
Extracting Sensory Preferability from Motor Streams


Abstract
1. Introduction
1.1. Reafference Principle and Sensory Input in Motor Stream
“Can we detect sensory variations within motor streams?”
“Is it possible to separate sensory and motor components automatically?”
“Could researchers develop methods to extract sensory information from continuous motor data?”
1.2. Interfaces in Need of “Understanding” the Preferability of Our Peripheral Nervous System
1.3. Baseline Research
1.4. Aims and Contributions
2. Experimental and Computational Methods
2.1. Participants
2.2. Experimental Task
Sensory Stimuli
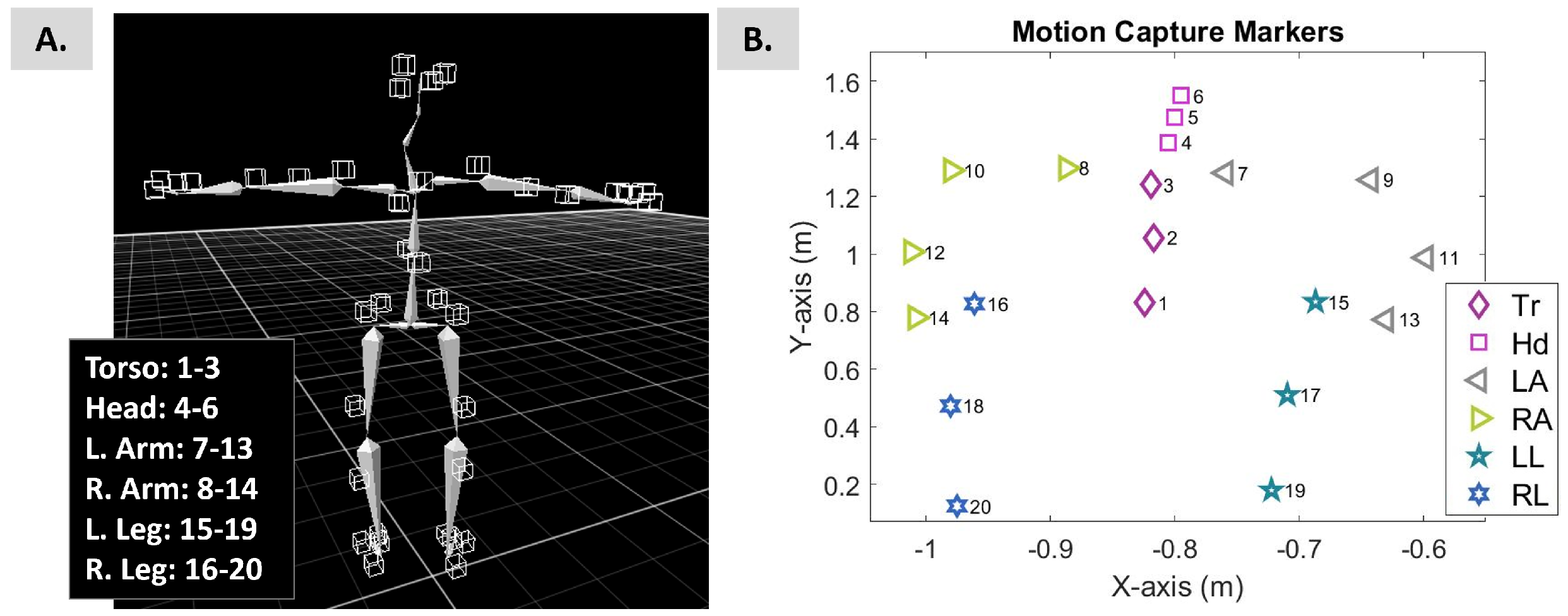
2.3. Instrumentation
