Impact of S1P Mimetics on Mesenteric Ischemia/Reperfusion Injury

 ,
,  and
and 
Abstract
1. Introduction
2. Results
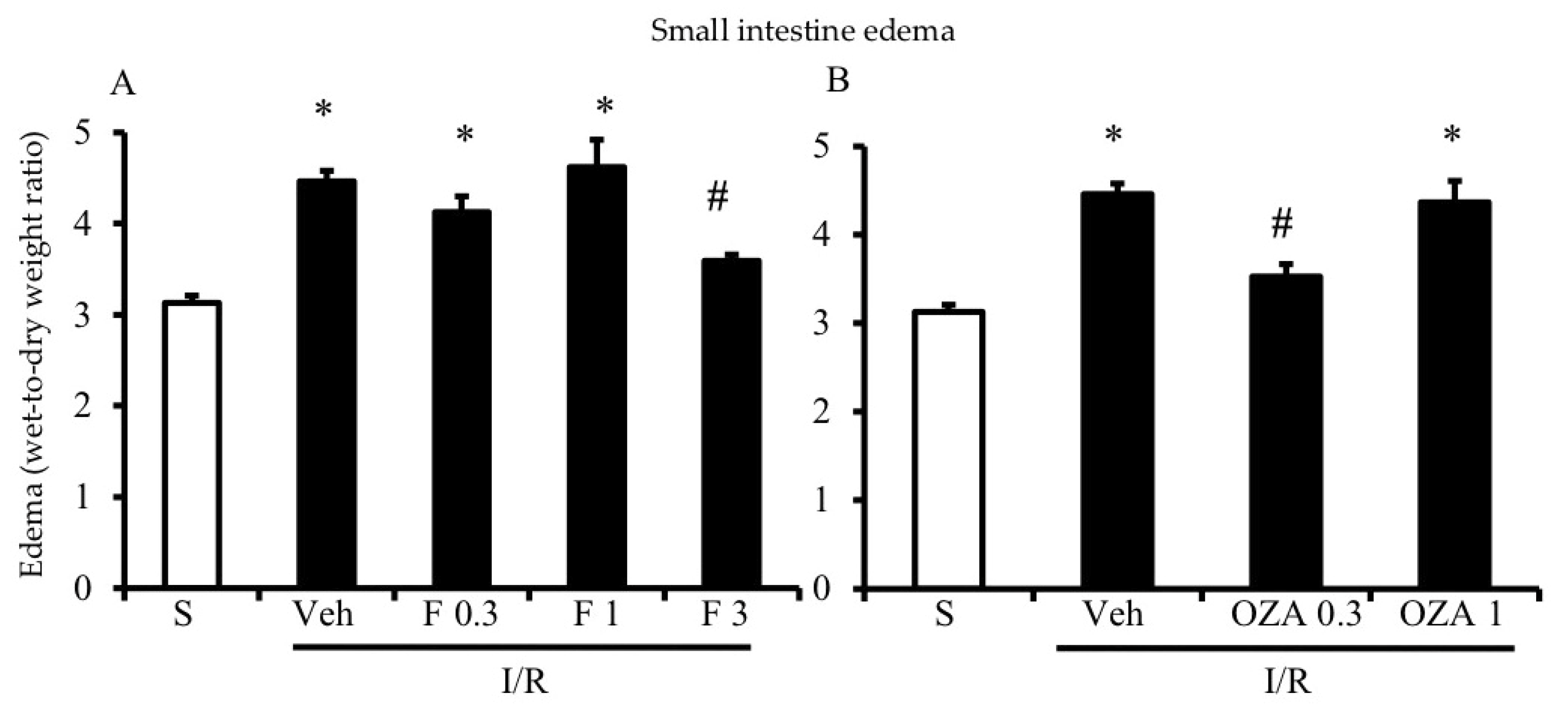
2.1. S1P Mimetics Decreased Vascular Permeability during Intestine I/R
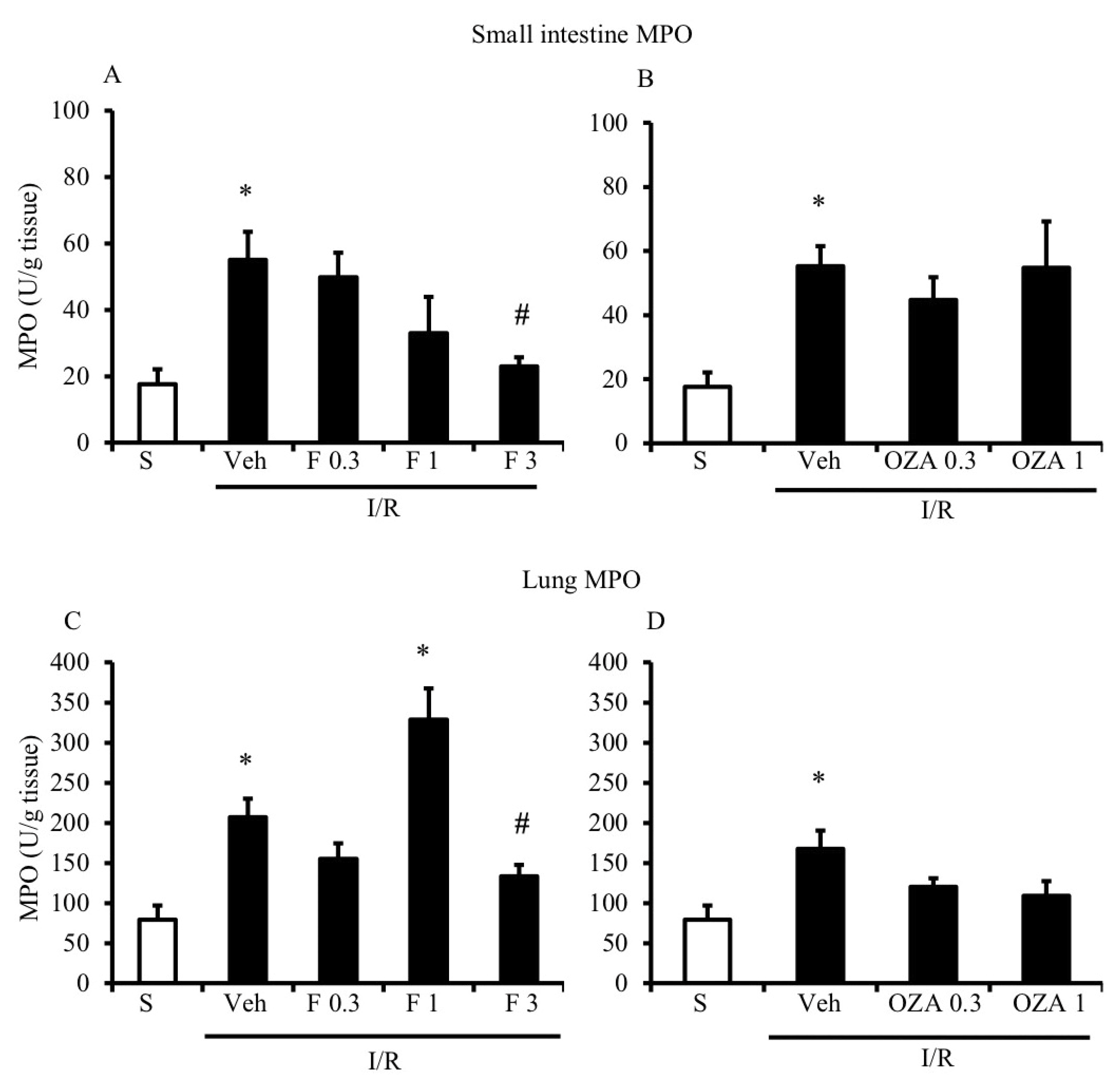
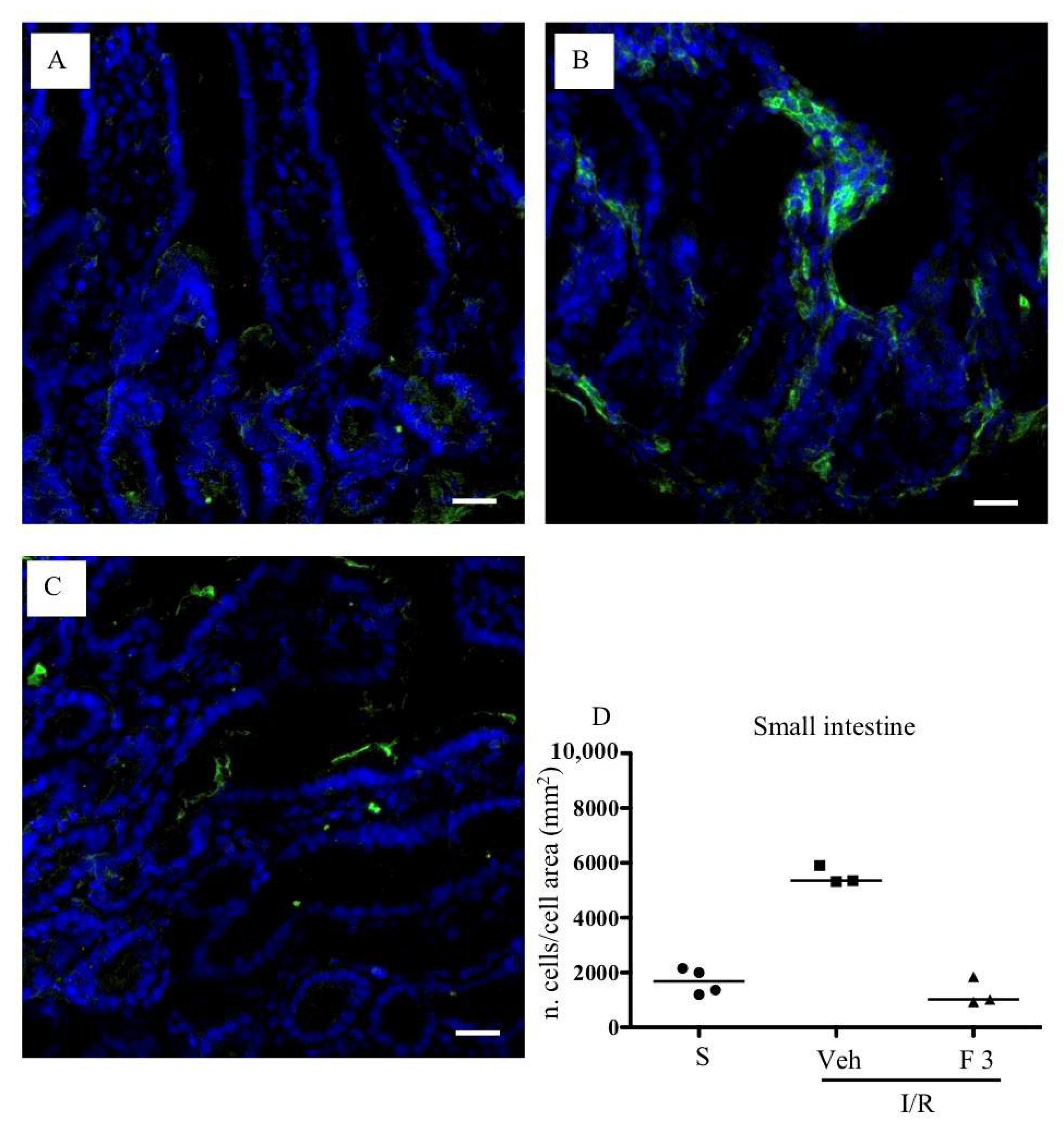
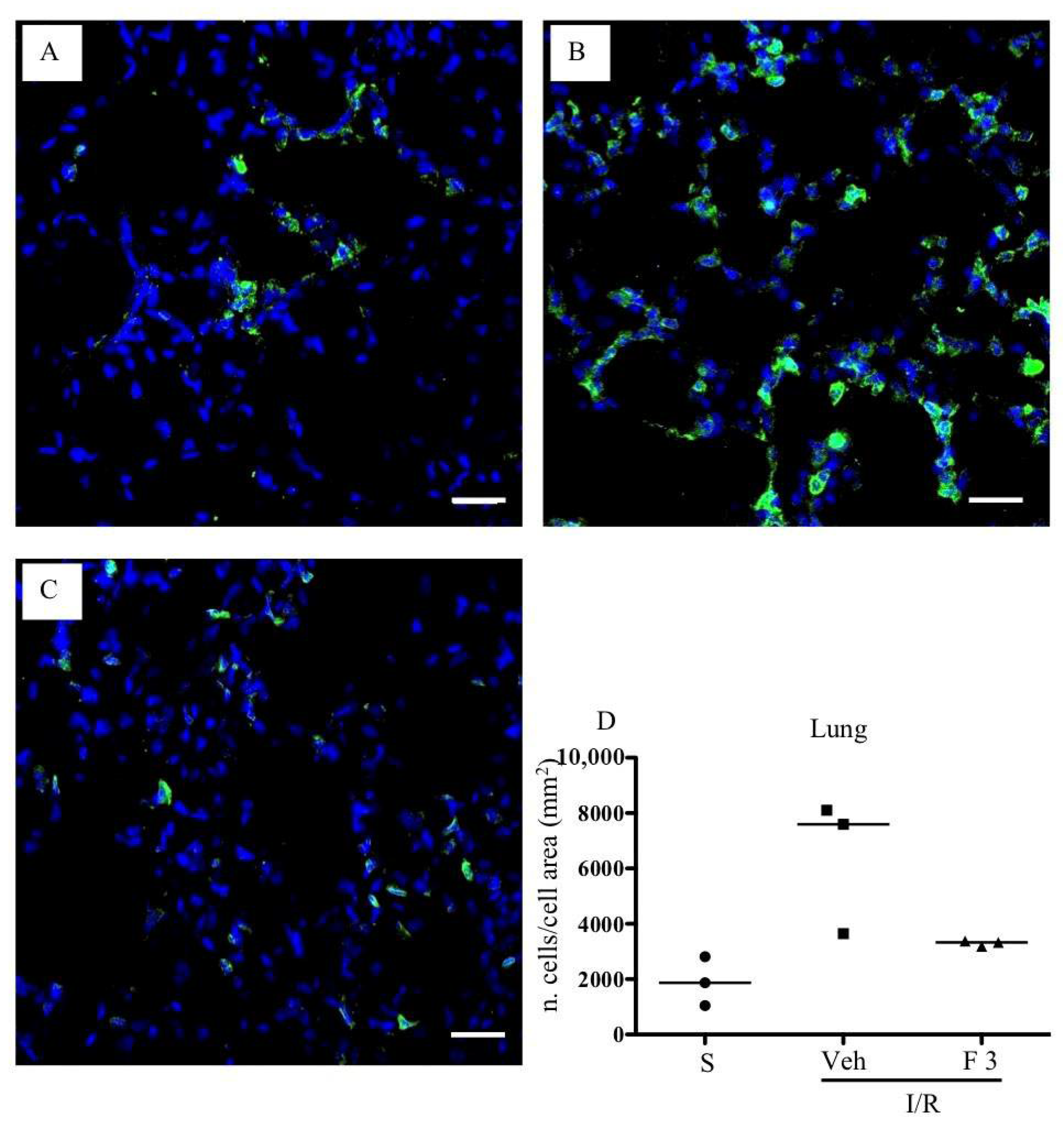
2.2. FTY720 but Not Ozanimod Mitigated I/R-Induced Recruitment of Leukocytes
2.3. S1P Mimetics Failed to Affect Lipoperoxidation during Intestine I/R
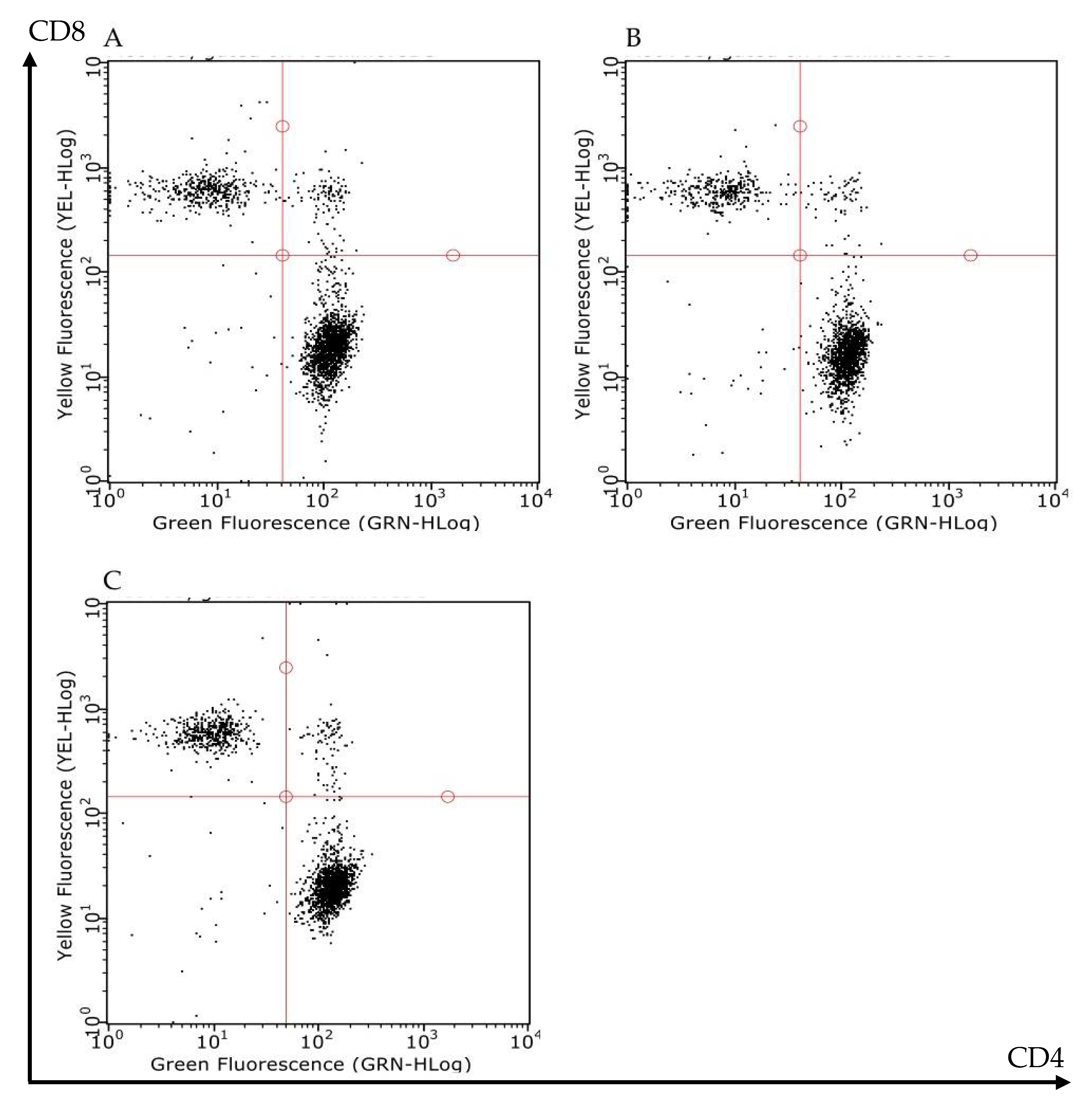
2.4. Mesenteric I/R Did Not Affect Mesenteric Lymph Nodes (MLNs) Lymphocytes Count
3. Discussion
4. Materials and Methods
4.1. Experimental Procedures
4.2. Drugs
4.3. Ischemia/Reperfusion
4.4. Myeloperoxidase Activity
4.5. Malondialdehyde Assay
4.6. Vascular Permeability
4.7. Flow Cytometric Assays
4.7.1. Isolation of Mesenteric Lymph Nodes (MLNs)
4.7.2. Immunofluorescent Staining
4.8. Histology and Immunofluorescence Analysis
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Maceyka, M.; Harikumar, K.B.; Milstien, S.; Spiegel, S. Sphingosine-1-phosphate signaling and its role in disease. Trends Cell Biol. 2012, 22, 50–60. [Google Scholar] [CrossRef] [PubMed]
- Sukocheva, O.A.; Furuya, H.; Ng, M.L.; Friedemann, M.; Menschikowski, M.; Tarasov, V.V.; Chubarev, V.N.; Klochkov, S.G.; Neganova, M.E.; Mangoni, A.A.; et al. Sphingosine kinase and sphingosine-1-phosphate receptor signaling pathway in inflammatory gastrointestinal disease and cancers: A novel therapeutic target. Pharmacol. Ther. 2020, 207, 107464. [Google Scholar] [CrossRef] [PubMed]
