
Antagonist Activation Measurement in Triceps Surae Using High-Density and Bipolar Surface EMG in Chronic Hemiparesis

 , , , ,
, , , , 

Abstract
1. Introduction
2. Materials and Methods
2.1. Participants
2.2. Experimental Protocol
2.3. Positioning of Surface Electrodes and EMG Recording
- -
- A matrix of 64 electrodes (13 × 5 with a missing corner electrode and an 8-mm inter-electrode distance) placed over the GM. The grid was centered halfway between the medial and lateral boundaries of the muscle, with the top row being placed about 2 cm distally to the popliteal fossa.
- -
- Two arrays of 8 electrodes (a 5-mm inter-electrode distance) were placed over the medial and lateral portions of SO. Each array was rotated ~45° outward with respect to the leg axis and centered 30 mm distally to the GM myotendinous junctions.
2.4. EMG Analysis
- -
- CAN measurement from bipolar EMG.
- -
- CAN measurement from HD-EMG.
- (i)
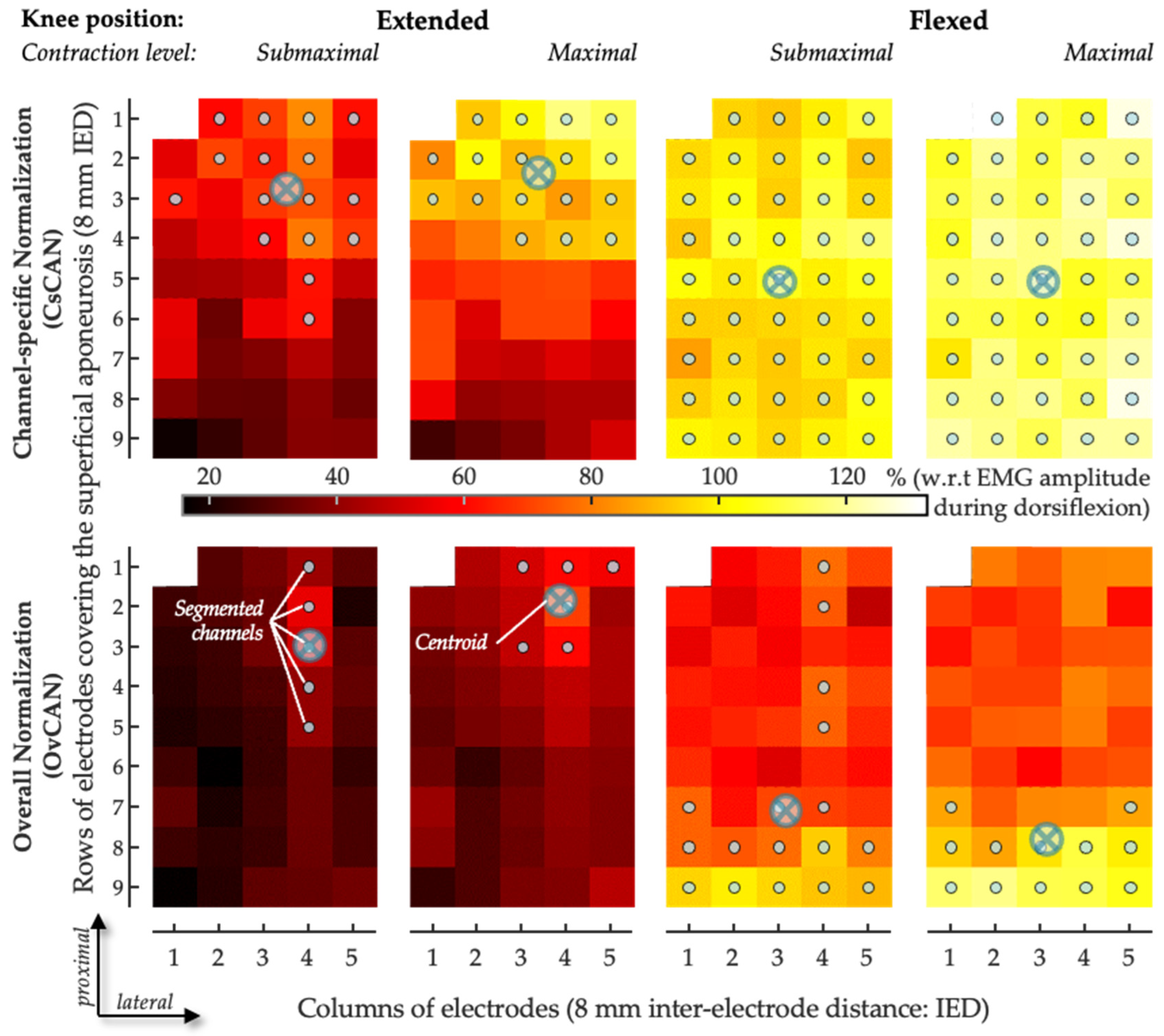
- The ratio of the RMS value was obtained for a given muscle, acting as an antagonist, and the RMS value obtained for the same muscle while it acted as an agonist at a maximal contraction level on a channel-specific basis—a procedure named channel-specific normalization. Using this approach, we computed the channel-specific CAN (CsCAN) in the GM and SO (CsCANGM and CsCANSO).
- (ii)
- The ratio between RMS values was obtained using the maximal RMS value across EMGs in the grid in the denominator—a procedure named overall normalization. Using this approach, we computed the overall CAN (OvCAN) for the two muscles (OvCANGM and OvCANSO).
- -
- Spatial distribution of antagonist activation from HD-EMG.
2.5. Statistics
3. Results
3.1. Participants
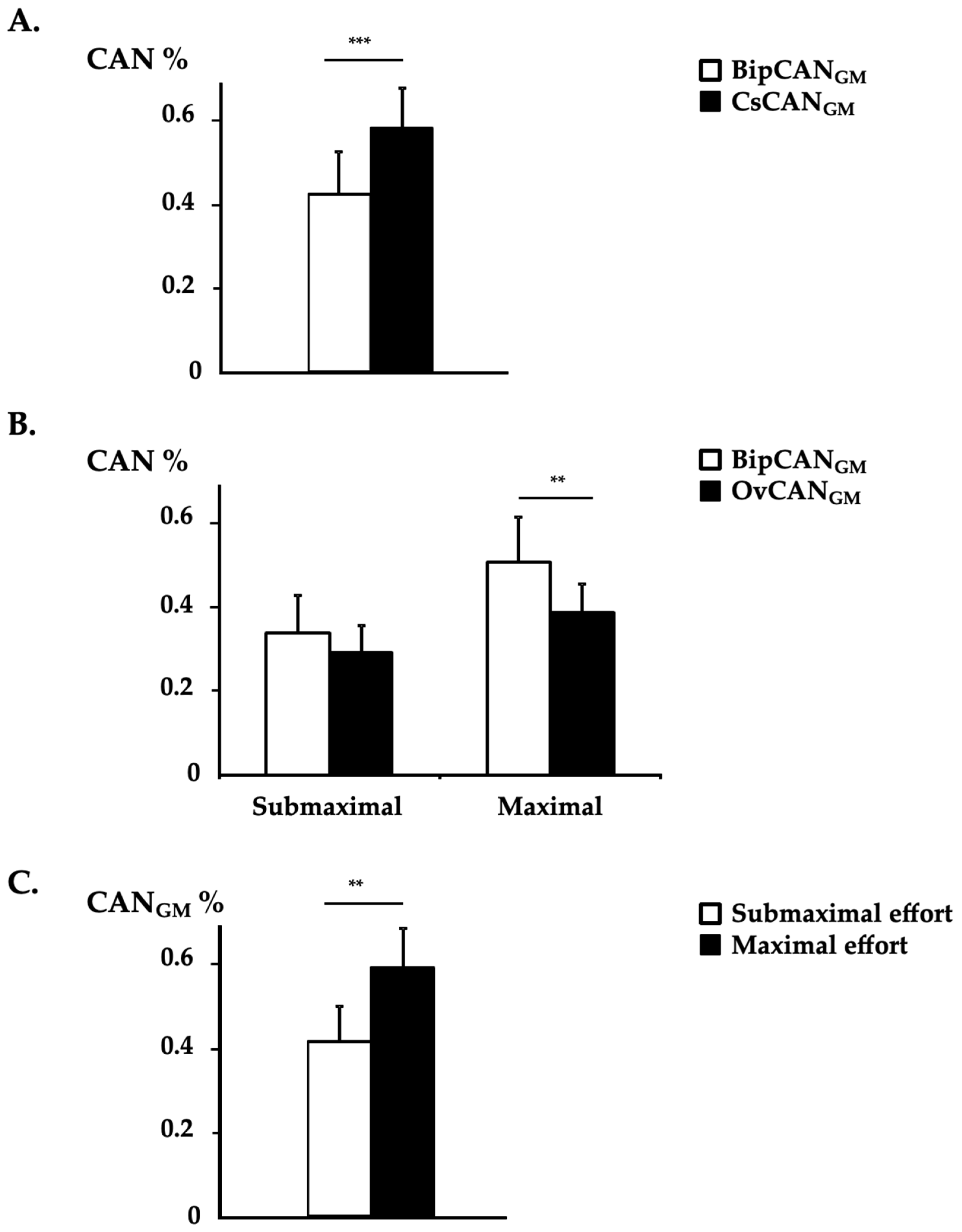
3.2. CAN Measurement in GM Using Bipolar EMG vs. HD-EMG
3.3. CAN Distribution in GM and SO
4. Discussion
4.1. Why Considering Two Normalization Procedures in HD-EMG Process?
4.2. Limitations and Differences in Estimations of Antagonist Activation: Detection Modes and Normalization Procedures
4.3. Antagonist Activation Equally Estimated in Medial and Lateral Portions of Soleus
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Knutsson, E.; Mårtensson, A. Dynamic motor capacity in spastic paresis and its relation to primer mover dysfunction, spastic reflexes and antagonistic coactivation. Scand. J. Rehab. Med. 1980, 12, 93–106. [Google Scholar]