2.4. Data Processing
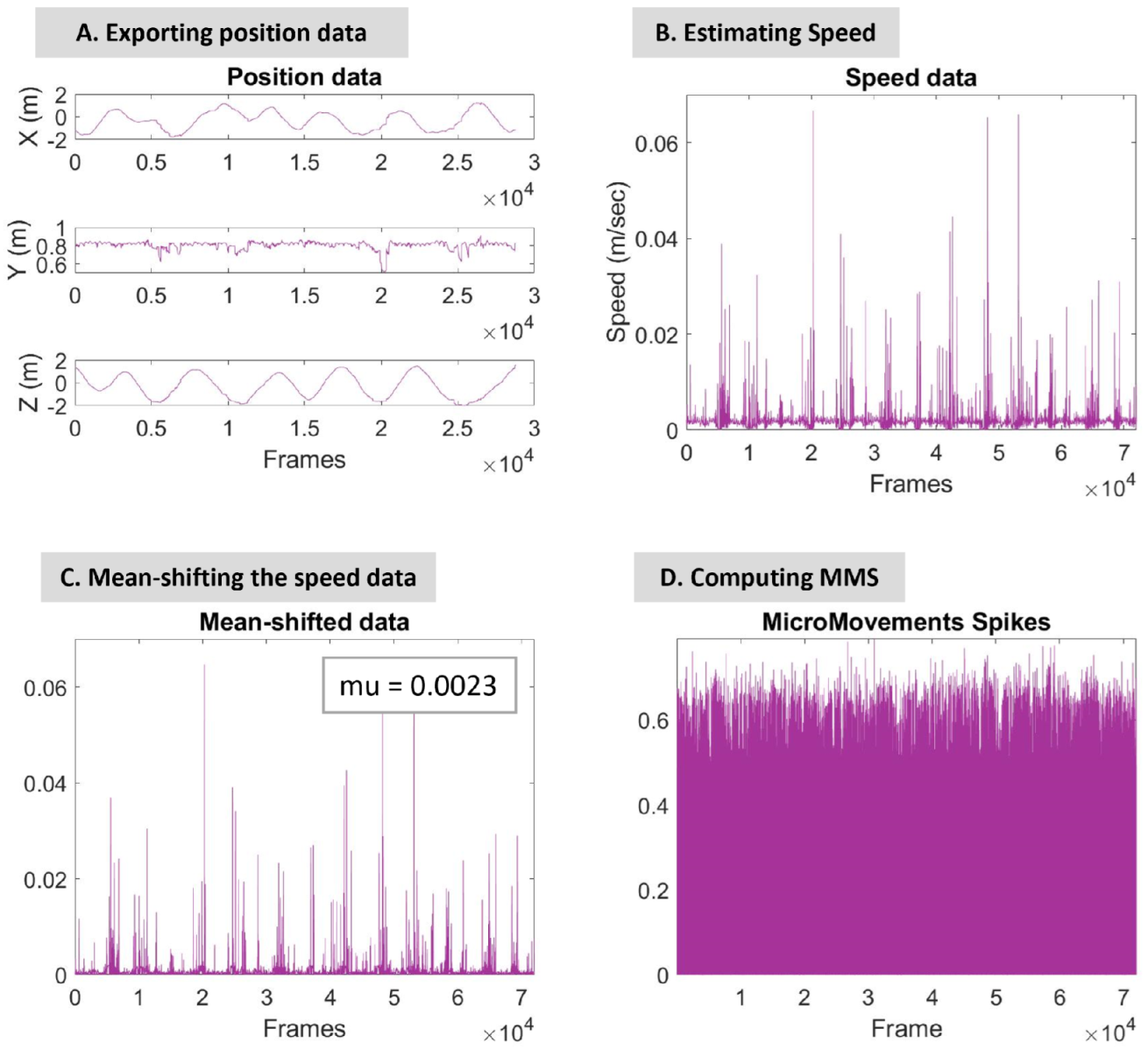
2.4.1. Extraction and Computation of Motor Stream
2.4.2. Micro-Movement Spikes
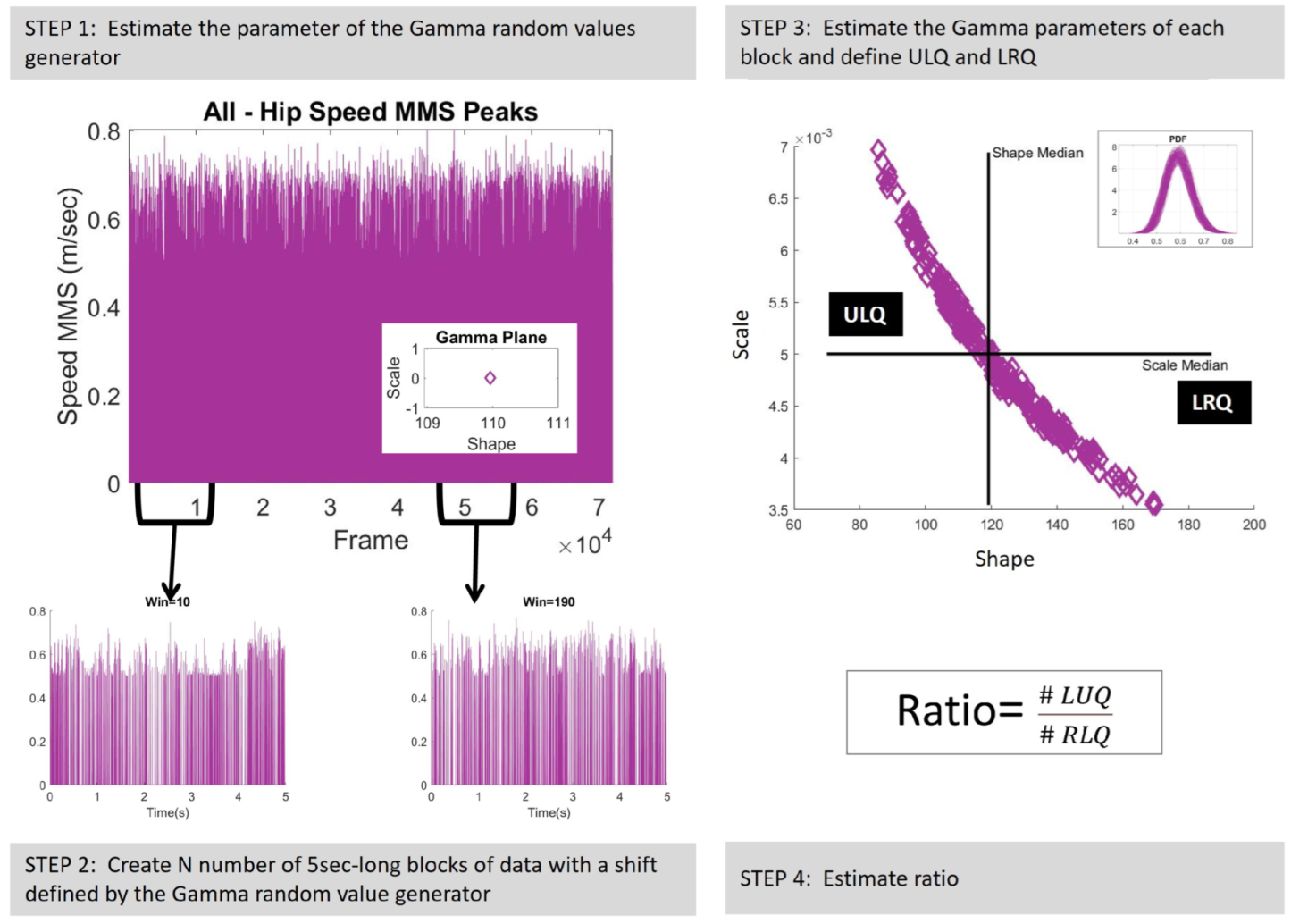
2.4.3. Micro-Movement Spikes Represented by Continuous Gamma Process
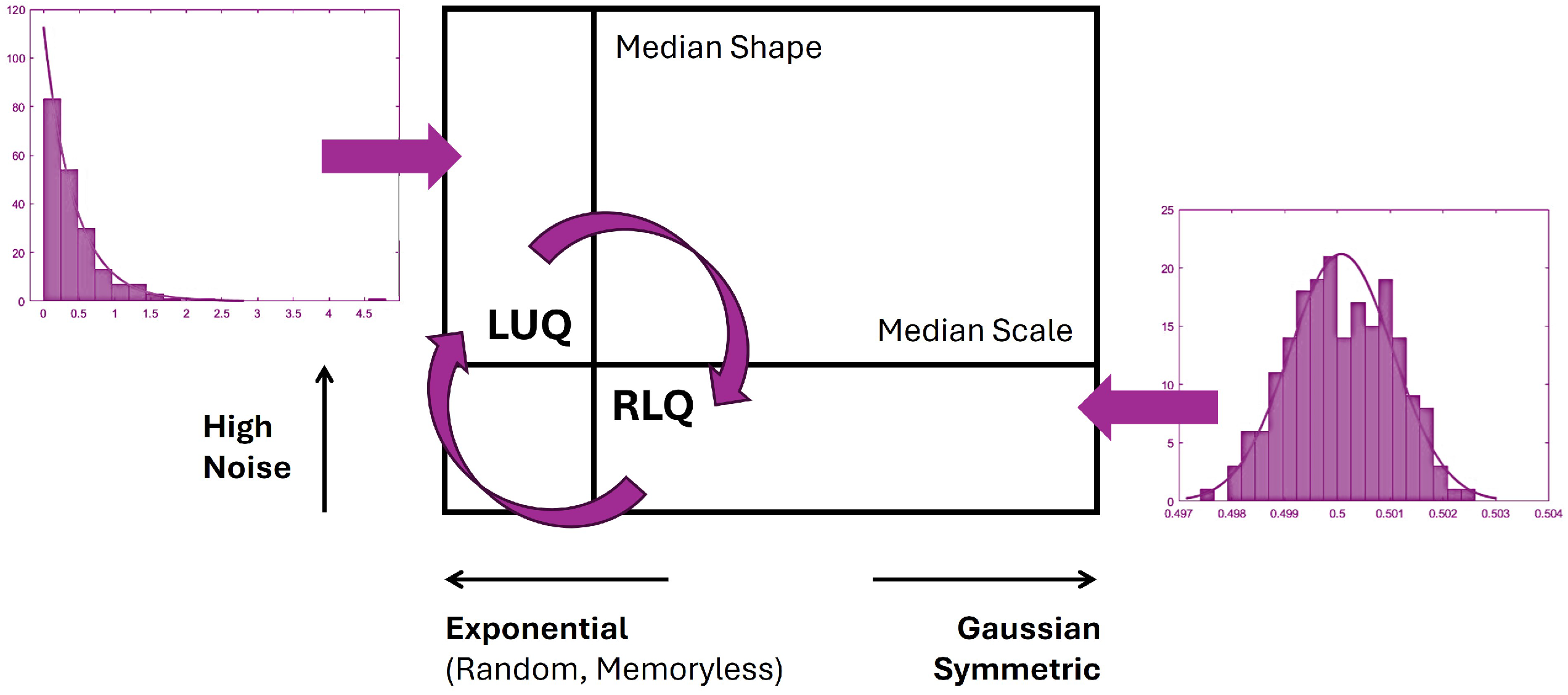
2.4.4. Tracking NSR and Predictive Regimes
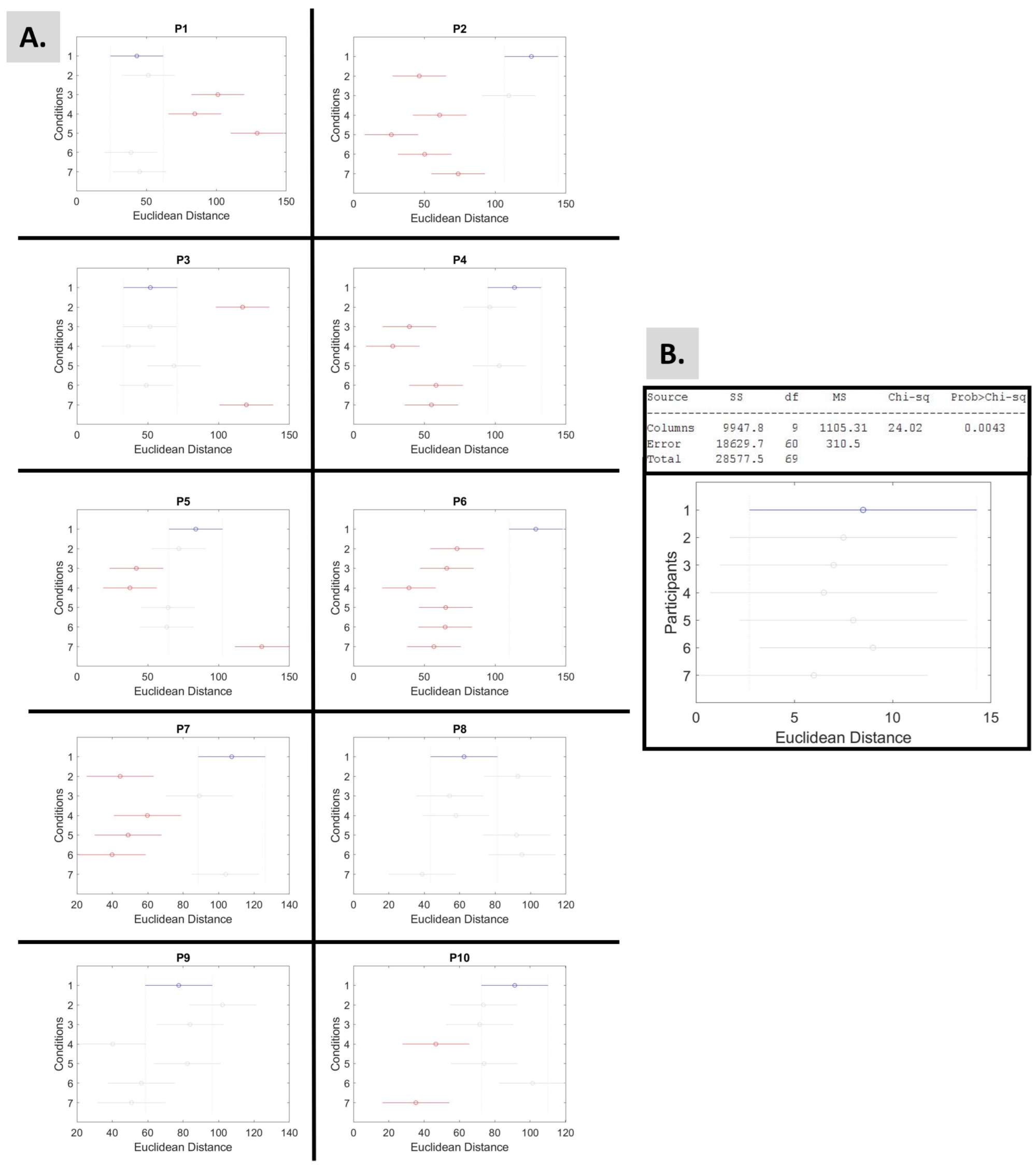
2.4.5. Euclidean Distance of the Extreme Signatures
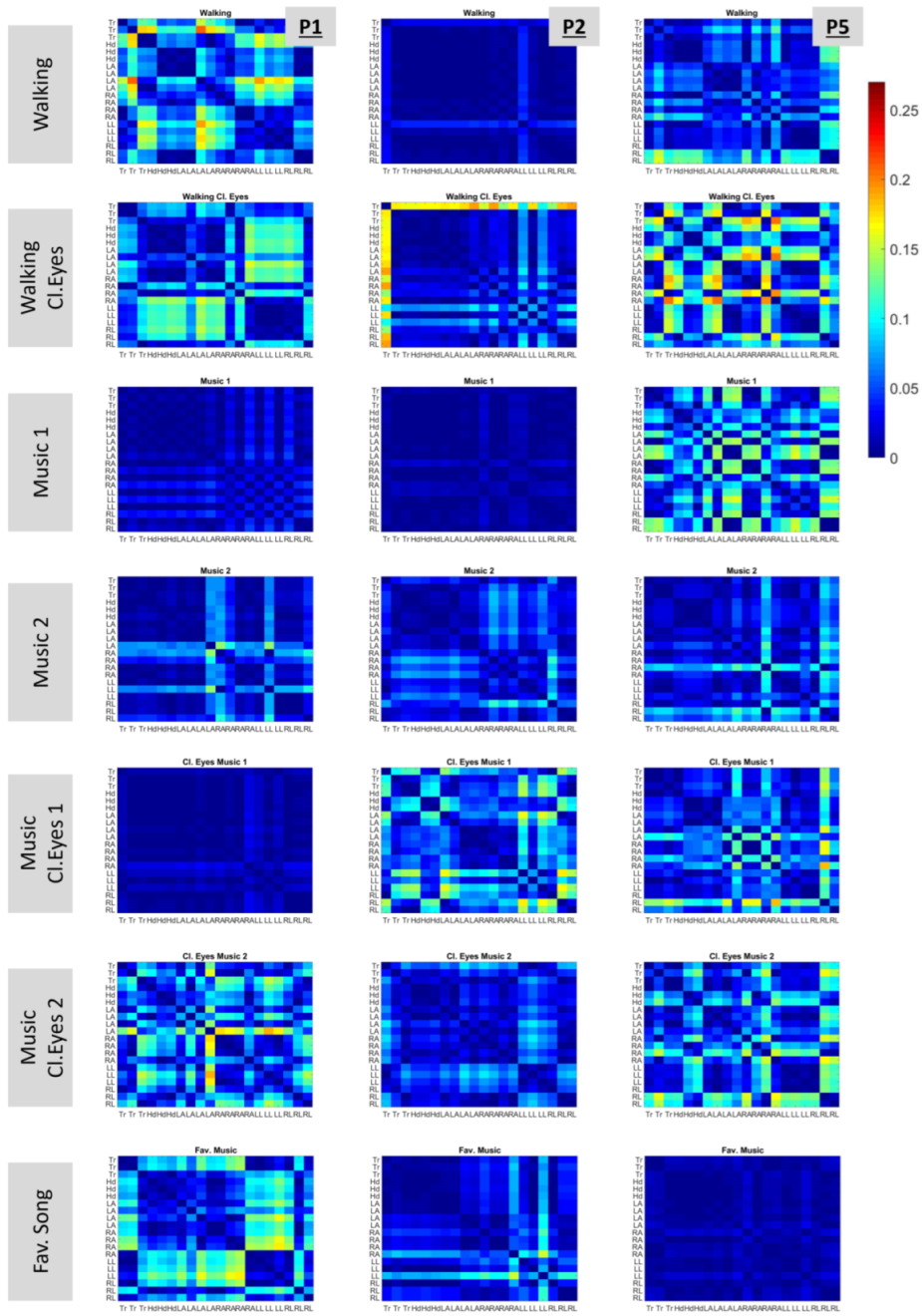
2.4.6. Rate of Change in Stochastic Transitions
2.4.7. Cumulative Information
2.4.8. Statistical Significance
3. Results
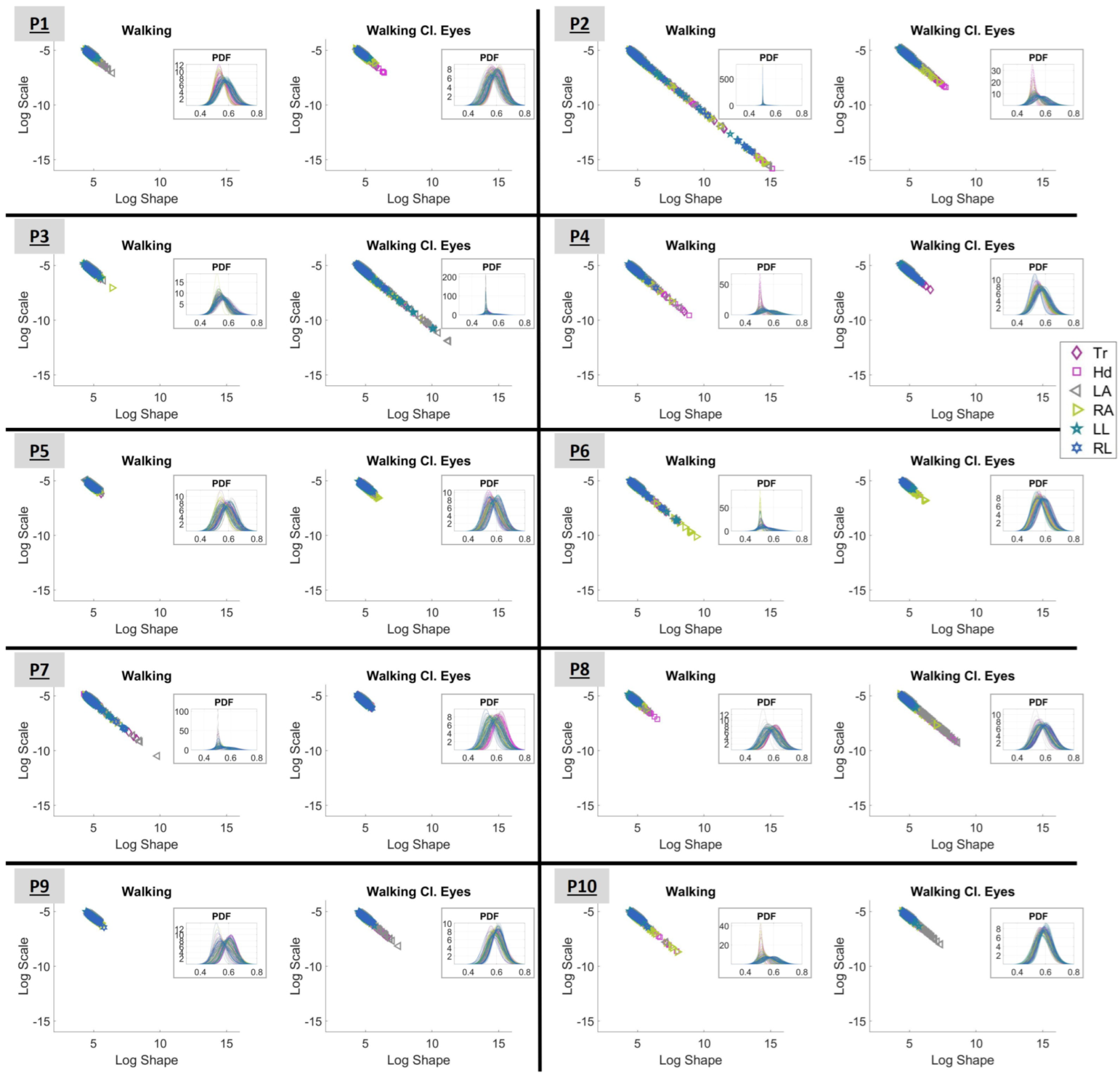
3.1. Investigating the Gamma Stochastic Signatures in Logarithmic Space
