


Perceptual Integration Compensates for Attention Deficit in Elderly during Repetitive Auditory-Based Sensorimotor Task

 , , , , , ,
, , , , , ,  and
and 
Abstract
1. Introduction
2. Materials and Methods
2.1. Participants
2.2. Neurophysiological Assessment
2.3. Sensorimotor Integration Session
2.4. EEG Preprocessing and Epochs Segmentation
2.5. Sensor-Level Analysis of Spectral Power
- prestimulus activity: the SP was averaged within the frequency bands of interest over the interval s before stimulus presentation and the problem was addressed only in a spatial domain;
- poststimulus activity: the SP was considered on the interval s after stimulus presentation, i.e., a problem was addressed in both spatial and temporal domains.
2.6. Sensor-Level Connectivity Analysis
3. Results
3.1. Effect of Repetition and Age on the Prestimulus Spectral Power
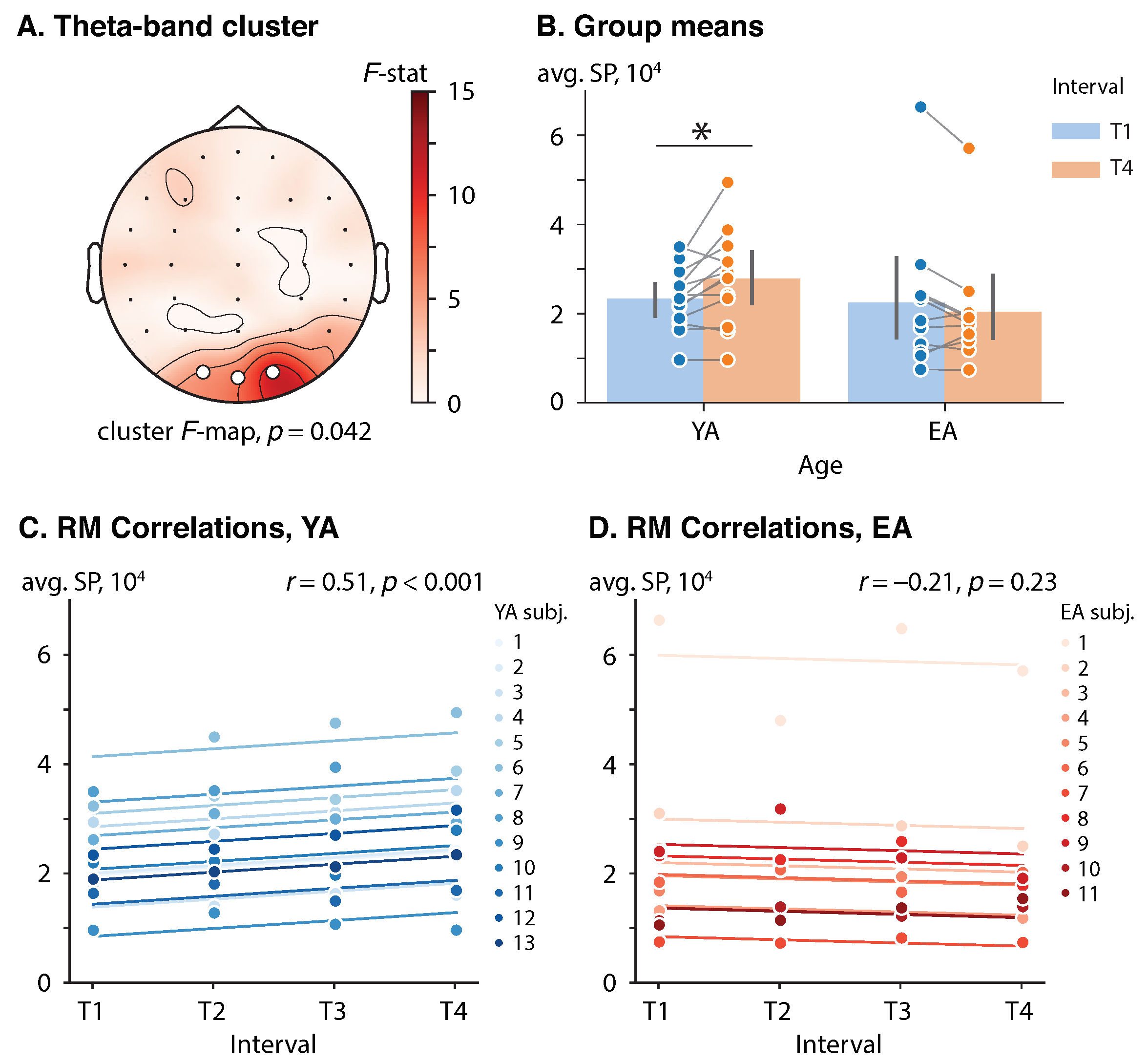
3.2. Effect of Repetition and Age on the Poststimulus Spectral Power
3.3. Effect of Repetition and Age on Functional Connectivity
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Golub, J.S. Brain changes associated with age-related hearing loss. Curr. Opin. Otolaryngol. Head Neck Surg. 2017, 25, 347–352. [Google Scholar] [CrossRef] [PubMed]
- Vaqué-Alcázar, L.; Sala-Llonch, R.; Valls-Pedret, C.; Vidal-Piñeiro, D.; Fernández-Cabello, S.; Bargalló, N.; Ros, E.; Bartrés-Faz, D. Differential age-related gray and white matter impact mediates educational influence on elders’ cognition. Brain Imaging Behav. 2017, 11, 318–332. [Google Scholar] [CrossRef] [PubMed]
- Oschwald, J.; Guye, S.; Liem, F.; Rast, P.; Willis, S.; Röcke, C.; Jäncke, L.; Martin, M.; Mérillat, S. Brain structure and cognitive ability in healthy aging: A review on longitudinal correlated change. Rev. Neurosci. 2020, 31, 1–57. [Google Scholar] [CrossRef] [PubMed]
- Zhai, F.; Liu, J.; Su, N.; Han, F.; Zhou, L.; Ni, J.; Yao, M.; Zhang, S.; Jin, Z.; Cui, L.; et al. Disrupted white matter integrity and network connectivity are related to poor motor performance. Sci. Rep. 2020, 10, 18369. [Google Scholar] [CrossRef] [PubMed]
- Degardin, A.; Devos, D.; Cassim, F.; Bourriez, J.L.; Defebvre, L.; Derambure, P.; Devanne, H. Deficit of sensorimotor integration in normal aging. Neurosci. Lett. 2011, 498, 208–212. [Google Scholar] [CrossRef]
- Yoshimura, N.; Tsuda, H.; Aquino, D.; Takagi, A.; Ogata, Y.; Koike, Y.; Minati, L. Age-related decline of sensorimotor integration influences resting-state functional brain connectivity. Brain Sci. 2020, 10, 966. [Google Scholar] [CrossRef]
