
Black-Box Mathematical Model for Net Photosynthesis Estimation and Its Digital IoT Implementation Based on Non-Invasive Techniques: Capsicum annuum L. Study Case

 ,
,  ,
,  ,
,  , ,
, , 
Abstract
:1. Introduction
- Leaf temperature (Tl) is one of the main factors related to NP. The increase in photosynthetic capacity is faster as the temperature of the leaves increases.
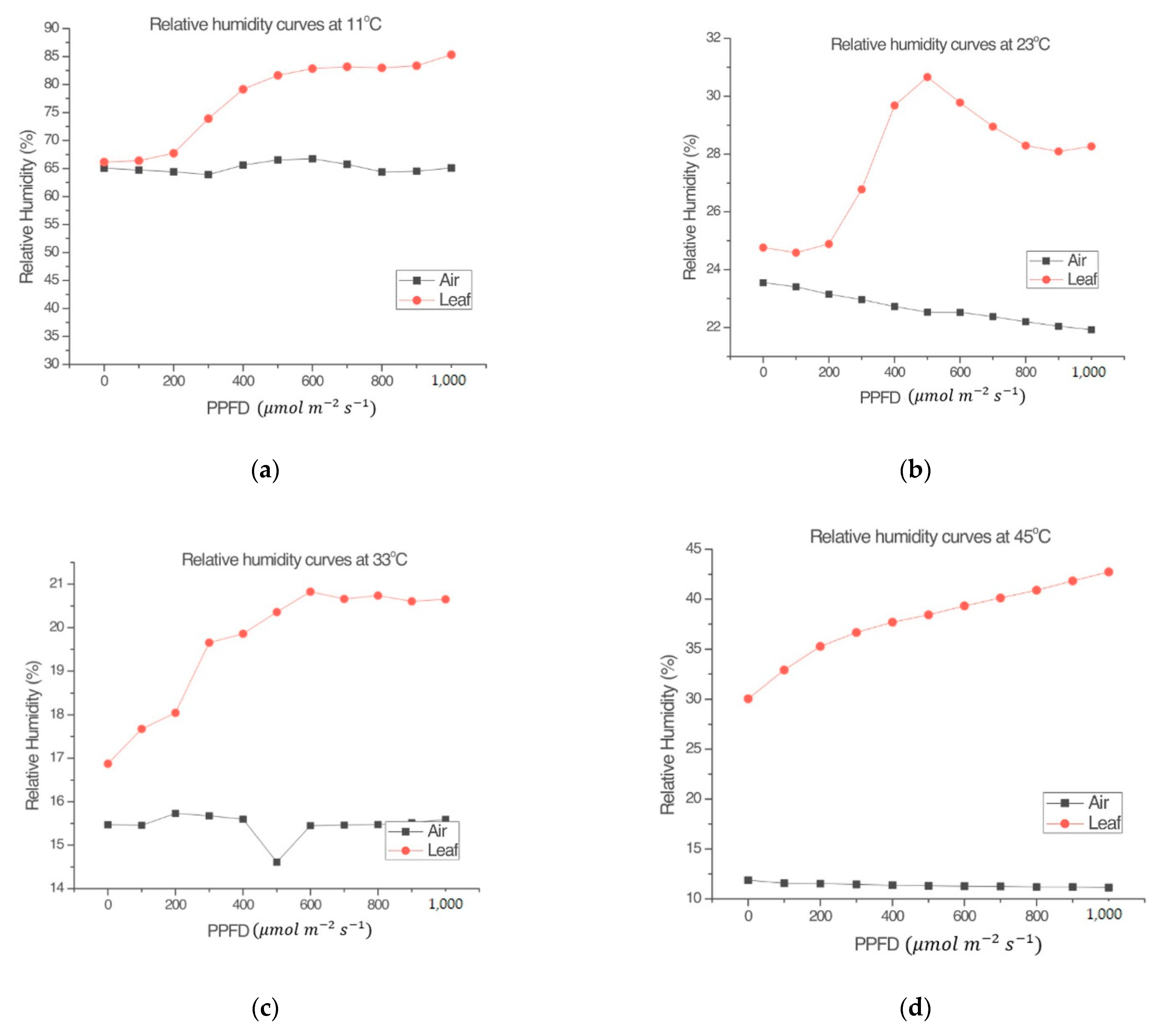
- Leaf relative humidity (RHl) is a plant response related to its transpiration.
- Incident radiation (R) is a reference for climatic conditions and a key factor for internal processes such as photosynthesis, temperature regulation, and transpiration.
- Propose a mathematical model that describes this relationship in different climatic conditions (air temperature and radiation). The mathematical model proposed in this article was obtained from measurements made in Capsicum annuum L. plants and validated in Capsicum chinense Jacq. and Capsicum annuum L. plants.
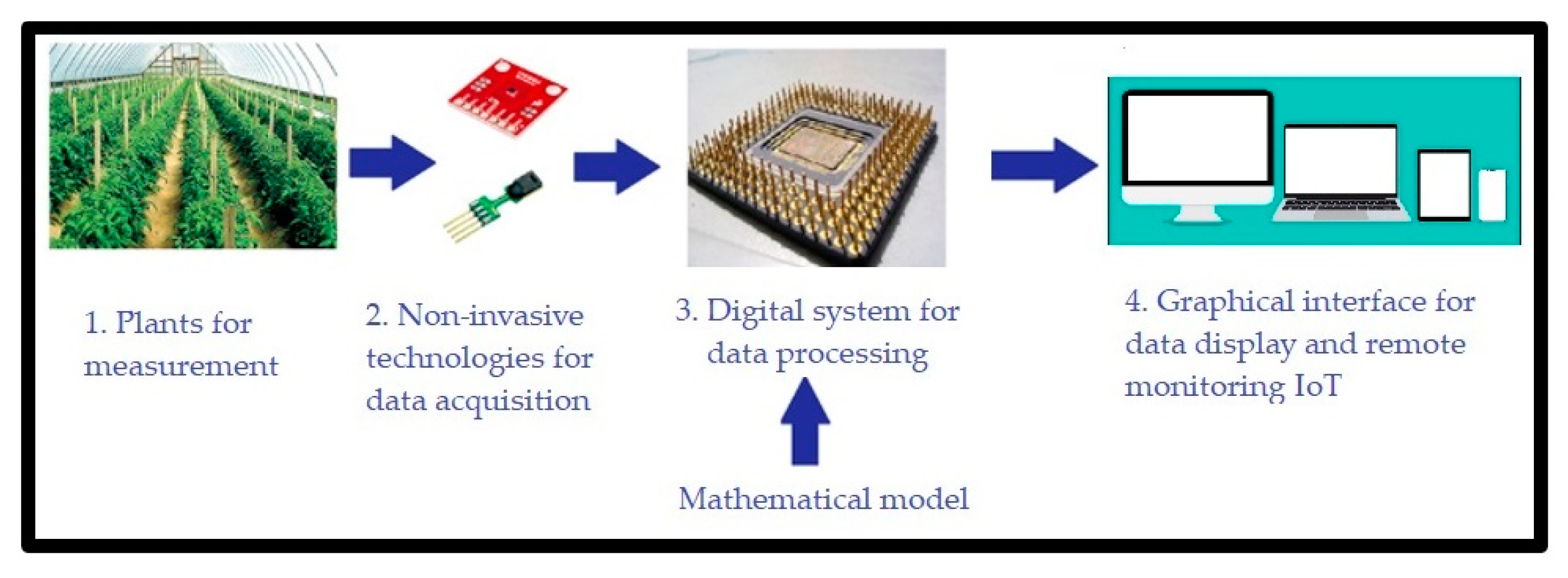
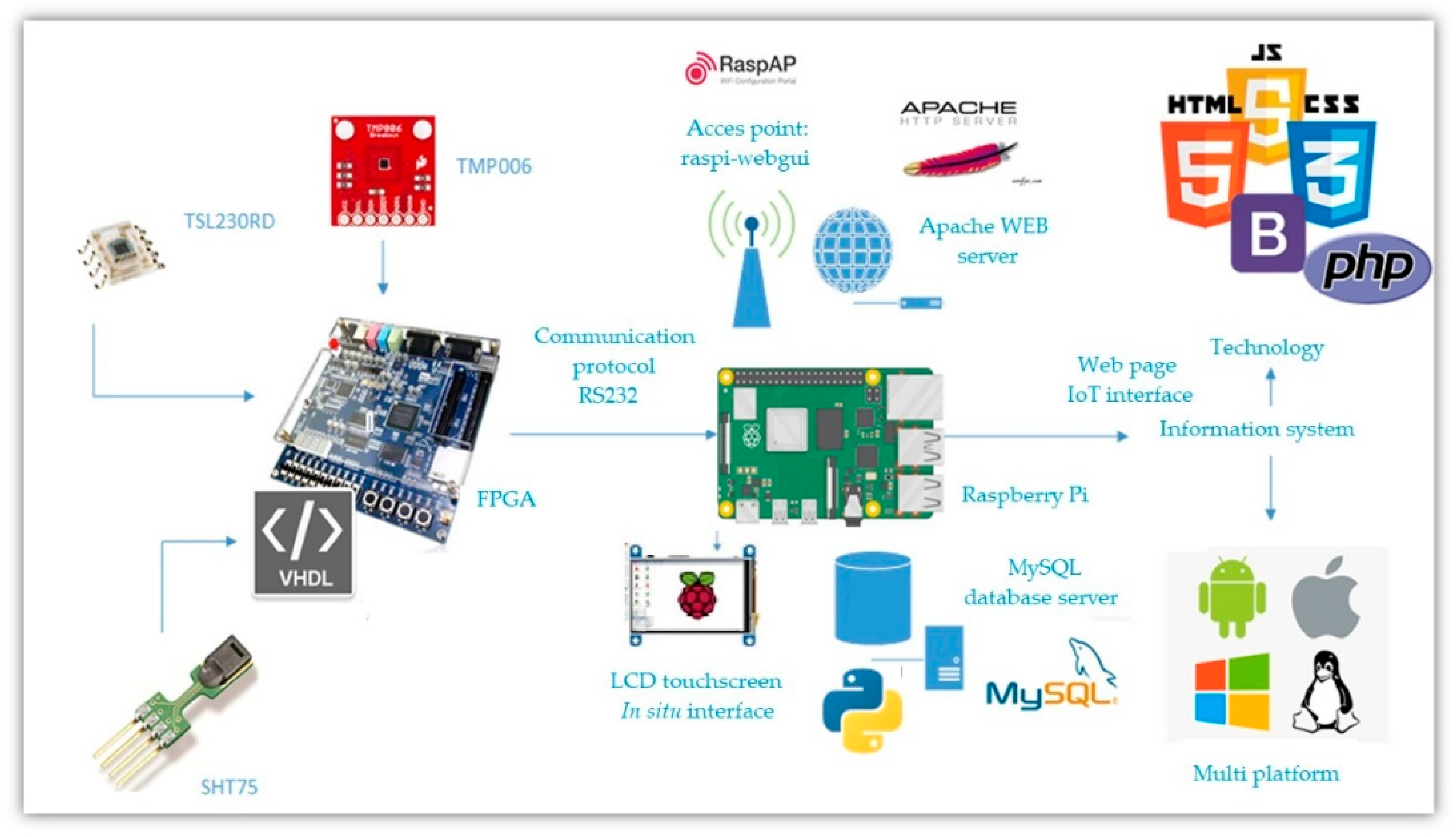