- Berger, W.; Horstmann, G.; Dietz, V. Tension development and muscle activation in the leg during gait in spastic hemiparesis: Independence of muscle hypertonia and exaggerated stretch reflexes. J. Neurol. Neurosurg. Psychiatry 1984, 47, 1029–1033. [Google Scholar] [CrossRef] [PubMed]
- Hammond, M.C.; Fitts, S.S.; Kraft, G.H.; Nutter, P.B.; Trotter, M.J.; Robinson, L.M. Co-contraction in the hemiparetic forearm: Quantitative EMG evaluation. Arch. Phys. Med. Rehabil. 1988, 69, 348–351. [Google Scholar] [PubMed]
- Ghédira, M.; Pradines, M.; Mardale, V.; Gracies, J.M.; Bayle, N.; Hutin, E. Quantified clinical measures linked to ambulation speed in hemiparesis. Top. Stroke Rehabil. 2022, 29, 411–422. [Google Scholar] [CrossRef] [PubMed]
- Souissi, H.; Zory, R.; Bredin, J.; Gerus, P. Comparison of methodologies to assess muscle co-contraction during gait. J. Biomech. 2017, 57, 141–145. [Google Scholar] [CrossRef] [PubMed]
- Vinti, M.; Couillandre, A.; Hausselle, J.; Bayle, N.; Primerano, A.; Merlo, A.; Hutin, E.; Gracies, J.M. Influence of effort intensity and gastrocnemius stretch on co-contraction and torque production in the healthy and paretic ankle. Clin. Neurophysiol. 2013, 124, 528–535. [Google Scholar] [CrossRef]
- Ghédira, M.; Albertsen, I.M.; Mardale, V.; Loche, C.M.; Vinti, M.; Gracies, J.M.; Bayle, N.; Hutin, E. Agonist and antagonist activation at the ankle monitored along the swing phase in hemiparetic gait. Clin. Biomech. 2021, 89, 105459. [Google Scholar] [CrossRef] [PubMed]
- Hermens, H.J.; Freriks, B.; Disselhorst-Klug, C.; Rau, G. Development of recommendations for SEMG sensors and sensor placement procedures. J. Electromyogr. Kinesiol. 2000, 10, 361–374. [Google Scholar] [CrossRef]
- Merletti, R.; Hermens, H. Introduction to the special issue on the SENIAM European Concerted Action. J. Electromyogr. Kinesiol. 2000, 10, 283–286. [Google Scholar] [CrossRef]
- Vigotsky, A.D.; Halperin, I.; Lehman, G.J.; Trajano, G.S.; Vieira, T.M. Interpreting Signal Amplitudes in Surface Electromyography Studies in Sport and Rehabilitation Sciences. Front. Physiol. 2018, 8, 985. [Google Scholar] [CrossRef]