- de Dieuleveult, A.L.; Siemonsma, P.C.; van Erp, J.B.; Brouwer, A.M. Effects of aging in multisensory integration: A systematic review. Front. Aging Neurosci. 2017, 9, 80. [Google Scholar] [CrossRef]
- Young-Bernier, M.; Davidson, P.S.; Tremblay, F. Paired-pulse afferent modulation of TMS responses reveals a selective decrease in short latency afferent inhibition with age. Neurobiol. Aging 2012, 33, 835.e1–835.e11. [Google Scholar] [CrossRef]
- Brown, K.; Neva, J.; Feldman, S.; Staines, W.; Boyd, L. Sensorimotor integration in healthy aging: Baseline differences and response to sensory training. Exp. Gerontol. 2018, 112, 1–8. [Google Scholar] [CrossRef]
- Ferreri, F.; Guerra, A.; Vollero, L.; Ponzo, D.; Maatta, S.; Mervaala, E.; Iannello, G.; Di Lazzaro, V. Age-related changes of cortical excitability and connectivity in healthy humans: Non-invasive evaluation of sensorimotor network by means of TMS-EEG. Neuroscience 2017, 357, 255–263. [Google Scholar] [CrossRef]
- Ferreri, F.; Vecchio, F.; Guerra, A.; Miraglia, F.; Ponzo, D.; Vollero, L.; Iannello, G.; Maatta, S.; Mervaala, E.; Rossini, P.M.; et al. Age related differences in functional synchronization of EEG activity as evaluated by means of TMS-EEG coregistrations. Neurosci. Lett. 2017, 647, 141–146. [Google Scholar] [CrossRef] [PubMed]
- Murray, M.M.; Eardley, A.F.; Edginton, T.; Oyekan, R.; Smyth, E.; Matusz, P.J. Sensory dominance and multisensory integration as screening tools in aging. Sci. Rep. 2018, 8, 8901. [Google Scholar] [CrossRef] [PubMed]
- Peiffer, A.M.; Hugenschmidt, C.E.; Maldjian, J.A.; Casanova, R.; Srikanth, R.; Hayasaka, S.; Burdette, J.H.; Kraft, R.A.; Laurienti, P.J. Aging and the interaction of sensory cortical function and structure. Hum. Brain Mapp. 2009, 30, 228–240. [Google Scholar] [CrossRef] [PubMed]
- Redel, P.; Bublak, P.; Sorg, C.; Kurz, A.; Förstl, H.; Müller, H.; Schneider, W.X.; Perneczky, R.; Finke, K. Deficits of spatial and task-related attentional selection in mild cognitive impairment and Alzheimer’s disease. Neurobiol. Aging 2012, 33, 195.e27–195.e42. [Google Scholar] [CrossRef]
- Dushanova, J.; Christov, M. The effect of aging on EEG brain oscillations related to sensory and sensorimotor functions. Adv. Med. Sci. 2014, 59, 61–67. [Google Scholar] [CrossRef]
- Frolov, N.S.; Pitsik, E.N.; Maksimenko, V.A.; Grubov, V.V.; Kiselev, A.R.; Wang, Z.; Hramov, A.E. Age-related slowing down in the motor initiation in elderly adults. PLoS ONE 2020, 15, e0233942. [Google Scholar] [CrossRef]
- Frolov, N.; Pitsik, E.; Maksimenko, V.; Hramov, A. Applying recurrence time entropy to identify changes in event-related potentials. Eur. Phys. J. Spec. Top. 2022, 232, 161–168. [Google Scholar] [CrossRef]