- Stepanovska, B.; Huwiler, A. Targeting the S1P receptor signaling pathways as a promising approach for treatment of autoimmune and inflammatory diseases. Pharmacol. Res. 2020, 154, 104170. [Google Scholar] [CrossRef]
- Huwiler, A.; Zangemeister-Wittke, U. The sphingosine 1-phosphate receptor modulator fingolimod as a therapeutic agent: Recent findings and new perspectives. Pharmacol. Ther. 2018, 185, 34–49. [Google Scholar] [CrossRef] [PubMed]
- Bertoni, S.; Ballabeni, V.; Barocelli, E.; Tognolini, M. Mesenteric ischemia-reperfusion: An overview of preclinical drug strategies. Drug Discov. Today 2018, 23, 1416–1425. [Google Scholar] [CrossRef] [PubMed]
- Troncoso, P.; Ortíz, M.; Martínez, L.; Kahan, B. FTY 720 prevents ischemic reperfusion damage in rat kidneys. Transplant. Proc. 2001, 33, 857–859. [Google Scholar] [CrossRef]
- Awad, A.S.; Ye, H.; Huang, L.; Li, L.; Foss, F.W.; Macdonald, T.L.; Lynch, K.R.; Okusa, M.D. Selective sphingosine 1-phosphate 1 receptor activation reduces ischemia-reperfusion injury in mouse kidney. Am. J. Physiol. Physiol. 2006, 290, F1516–F1524. [Google Scholar] [CrossRef]
- Man, K.; Ng, K.T.; Lee, T.K.W.; Lo, C.; Sun, C.K.; Li, X.L.; Zhao, Y.; Ho, J.W.; Fan, S.T. FTY720 Attenuates Hepatic Ischemia-Reperfusion Injury in Normal and Cirrhotic Livers. Arab. Archaeol. Epigr. 2005, 5, 40–49. [Google Scholar] [CrossRef]