- Vieira, T.M.; Botter, A.; Muceli, S.; Farina, D. Specificity of surface EMG recordings for gastrocnemius during upright standing. Sci. Rep. 2017, 7, 13300. [Google Scholar] [CrossRef] [PubMed]
- Merletti, R.; Holobar, A.; Farina, D. Analysis of motor units with high-density surface electromyography. J. Electromyogr. Kinesiol. 2008, 18, 879–890. [Google Scholar] [CrossRef] [PubMed]
- Vieira, T.M.; Botter, A. The Accurate Assessment of Muscle Excitation Requires the Detection of Multiple Surface Electromyograms. Exerc. Sport. Sci. Rev. 2021, 49, 23–34. [Google Scholar] [CrossRef] [PubMed]
- Miller, L.C.; Thompson, C.K.; Negro, F.; Heckman, C.J.; Farina, D.; Dewald, J.P. High-density surface EMG decomposition allows for recording of motor unit discharge from proximal and distal flexion synergy muscles simultaneously in individuals with stroke. Annu. Int. Conf. IEEE Eng. Med. Biol. Soc. 2014, 2014, 5340–5344. [Google Scholar] [CrossRef] [PubMed]
- Gallina, A.; Pollock, C.L.; Vieira, T.M.; Ivanova, T.D.; Garland, S.J. Between-day reliability of triceps surae responses to standing perturbations in people post-stroke and healthy controls: A high-density surface EMG investigation. Gait. Posture 2016, 44, 103–109. [Google Scholar] [CrossRef] [PubMed]
- Kim, E.H.; Wilson, J.M.; Thompson, C.K.; Heckman, C.J. Differences in estimated persistent inward currents between ankle flexors and extensors in humans. J. Neurophysiol. 2020, 124, 525–535. [Google Scholar] [CrossRef] [PubMed]
- Holobar, A.; Minetto, M.A.; Botter, A.; Negro, F.; Farina, D. Experimental analysis of accuracy in the identification of motor unit spike trains from high-density surface EMG. IEEE Trans. Neural Syst. Rehabil. Eng. 2010, 18, 221–229. [Google Scholar] [CrossRef] [PubMed]
- Marateb, H.R.; McGill, K.C.; Holobar, A.; Lateva, Z.C.; Mansourian, M.; Merletti, R. Accuracy assessment of CKC high-density surface EMG decomposition in biceps femoris muscle. J. Neural Eng. 2011, 8, 066002. [Google Scholar] [CrossRef] [PubMed]
- Martinez-Valdes, E.; Laine, C.; Falla, D.; Mayer, F.; Farina, D. High-density surface electromyography provides reliable estimates of motor unit behaviour. Clin. Neurophysiol. 2016, 127, 2534–2541. [Google Scholar] [CrossRef]
- Ning, Y.; Zhu, X.; Zhu, S.; Zhang, Y. Surface EMG decomposition based on k-means clustering and convolution kernel compensation. IEEE J. Biomed. Health Inform. 2015, 19, 471–477. [Google Scholar] [CrossRef]