- Pitsik, E.N.; Frolov, N.S.; Shusharina, N.; Hramov, A.E. Age-related changes in functional connectivity during the sensorimotor integration detected by artificial neural network. Sensors 2022, 22, 2537. [Google Scholar] [CrossRef]
- Bock, O.; Schneider, S. Sensorimotor adaptation in young and elderly humans. Neurosci. Biobehav. Rev. 2002, 26, 761–767. [Google Scholar] [CrossRef]
- Bock, O.; Girgenrath, M. Relationship between sensorimotor adaptation and cognitive functions in younger and older subjects. Exp. Brain Res. 2006, 169, 400–406. [Google Scholar] [CrossRef]
- Freyer, F.; Reinacher, M.; Nolte, G.; Dinse, H.R.; Ritter, P. Repetitive tactile stimulation changes resting-state functional connectivity—Implications for treatment of sensorimotor decline. Front. Hum. Neurosci. 2012, 6, 144. [Google Scholar] [CrossRef]
- Blais, M.; Martin, E.; Albaret, J.M.; Tallet, J. Preservation of perceptual integration improves temporal stability of bimanual coordination in the elderly: An evidence of age-related brain plasticity. Behav. Brain Res. 2014, 275, 34–42. [Google Scholar] [CrossRef] [PubMed]
- Predel, C.; Kaminski, E.; Hoff, M.; Carius, D.; Villringer, A.; Ragert, P. Motor Skill Learning-Induced Functional Plasticity in the Primary Somatosensory Cortex: A Comparison Between Young and Older Adults. Front. Aging Neurosci. 2020, 12, 596438. [Google Scholar] [CrossRef] [PubMed]
- Vieweg, J.; Panzer, S.; Schaefer, S. Effects of age simulation and age on motor sequence learning: Interaction of age-related cognitive and motor decline. Hum. Mov. Sci. 2023, 87, 103025. [Google Scholar] [CrossRef] [PubMed]
- King, B.R.; Fogel, S.M.; Albouy, G.; Doyon, J. Neural correlates of the age-related changes in motor sequence learning and motor adaptation in older adults. Front. Hum. Neurosci. 2013, 7, 142. [Google Scholar] [CrossRef]
- Maria Rossini, P.; Ada Noris Ferilli, M.; Rossini, L.; Ferreri, F. Clinical neurophysiology of brain plasticity in aging brain. Curr. Pharm. Des. 2013, 19, 6426–6439. [Google Scholar] [CrossRef]
- Degen, C.; Schröder, J. Training-induced cerebral changes in the elderly. Restor. Neurol. Neurosci. 2014, 32, 213–221. [Google Scholar] [CrossRef]
- Cheng, C.H.; Lin, M.Y.; Yang, S.H. Age effect on automatic inhibitory function of the somatosensory and motor cortex: An MEG study. Front. Aging Neurosci. 2018, 10, 53. [Google Scholar] [CrossRef]
- Nuwer, M.R.; Comi, G.; Emerson, R.; Fuglsang-Frederiksen, A.; Guérit, J.M.; Hinrichs, H.; Ikeda, A.; Luccas, F.J.C.; Rappelsburger, P. IFCN standards for digital recording of clinical EEG. Electroencephalogr. Clin. Neurophysiol. 1998, 106, 259–261. [Google Scholar] [CrossRef]