- Implement the developed mathematical model in a digital system. Such system should estimate net photosynthesis through non-invasive techniques, besides being able to work for in vivo, in situ, in vitro, portable, and on a small-scale measurement. In order to implement this, we propose the use of a digital system combined with a communication system capable of measuring greenhouse variables. The system records the measured magnitudes in a database with remote IoT access. A general scheme of this proposal is observed in Figure 1.
2. Materials and Methods
2.1. Experimentation for Data Collection
2.1.1. The Experimental Organism
- They have the C3 metabolism which is the most common type of photosynthesis [36].
- They are small and portable.
- These chili plants are the most widespread and cultivated species in subtropical and temperate countries. They are produced all year round, and they are cultivated all over the world.
- These chili plants are mainly used for food preparation due to their taste and nutritional properties, but they are also used in the pharmaceutical, cosmetic, and military industries around the world [37].
2.1.2. Gas Analysis, Leaf Temperature, Relative Humidity, and Incident Radiation Measurements
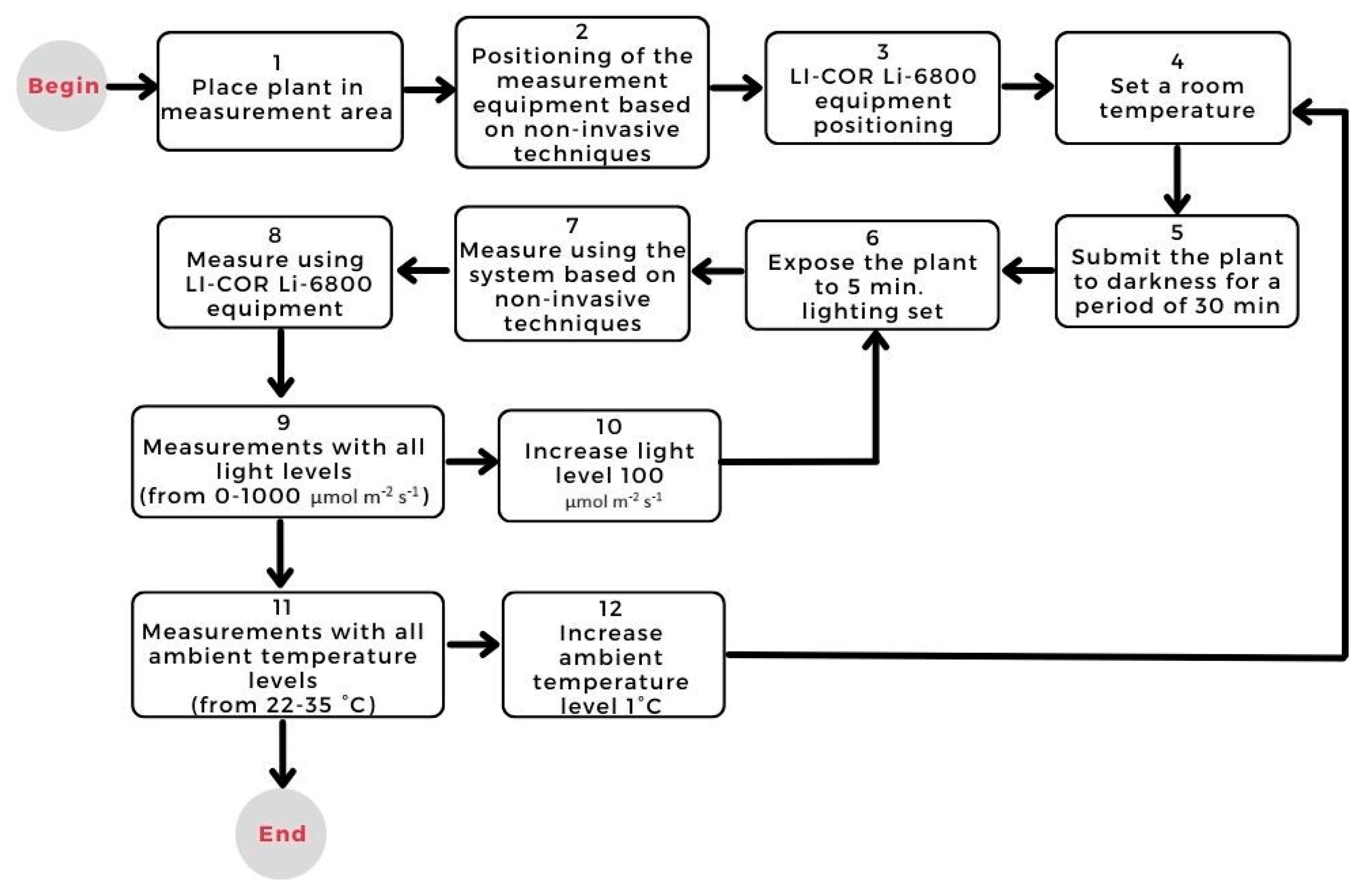
2.1.3. Stabilization Process
2.1.4. Light–Response Curves