- Blok, J.H.; van Dijk, J.P.; Drost, G.; Zwarts, M.J.; Stegeman, D.F. A high-density multichannel surface electromyography system for the characterization of single motor units. Rev. Sci. Instrum. 2002, 73, 1887–1897. [Google Scholar] [CrossRef]
- Farina, D.; Negro, F.; Gazzoni, M.; Enoka, R.M. Detecting the unique representation of motor-unit action potentials in the surface electromyogram. J. Neurophysiol. 2008, 100, 1223–1233. [Google Scholar] [CrossRef]
- Power, K.E.; Lockyer, E.J.; Botter, A.; Vieira, T.; Button, D.C. Endurance-exercise training adaptations in spinal motoneurones: Potential functional relevance to locomotor output and assessment in humans. Eur. J. Appl. Physiol. 2022, 122, 1367–1381. [Google Scholar] [CrossRef] [PubMed]
- Radeke, J.; van Dijk, J.P.; Holobar, A.; Lapatki, B.G. Electrophysiological method to examine muscle fiber architecture in the upper lip in cleft-lip patients. J. Orofacial Orthop. 2014, 74, 51–61. [Google Scholar] [CrossRef] [PubMed]
- Hu, X.; Rymer, W.Z.; Suresh, N.L. Motor unit pool organization examined via spike-triggered averaging of the surface electromyogram. J. Neurophysiol. 2013, 110, 1205–1220. [Google Scholar] [CrossRef] [PubMed]
- Olver, J.; Esquenazi, A.; Fung, V.S.; Singer, B.J.; Ward, A.B.; Cerebral Palsy Institute. Botulinum toxin assessment, intervention and aftercare for lower limb disorders of movement and muscle tone in adults: International consensus statement. Eur. J. Neurol. 2010, 17 (Suppl. S2), 57–73. [Google Scholar] [CrossRef]
- Gupta, A.D.; Chu, W.H.; Howell, S.; Chakraborty, S.; Koblar, S.; Visvanathan, R.; Cameron, I.; Wilson, D. A systematic review: Efficacy of botulinum toxin in walking and quality of life in post-stroke lower limb spasticity. Syst. Rev. 2018, 7, 1. [Google Scholar] [CrossRef] [PubMed]
- Vinti, M.; Gracies, J.M.; Gazzoni, M.; Vieira, T. Localised sampling of myoelectric activity may provide biased estimates of antagonist activation for gastrocnemius though not for soleus and tibialis anterior muscles. J. Electromyogr. Kinesiol. 2018, 38, 34–43. [Google Scholar] [CrossRef] [PubMed]
- Yang, J.F.; Winter, D.A. Electromyographic amplitude normalization methods: Improving their sensitivity as diagnostic tools in gait analysis. Arch. Phys. Med. Rehabil. 1984, 65, 517–521. [Google Scholar]