3.2. Observing Variations Within the Kinematic Chain of Each Participant
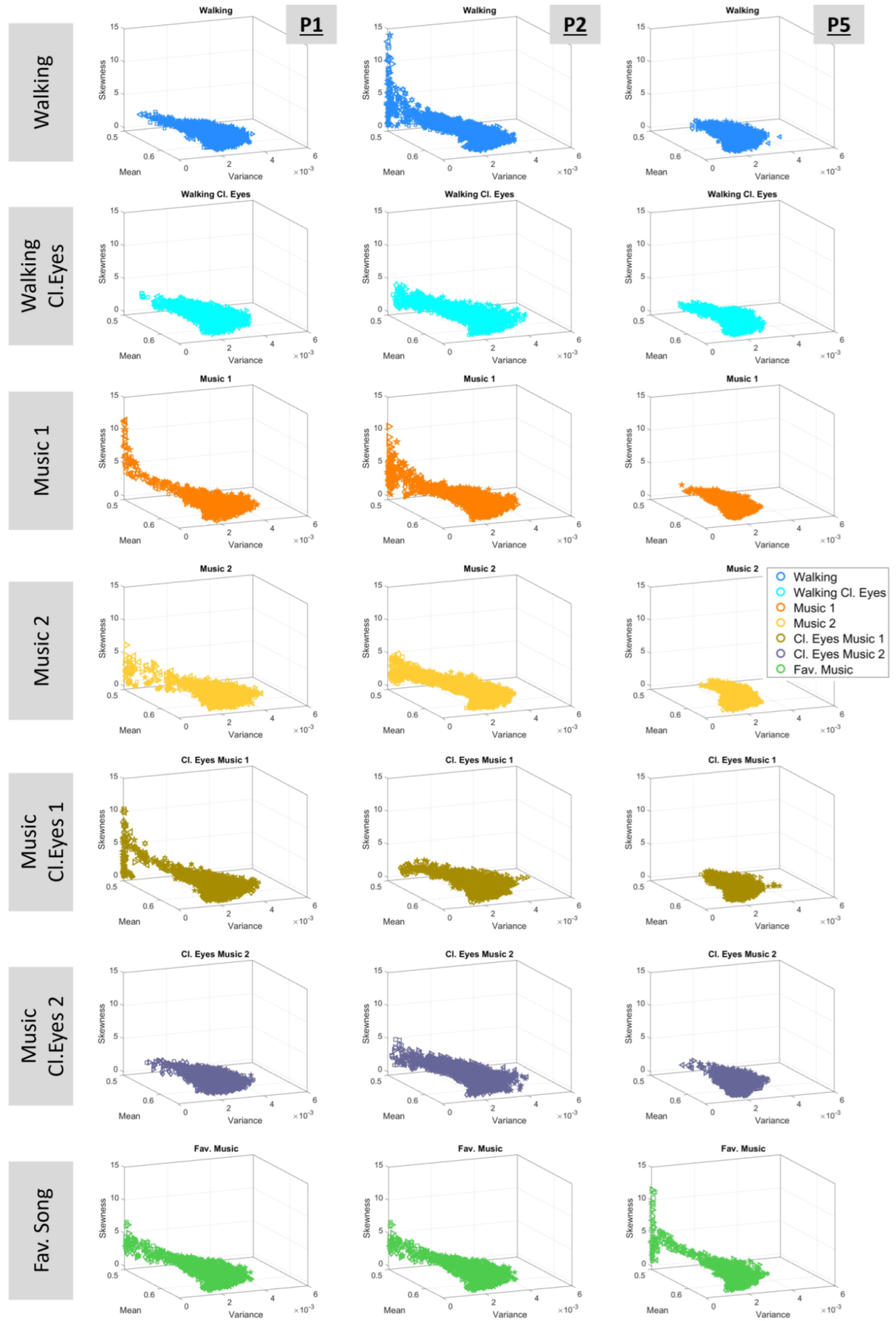
3.3. Expanding Investigations to the Four Moments of the Gamma Distribution
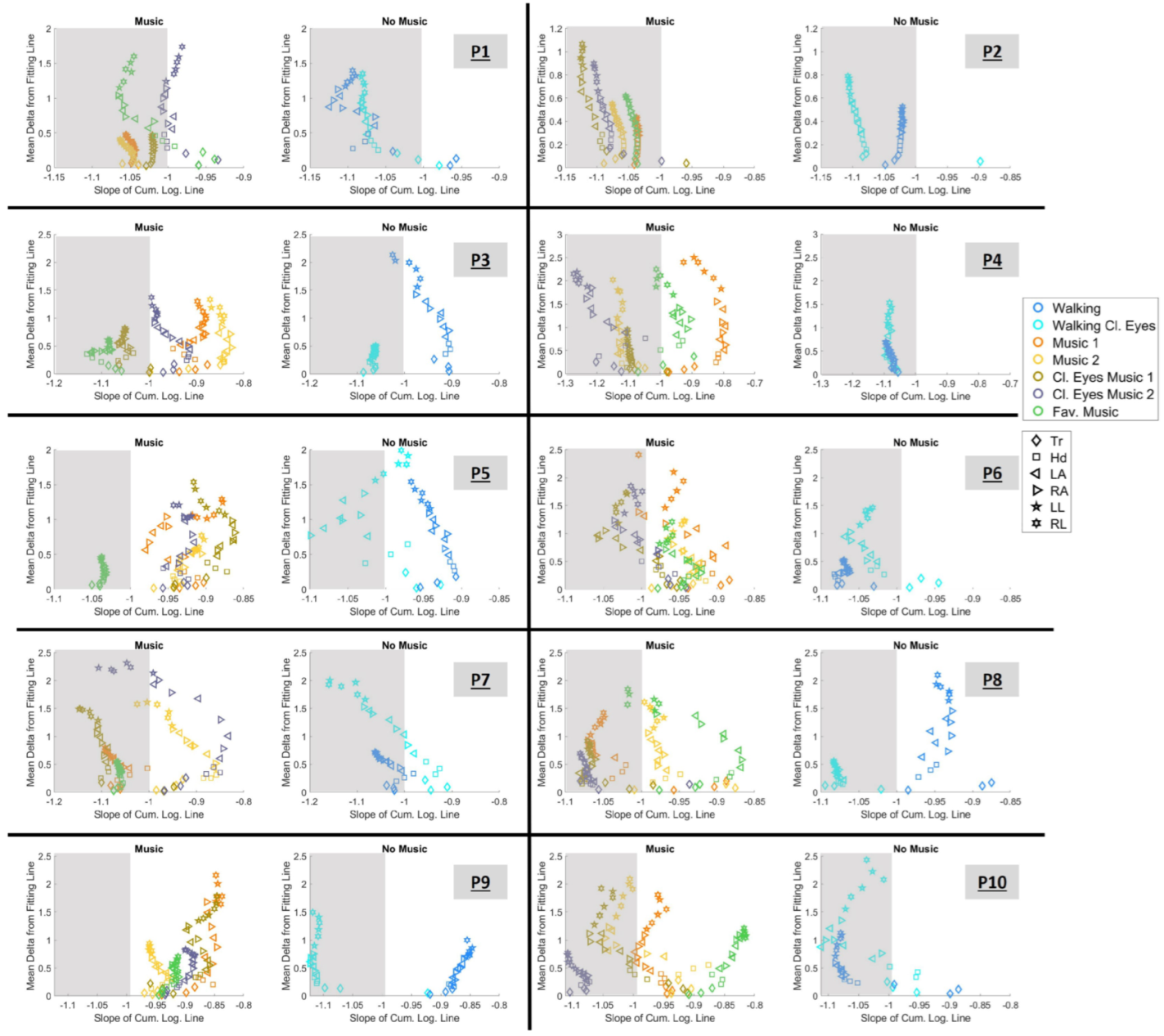
3.4. Exploring the Cumulative Space
3.4.1. Choosing Sensory Preferability Through Parameter Space 1
- P1: “Music 1”, “Music 2”, “Music Cl. Eyes 1”, and “Fav. music”
- P2: for all conditions
- P3: “Music Cl. Eyes 1” and “Fav. music”
- P4: “Music 2”, “Music Cl. Eyes 1”, and “Music Cl. Eyes 2”
- P5: “Fav. music”
- P6: “Cl. eyes 1” and “Cl. eyes 2”
- P7: “Music 1”, “Music Cl. Eyes 1”, and “Fav. music”
