
Age-Related Changes in Functional Connectivity during the Sensorimotor Integration Detected by Artificial Neural Network

 , , and
, , and 
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
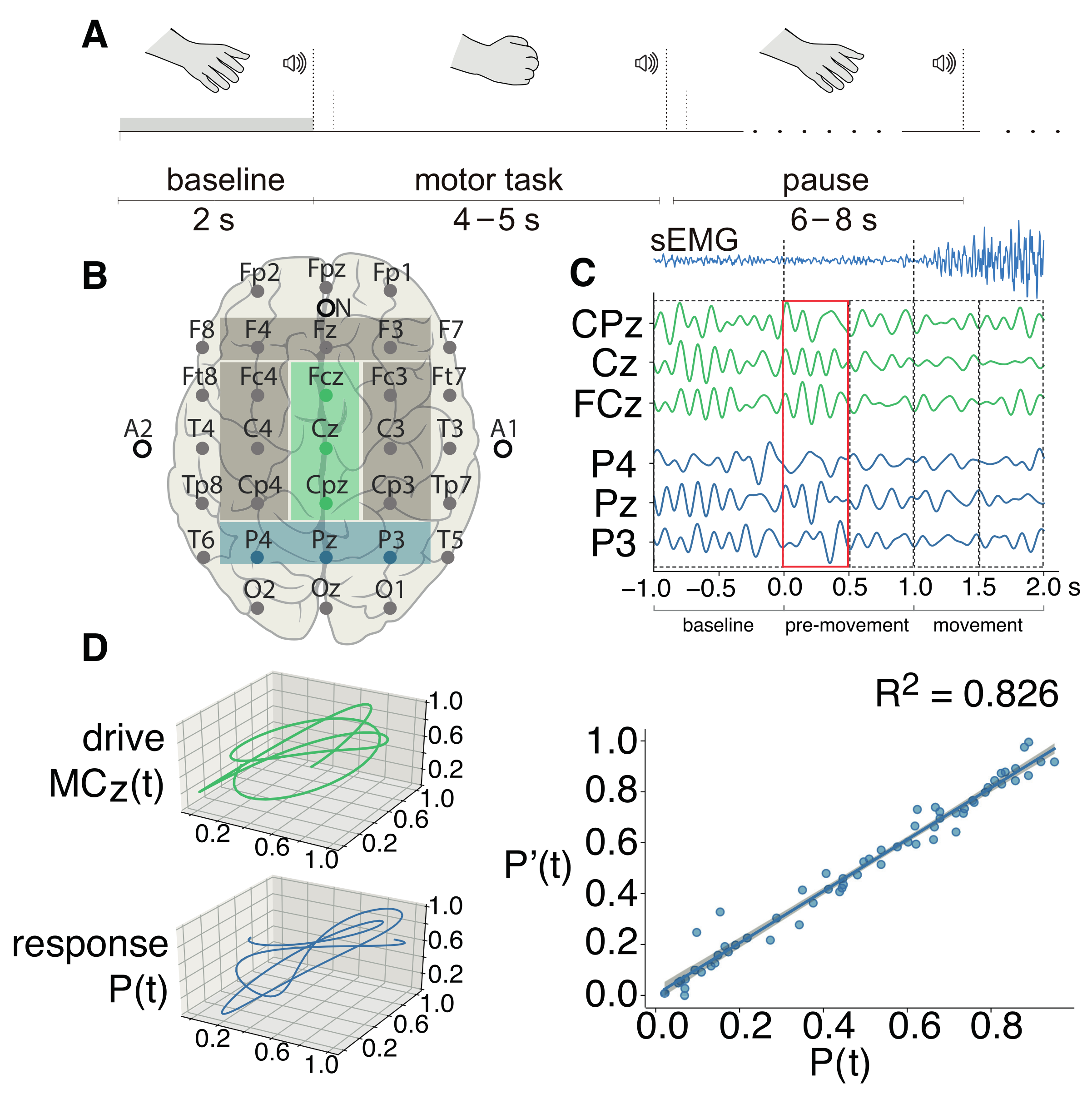
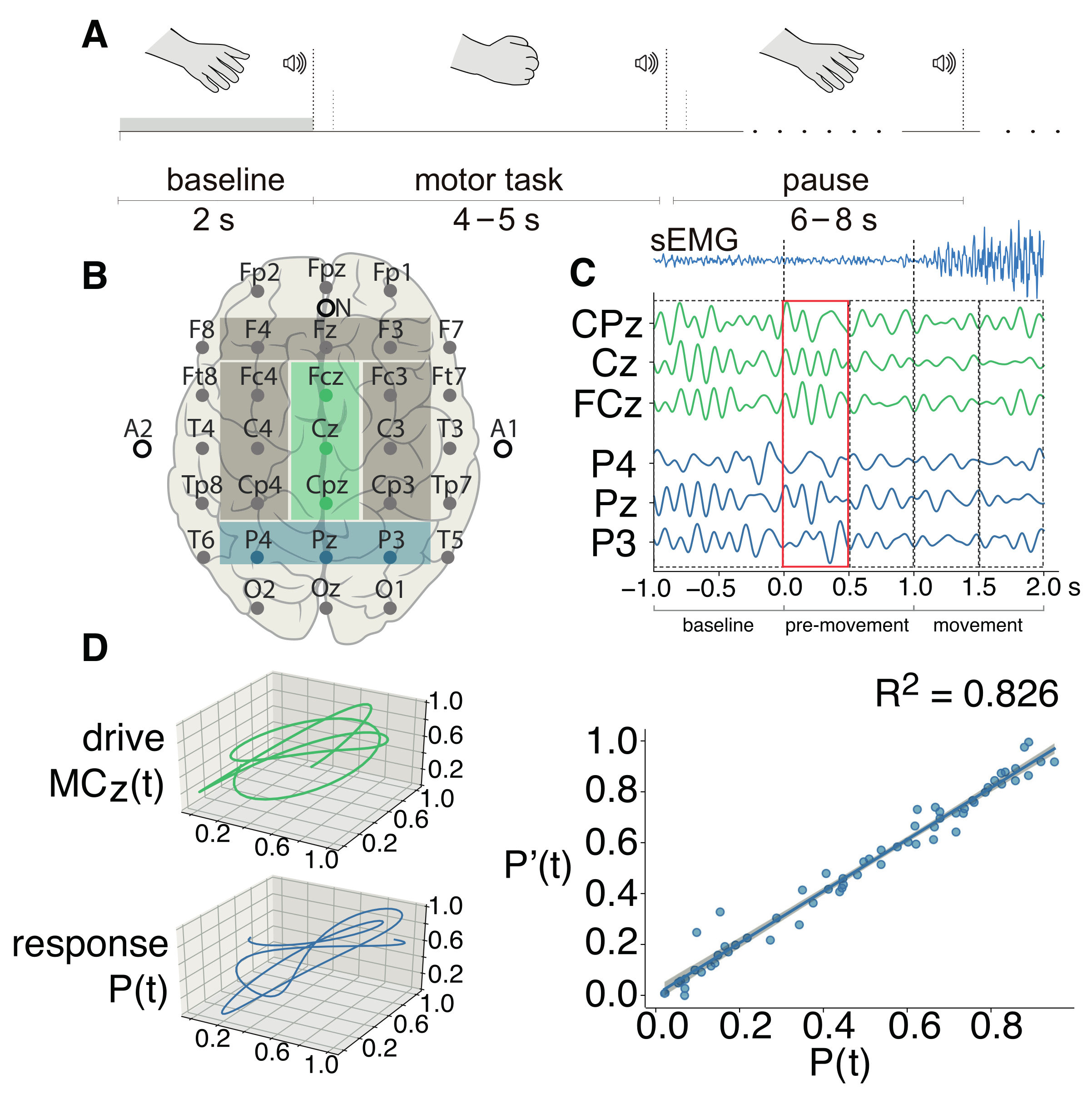
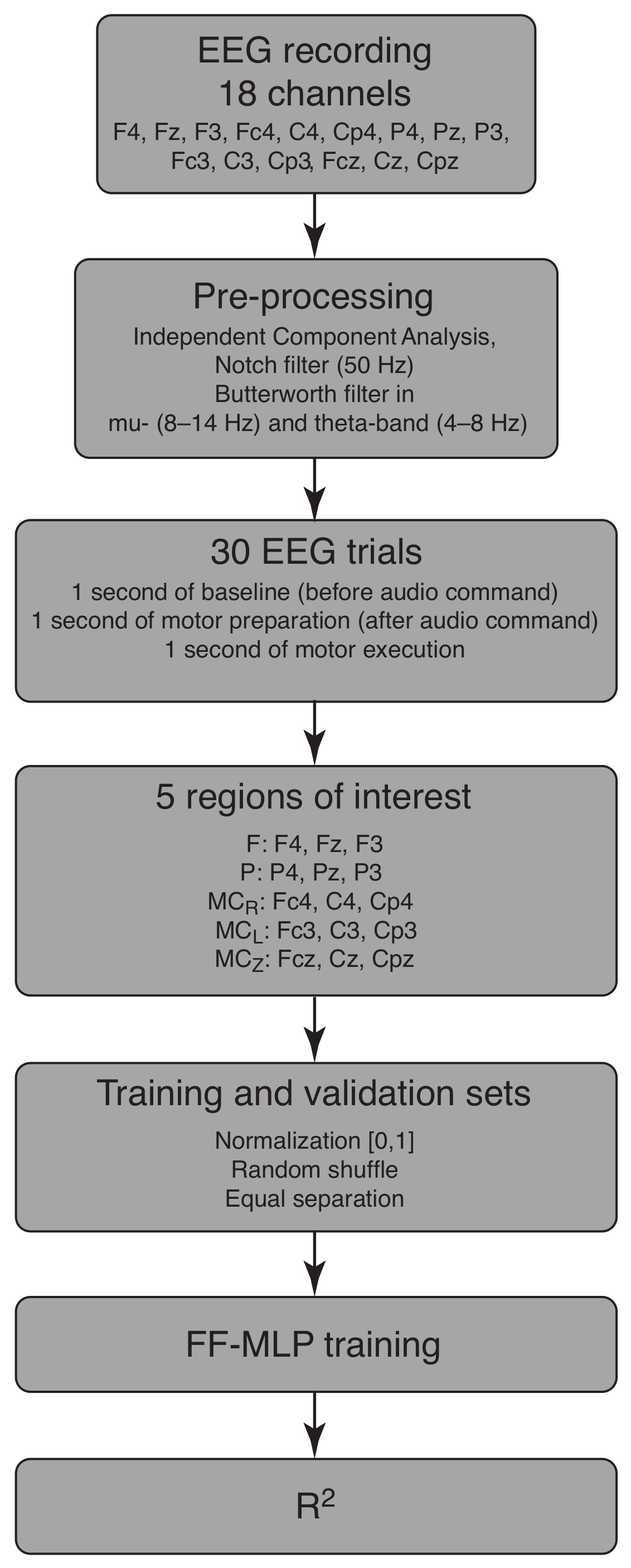
2. Materials and Methods
2.1. Experimental Dataset
2.2. FF-MLP Model
2.2.1. Training and Validation Datasets
2.2.2. MLP Model Configuration
2.3. Statistical Analysis
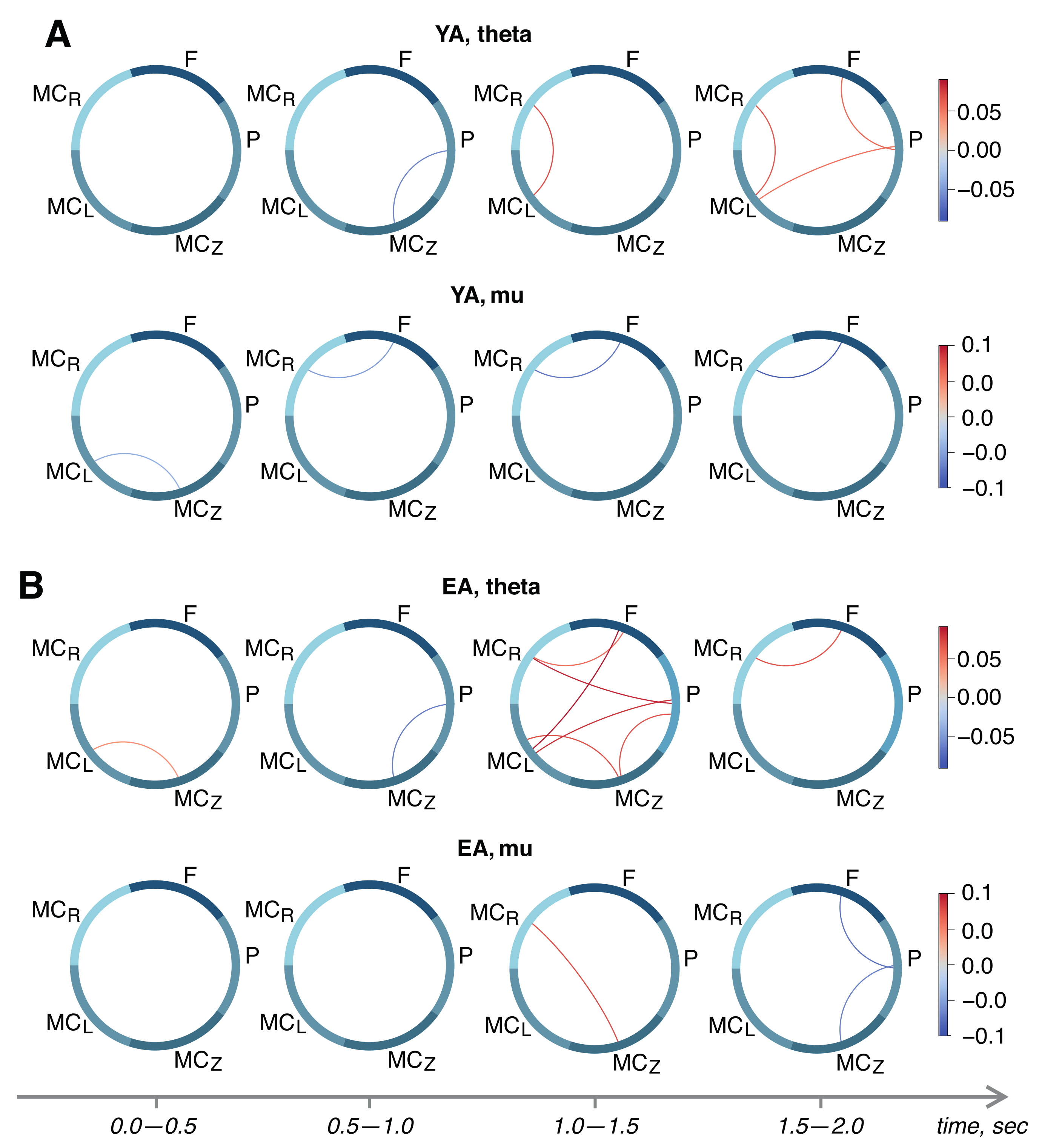
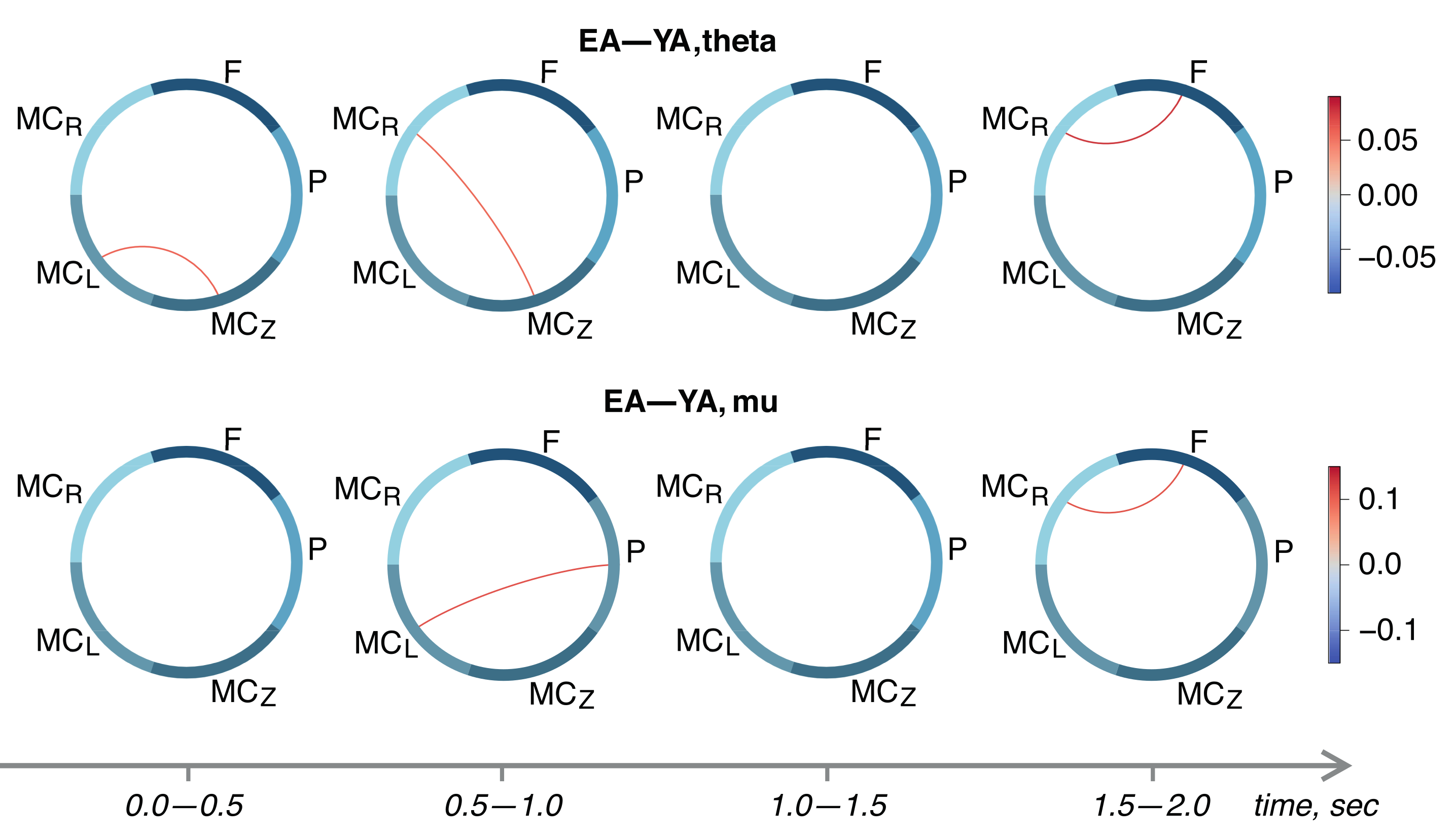
3. Results
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Pisarchik, A.N.; Maksimenko, V.A.; Andreev, A.V.; Frolov, N.S.; Makarov, V.V.; Zhuravlev, M.O.; Runnova, A.E.; Hramov, A.E. Coherent resonance in the distributed cortical network during sensory information processing. Sci. Rep. 2019, 9, 18325. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cai, R.L.; Shen, G.M.; Wang, H.; Guan, Y.Y. Brain functional connectivity network studies of acupuncture: A systematic review on resting-state fMRI. J. Integr. Med. 2018, 16, 26–33. [Google Scholar] [CrossRef] [PubMed]
- Maksimenko, V.A.; Runnova, A.E.; Frolov, N.S.; Makarov, V.V.; Nedaivozov, V.; Koronovskii, A.A.; Pisarchik, A.; Hramov, A.E. Multiscale neural connectivity during human sensory processing in the brain. Phys. Rev. E 2018, 97, 052405. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chang, C.; Liu, Z.; Chen, M.C.; Liu, X.; Duyn, J.H. EEG correlates of time-varying BOLD functional connectivity. Neuroimage 2013, 72, 227–236. [Google Scholar] [CrossRef] [Green Version]
- Baker, J.M.; Bruno, J.L.; Gundran, A.; Hosseini, S.H.; Reiss, A.L. fNIRS measurement of cortical activation and functional connectivity during a visuospatial working memory task. PLoS ONE 2018, 13, e0201486. [Google Scholar]
- Rieck, J.R.; DeSouza, B.; Baracchini, G.; Grady, C.L. Reduced modulation of BOLD variability as a function of cognitive load in healthy aging. Neurobiol. Aging 2022, 112, 215–230. [Google Scholar] [CrossRef]