- von Elm, E.; Altman, D.G.; Egger, M.; Pocock, S.J.; Gøtzsche, P.C.; Vandenbroucke, J.P.; STROBE Initiative. Strengthening the Reporting of Observational Studies in Epidemiology (STROBE) statement: Guidelines for reporting observational studies. BMJ 2007, 335, 806–808. [Google Scholar] [CrossRef]
- Vieira, T.M.; Merletti, R.; Mesin, L. Automatic segmentation of surface EMG images: Improving the estimation of neuromuscular activity. J. Biomech. 2010, 43, 2149–2158. [Google Scholar] [CrossRef]
- Burden, A. How should we normalize electromyograms obtained from healthy participants? What we have learned from over 25 years of research. J. Electromyogr. Kinesiol. 2010, 20, 1023–1035. [Google Scholar] [CrossRef] [PubMed]
- Cruz Martínez, A.; del Campo, F.; Mingo, M.R.; Pérez Conde, M.C. Altered motor unit architecture in hemiparetic patients. A single fibre EMG study. J. Neurol. Neurosurg. Psychiatry 1982, 45, 756–757. [Google Scholar] [CrossRef] [PubMed]
- Ramsay, J.W.; Buchanan, T.S.; Higginson, J.S. Differences in plantar flexor fascicle length and pennation angle between healthy and poststroke individuals and implications for poststroke plantar flexor force contributions. Stroke Res. Treat. 2014, 2014, 919486. [Google Scholar] [CrossRef]
- Gao, F.; Zhang, L.Q. Altered contractile properties of the gastrocnemius muscle poststroke. J. Appl. Physiol. 2008, 105, 1802–1808. [Google Scholar] [CrossRef]
- Kwah, L.K.; Herbert, R.D.; Harvey, L.A.; Diong, J.; Clarke, J.L.; Martin, J.H.; Clarke, E.C.; Hoang, P.D.; Bilston, L.E.; Gandevia, S.C. Passive mechanical properties of gastrocnemius muscles of people with ankle contracture after stroke. Arch. Phys. Med. Rehabil. 2012, 93, 1185–1190. [Google Scholar] [CrossRef] [PubMed]
- Lukács, M. Electrophysiological signs of changes in motor units after ischaemic stroke. Clin. Neurophysiol. 2005, 116, 1566–1570. [Google Scholar] [CrossRef]
- Lukács, M.; Vécsei, L.; Beniczky, S. Changes in muscle fiber density following a stroke. Clin. Neurophysiol. 2009, 120, 1539–1542. [Google Scholar] [CrossRef]
- Vieira, T.M.; Lemos, T.; Oliveira, L.A.S.; Horsczaruk, C.H.R.; Freitas, G.R.; Tovar-Moll, F.; Rodrigues, E.C. Postural Muscle Unit Plasticity in Stroke Survivors: Altered Distribution of Gastrocnemius’ Action Potentials. Front. Neurol. 2019, 10, 686. [Google Scholar] [CrossRef]