- Stone, M.L.; Sharma, A.K.; Zhao, Y.; Charles, E.J.; Huerter, M.E.; Johnston, W.F.; Kron, I.L.; Lynch, K.R.; Laubach, V.E. Sphingosine-1-phosphate receptor 1 agonism attenuates lung ischemia-reperfusion injury. Am. J. Physiol. Cell. Mol. Physiol. 2015, 308, L1245–L1252. [Google Scholar] [CrossRef]
- Bonitz, J.A.; Son, J.Y.; Chandler, B.; Tomaio, J.N.; Qin, Y.; Prescott, L.M.; Feketeova, E.; Deitch, E.A. A Sphingosine-1 Phosphate Agonist (FTY720) Limits Trauma/Hemorrhagic Shock–Induced Multiple Organ Dysfunction Syndrome. Shock 2014, 42, 448–455. [Google Scholar] [CrossRef]
- Lamb, Y.N. Ozanimod: First Approval. Drugs 2020, 80, 841–848. [Google Scholar] [CrossRef] [PubMed]
- O’Sullivan, S.; Dev, K.K. Sphingosine-1-phosphate receptor therapies: Advances in clinical trials for CNS-related diseases. Neuropharmacology 2017, 113, 597–607. [Google Scholar] [CrossRef] [PubMed]
- Chiba, K.; Matsuyuki, H.; Maeda, Y.; Sugahara, K. Role of sphingosine 1-phosphate receptor type 1 in lymphocyte egress from secondary lymphoid tissues and thymus. Cell. Mol. Immunol. 2006, 3, 11–19. [Google Scholar] [PubMed]
- Raza, Z.; Saleem, U.; Naureen, Z. Sphingosine 1-phosphate signaling in ischemia and reperfusion injury. Prostaglandins Other Lipid Mediat. 2020, 149, 106436. [Google Scholar] [CrossRef] [PubMed]