- Yamashita, M.; Yoshihara, Y.; Hashimoto, R.; Yahata, N.; Ichikawa, N.; Sakai, Y.; Yamada, T.; Matsukawa, N.; Okada, G.; Tanaka, S.C.; et al. A prediction model of working memory across health and psychiatric disease using whole-brain functional connectivity. eLife 2018, 7, e38844. [Google Scholar] [CrossRef]
- Zuber, P.; Gaetano, L.; Griffa, A.; Huerbin, M.; Pedullà, L.; Bonzano, L.; Altermatt, A.; Tsagkas, C.; Parmar, K.; Hagmann, P.; et al. Additive and interaction effects of working memory and motor sequence training on brain functional connectivity. Sci. Rep. 2021, 11, 23089. [Google Scholar] [CrossRef]
- Bubbico, G.; Chiacchiaretta, P.; Parenti, M.; Di Marco, M.; Panara, V.; Sepede, G.; Ferretti, A.; Perrucci, M.G. Effects of second language learning on the plastic aging brain: Functional connectivity, cognitive decline, and reorganization. Front. Neurosci. 2019, 13, 423. [Google Scholar] [CrossRef]
- Marins, T.; Rodrigues, E.; Bortolini, T.; Melo, B.; Moll, J.; Tovar-Moll, F. Structural and functional connectivity changes in response to short-term neurofeedback training with motor imagery. Neuroimage 2019, 194, 283–290. [Google Scholar] [CrossRef]
- Sugata, H.; Yagi, K.; Yazawa, S.; Nagase, Y.; Tsuruta, K.; Ikeda, T.; Nojima, I.; Hara, M.; Matsushita, K.; Kawakami, K.; et al. Role of beta-band resting-state functional connectivity as a predictor of motor learning ability. NeuroImage 2020, 210, 116562. [Google Scholar] [CrossRef] [PubMed]
- Zhou, L.; Zhu, Q.; Wu, B.; Qin, B.; Hu, H.; Qian, Z. A comparison of directed functional connectivity among fist-related brain activities during movement imagery, movement execution, and movement observation. Brain Res. 2022, 1777, 147769. [Google Scholar] [CrossRef] [PubMed]
- Feng, N.; Hu, F.; Wang, H.; Zhou, B. Motor Intention Decoding from the Upper Limb by Graph Convolutional Network Based on Functional Connectivity. Int. J. Neural Syst. 2021, 31, 2150047. [Google Scholar] [CrossRef] [PubMed]