- Watanabe, K.; Kouzaki, M.; Moritani, T. Task-dependent spatial distribution of neural activation pattern in human rectus femoris muscle. J. Electromyogr. Kinesiol. 2012, 22, 251–258. [Google Scholar] [CrossRef]
- De Luca, C.J.; Kuznetsov, M.; Gilmore, L.D.; Roy, S.H. Inter-electrode spacing of surface EMG sensors: Reduction of crosstalk contamination during voluntary contractions. J. Biomech. 2012, 45, 555–561. [Google Scholar] [CrossRef] [PubMed]
- De Luca, C.J.; Merletti, R. Surface myoelectric signal cross-talk among muscles of the leg. Electroencephalogr. Clin. Neurophysiol. 1988, 69, 568–575. [Google Scholar] [CrossRef] [PubMed]
- Lynn, P.A.; Bettles, N.D.; Hughes, A.D.; Johnson, S.W. Influences of electrode geometry on bipolar recordings of the surface electromyogram. Med. Biol. Eng. Comput. 1978, 16, 651–660. [Google Scholar] [CrossRef] [PubMed]
- Roeleveld, K.; Stegeman, D.F.; Vingerhoets, H.M.; Oosterom, A.V. Motor unit potential contribution to surface electromyography. Acta Physiol. Scand. 1997, 160, 175–183. [Google Scholar] [CrossRef] [PubMed]
- Rutkove, S.B. Introduction to Volume Conduction. In The Clinical Neurophysiology Primer; Humana Press: Totowa, NJ, USA, 2007; pp. 43–53. [Google Scholar]
- Edgerton, V.R.; Smith, J.L.; Simpson, D.R. Muscle fibre type populations of human leg muscles. Histochem. J. 1975, 7, 259–266. [Google Scholar] [CrossRef]
- Rainoldi, A.; Melchiorri, G.; Caruso, I. A method for positioning electrodes during surface EMG recordings in lower limb muscles. J. Neurosci. Methods. 2004, 134, 37–43. [Google Scholar] [CrossRef]




{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Subjects (n) | 12 |
| Age (y) | 51 ± 14 |
| Time since paresis onset (y) | 6 ± 6 |
| Gender | |
| Female (n) | 3 |
| Male (n) | 9 |
| Paretic side | |
| Left (n) | 7 |
| Right (n) | 5 |
| Cause | |
| Ischemic stroke (n) | 9 |
| Hemorrhagic stroke (n) | 3 |
| Muscle | Knee Position | Effort Level | BipCAN | ReCAN | AbCAN |
|---|---|---|---|---|---|
| Gastrocnemius medialis | |||||
| n = 12 | Flexed | Submaximal | 0.33 ± 0.30 | 0.51 ± 0.33 | 0.30 ± 0.21 |
| Maximal | 0.48 ± 0.42 | 0.71 ± 0.33 | 0.41 ± 0.23 | ||
| Extended | Submaximal | 0.34 ± 0.33 | 0.47 ± 0.32 | 0.28 ± 0.23 | |