- Karliner, J.S. Sphingosine kinase regulation and cardioprotection. Cardiovasc. Res. 2008, 82, 184–192. [Google Scholar] [CrossRef] [PubMed]
- Sanchez, T. Sphingosine-1-Phosphate Signaling in Endothelial Disorders. Curr. Atheroscler. Rep. 2016, 18, 31. [Google Scholar] [CrossRef]
- Ding, R.; Han, J.; Tian, Y.; Guo, R.; Ma, X. Sphingosine-1-Phosphate Attenuates Lung Injury Induced by Intestinal Ischemia/Reperfusion in Mice: Role of Inducible Nitric-Oxide Synthase. Inflammation 2011, 35, 158–166. [Google Scholar] [CrossRef]
- Park, S.W.; Kim, M.; Kim, M.; D’Agati, V.D.; Lee, H.T. Sphingosine kinase 1 protects against renal ischemia–reperfusion injury in mice by sphingosine-1-phosphate1 receptor activation. Kidney Int. 2011, 80, 1315–1327. [Google Scholar] [CrossRef]
- Henry, L.; Fransolet, M.; Labied, S.; Blacher, S.; Masereel, M.-C.; Foidart, J.-M.; Noël, A.; Nisolle, M.; Munaut, C. Supplementation of transport and freezing media with anti-apoptotic drugs improves ovarian cortex survival. J. Ovarian Res. 2016, 9, 4. [Google Scholar] [CrossRef]
- Bajwa, A.; Huang, L.; Kurmaeva, E.; Gigliotti, J.C.; Ye, H.; Miller, J.; Rosin, D.L.; Lobo, P.I.; Okusa, M.D. Sphingosine 1-Phosphate Receptor 3–Deficient Dendritic Cells Modulate Splenic Responses to Ischemia-Reperfusion Injury. J. Am. Soc. Nephrol. 2015, 27, 1076–1090. [Google Scholar] [CrossRef]