- Holiga, Š.; Hipp, J.F.; Chatham, C.H.; Garces, P.; Spooren, W.; D’Ardhuy, X.L.; Bertolino, A.; Bouquet, C.; Buitelaar, J.K.; Bours, C.; et al. Patients with autism spectrum disorders display reproducible functional connectivity alterations. Sci. Transl. Med. 2019, 11, eaat9223. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Xu, J.; Wang, C.; Xu, Z.; Li, T.; Chen, F.; Chen, K.; Gao, J.; Wang, J.; Hu, Q. Specific functional connectivity patterns of middle temporal gyrus subregions in children and adults with autism spectrum disorder. Autism Res. 2020, 13, 410–422. [Google Scholar] [CrossRef]
- Gu, Y.; Lin, Y.; Huang, L.; Ma, J.; Zhang, J.; Xiao, Y.; Dai, Z.; Initiative, A.D.N. Abnormal dynamic functional connectivity in Alzheimer’s disease. CNS Neurosci. Ther. 2020, 26, 962–971. [Google Scholar] [CrossRef]
- Smith, R.X.; Strain, J.F.; Tanenbaum, A.; Fagan, A.M.; Hassenstab, J.; McDade, E.; Schindler, S.E.; Gordon, B.A.; Xiong, C.; Chhatwal, J.; et al. Resting-state functional connectivity disruption as a pathological biomarker in autosomal dominant Alzheimer disease. Brain Connect. 2021, 11, 239–249. [Google Scholar] [CrossRef]
- Sharaev, M.; Artemov, A.; Kondrateva, E.; Ivanov, S.; Sushchinskaya, S.; Bernstein, A.; Cichocki, A.; Burnaev, E. Learning connectivity patterns via graph kernels for fmri-based depression diagnostics. In Proceedings of the 2018 IEEE International Conference on Data Mining Workshops (ICDMW), Singapore, 17–20 November 2018; pp. 308–314. [Google Scholar]
- Chen, M.H.; DeLuca, J.; Genova, H.M.; Yao, B.; Wylie, G.R. Cognitive fatigue is associated with altered functional connectivity in interoceptive and reward pathways in multiple sclerosis. Diagnostics 2020, 10, 930. [Google Scholar] [CrossRef]
- Nobukawa, S.; Kikuchi, M.; Takahashi, T. Changes in functional connectivity dynamics with aging: A dynamical phase synchronization approach. Neuroimage 2019, 188, 357–368. [Google Scholar] [CrossRef]