- P8: “Music 1”, “Cl. eyes 1”, “Cl. eyes 2”
- P9: for no condition
- P10: “Music 1”, “Cl. eyes 2”, and “Cl. eyes 2”.
3.4.2. Choosing Sensory Preferability Through Parameter Space 2
- P1: during condition “Fav. music”
- P2: during condition “Cl. eyes 2”
- P3: during condition “Fav. music”
- P4: during condition “Cl. eyes 2”
- P6: during condition “Cl. eyes 1”
- P7: during condition “Fav. music”
- P8: during condition “Cl. eyes 2”.
4. Discussion and Possible Applications
4.1. Metrics, Parameter Space, and Findings
4.2. Key Elements of the Study Design
4.3. Affecting Temporal and Spatial Features of the Motor Stream
4.4. Theory and Possible Applications
5. Conclusions
Supplementary Materials
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- King, B.A.; Paulson, L.D. Motion capture moves into new realms. IEEE Comput. 2007, 40, 13–16. [Google Scholar]
- Pasquet, M.O.; Tihy, M.; Gourgeon, A.; Pompili, M.N.; Godsil, B.P.; Léna, C.; Dugué, G.P. Wireless inertial measurement of head kinematics in freely-moving rats. Sci. Rep. 2016, 6, 35689. [Google Scholar] [PubMed]
- Debauche, O.; Mahmoudi, S.; Andriamandroso, A.L.H.; Manneback, P.; Bindelle, J.; Lebeau, F. Cloud services integration for farm animals’ behavior studies based on smartphones as activity sensors. J. Ambient Intell. Humaniz. Comput. 2019, 10, 4651–4662. [Google Scholar]
- Kutilek, P.; Socha, V.; Schlenker, J.; Skoda, D.; Hybl, J.; Cerny, R.; Frynta, D.; Landova, E.; Haskova, T.; Kurali, A. System for measuring movement response of small animals to changes in their orientation. In Proceedings of the 2015 International Conference on Applied Electronics (AE), Pilsen, Czech Republic, 8–9 September 2015; pp. 139–144. [Google Scholar]
- Ewert, J.P. Reafference Principle. In Encyclopedia of Sciences and Religions; Runehov, A.L.C., Oviedo, L., Eds.; Springer: Dordrecht, The Netherlands, 2013; p. 1950. [Google Scholar]
- Reafference principle. In APA Dictionary of Psychology; American Psychological Association: Washington, DC, USA.