- Qiang, G.-H.; Wang, Z.-X.; Ji, A.-L.; Wu, J.-Y.; Cao, Y.; Zhang, G.; Zhang, Y.-Y.; Jiang, C. Sphingosine kinase 1 knockout alleviates hepatic ischemia/reperfusion injury by attenuating inflammation and oxidative stress in mice. Hepatobiliary Pancreat. Dis. Int. 2019, 18, 255–265. [Google Scholar] [CrossRef] [PubMed]
- Scott, F.L.; Clemons, B.; Brooks, J.; Brahmachary, E.; Powell, R.; Dedman, H.; Desale, H.G.; Timony, G.; Martinborough, E.; Rosen, H.; et al. Ozanimod (RPC1063) is a potent sphingosine-1-phosphate receptor-1 (S1P1) and receptor-5 (S1P5) agonist with autoimmune disease-modifying activity. Br. J. Pharmacol. 2016, 173, 1778–1792. [Google Scholar] [CrossRef] [PubMed]
- Billich, A.; Bornancin, F.; Dévay, P.; Mechtcheriakova, D.; Urtz, N.; Baumruker, T. Phosphorylation of the Immunomodulatory Drug FTY720 by Sphingosine Kinases. J. Biol. Chem. 2003, 278, 47408–47415. [Google Scholar] [CrossRef] [PubMed]
- Radeva, M.Y.; Waschke, J. Mind the gap: Mechanisms regulating the endothelial barrier. Acta Physiol. 2017, 222, e12860. [Google Scholar] [CrossRef]
- Brinkmann, V. Sphingosine 1-phosphate receptors in health and disease: Mechanistic insights from gene deletion studies and reverse pharmacology. Pharmacol. Ther. 2007, 115, 84–105. [Google Scholar] [CrossRef]
- Olesch, C.; Ringel, C.; Brüne, B.; Weigert, A. Beyond Immune Cell Migration: The Emerging Role of the Sphingosine-1-phosphate Receptor S1PR4 as a Modulator of Innate Immune Cell Activation. Mediat. Inflamm. 2017, 2017, 1–12. [Google Scholar] [CrossRef]