- Stillman, C.M.; Donofry, S.D.; Erickson, K.I. Exercise, fitness and the aging brain: A review of functional connectivity in aging. Arch. Psychol. 2019, 3. [Google Scholar] [CrossRef]
- Boa Sorte Silva, N.C.; Nagamatsu, L.S.; Gill, D.P.; Owen, A.M.; Petrella, R.J. Memory function and brain functional connectivity adaptations following multiple-modality exercise and mind–motor training in older adults at risk of dementia: An exploratory sub-study. Front. Aging Neurosci. 2020, 12, 22. [Google Scholar] [CrossRef] [PubMed]
- Larivière, S.; Xifra-Porxas, A.; Kassinopoulos, M.; Niso, G.; Baillet, S.; Mitsis, G.D.; Boudrias, M.H. Functional and effective reorganization of the aging brain during unimanual and bimanual hand movements. Hum. Brain Mapp. 2019, 40, 3027–3040. [Google Scholar] [CrossRef] [PubMed]
- Uddin, L.Q.; Yeo, B.; Spreng, R.N. Towards a universal taxonomy of macro-scale functional human brain networks. Brain Topogr. 2019, 32, 926–942. [Google Scholar] [CrossRef] [PubMed]
- Hramov, A.E.; Frolov, N.S.; Maksimenko, V.A.; Kurkin, S.A.; Kazantsev, V.B.; Pisarchik, A.N. Functional networks of the brain: From connectivity restoration to dynamic integration. Physics-Uspekhi 2021, 64, 584. [Google Scholar] [CrossRef]
- Gulbinaite, R.; van Rijn, H.; Cohen, M.X. Fronto-parietal network oscillations reveal relationship between working memory capacity and cognitive control. Front. Hum. Neurosci. 2014, 8, 761. [Google Scholar] [CrossRef] [Green Version]
- Darki, F.; Klingberg, T. The role of fronto-parietal and fronto-striatal networks in the development of working memory: A longitudinal study. Cereb. Cortex 2015, 25, 1587–1595. [Google Scholar] [CrossRef] [PubMed]
- Zanto, T.P.; Gazzaley, A. Fronto-parietal network: Flexible hub of cognitive control. Trends Cogn. Sci. 2013, 17, 602–603. [Google Scholar] [CrossRef] [Green Version]