2.1.5. Response Curves to Reference Light
2.2. Mathematical Model
2.3. Implementation in Digital System
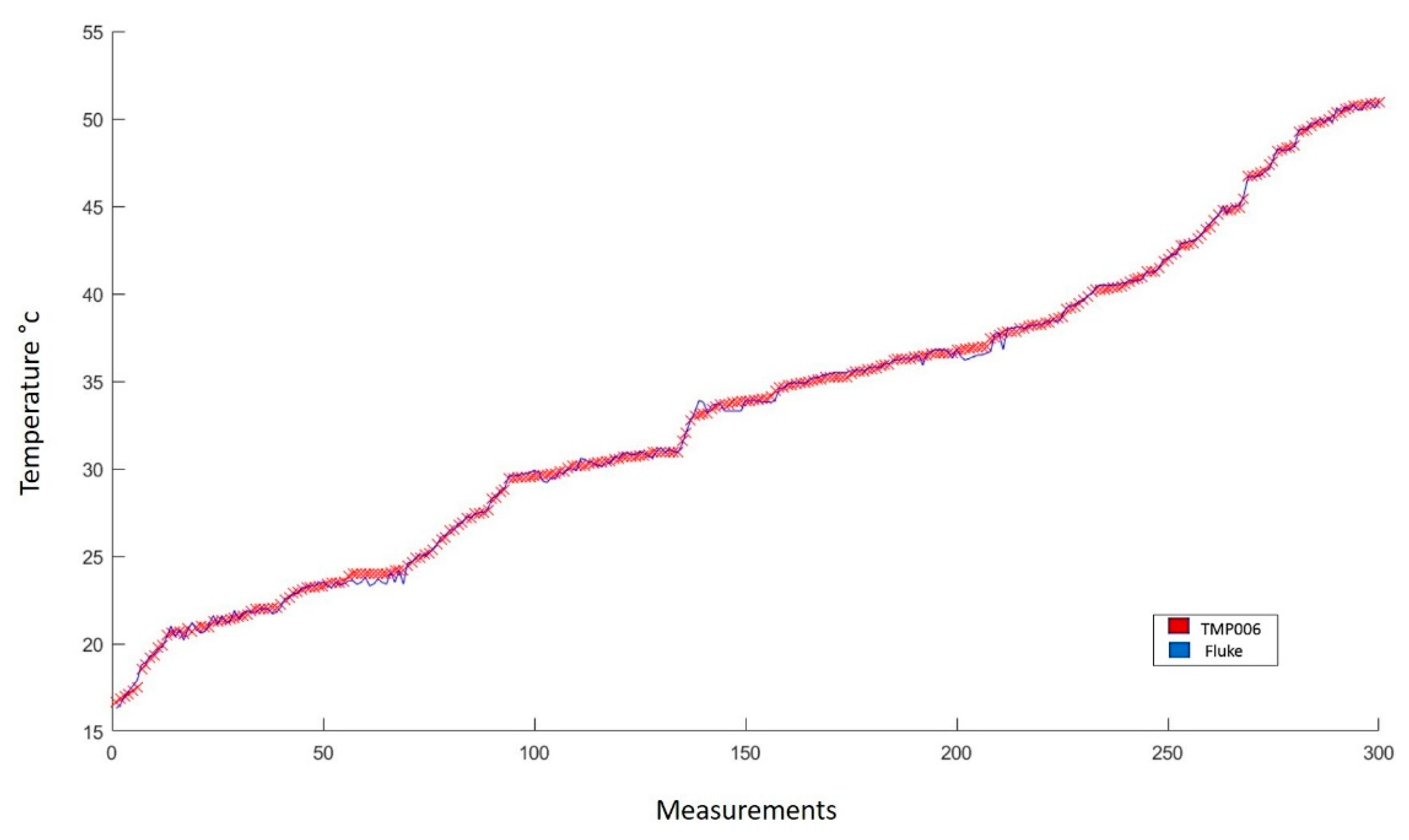
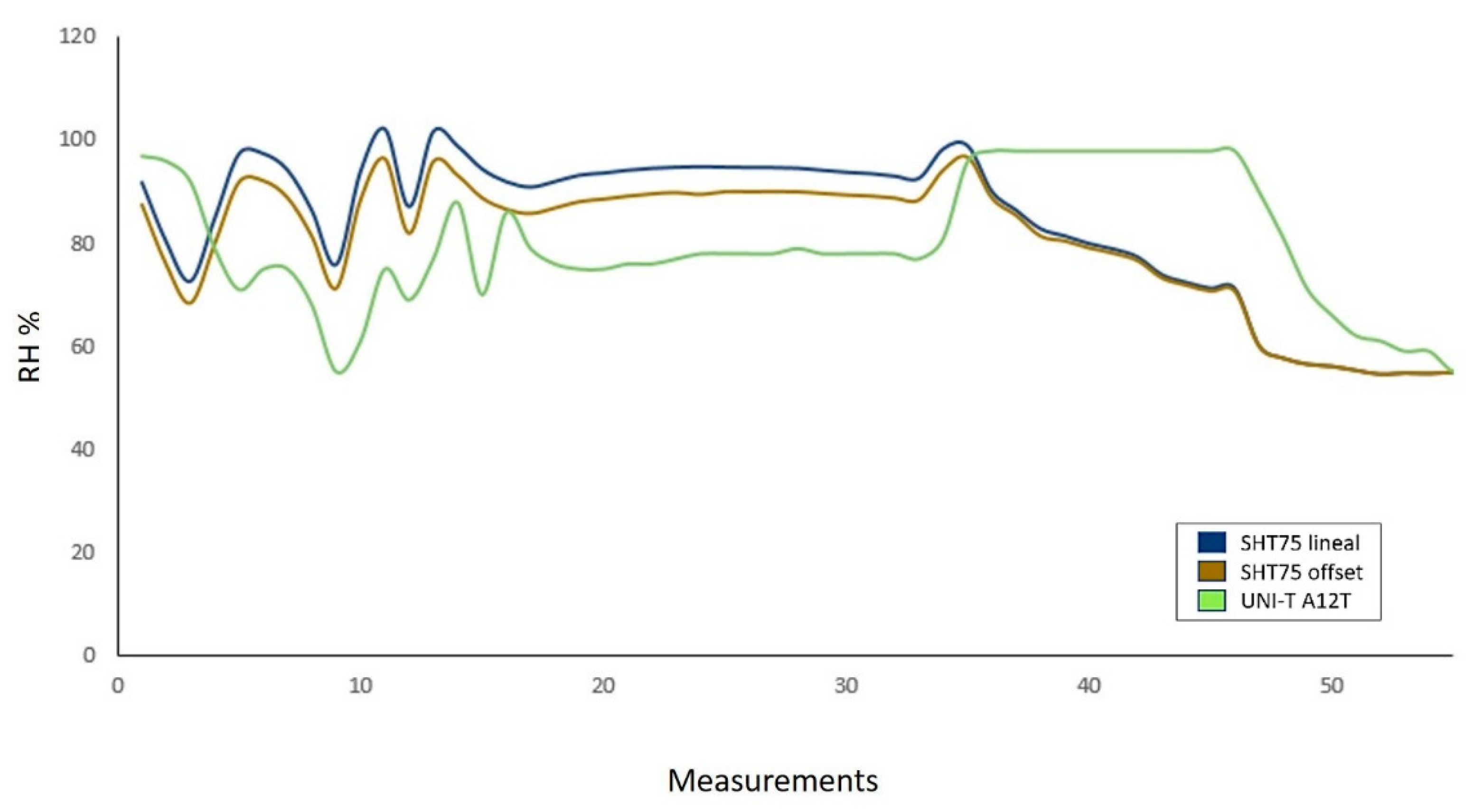
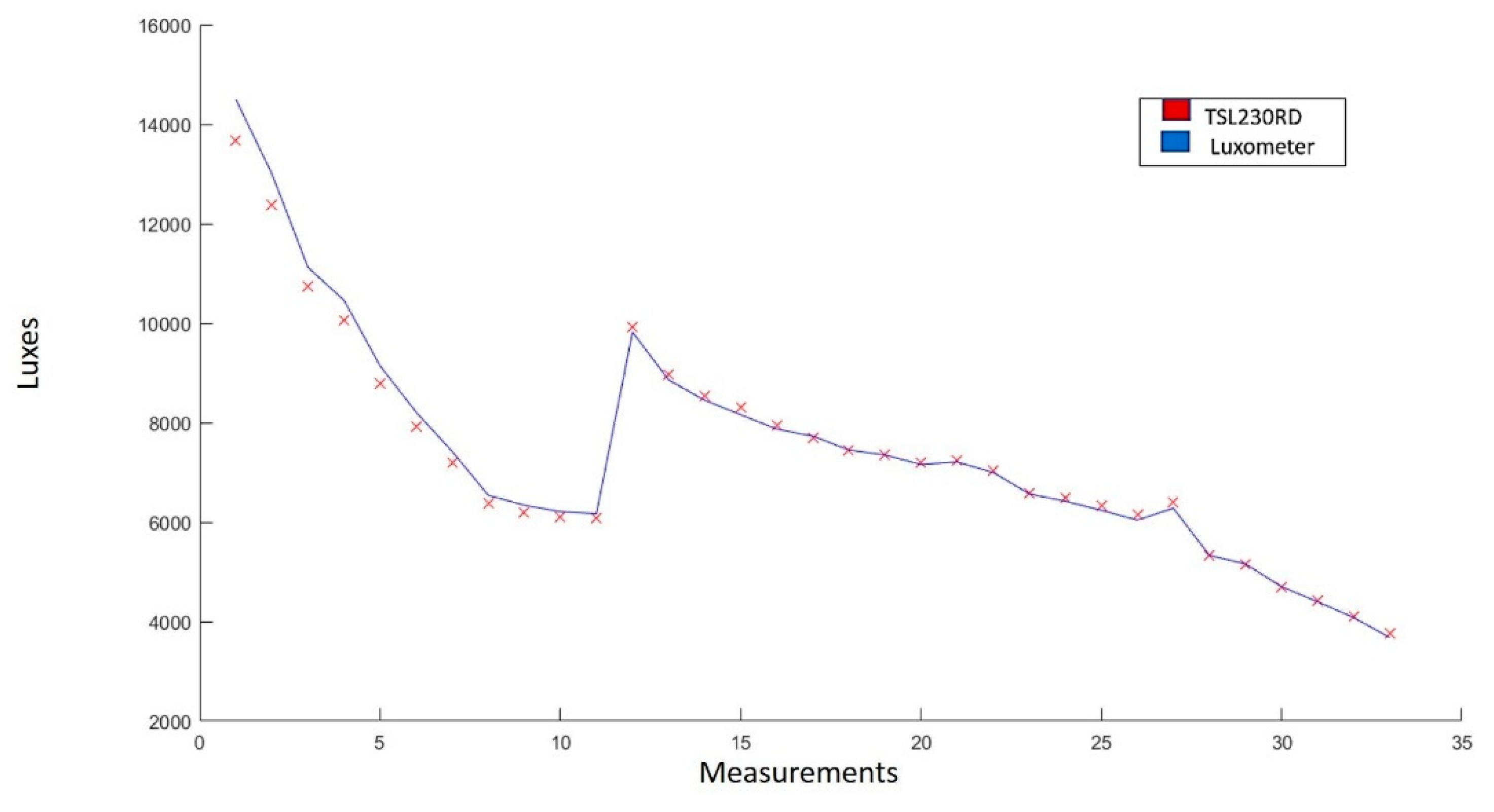
2.3.1. Sensors
2.3.2. Digital System
2.4. IoT
2.5. Methodology for the Mathematical Model
- Experimental conditions of the case study plant (Section 2.1.1).
- Steps to homogenize the net photosynthesis estimate of the case study plants (Section 2.1.3).
- Steps to obtain the measurements of Ta, Tl, R, RHa, RHl, and NP (Section 2.1.4) in both plants in the case study, Capsicum annuum L. and Capsicum chinense Jacq. The measurements were obtained with the LI-COR Li-6800.
- Steps to demonstrate that the LI-COR Li-6800 lamp, by itself, does not affect the measurements obtained (Section 2.1.5) [4].
- The general description for the mathematical model generation (Section 2.2).
- Once the variables of interest were obtained, a set of measurements was selected (5 plants with 5 repetitions each) of Capsicum annuum L. to generate the mathematical model.
- To validate the obtained model, it was compared with another group of measurements in Capsicum annuum L. and Capsicum chinense Jacq. (5 plants of each species, with 5 re-requests each).
2.6. Methodology for Implementation in a Digital System
- Communication and information processing of each of the sensors (temperature, relative humidity, and lighting) to obtain the measurements required by the mathematical model.
- Implementation of the black-box mathematical model through the hardware description to estimate net photosynthesis.
- Synchronization of each device for communication, operation, storage, and transmission.
- Creation of a graphical interface to make it easier for the user to understand the data acquired by the sensors, as well as the estimation of net photosynthesis determined by the mathematical model.
- Communication between the FPGA and Raspberry Pi following the structure that is applied for an I2C protocol.
- Synchronization and transmission between the FPGA and Raspberry Pi so that the information from the sensors is displayed through the IoT interface.
- Unilateral transmission, for sending serial data to the Raspberry Pi through a UART pin. For serial transmission (Tx), the data contained in a vector is received as input, which is then broken down and sent serially in a timed manner.