- Hyvärinen, A.; Oja, E. Independent component analysis: Algorithms and applications. Neural Netw. 2000, 13, 411–430. [Google Scholar] [CrossRef]
- Gramfort, A.; Luessi, M.; Larson, E.; Engemann, D.A.; Strohmeier, D.; Brodbeck, C.; Goj, R.; Jas, M.; Brooks, T.; Parkkonen, L.; et al. MEG and EEG data analysis with MNE-Python. Front. Neurosci. 2013, 7, 267. [Google Scholar] [CrossRef]
- Hramov, A.E.; Koronovskii, A.A.; Makarov, V.A.; Pavlov, A.N.; Sitnikova, E. Wavelets in Neuroscience; Springer: New York, NY, USA, 2015. [Google Scholar]
- Pitsik, E.; Frolov, N.; Hauke Kraemer, K.; Grubov, V.; Maksimenko, V.; Kurths, J.; Hramov, A. Motor execution reduces EEG signals complexity: Recurrence quantification analysis study. Chaos Interdiscip. J. Nonlinear Sci. 2020, 30, 023111. [Google Scholar] [CrossRef] [PubMed]
- Klimesch, W. EEG alpha and theta oscillations reflect cognitive and memory performance: A review and analysis. Brain Res. Rev. 1999, 29, 169–195. [Google Scholar] [CrossRef] [PubMed]
- Maris, E.; Oostenveld, R. Nonparametric statistical testing of EEG-and MEG-data. J. Neurosci. Methods 2007, 164, 177–190. [Google Scholar] [CrossRef]
- Bakdash, J.Z.; Marusich, L.R. Repeated measures correlation. Front. Psychol. 2017, 8, 456. [Google Scholar] [CrossRef] [PubMed]
- Vallat, R. Pingouin: Statistics in Python. J. Open Source Softw. 2018, 3, 1026. [Google Scholar] [CrossRef]
- Hramov, A.E.; Frolov, N.S.; Maksimenko, V.A.; Kurkin, S.A.; Kazantsev, V.B.; Pisarchik, A.N. Functional networks of the brain: From connectivity restoration to dynamic integration. Physics-Uspekhi 2021, 64, 584. [Google Scholar] [CrossRef]
- Bastos, A.M.; Schoffelen, J.M. A tutorial review of functional connectivity analysis methods and their interpretational pitfalls. Front. Syst. Neurosci. 2016, 9, 175. [Google Scholar] [CrossRef]
- Lachaux, J.P.; Rodriguez, E.; Martinerie, J.; Varela, F.J. Measuring phase synchrony in brain signals. Hum. Brain Mapp. 1999, 8, 194–208. [Google Scholar] [CrossRef]
- Zalesky, A.; Fornito, A.; Bullmore, E.T. Network-based statistic: Identifying differences in brain networks. Neuroimage 2010, 53, 1197–1207. [Google Scholar] [CrossRef]
- Pitsik, E.N. Recurrence quantification analysis provides the link between age-related decline in motor brain response and complexity of the baseline EEG. Appl. Nonlinear Dyn. 2021, 29, 386–397. [Google Scholar]
