef] [Green Version]
- Liu, X. Transporter-mediated drug-drug interactions and their significance. Adv. Exp. Med. Biol. 2019, 1141, 241–291. [Google Scholar]
- Gessner, A.; König, J.; Fromm, M.F. Clinical aspects of transporter-mediated drug-drug interactions. Clin. Pharmacol. Ther. 2019, 105, 1386–1394. [Google Scholar] [CrossRef] [PubMed]
- Nakanishi, T.; Tamai, I. Interaction of drug or food with drug transporters in intestine and liver. Curr. Drug Metab. 2015, 16, 753–764. [Google Scholar] [CrossRef] [PubMed]
- Nigam, S.K. What do drug transporters really do? Nat. Rev. Drug Discov. 2015, 14, 29–44. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lai, R.E.; Jay, C.E.; Sweet, D.H. Organic solute carrier 22 (SLC22) family: Potential for interactions with food, herbal/dietary supplements, endogenous compounds, and drugs. J. Food Drug Anal. 2018, 26, S45–S60. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Peng, Y.H.; Lin, S.P.; Yu, C.P.; Tsai, S.Y.; Chen, M.Y.; Hou, Y.C.; Chao, P.D. Serum concentrations of anthraquinones after intake of Folium Sennae and potential modulation on P-glycoprotein. Planta Med. 2014, 80, 1291–1297. [Google Scholar] [CrossRef] [Green Version]
- Yang, M.S.; Yu, C.P.; Huang, C.Y.; Chao, P.L.; Lin, S.P.; Hou, Y.C. Aloe activated P-glycoprotein and CYP 3A: A study on the serum kinetics of aloe and its interaction with cyclosporine in rats. Food Funct. 2017, 8, 315–322. [Google Scholar] [CrossRef]
- Dai, Y.; Ma, B.L.; Zheng, M.; Shi, R.; Li, Y.Y.; Wang, T.M.; Ma, Y.M. Identification of drug transporters involved in the uptake and efflux of rhein in hepatocytes. RSC Adv. 2017, 7, 15236–15245. [Google Scholar] [CrossRef] [Green Version]
- Ma, L.; Zhao, L.; Hu, H.; Qin, Y.; Bian, Y.; Jiang, H.; Zhou, H.; Yu, L.; Zeng, S. Interaction of five anthraquinones from rhubarb with human organic anion transporter 1 (SLC22A6) and 3 (SLC22A8) and drug-drug interaction in rats. J. Ethnopharmacol. 2014, 153, 864–871. [Google Scholar] [CrossRef]
- Lin, S.P.; Hou, Y.C.; Tsai, S.Y.; Wang, M.J.; Chao, P.D. Tissue distribution of naringenin conjugated metabolites following repeated dosing of naringin to rats. Biomedicine 2014, 4, 16. [Google Scholar] [CrossRef]
- Dabeek, W.M.; Marra, M.V. Dietary quercetin and kaempferol: Bioavailability and potential cardiovascular-related bioactivity in humans. Nutrients 2019, 11, 2288. [Google Scholar] [CrossRef] [Green Version]
- Manach, C.; Scalbert, A.; Morand, C.; Rémésy, C.; Jiménez, L. Polyphenols: Food sources and bioavailability. Am. J. Clin. Nutr. 2004, 79, 727–747. [Google Scholar] [CrossRef] [Green Version]
- Ye, L.; Lu, L.; Li, Y.; Zeng, S.; Yang, X.; Chen, W.; Feng, Q.; Liu, W.; Tang, L.; Liu, Z. Potential role of ATP-binding cassette transporters in the intestinal transport of rhein. Food Chem. Toxicol. 2013, 58, 301–305. [Google Scholar] [CrossRef] [PubMed]
- Deeley, R.G.; Westlake, C.; Cole, S.P. Transmembrane transport of endo- and xenobiotics by mammalian ATP-binding cassette multidrug resistance proteins. Physiol. Rev. 2006, 86, 849–899. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, Y.K.; Wang, Y.J.; Gupta, P.; Chen, Z.S. Multidrug resistance proteins (MRPs) and cancer therapy. AAPS J. 2015, 17, 802–812. [Google Scholar] [CrossRef] [Green Version]
- Murakami, T.; Mori, N. Involvement of multiple transporters-mediated transports in mizoribine and methotrexate pharmacokinetics. Pharmaceuticals 2012, 5, 802–836. [Google Scholar] [CrossRef] [Green Version]
- Mao, Q.; Unadkat, J.D. Role of the breast cancer resistance protein (BCRP/ABCG2) in drug transport—An update. AAPS J. 2015, 17, 65–82. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Stamp, L.K.; Hazlett, J.; Highton, J.; Hessian, P.A. Expression of methotrexate transporters and metabolizing enzymes in rheumatoid synovial tissue. J. Rheumatol. 2013, 40, 1519–1522. [Google Scholar] [CrossRef] [PubMed]