2.7. Methodology for Digital IoT Implementation
- Reception of the data in the Raspberry Pi to be read and stored in a corresponding matrix, to be later characterized by means of the Python programming language [67].
- Log storage, using the MySQL database manager [68]. The saved data contains an id, date, time, value, and the user who made it.
- Sensor ID, date, time, value, and number of measurements are uploaded to the phpMyAdmin manager by accessing the local host from a web browser [69].
- Use of the Bootstrap libraries [70] to develop the website (https://fotosintesisproject.000webhostapp.com/, accessed on 2 May 2022) where the data of all greenhouse variables and the estimation of net photosynthesis will be displayed, as well as its graphic behavior.
- Adaptation of a 7-inch LCD screen, to visualize in situ the information from the sensors and the net photosynthesis estimation.
2.8. Plant Experimentation
3. Results
3.1. Experimentation for Data Collection
3.1.1. Light–Response Curves
3.1.2. Reference Light–Response Curves
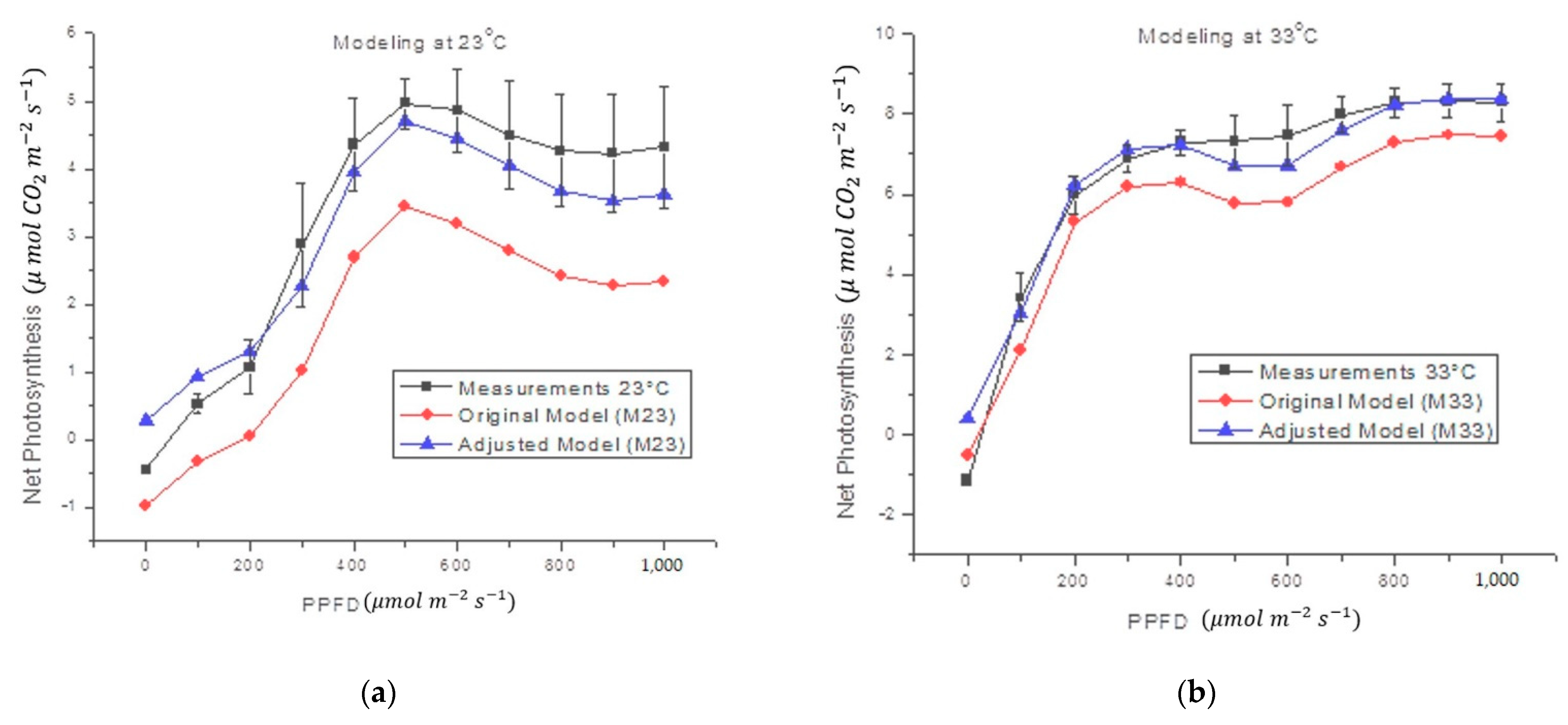
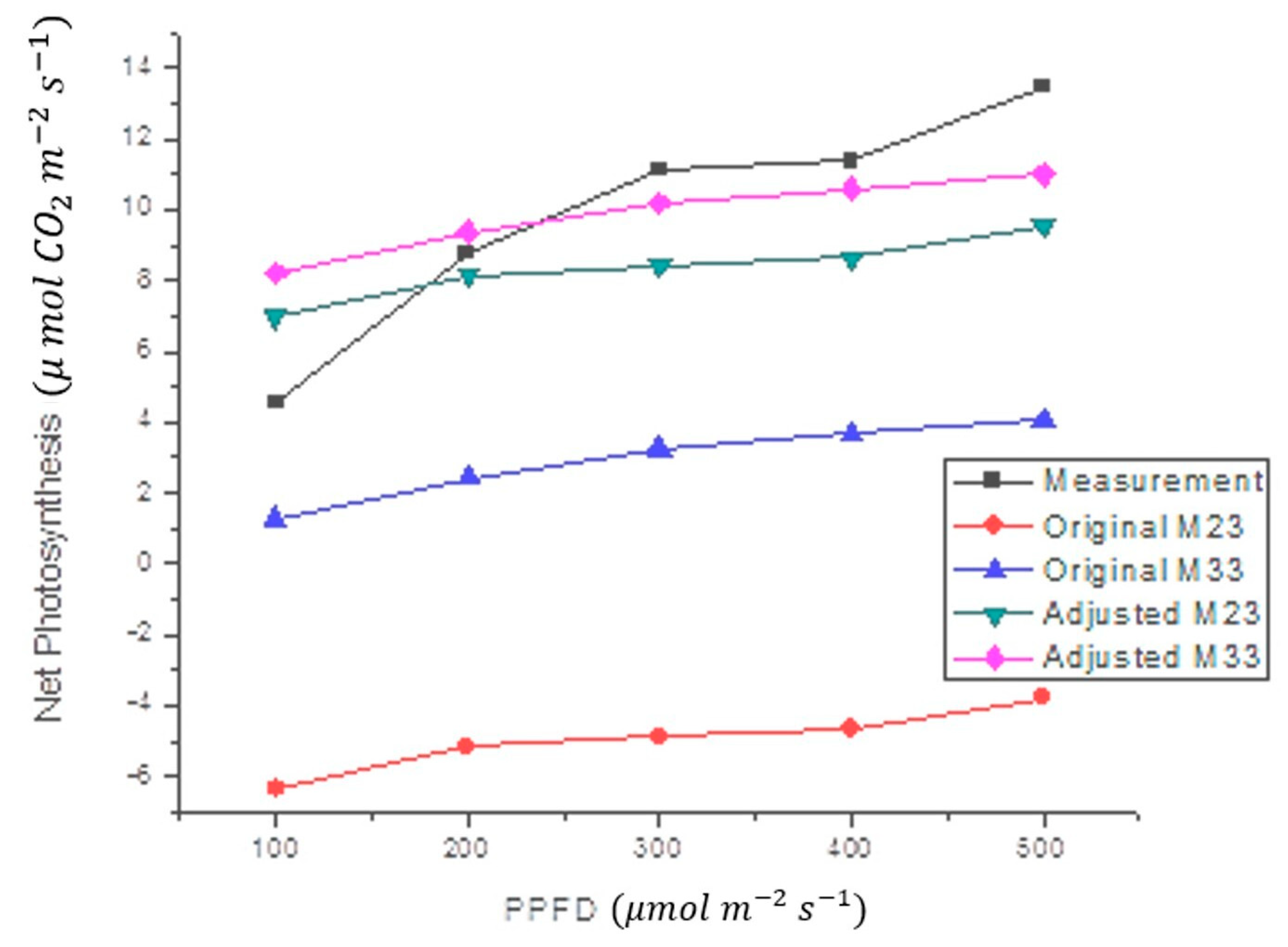
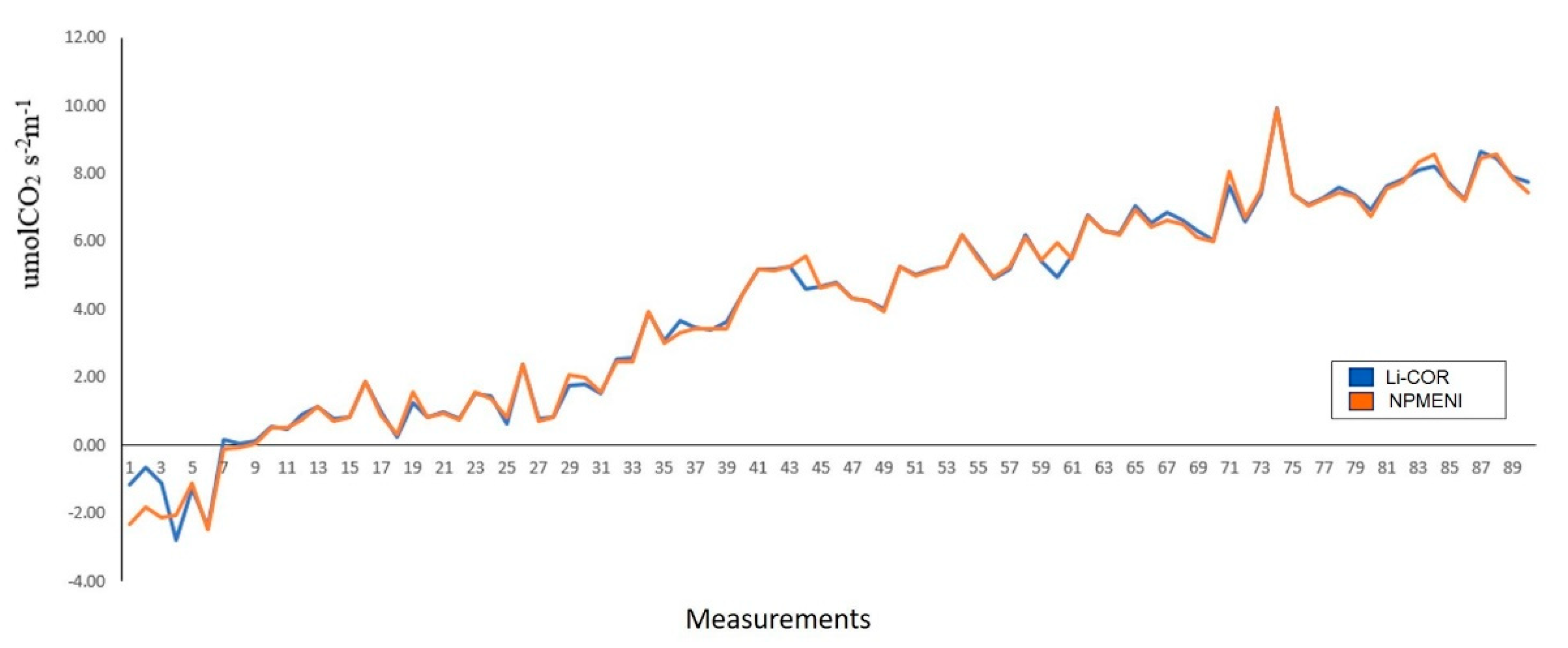
3.2. Mathematical Model
3.3. Implementation in Digital System
3.3.1. Sensors
3.3.2. Digital System
3.4. IoT
3.5. Plant Experimentation
4. Discussion
5. Conclusions
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Vázquez-Cruz, M.A.; Espinosa-Calderón, A.; Jiménez-Sánchez, A.R.; Guzmán-Cruz, R. Mathematical Modeling of Biosystems. In Biosystems Engineering: Biofactories for Food Production in the Century XXI; Guevara-Gonzalez, R., Torres-Pacheco, I., Eds.; Springer International Publishing: Cham, Switzerland, 2014; pp. 51–76. ISBN 978-3-319-03880-3. [Google Scholar]
- Guzmán-Cruz, R.; Castañeda-Miranda, R.; García-Escalante, J.J.; López-Cruz, I.L.; Lara-Herrera, A.; de la Rosa, J.I. Calibration of a Greenhouse Climate Model Using Evolutionary Algorithms. Biosyst. Eng. 2009, 104, 135–142. [Google Scholar] [CrossRef]