- Solomon, E.A.; Kragel, J.E.; Sperling, M.R.; Sharan, A.; Worrell, G.; Kucewicz, M.; Inman, C.S.; Lega, B.; Davis, K.A.; Stein, J.M.; et al. Widespread theta synchrony and high-frequency desynchronization underlies enhanced cognition. Nat. Commun. 2017, 8, 1704. [Google Scholar] [CrossRef] [PubMed]
- Postuma, R.B. Biomarkers of Neurodegenerative Disease in Idiopathic RBD. In Rapid-Eye-Movement Sleep Behavior Disorder; Springer: New York, NY, USA, 2019; pp. 527–540. [Google Scholar]
- Addante, R.J.; Watrous, A.J.; Yonelinas, A.P.; Ekstrom, A.D.; Ranganath, C. Prestimulus theta activity predicts correct source memory retrieval. Proc. Natl. Acad. Sci. USA 2011, 108, 10702–10707. [Google Scholar] [CrossRef] [PubMed]
- Kurkin, S.; Gordleeva, S.; Savosenkov, A.; Grigorev, N.; Smirnov, N.; Grubov, V.V.; Udoratina, A.; Maksimenko, V.; Kazantsev, V.; Hramov, A.E. Transcranial Magnetic Stimulation of the Dorsolateral Prefrontal Cortex Increases Posterior Theta Rhythm and Reduces Latency of Motor Imagery. Sensors 2023, 23, 4661. [Google Scholar] [CrossRef]
- White, T.P.; Jansen, M.; Doege, K.; Mullinger, K.J.; Park, S.B.; Liddle, E.B.; Gowland, P.A.; Francis, S.T.; Bowtell, R.; Liddle, P.F. Theta power during encoding predicts subsequent-memory performance and default mode network deactivation. Hum. Brain Mapp. 2013, 34, 2929–2943. [Google Scholar] [CrossRef]
- Han, H.B.; Lee, K.E.; Choi, J.H. Functional dissociation of θ oscillations in the frontal and visual cortices and their long-range network during sustained attention. Eneuro 2019, 6, 1–12. [Google Scholar] [CrossRef]
- Reiner, M.; Rozengurt, R.; Barnea, A. Better than sleep: Theta neurofeedback training accelerates memory consolidation. Biol. Psychol. 2014, 95, 45–53. [Google Scholar] [CrossRef]
- McNab, F.; Klingberg, T. Prefrontal cortex and basal ganglia control access to working memory. Nat. Neurosci. 2008, 11, 103–107. [Google Scholar] [CrossRef]
- Sauseng, P.; Klimesch, W.; Schabus, M.; Doppelmayr, M. Fronto-parietal EEG coherence in theta and upper alpha reflect central executive functions of working memory. Int. J. Psychophysiol. 2005, 57, 97–103. [Google Scholar] [CrossRef]
- Kawasaki, M.; Kitajo, K.; Yamaguchi, Y. Fronto-parietal and fronto-temporal theta phase synchronization for visual and auditory-verbal working memory. Front. Psychol. 2014, 5, 200. [Google Scholar] [CrossRef]
- Kawasaki, M.; Kitajo, K.; Yamaguchi, Y. Dynamic links between theta executive functions and alpha storage buffers in auditory and visual working memory. Eur. J. Neurosci. 2010, 31, 1683–1689. [Google Scholar] [CrossRef] [PubMed]
- Cabeza, R.; Dennis, N.A. Frontal lobes and aging: Deterioration and compensation. Princ. Front. Lobe Funct. 2012, 2, 628–652. [Google Scholar]