- Fettel, J.; Kühn, B.; Guillen, N.A.; Sürün, D.; Peters, M.; Bauer, R.; Angioni, C.; Geisslinger, G.; Schnütgen, F.; Zu Heringdorf, D.M.; et al. Sphingosine-1-phosphate (S1P) induces potent anti-inflammatory effects in vitro and in vivo by S1P receptor 4-mediated suppression of 5-lipoxygenase activity. FASEB J. 2018, 33, 1711–1726. [Google Scholar] [CrossRef]
- Imeri, F.; Blanchard, O.; Jenni, A.; Schwalm, S.; Wünsche, C.; Živković, A.; Stark, H.; Pfeilschifter, J.; Huwiler, A. FTY720 and two novel butterfly derivatives exert a general anti-inflammatory potential by reducing immune cell adhesion to endothelial cells through activation of S1P3 and phosphoinositide 3-kinase. Naunyn-Schmiedeberg’s Arch. Pharmacol. 2015, 388, 1283–1292. [Google Scholar] [CrossRef]
- Shigematsu, T.; Wolf, R.; Granger, D.N. T-lymphocytes modulate the microvascular and inflammatory responses to intestinal ischemia-reperfusion. Microcirculation 2002, 9, 99–109. [Google Scholar] [CrossRef]
- Ebertoni, S.; Arcaro, V.; Vivo, V.; Rapalli, A.; Tognolini, M.; Cantoni, A.M.; Saccani, F.; Flammini, L.; Domenichini, G.; Ballabeni, V.; et al. Suppression of inflammatory events associated to intestinal ischemia–reperfusion by 5-HT1A blockade in mice. Pharmacol. Res. 2014, 81, 17–25. [Google Scholar] [CrossRef]