- Farquhar, G.D.; von Caemmerer, S.; Berry, J.A. A Biochemical Model of Photosynthetic CO2 Assimilation in Leaves of C3 Species. Planta 1980, 149, 78–90. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Espinosa-Calderon, A.; Torres-Pacheco, I.; Padilla-Medina, J.A.; Chavaro-Ortiz, M.; Xoconostle-Cazares, B.; Gomez-Silva, L.; Ruiz-Medrano, R.; Guevara-Gonzalez, R.G. Relationship between Leaf Temperature and Photosynthetic Carbon in Capsicum Annuum L. in Controlled Climates. J. Sci. Ind. Res. 2012, 71, 528–533. [Google Scholar]
- García-Camacho, F.; Sánchez-Mirón, A.; Molina-Grima, E.; Camacho-Rubio, F.; Merchuck, J.C. A Mechanistic Model of Photosynthesis in Microalgae Including Photoacclimation Dynamics. J. Theor. Biol. 2012, 304, 1–15. [Google Scholar] [CrossRef]
- Chen, D.-X.; Coughenour, M.B.; Knapp, A.K.; Owensby, C.E. Mathematical Simulation of C4 Grass Photosynthesis in Ambient and Elevated CO2. Ecol. Model. 1994, 73, 63–80. [Google Scholar] [CrossRef]
- Zufferey, V.; Murisier, F.; Schultz, H. A Model Analysis of the Photosynthetic Response of Vitis vinifera L. Cvs Riesling and Chasselas Leaves in the Field: I. Interaction of Age, Light and Temperature. Vitis 2000, 39, 19–26. [Google Scholar]
- Boonen, C.; Samson, R.; Janssens, K.; Pien, H.; Lemeur, R.; Berckmans, D. Scaling the Spatial Distribution of Photosynthesis from Leaf to Canopy in a Plant Growth Chamber. Ecol. Model. 2002, 156, 201–212. [Google Scholar] [CrossRef]
- Ye, Z.P. A New Model for Relationship between Irradiance and the Rate of Photosynthesis in Oryza Sativa. Photosynthetica 2007, 45, 637–640. [Google Scholar] [CrossRef]
- Bernacchi, C.J.; Rosenthal, D.M.; Pimentel, C.; Long, S.P.; Farquhar, G.D. Modeling the Temperature Dependence of C3 Photosynthesis. In Photosynthesis In Silico: Understanding Complexity from Molecules to Ecosystems; Laisk, A., Nedbal, L., Govindjee, Eds.; Advances in Photosynthesis and Respiration; Springer: Dordrecht, The Netherlands, 2009; pp. 231–246. ISBN 978-1-4020-9237-4. [Google Scholar]
- LI-COR. Using the LI-6800, Portable Photosynthesis: User Manual. LI-6800 Instruction Manuals. Available online: https://www.licor.com/env/support/LI-6800/manuals.html (accessed on 11 February 2022).