- Boemio, A.; Fromm, S.; Braun, A.; Poeppel, D. Hierarchical and asymmetric temporal sensitivity in human auditory cortices. Nat. Neurosci. 2005, 8, 389–395. [Google Scholar] [CrossRef] [PubMed]
- Flinker, A.; Doyle, W.K.; Mehta, A.D.; Devinsky, O.; Poeppel, D. Spectrotemporal modulation provides a unifying framework for auditory cortical asymmetries. Nat. Hum. Behav. 2019, 3, 393–405. [Google Scholar] [CrossRef]
- Hickok, G.; Poeppel, D. Dorsal and ventral streams: A framework for understanding aspects of the functional anatomy of language. Cognition 2004, 92, 67–99. [Google Scholar] [CrossRef]
- Rauschecker, J.P.; Scott, S.K. Maps and streams in the auditory cortex: Nonhuman primates illuminate human speech processing. Nat. Neurosci. 2009, 12, 718–724. [Google Scholar] [CrossRef]
- Martin, J.S.; Jerger, J.F. Some effects of aging on central auditory processing. J. Rehabil. Res. Dev. 2005, 42, 25–44. [Google Scholar] [CrossRef]
- Profant, O.; Škoch, A.; Balogová, Z.; Tintěra, J.; Hlinka, J.; Syka, J. Diffusion tensor imaging and MR morphometry of the central auditory pathway and auditory cortex in aging. Neuroscience 2014, 260, 87–97. [Google Scholar] [CrossRef]
- Sun, J.; Wang, B.; Niu, Y.; Tan, Y.; Fan, C.; Zhang, N.; Xue, J.; Wei, J.; Xiang, J. Complexity analysis of EEG, MEG, and fMRI in mild cognitive impairment and Alzheimer’s disease: A review. Entropy 2020, 22, 239. [Google Scholar] [CrossRef]
- Pappalettera, C.; Miraglia, F.; Cotelli, M.; Rossini, P.M.; Vecchio, F. Analysis of complexity in the EEG activity of Parkinson’s disease patients by means of approximate entropy. GeroScience 2022, 44, 1599–1607. [Google Scholar] [CrossRef]
- Cancela, J.; Moreno, E.M.; Arredondo, M.T.; Bonato, P. Designing auditory cues for Parkinson’s disease gait rehabilitation. In Proceedings of the 2014 36th Annual International Conference of the IEEE Engineering in Medicine and Biology Society, Chicago, IL, USA, 26–30 August 2014; pp. 5852–5855. [Google Scholar]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Group, No. | IAF | Theta | Lower-1 Alpha | Lower-2 Alpha | Upper Alpha |
|---|---|---|---|---|---|
| YA, 1 | 10.4 Hz | 4.4–6.4 Hz | 6.4–8.4 Hz | 8.4–10.4 Hz | 10.4–12.4 Hz |
| YA, 2 | 11.0 Hz | 5.0–7.0 Hz | 7.0–9.0 Hz | 9.0–11.0 Hz | 11.0–13.0 Hz |
| YA, 3 | 10.3 Hz | 4.3–6.3 Hz | 6.3–8.3 Hz | 8.3–10.3 Hz | 10.3–12.3 Hz |
| YA, 4 | 10.5 Hz | 4.5–6.5 Hz | 6.5–8.5 Hz | 8.5–10.5 Hz | 10.5–12.5 Hz |
| YA, 5 | 9.4 Hz | 3.4–5.4 Hz | 5.4–7.4 Hz | 7.4–9.4 Hz | 9.4–11.4 Hz |
| YA, 6 | 9.4 Hz | 3.4–5.4 Hz | 5.4–7.4 Hz | 7.4–9.4 Hz | 9.4–11.4 Hz |