- Ito, M. Cerebellar circuitry as a neuronal machine. Prog. Neurobiol. 2021, 196, 101907. [Google Scholar] [CrossRef]
- Shadmehr, R.; Krakauer, J.W. A computational neuroanatomy for motor control. Exp. Brain Res. 2020, 238, 1509–1531. [Google Scholar] [CrossRef]
- Wolpert, D.M.; Miall, R.C.; Kawato, M. Internal models in the cerebellum. Trends Cogn. Sci. 1998, 2, 338–347. [Google Scholar] [PubMed]
- Kawato, M.; Wolpert, D. Internal models for motor control. Sens. Guid. Mov. 1998, 218, 291–307. [Google Scholar]
- Berntson, G.G.; Cacioppo, J.T. Handbook of Neuroscience for the Behavioral Sciences; John Wiley & Sons: Hoboken, NJ, USA, 2009; Volume 2. [Google Scholar]
- López-Larraz, E.; Trincado-Alonso, F.; Rajasekaran, V.; Pérez-Nombela, S.; Del-Ama, A.J.; Aranda, J.; Minguez, J.; Gil-Agudo, A.; Montesano, L. Control of an ambulatory exoskeleton with a brain–machine interface for spinal cord injury gait rehabilitation. Front. Neurosci. 2016, 10, 359. [Google Scholar]
- Benabid, A.L.; Costecalde, T.; Eliseyev, A.; Charvet, G.; Verney, A.; Karakas, S.; Foerster, M.; Lambert, A.; Morinière, B.; Abroug, N.; et al. An exoskeleton controlled by an epidural wireless brain–machine interface in a tetraplegic patient: A proof-of-concept demonstration. Lancet Neurol. 2019, 18, 1112–1122. [Google Scholar]
- Tang, Z.; Sun, S.; Zhang, S.; Chen, Y.; Li, C.; Chen, S. A brain-machine interface based on ERD/ERS for an upper-limb exoskeleton control. Sensors 2016, 16, 2050. [Google Scholar] [CrossRef]
- Nef, T.; Mihelj, M.; Kiefer, G.; Perndl, C.; Muller, R.; Riener, R. ARMin-Exoskeleton for arm therapy in stroke patients. In Proceedings of the 2007 IEEE 10th International Conference on Rehabilitation Robotics, Noordwijk, The Netherlands, 12–15 June 2007; pp. 68–74. [Google Scholar]
- Balasubramanian, S.; Wei, R.; Perez, M.; Shepard, B.; Koeneman, E.; Koeneman, J.; He, J. RUPERT: An exoskeleton robot for assisting rehabilitation of arm functions. In Proceedings of the 2008 Virtual Rehabilitation, Vancouver, BC, Canada, 25–27 August 2008; pp. 163–167. [Google Scholar]
- Kalampratsidou, V.; Kemper, S.; Torres, E. Real-Time Proxy-Control of Re-Parameterized Peripheral Signals using a Close-Loop Interface. J. Vis. Exp 2021, 171, e61943. [Google Scholar]
- Kalampratsidou, V.; Torres, E.B. Body-brain-avatar interface: A tool to study sensory-motor integration and neuroplasticity. In Proceedings of the International Symposium on Movement and Computing, MOCO, London, UK, 28–30 June 2017; Volume 17. [Google Scholar]
- Kalampratsidou, V.; Zavorskas, M.; Albano, J.; Kemper, S.; Torres, E.B. Dance from the heart: A dance performance of sounds led by the dancer’s heart. In Proceedings of the Sixth International Symposium on Movement and Computing, Tempe, AZ, USA, 10–12 October 2019. [Google Scholar]
- Kalampratsidou, V.; Torres, E.B. Sonification of heart rate variability can entrain bodies in motion. In Proceedings of the 7th International Conference on Movement and Computing, Virtual, 15–17 July 2020; pp. 1–8. [Google Scholar]
- Kalampratsidou, V.; Kemper, S.; Torres, B.E. Real time streaming and closed loop co-adaptive interface to steer multi-layered nervous systems performance. In Proceedings of the 48th Annual Meeting of Society for Neuroscience, San Diego, CA, USA, 3–7 November 2018. [Google Scholar]
- Kalampratsidou, V. Co-Adaptive Multimodal Interface Guided by Real-Time Multisensory Stochastic Feedback. Ph.D. Thesis, Rutgers University-School of Graduate Studies, New Brunswick, NJ, USA, 2018. [Google Scholar]
- Torres, E.B.; Brincker, M.; Isenhower, R.W.; Yanovich, P.; Stigler, K.A.; Nurnberger, J.I.; Metaxas, D.N.; José, J.V. Autism: The micro-movement perspective. Front. Integr. Neurosci. 2013, 7, 32. [Google Scholar] [CrossRef] [PubMed]
- Torres, E.; Jose, J. Novel Diagnostic Tool to Quantify Signatures of Movement in Subjects with Neurobiological Disorders, Autism and Autism Spectrum Disorders; US Patent Application; Office of Technology Commercialization, Rutgers, The State University of New Jersey: New Brunswick, NJ, USA, 2012. [Google Scholar]
- Torres, E.B. Rethinking the study of volition for clinical use. In Progress in Motor Control; Springer: Cham, Switzerland, 2016; pp. 229–254. [Google Scholar]
- Torres, E.B.; Nguyen, J.; Mistry, S.; Whyatt, C.; Kalampratsidou, V.; Kolevzon, A. Characterization of the statistical signatures of micro-movements underlying natural gait patterns in children with Phelan McDermid syndrome: Towards precision-phenotyping of behavior in ASD. Front. Integr. Neurosci. 2016, 10, 22. [Google Scholar] [PubMed]
- Wu, D.; Torres, E.; Nguyen, J.; Mistry, S.; Whyatt, C.; Kalampratsidou, V.; Kolevzon, A.; Jose, J. How doing a dynamical analysis of gait movement may provide information about Autism. In Bulletin of the American Physical Society; American Physical Society (APS): Washington, DC, USA, 2017; Volume 62. [Google Scholar]