- Müller, J.; Braune, H.; Diepenbrock, W. Complete Parameterization of Photosynthesis Models—An Example for Barley. In Proceedings of the Crop Modeling and Decision Support; Cao, W., White, J.W., Wang, E., Eds.; Springer: Berlin/Heidelberg, Germany, 2009; pp. 12–23. [Google Scholar]
- Johnson, I.R.; Thornley, J.H.M.; Frantz, J.M.; Bugbee, B. A Model of Canopy Photosynthesis Incorporating Protein Distribution through the Canopy and Its Acclimation to Light, Temperature and CO2. Ann. Bot. 2010, 106, 735–749. [Google Scholar] [CrossRef] [Green Version]
- Lombardozzi, D.; Sparks, J.P.; Bonan, G.; Levis, S. Ozone Exposure Causes a Decoupling of Conductance and Photosynthesis: Implications for the Ball-Berry Stomatal Conductance Model. Oecologia 2012, 169, 651–659. [Google Scholar] [CrossRef]
- Von Caemmerer, S. Steady-State Models of Photosynthesis. Plant Cell Environ. 2013, 36, 1617–1630. [Google Scholar] [CrossRef] [PubMed]
- Serbin, S.P.; Singh, A.; Desai, A.R.; Dubois, S.G.; Jablonski, A.D.; Kingdon, C.C.; Kruger, E.L.; Townsend, P.A. Remotely Estimating Photosynthetic Capacity, and Its Response to Temperature, in Vegetation Canopies Using Imaging Spectroscopy. Remote Sens. Environ. 2015, 167, 78–87. [Google Scholar] [CrossRef]
- Janka, E.; Körner, O.; Rosenqvist, E.; Ottosen, C.-O. A Coupled Model of Leaf Photosynthesis, Stomatal Conductance, and Leaf Energy Balance for Chrysanthemum (Dendranthema grandiflora). Comput. Electron. Agric. 2016, 123, 264–274. [Google Scholar] [CrossRef]
- Field, C.B.; Ball, J.T.; Berry, J.A. Photosynthesis: Principles and Field Techniques. In Plant Physiological Ecology: Field Methods and Instrumentation; Pearcy, R.W., Ehleringer, J.R., Mooney, H.A., Rundel, P.W., Eds.; Springer: Dordrecht, The Netherlands, 2000; pp. 209–253. ISBN 978-94-010-9013-1. [Google Scholar]
- Wolfbeis, O.S. Materials for Fluorescence-Based Optical Chemical Sensors. J. Mater. Chem. 2005, 15, 2657–2669. [Google Scholar] [CrossRef]
- Millan-Almaraz, J.R.; Guevara-Gonzalez, R.G.; Troncoso, R.R.; Osornio-Rios, R.A.; Torres-Pacheco, I. Advantages and Disadvantages on Photosynthesis Measurement Techniques: A Review. Afr. J. Biotechnol. 2009, 8, 7340–7349. [Google Scholar] [CrossRef]
- Hunt, S. Measurements of Photosynthesis and Respiration in Plants. Physiol. Plant. 2003, 117, 314–325. [Google Scholar] [CrossRef]
- Schulze, E.-D. A New Type of Climatized Gas Exchange Chamber for Net Photosynthesis and Transpiration Measurements in the Field. Oecologia 1972, 10, 243–251. [Google Scholar] [CrossRef]
- Takahashi, M.; Ishiji, T.; Kawashima, N. Handmade Oxygen and Carbon Dioxide Sensors for Monitoring the Photosynthesis Process as Instruction Material for Science Education. Sens. Actuators B Chem. 2001, 77, 237–243. [Google Scholar] [CrossRef]
- Kawachi, N.; Sakamoto, K.; Ishii, S.; Fujimaki, S.; Suzui, N.; Ishioka, N.S.; Matsuhashi, S. Kinetic Analysis of Carbon-11-Labeled Carbon Dioxide for Studying Photosynthesis in a Leaf Using Positron Emitting Tracer Imaging System. IEEE Trans. Nucl. Sci. 2006, 53, 2991–2997. [Google Scholar] [CrossRef]
- Taiz, L.; Zeiger, E.; Møller, I.M.; Murphy, A. Plant Physiology and Development; University of California: Santa Cruz, CA, USA, 2015. [Google Scholar]
- Hermand, J.-P. Photosynthesis of Seagrasses Observed in Situ from Acoustic Measurements. In Proceedings of the Oceans ’04 MTS/IEEE Techno-Ocean ’04, Kobe, Japan, 9–12 November 2004; IEEE Cat. No.04CH37600. Volume 1, pp. 433–437. [Google Scholar]
- Azcon-Bieto, J.; Talón, M. Fundamentos de Fisiología Vegetal; McGraw-Hill: New York, NY, USA, 2000. [Google Scholar]
- Espinosa-Calderon, A.; Torres-Pacheco, I.; Padilla-Medina, J.A.; Osornio-Rios, R.A.; Romero-Troncoso, R.D.J.; Villaseñor-Mora, C.; Rico-Garcia, E.; Guevara-Gonzalez, R.G. Description of Photosynthesis Measurement Methods in Green Plants Involving Optical Techniques, Advantages and Limitations. Int. J. Anim. Breed. Genet. 2014, 4, 1–10. [Google Scholar]
- Yamori, W. Photosynthetic Response to Fluctuating Environments and Photoprotective Strategies under Abiotic Stress. J. Plant Res. 2016, 129, 379–395. [Google Scholar] [CrossRef] [PubMed]
- McCree, K.J. Test of Current Definitions of Photosynthetically Active Radiation against Leaf Photosynthesis Data. Agric. Meteorol. 1972, 10, 443–453. [Google Scholar] [CrossRef]
- Han, B.-P. Photosynthesis–Irradiance Response at Physiological Level: A Mechanistic Model. J. Theor. Biol. 2001, 213, 121–127. [Google Scholar] [CrossRef] [PubMed]
- Vega, M.C.; Vivas, P.O.; Rios, C.M.; Luis, C.G.; Martín, B.C.; Seco, A.H. Las Tecnologías IOT Dentro de la Industria Conectada: Internet of Things; EOI Escuela de Organización Industrial: Madrid, Spain, 2015. [Google Scholar]
- Hamerlynck, E.P.; Knapp, A.K. Leaf-Level Responses to Light and Temperature in Two Co-Occurring Quercus (Fagaceae) Species: Implications for Tree Distribution Patterns. For. Ecol. Manag. 1994, 68, 149–159. [Google Scholar] [CrossRef]
- Sage, R.F.; Kubien, D.S. The Temperature Response of C3 and C4 Photosynthesis. Plant Cell Environ. 2007, 30, 1086–1106. [Google Scholar] [CrossRef]
- Helliker, B.R.; Richter, S.L. Subtropical to Boreal Convergence of Tree-Leaf Temperatures. Nature 2008, 454, 511–514. [Google Scholar] [CrossRef]
- West-Eberhard, M.J.; Smith, J.A.C.; Winter, K. Photosynthesis, Reorganized. Science 2011, 332, 311–312. [Google Scholar] [CrossRef]
- Kothari, S.L.; Joshi, A.; Kachhwaha, S.; Ochoa-Alejo, N. Chilli Peppers—A Review on Tissue Culture and Transgenesis. Biotechnol. Adv. 2010, 28, 35–48. [Google Scholar] [CrossRef]
- Steiner, A.A. The Universal Nutrient Solution. In Proceedings of the Sixth International Congress on Soilless Culture, Lunteren, The Netherlands, 29 April–5 May 1984. [Google Scholar]
- Singh, M.P.; Erickson, J.E.; Boote, K.J.; Tillman, B.L.; van Bruggen, A.H.C.; Jones, J.W. Photosynthetic Consequences of Late Leaf Spot Differ between Two Peanut Cultivars with Variable Levels of Resistance. Crop Sci. 2011, 51, 2741–2748. [Google Scholar] [CrossRef]
- Snider, J.L.; Choinski, J.S.; Wise, R.R. Juvenile Rhus Glabra Leaves Have Higher Temperatures and Lower Gas Exchange Rates than Mature Leaves When Compared in the Field during Periods of High Irradiance. J. Plant Physiol. 2009, 166, 686–696. [Google Scholar] [CrossRef]
- Caron Environmental Test Chambers. Available online: https://www.manualslib.com/manual/1370415/Caron-6010.html?page=2#manual (accessed on 15 February 2022).