- Vlaming, M.L.; van Esch, A.; van de Steeg, E.; Pala, Z.; Wagenaar, E.; van Tellingen, O.; Schinkel, A.H. Impact of abcc2 [multidrug resistance-associated protein (MRP) 2], abcc3 (MRP3), and abcg2 (breast cancer resistance protein) on the oral pharmacokinetics of methotrexate and its main metabolite 7-hydroxymethotrexate. Drug Metab. Dispos. 2011, 39, 1338–1344. [Google Scholar] [CrossRef] [PubMed]
- Chen, Z.; Shi, T.; Zhang, L.; Zhu, P.; Deng, M.; Huang, C.; Hu, T.; Jiang, L.; Li, J. Mammalian drug efflux transporters of the ATP binding cassette (ABC) family in multidrug resistance: A review of the past decade. Cancer Lett. 2016, 370, 153–164. [Google Scholar] [CrossRef]
- Fardel, O.; Jigorel, E.; Le Vee, M.; Payen, L. Physiological, pharmacological and clinical features of the multidrug resistance protein 2. Biomed. Pharmacother. 2005, 59, 104–114. [Google Scholar] [CrossRef] [PubMed]
- Lemli, J.; Lemmens, L. Metabolism of sennosides and rhein in the rat. Pharmacology 1980, 20 (Suppl. 1), 50–57. [Google Scholar] [CrossRef]
- Dreessen, M.; Eyssen, H.; Lemli, J. The metabolism of sennosides A and B by the intestinal microflora: In vitro and in vivo studies on the rat and the mouse. J. Pharm. Pharmacol. 1981, 33, 679–681. [Google Scholar] [CrossRef]
- Mosmann, T. Rapid colorimetric assay for cellular growth and survival: Application to proliferation and cytotoxicity assays. J. Immunol. Methods 1983, 65, 55–63. [Google Scholar] [CrossRef]
- Hsu, P.W.; Shia, C.S.; Wu, C.T.; Chang, N.W.; Chao, P.D.L.; Hou, Y.C. Noni increased the systemic exposure of methotrexate in rats through inhibition on multi-drug resistance protein 2 and breast cancer resistance protein (BCRP). J. Funct. Foods 2013, 5, 1414–1420. [Google Scholar] [CrossRef]
- Yang, S.Y.; Juang, S.H.; Tsai, S.Y.; Chao, P.D.; Hou, Y.C. St. John’s wort significantly increased the systemic exposure and toxicity of methotrexate in rats. Toxicol. Appl. Pharmacol. 2012, 263, 39–43. [Google Scholar] [CrossRef] [PubMed]
- Yu, C.P.; Hsieh, Y.C.; Shia, C.S.; Hsu, P.W.; Chen, J.Y.; Hou, Y.C.; Hsieh, Y.W. Increased systemic exposure of methotrexate by a polyphenol-rich herb via modulation on efflux transporters multidrug resistance-associated protein 2 and breast cancer resistance protein. J. Pharm. Sci. 2016, 105, 343–349. [Google Scholar] [CrossRef] [PubMed]

{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Parameters | MTX Alone | MTX + FS (5.0 g/kg, Single Dose) | MTX + FS (2.5 g/kg, 7 Doses) | ||||
|---|---|---|---|---|---|---|---|
| Treatments | |||||||
| Cmax | 0.2 | ±0.02 | 0.2 | ±0.02 | 0.2 | ±0.03 | |
| AUC 0–240 | 32.9 | ±3.0 | 24.3 | ±2.5 | 24.8 | ±4.5 | |
| (−26%) | (−25%) | ||||||
| AUC 720–2880 | 73.4 | ±14.9 a | 148.3 | ±15.6 b | 131.7 | ±4.8 b | |
| (+102%) | (+78%) | ||||||
| AUC 0–2880 | 147.1 | ±15.6 a | 213.7 | ±18.6 b | 196.2 | ±10.4 ab | |
| (+45%) | (+33%) | ||||||
| MRT 0–2880 | 841.9 | ±103.6 a | 1199.2 | ±68.3 b | 1195.8 | ±88.6 b | |
| (+42%) | (+42%) | ||||||
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Yu, C.-P.; Peng, Y.-H.; Huang, C.-Y.; Hsieh, Y.-W.; Hou, Y.-C.; Lin, S.-P. Folium Sennae Increased the Bioavailability of Methotrexate through Modulation on MRP 2 and BCRP. Pharmaceuticals 2021, 14, 1036. https://doi.org/10.3390/ph14101036
Yu C-P, Peng Y-H, Huang C-Y, Hsieh Y-W, Hou Y-C, Lin S-P. Folium Sennae Increased the Bioavailability of Methotrexate through Modulation on MRP 2 and BCRP. Pharmaceuticals. 2021; 14(10):1036. https://doi.org/10.3390/ph14101036
Chicago/Turabian StyleYu, Chung-Ping, Yu-Hsuan Peng, Ching-Ya Huang, Yow-Wen Hsieh, Yu-Chi Hou, and Shiuan-Pey Lin. 2021. "Folium Sennae Increased the Bioavailability of Methotrexate through Modulation on MRP 2 and BCRP" Pharmaceuticals 14, no. 10: 1036. https://doi.org/10.3390/ph14101036
APA StyleYu, C.-P., Peng, Y.-H., Huang, C.-Y., Hsieh, Y.-W., Hou, Y.-C., & Lin, S.-P. (2021). Folium Sennae Increased the Bioavailability of Methotrexate through Modulation on MRP 2 and BCRP. Pharmaceuticals, 14(10), 1036. https://doi.org/10.3390/ph14101036