- Markett, S.; Reuter, M.; Montag, C.; Voigt, G.; Lachmann, B.; Rudorf, S.; Elger, C.E.; Weber, B. Assessing the function of the fronto-parietal attention network: Insights from resting-state fMRI and the attentional network test. Hum. Brain Mapp. 2014, 35, 1700–1709. [Google Scholar] [CrossRef]
- Zhao, M.; Marino, M.; Samogin, J.; Swinnen, S.P.; Mantini, D. Hand, foot and lip representations in primary sensorimotor cortex: A high-density electroencephalography study. Sci. Rep. 2019, 9, 19464. [Google Scholar] [CrossRef]
- Dong, D.; Wang, Y.; Chang, X.; Luo, C.; Yao, D. Dysfunction of large-scale brain networks in schizophrenia: A meta-analysis of resting-state functional connectivity. Schizophr. Bull. 2018, 44, 168–181. [Google Scholar] [CrossRef]
- Joo, S.H.; Lim, H.K.; Lee, C.U. Three large-scale functional brain networks from resting-state functional MRI in subjects with different levels of cognitive impairment. Psychiatry Investig. 2016, 13, 1. [Google Scholar] [CrossRef] [PubMed]
- Qayyum, A.; Khan, M.A.; Mazher, M.; Suresh, M. Classification of eeg learning and resting states using 1d-convolutional neural network for cognitive load assesment. In Proceedings of the 2018 IEEE Student Conference on Research and Development (SCOReD), Bangi, Malaysia, 26–28 November 2018; pp. 1–5. [Google Scholar]
- Sagga, D.; Echtioui, A.; Khemakhem, R.; Ghorbel, M. Epileptic seizure detection using EEG signals based on 1D-CNN Approach. In Proceedings of the 2020 20th IEEE International Conference on Sciences and Techniques of Automatic Control and Computer Engineering (STA), Sfax, Tunisia, 20–22 December 2020; pp. 51–56. [Google Scholar]
- Schoffelen, J.M.; Gross, J. Source connectivity analysis with MEG and EEG. Hum. Brain Mapp. 2009, 30, 1857–1865. [Google Scholar] [CrossRef] [PubMed]
- Sosnik, R.; Zheng, L. Reconstruction of hand, elbow and shoulder actual and imagined trajectories in 3D space using EEG current source dipoles. J. Neural Eng. 2021, 18, 056011. [Google Scholar] [CrossRef]