- Kalampratsidou, V.; Torres, E.B. Outcome measures of deliberate and spontaneous motions. In Proceedings of the International Symposium on Movement and Computing, Thessaloniki, Greece, 5–6 July 2016; p. 9. [Google Scholar]
- Kalabratsidou, V.; Torres, E.B. Peripheral Network Connectivity Analyses for the Real-Time Tracking of Coupled Bodies in Motion. Sensors 2018, 18, 3117. [Google Scholar] [CrossRef] [PubMed]
- Kalampratsidou, V.; Torres, E.B. Bodily signals entrainment in the presence of music. In Proceedings of the International Conference on Movement and Computing, Tempe, AZ, USA, 10–12 October 2019; pp. 1–8. [Google Scholar]
- Lleonart, J.; Salat, J.; Torres, G.J. Removing allometric effects of body size in morphological analysis. J. Theor. Biol. 2000, 205, 85–93. [Google Scholar]
- Kozłowski, J.; Konarzewski, M. Is West, Brown and Enquist’s model of allometric scaling mathematically correct and biologically relevant? Funct. Ecol. 2004, 18, 283–289. [Google Scholar] [CrossRef]
- Torres, E.B. Two classes of movements in motor control. Exp. Brain Res. 2011, 215, 269–283. [Google Scholar]
- Torres, E.B. Signatures of movement variability anticipate hand speed according to levels of intent. Behav. Brain Funct. 2013, 9, 10. [Google Scholar] [CrossRef]
- Torres, E.B.; Smith, B.; Mistry, S.; Brincker, M.; Whyatt, C. Neonatal diagnostics: Toward dynamic growth charts of neuromotor control. Front. Pediatr. 2016, 4, 121. [Google Scholar]
- Wolpert, D.M.; Flanagan, J.R. Computations underlying predictive control. Nat. Neurosci. 2016, 19, 369–377. [Google Scholar] [CrossRef]
- Herzfeld, D.J.; Vaswani, P.A.; Marko, M.K.; Shadmehr, R. A memory of errors in sensorimotor learning. Science 2014, 345, 1349–1353. [Google Scholar] [CrossRef] [PubMed]
- Dhawale, A.K.; Smith, M.A.; Ölveczky, B.P. The role of variability in motor learning. Annu. Rev. Neurosci. 2017, 40, 479–498. [Google Scholar] [CrossRef] [PubMed]
- Patel, R.; Park, H.; Bonato, P.; Chan, L.; Rodgers, M. A review of wearable sensors for monitoring human motion. Sensors 2020, 20, 586. [Google Scholar] [CrossRef]
- López-Moliner, J.; Coats, R.O. Perceptual-motor recalibration and real-time adaptation: A review. Neurosci. Biobehav. Rev. 2021, 125, 13–26. [Google Scholar] [CrossRef]




{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Condition | Description | Duration |
|---|---|---|
| # 1 | Walking | 5 min |
| # 2 | Walking Cl. Eyes | 5 min |
| Walking Open Eyes & Music | 5 min | |
| # 3 | Music 1 | 2.17 min |
| # 4 | Music 2 | 2.43 min |
| Walking Closed Eyes & Music | 5 min | |
| # 5 | Music Cl. Eyes 1 | 2.17 min |
| # 6 | Music Cl. Eyes 2 | 2.43 min |
| # 7 | Fav. Music | 5 min |
| Walking | Cl.Eyes | Music 1 | Music 2 | M.Cl.E.1 | M.Cl.E.2 | Fav.M. | Mean | |
|---|---|---|---|---|---|---|---|---|
| P1 | 2.6153 | 3.0803 | 8.3027 | 9.9585 | ||||
| P2 | 15.4041 | 5.0433 | 9.8127 | 7.5092 | 3.8996 | 6.1839 | 7.9351 | 7.96 |
| P3 | 3.0795 | 9.9164 | 3.4311 | 2.2010 | 4.0664 | 2.7714 | 10.8285 | 5.18 |
| P4 | 6.5625 | 3.3152 | 1.8821 | 1.3742 | 4.5432 | 1.8562 | 2.3375 | 3.12 |
| P5 | 1.7314 | 2.1357 | 1.5971 | 1.5458 | 1.7745 | 1.9206 | 12.8226 | 3.36 |
| P6 | 7.1734 | 2.5319 | 2.2215 | 1.3054 | 2.2047 | 1.7081 | 1.9690 | 2.73 |
| P7 | 7.9360 | 1.6930 | 6.1193 | 2.3235 | 3.3887 | 2.1550 | 8.3185 | 4.56 |
| P8 | 3.1501 | 6.2031 | 3.1024 | 4.2011 | 5.1410 | 6.3765 | 1.6493 | 4.26 |
| P9 | 1.9820 | 4.3195 | 2.5957 | 1.4136 | 2.2465 | 1.5484 | 1.5303 | 2.23 |
| P10 | 5.1849 | 4.2032 | 2.9588 | 2.3339 | 4.1045 | 6.4769 | 1.7299 | 3.86 |
| KW Test | P1 | P2 | P3 | P4 | P5 |
| p-values | 3.78 | 1.08 | 4.95 | 1.29 | 3.59 |
| P6 | P7 | P8 | P9 | P10 | |
| p-values | 2.25 | 2.54 | 1.59 | 4.39 | 6.99 |
| KW Test | P1 | P2 | P3 | P4 | P5 |
| Mean | 8.02 | 8.12 | 1.64 | 0 | 1.98 |
| Variance | 2.79 | 1.33 | 4.86 | 8.6 | 2.60 |
| Skewness | 7.81 | 6.18 | 2.48 | 0 | 2.62 |
| Kurtosis | 5.20 | 2.56 | 6.61 | 1.38 | 1.35 |
| P6 | P7 | P8 | P9 | P10 | |
| Mean | 0 | 2.29 | 0 | 0 | 0 |
| Variance | 4.05 | 2.09 | 1.73 | 1.88 | 2.88 |
| Skewness | 0 | 7.97 | 0 | 0 | 0 |
| Kurtosis | 6.83 | 1.33 | 4.82 | 3.59 | 3.64 |
| Wilcoxon Rank-Sum Test | Ratio | Cum. Log. Gamma Slope |
|---|---|---|
| Music vs. No Music | >0.01 | |
| Op.Eyes vs. Cl.Eyes | >0.01 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the author. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kalampratsidou, V. Extracting Sensory Preferability from Motor Streams. Sensors 2025, 25, 2087. https://doi.org/10.3390/s25072087
Kalampratsidou V. Extracting Sensory Preferability from Motor Streams. Sensors. 2025; 25(7):2087. https://doi.org/10.3390/s25072087
Chicago/Turabian StyleKalampratsidou, Vilelmini. 2025. "Extracting Sensory Preferability from Motor Streams" Sensors 25, no. 7: 2087. https://doi.org/10.3390/s25072087
APA StyleKalampratsidou, V. (2025). Extracting Sensory Preferability from Motor Streams. Sensors, 25(7), 2087. https://doi.org/10.3390/s25072087