- Daniel, C.; Sartory, N.; Zahn, N.; Geisslinger, G.; Radeke, H.H.; Stein, J. FTY720 ameliorates Th1-mediated colitis in mice by directly affecting the functional activity of CD4+CD25+ regulatory T cells. J. Immunol. 2007, 178, 2458–2468. [Google Scholar] [CrossRef]
- Krawisz, J.E.; Sharon, P.; Stenson, W.F. Quantitative assay for acute intestinal inflammation based on myeloperoxidase activity. Assessment of inflammation in rat and hamster models. Gastroenterology 1984, 87, 1344–1350. [Google Scholar] [CrossRef]
- Ohkawa, H.; Ohishi, N.; Yagi, K. Assay for lipid peroxides in animal tissues by thiobarbituric acid reaction. Anal. Biochem. 1979, 95, 351–358. [Google Scholar] [CrossRef]
- Moore-Olufemi, S.D.; Kozar, R.A.; Moore, F.A.; Sato, N.; Hassoun, H.T.; Cox, C.S., Jr.; Kone, B.C. Ischemic preconditioning protects against gut dysfunction and mucosal injury after ischemia/reperfusion injury. Shock 2005, 23, 258–263. [Google Scholar]
- Schindelin, J.; Arganda-Carreras, I.; Frise, E.; Kaynig, V.; Longair, M.; Pietzsch, T.; Preibisch, S.; Rueden, C.; Saalfeld, S.; Schmid, B.; et al. Fiji: An open-source platform for biological-image analysis. Nat. Methods 2012, 9, 676–682. [Google Scholar] [CrossRef] [PubMed]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Treatment | MDA (nmol/g) |
|---|---|
| S | 304.1 ± 60.6 |
| I/R + vehicle | 686.9 ± 88.4 |
| I/R + FTY720 0.3 mg/kg | 1813.0 ± 315.6 *,# |
| I/R + FTY720 1.0 mg/kg | 525.3 ± 76.7 |
| I/R + FTY720 3.0 mg/kg | 1020.0 ± 225.8 |
| I/R + ozanimod 0.3 mg/kg | 539.1 ± 79.9 |