- Frolov, N.; Maksimenko, V.; Lüttjohann, A.; Koronovskii, A.; Hramov, A. Feed-forward artificial neural network provides data-driven inference of functional connectivity. Chaos Interdiscip. J. Nonlinear Sci. 2019, 29, 091101. [Google Scholar] [CrossRef] [PubMed]
- Rulkov, N.F.; Sushchik, M.M.; Tsimring, L.S.; Abarbanel, H.D. Generalized synchronization of chaos in directionally coupled chaotic systems. Phys. Rev. E 1995, 51, 980. [Google Scholar] [CrossRef] [PubMed]
- Taud, H.; Mas, J. Multilayer perceptron (MLP). In Geomatic Approaches for Modeling Land Change Scenarios; Springer: Berlin/Heidelberg, Germany, 2018; pp. 451–455. [Google Scholar]
- Pitsik, E.; Frolov, N. Artificial neural network predicts inter-areal functional connectivity. In Saratov Fall Meeting 2020: Computations and Data Analysis: From Molecular Processes to Brain Functions; International Society for Optics and Photonics: San Diego, CA, USA, 2021; Volume 11847, p. 118470U. [Google Scholar]
- Frolov, N.S.; Pitsik, E.N.; Maksimenko, V.A.; Grubov, V.V.; Kiselev, A.R.; Wang, Z.; Hramov, A.E. Age-related slowing down in the motor initiation in elderly adults. PLoS ONE 2020, 15, e0233942. [Google Scholar] [CrossRef]
- Velasques, B.; Cagy, M.; Piedade, R.; Ribeiro, P. Sensorimotor integration and attention: An electrophysiological analysis. In Functional Brain Mapping and the Endeavor to Understand the Working Brain; IntechOpen: London, UK, 2013. [Google Scholar]
- Park, I.M.; Meister, M.L.; Huk, A.C.; Pillow, J.W. Encoding and decoding in parietal cortex during sensorimotor decision-making. Nat. Neurosci. 2014, 17, 1395–1403. [Google Scholar] [CrossRef]
- Desmurget, M.; Richard, N.; Beuriat, P.A.; Szathmari, A.; Mottolese, C.; Duhamel, J.R.; Sirigu, A. Selective inhibition of volitional hand movements after stimulation of the dorsoposterior parietal cortex in humans. Curr. Biol. 2018, 28, 3303–3309. [Google Scholar] [CrossRef] [Green Version]