| YA, 7 | 10.1 Hz | 4.1–6.1 Hz | 6.1–8.1 Hz | 8.1–10.1 Hz | 10.1–12.1 Hz |
| YA, 8 | 9.0 Hz | 3.0–5.0 Hz | 5.0–7.0 Hz | 7.0–9.0 Hz | 9.0–11.0 Hz |
| YA, 9 | 10.5 Hz | 4.5–6.5 Hz | 6.5–8.5 Hz | 8.5–10.5 Hz | 10.5–12.5 Hz |
| YA, 10 | 10.9 Hz | 4.9–6.9 Hz | 6.9–8.9 Hz | 8.9–10.9 Hz | 10.9–12.9 Hz |
| YA, 11 | 10.8 Hz | 4.8–6.8 Hz | 6.8–8.8 Hz | 8.8–10.8 Hz | 10.8–12.8 Hz |
| YA, 12 | 9.5 Hz | 3.5–5.5 Hz | 5.5–7.5 Hz | 7.5–9.5 Hz | 9.5–11.5 Hz |
| YA, 13 | 10.6 Hz | 4.6–6.6 Hz | 6.6–8.6 Hz | 8.6–10.6 Hz | 10.6–12.6 Hz |
| EA, 1 | 8.9 Hz | 2.9–4.9 Hz | 4.9–6.9 Hz | 6.9–8.9 Hz | 8.9–10.9 Hz |
| EA, 2 | 10.3 Hz | 4.3–6.3 Hz | 6.3–8.3 Hz | 8.3–10.3 Hz | 10.3–12.3 Hz |
| EA, 3 | 8.7 Hz | 2.7–4.7 Hz | 4.7–6.7 Hz | 6.7–8.7 Hz | 8.7–10.7 Hz |
| EA, 4 | 8.5 Hz | 2.5–4.5 Hz | 4.5–6.5 Hz | 6.5–8.5 Hz | 8.5–10.5 Hz |
| EA, 5 | 10.0 Hz | 4.0–6.0 Hz | 6.0–8.0 Hz | 8.0–10.0 Hz | 10.0–12.0 Hz |
| EA, 6 | 9.1 Hz | 3.1–5.1 Hz | 5.1–7.1 Hz | 7.1–9.1 Hz | 9.1–11.1 Hz |
| EA, 7 | 10.4 Hz | 4.4–6.4 Hz | 6.4–8.4 Hz | 8.4–10.4 Hz | 10.4–12.4 Hz |
| EA, 8 | 11.5 Hz | 5.5–7.5 Hz | 7.5–9.5 Hz | 9.5–11.5 Hz | 11.5–13.5 Hz |
| EA, 9 | 9.1 Hz | 3.1–5.1 Hz | 5.1–7.1 Hz | 7.1–9.1 Hz | 9.1–11.1 Hz |
| EA, 10 | 11.1 Hz | 5.1–7.1 Hz | 7.1–9.1 Hz | 9.1–11.1 Hz | 11.1–13.1 Hz |
| EA, 11 | 10.1 Hz | 4.1–6.1 Hz | 6.1–8.1 Hz | 8.1–10.1 Hz | 10.1–12.1 Hz |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Frolov, N.; Pitsik, E.; Grubov, V.; Badarin, A.; Maksimenko, V.; Zakharov, A.; Kurkin, S.; Hramov, A. Perceptual Integration Compensates for Attention Deficit in Elderly during Repetitive Auditory-Based Sensorimotor Task. Sensors 2023, 23, 6420. https://doi.org/10.3390/s23146420
Frolov N, Pitsik E, Grubov V, Badarin A, Maksimenko V, Zakharov A, Kurkin S, Hramov A. Perceptual Integration Compensates for Attention Deficit in Elderly during Repetitive Auditory-Based Sensorimotor Task. Sensors. 2023; 23(14):6420. https://doi.org/10.3390/s23146420
Chicago/Turabian StyleFrolov, Nikita, Elena Pitsik, Vadim Grubov, Artem Badarin, Vladimir Maksimenko, Alexander Zakharov, Semen Kurkin, and Alexander Hramov. 2023. "Perceptual Integration Compensates for Attention Deficit in Elderly during Repetitive Auditory-Based Sensorimotor Task" Sensors 23, no. 14: 6420. https://doi.org/10.3390/s23146420
APA StyleFrolov, N., Pitsik, E., Grubov, V., Badarin, A., Maksimenko, V., Zakharov, A., Kurkin, S., & Hramov, A. (2023). Perceptual Integration Compensates for Attention Deficit in Elderly during Repetitive Auditory-Based Sensorimotor Task. Sensors, 23(14), 6420. https://doi.org/10.3390/s23146420