| I/R + ozanimod 1.0 mg/kg | 871.9 ± 253.6 |
| Treatment | Total Lymphocytes (×105) | CD4+CD8- (×105) | CD4-CD8+ (×105) |
|---|---|---|---|
| S | 25.5 ± 0.8 | 13.4 ± 2.0 | 4.9 ± 1.0 |
| I/R + vehicle | 24.0 ± 3.9 | 13.6 ± 2.0 | 4.1 ± 0.7 |
| I/R + FTY720 3.0 mg/kg | 24.3 ± 3.0 | 13.8 ± 1.7 | 3.9 ± 0.5 |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Potì, F.; Giorgio, C.; Zini, I.; Nofer, J.-R.; Vivo, V.; Palese, S.; Ballabeni, V.; Barocelli, E.; Bertoni, S. Impact of S1P Mimetics on Mesenteric Ischemia/Reperfusion Injury. Pharmaceuticals 2020, 13, 298. https://doi.org/10.3390/ph13100298
Potì F, Giorgio C, Zini I, Nofer J-R, Vivo V, Palese S, Ballabeni V, Barocelli E, Bertoni S. Impact of S1P Mimetics on Mesenteric Ischemia/Reperfusion Injury. Pharmaceuticals. 2020; 13(10):298. https://doi.org/10.3390/ph13100298
Chicago/Turabian StylePotì, Francesco, Carmine Giorgio, Irene Zini, Jerzy-Roch Nofer, Valentina Vivo, Simone Palese, Vigilio Ballabeni, Elisabetta Barocelli, and Simona Bertoni. 2020. "Impact of S1P Mimetics on Mesenteric Ischemia/Reperfusion Injury" Pharmaceuticals 13, no. 10: 298. https://doi.org/10.3390/ph13100298
APA StylePotì, F., Giorgio, C., Zini, I., Nofer, J.-R., Vivo, V., Palese, S., Ballabeni, V., Barocelli, E., & Bertoni, S. (2020). Impact of S1P Mimetics on Mesenteric Ischemia/Reperfusion Injury. Pharmaceuticals, 13(10), 298. https://doi.org/10.3390/ph13100298