| Maximal | 0.53 ± 0.36 | 0.63 ± 0.33 | 0.36 ± 0.24 | ||
| Medial Soleus | |||||
| n = 12 | Flexed | Submaximal | - | 0.40 ± 0.35 | 0.29 ± 0.23 |
| Maximal | - | 0.48 ± 0.38 | 0.35 ± 0.25 | ||
| Extended | Submaximal | - | 0.61 ± 0.76 | 0.54 ± 0.79 | |
| Maximal | - | 0.63 ± 0.43 | 0.49 ± 0.33 | ||
| Lateral Soleus | |||||
| n = 12 | Flexed | Submaximal | - | 0.37 ± 0.32 | 0.27 ± 0.23 |
| Maximal | - | 0.48 ± 0.35 | 0.34 ± 0.25 | ||
| Extended | Submaximal | - | 0.41 ± 0.42 | 0.40 ± 0.59 | |
| Maximal | - | 0.61 ± 0.38 | 0.47 ± 0.31 | ||
| Knee Position | Effort Level | Absolute | Relative | |
|---|---|---|---|---|
| Location of the CAN centroid along proximal-distal axis | ||||
| Flexed | Submaximal | 0.56 ± 0.16 | 0.49 ± 0.18 | |
| Maximal | 0.46 ± 0.13 | 0.42 ± 0.11 | ||
| Extended | Submaximal | 0.54 ± 0.17 | 0.37 ± 0.16 | |
| Maximal | 0.64 ± 0.25 | 0.42 ± 0.19 | ||
| Location of the CAN centroid along medial-lateral axis | ||||
| Flexed | Submaximal | 0.63 ± 0.17 | 0.68 ± 0.14 | |
| Maximal | 0.61 ± 0.14 | 0.64 ± 0.14 | ||
| Extended | Submaximal | 0.67 ± 0.22 | 0.60 ± 0.15 | |
| Maximal | 0.60 ± 0.17 | 0.57 ± 0.16 | ||
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ghédira, M.; Vieira, T.M.; Cerone, G.L.; Gazzoni, M.; Gracies, J.-M.; Hutin, E. Antagonist Activation Measurement in Triceps Surae Using High-Density and Bipolar Surface EMG in Chronic Hemiparesis. Sensors 2024, 24, 3701. https://doi.org/10.3390/s24123701
Ghédira M, Vieira TM, Cerone GL, Gazzoni M, Gracies J-M, Hutin E. Antagonist Activation Measurement in Triceps Surae Using High-Density and Bipolar Surface EMG in Chronic Hemiparesis. Sensors. 2024; 24(12):3701. https://doi.org/10.3390/s24123701
Chicago/Turabian StyleGhédira, Mouna, Taian Martins Vieira, Giacinto Luigi Cerone, Marco Gazzoni, Jean-Michel Gracies, and Emilie Hutin. 2024. "Antagonist Activation Measurement in Triceps Surae Using High-Density and Bipolar Surface EMG in Chronic Hemiparesis" Sensors 24, no. 12: 3701. https://doi.org/10.3390/s24123701
APA StyleGhédira, M., Vieira, T. M., Cerone, G. L., Gazzoni, M., Gracies, J.-M., & Hutin, E. (2024). Antagonist Activation Measurement in Triceps Surae Using High-Density and Bipolar Surface EMG in Chronic Hemiparesis. Sensors, 24(12), 3701. https://doi.org/10.3390/s24123701