- Gao, Z.; Davis, C.; Thomas, A.M.; Economo, M.N.; Abrego, A.M.; Svoboda, K.; De Zeeuw, C.I.; Li, N. A cortico-cerebellar loop for motor planning. Nature 2018, 563, 113–116. [Google Scholar] [CrossRef]
- Lee, S.H.; Jin, S.H.; An, J. The difference in cortical activation pattern for complex motor skills: A functional near-infrared spectroscopy study. Sci. Rep. 2019, 9, 14066. [Google Scholar] [CrossRef] [Green Version]
- Pitsik, E.; Frolov, N.; Hauke Kraemer, K.; Grubov, V.; Maksimenko, V.; Kurths, J.; Hramov, A. Motor execution reduces EEG signals complexity: Recurrence quantification analysis study. Chaos Interdiscip. J. Nonlinear Sci. 2020, 30, 023111. [Google Scholar] [CrossRef] [PubMed]
- Cruikshank, L.C.; Singhal, A.; Hueppelsheuser, M.; Caplan, J.B. Theta oscillations reflect a putative neural mechanism for human sensorimotor integration. J. Neurophysiol. 2012, 107, 65–77. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Van Der Cruijsen, J.; Manoochehri, M.; Jonker, Z.D.; Andrinopoulou, E.R.; Frens, M.A.; Ribbers, G.M.; Schouten, A.C.; Selles, R.W. Theta but not beta power is positively associated with better explicit motor task learning. NeuroImage 2021, 240, 118373. [Google Scholar] [CrossRef] [PubMed]
- Pellegrino, G.; Tomasevic, L.; Herz, D.M.; Larsen, K.M.; Siebner, H.R. Theta activity in the left dorsal premotor cortex during action re-evaluation and motor reprogramming. Front. Hum. Neurosci. 2018, 12, 364. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Reuter-Lorenz, P.A.; Park, D.C. How does it STAC up? Revisiting the scaffolding theory of aging and cognition. Neuropsychol. Rev. 2014, 24, 355–370. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Papitto, G.; Friederici, A.D.; Zaccarella, E. The topographical organization of motor processing: An ALE meta-analysis on six action domains and the relevance of Broca’s region. NeuroImage 2020, 206, 116321. [Google Scholar] [CrossRef]
- Solesio-Jofre, E.; Beets, I.A.; Woolley, D.G.; Pauwels, L.; Chalavi, S.; Mantini, D.; Swinnen, S.P. Age-dependent modulations of resting state connectivity following motor practice. Front. Aging Neurosci. 2018, 10, 25. [Google Scholar] [CrossRef]
- Wang, M. The Role of Dorsal Premotor Cortex in Decision-Making and Action Selection; Stanford University: Stanford, CA, USA, 2019. [Google Scholar]
- Quentin, R.; Cohen, L.G. Reversing working memory decline in the elderly. Nat. Neurosci. 2019, 22, 686–688. [Google Scholar] [CrossRef]
- Steiger, T.K.; Herweg, N.A.; Menz, M.M.; Bunzeck, N. Working memory performance in the elderly relates to theta-alpha oscillations and is predicted by parahippocampal and striatal integrity. Sci. Rep. 2019, 9, 706. [Google Scholar] [CrossRef]
- Matysiak, O.; Kroemeke, A.; Brzezicka, A. Working memory capacity as a predictor of cognitive training efficacy in the elderly population. Front. Aging Neurosci. 2019, 11, 126. [Google Scholar] [CrossRef] [Green Version]
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Pitsik, E.N.; Frolov, N.S.; Shusharina, N.; Hramov, A.E. Age-Related Changes in Functional Connectivity during the Sensorimotor Integration Detected by Artificial Neural Network. Sensors 2022, 22, 2537. https://doi.org/10.3390/s22072537
Pitsik EN, Frolov NS, Shusharina N, Hramov AE. Age-Related Changes in Functional Connectivity during the Sensorimotor Integration Detected by Artificial Neural Network. Sensors. 2022; 22(7):2537. https://doi.org/10.3390/s22072537
Chicago/Turabian StylePitsik, Elena N., Nikita S. Frolov, Natalia Shusharina, and Alexander E. Hramov. 2022. "Age-Related Changes in Functional Connectivity during the Sensorimotor Integration Detected by Artificial Neural Network" Sensors 22, no. 7: 2537. https://doi.org/10.3390/s22072537
APA StylePitsik, E. N., Frolov, N. S., Shusharina, N., & Hramov, A. E. (2022). Age-Related Changes in Functional Connectivity during the Sensorimotor Integration Detected by Artificial Neural Network. Sensors, 22(7), 2537. https://doi.org/10.3390/s22072537