- Tap, F. Economics-Based Optimal Control of Greenhouse Tomato Crop Production; Wageningen University and Research: Wageninge, The Netherlands, 2000. [Google Scholar]
- Van Henten, E.J. Greenhouse Climate Management: An Optical Control Approach; Wageningen University and Research: Wageninge, The Netherlands, 1994; ISBN 978-90-5485-321-3. [Google Scholar]
- Eiben, A.E.; Smith, J.E. Introduction to Evolutionary Computing; Natural Computing Series; Springer: Berlin/Heidelberg, Germany, 2015; ISBN 978-3-662-44873-1. [Google Scholar]
- Michalewicz, Z.; Fogel, D.B. How to Solve It: Modern Heuristics; Springer Science & Business Media: New York, NY, USA, 2013; ISBN 978-3-662-07807-5. [Google Scholar]
- The MathWorks, Inc. MATLAB—El Lenguaje del Cálculo Técnico. Available online: https://la.mathworks.com/products/matlab.html (accessed on 18 February 2022).
- Moore, H. MATLAB for Engineers, 3rd ed.; Pearson Prentice Hall: Boston, MA, USA, 2012; ISBN 978-0-13-210325-1. [Google Scholar]
- Texas Instruments Incorporated, TMP006/B Infrared Thermopile Sensor in Chip-Scale Package. 2015. Available online: https://www.findic.us/doc/browser/bLXq9kwex?doc_id=91490930#locale=en-US (accessed on 20 June 2021).
- Sensirion, Datasheet SHT7x (SHT71, SHT75) Humidity and Temperature Sensor IC. 2011. Available online: https://www.mouser.com/datasheet/2/682/Sensirion_Humidity_SHT7x_Datasheet_V5-469726.pdf (accessed on 1 January 2021).
- Texas Advanced Optoelectronic Solutions, TSL230RD, TSL230ARD, TSL230BRD, Programmable Light-Frequency Converters. 2007. Available online: https://www.mouser.com/datasheet/2/588/TSL230RDTSL230ARDTSL230BRD-P-519226.pdf (accessed on 8 March 2022).
- UNI-T. UNI-T Voltage Meter, Multimeter, Oscilloscope. Available online: https://www.uni-trend.com/meters/html/product/Environmental/Environmental_Tester/A10T_Temperature/A12T.html (accessed on 8 March 2022).
- Fluke Corporation. Termómetro de Infrarrojos sin Contacto. Termómetros IR de Contacto. Available online: https://www.fluke.com/es-mx/productos/medicion-de-temperatura/termometros-infrarrojos (accessed on 8 March 2022).
- Electronica Steren. Medidor Digital de Luminosidad (Luxómetro). Available online: https://www.steren.com.mx/medidor-digital-de-luminosidad-luxometro-her-408.html (accessed on 8 March 2022).
- Millan-Almaraz, J.R.; Torres-Pacheco, I.; Duarte-Galvan, C.; Guevara-Gonzalez, R.G.; Contreras-Medina, L.M.; Romero-Troncoso, R.D.J.; Rivera-Guillen, J.R. FPGA-Based Wireless Smart Sensor for Real-Time Photosynthesis Monitoring. Comput. Electron. Agric. 2013, 95, 58–69. [Google Scholar] [CrossRef]
- Duarte-Galvan, C.; Romero-Troncoso, R.D.J.; Torres-Pacheco, I.; Guevara-Gonzalez, R.G.; Fernandez-Jaramillo, A.A.; Contreras-Medina, L.M.; Carrillo-Serrano, R.V.; Millan-Almaraz, J.R. FPGA-Based Smart Sensor for Drought Stress Detection in Tomato Plants Using Novel Physiological Variables and Discrete Wavelet Transform. Sensors 2014, 14, 18650–18669. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Millan-Almaraz, J.R.; Romero-Troncoso, R.D.J.; Guevara-Gonzalez, R.G.; Contreras-Medina, L.M.; Carrillo-Serrano, R.V.; Osornio-Rios, R.A.; Duarte-Galvan, C.; Rios-Alcaraz, M.A.; Torres-Pacheco, I. FPGA-Based Fused Smart Sensor for Real-Time Plant-Transpiration Dynamic Estimation. Sensors 2010, 10, 8316–8331. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fernandez-Jaramillo, A.A.; Romero-Troncoso, R.D.J.; Duarte-Galvan, C.; Torres-Pacheco, I.; Guevara-Gonzalez, R.G.; Contreras-Medina, L.M.; Herrera-Ruiz, G.; Millan-Almaraz, J.R. FPGA-Based Chlorophyll Fluorescence Measurement System with Arbitrary Light Stimulation Waveform Using Direct Digital Synthesis. Measurement 2015, 75, 12–22. [Google Scholar] [CrossRef]
- Altera Corporation. Altera Cyclone II Reference Manual (Page 14 of 56). ManualsLib. Available online: https://www.manualslib.com/manual/1497884/Altera-Cyclone-Ii.html?page=14#manual (accessed on 18 February 2022).
- Raspberry Pi Foundation. Raspberry Pi Documentation. Available online: https://www.raspberrypi.com/documentation/ (accessed on 18 February 2022).
- Calderón, A.E.; Mendoza, B.G.R.; Del Carmen García Rodríguez, L.; López Romero, F.A.; Olivarez, J.P.; Licea, M.A.R.; Carmona, J.D.; Cruz, R.G.; Tavera, V.M. A Reconfigurable IoT System for the Measurement of Greenhouse Variables. In Proceedings of the 2021 IEEE International Autumn Meeting on Power, Electronics and Computing (ROPEC), Ixtapa, Mexico, 10–12 November 2021; Volume 5, pp. 1–7. [Google Scholar]
- Barba, C.J.B. Seguimiento de datos en tiempo real con Apache Kafka en Raspberry pi 3; Caso Práctico: IoT Floricultura. In Proceedings of the [2019-MADRID] Congreso Internacional de Tecnología, Ciencia y Sociedad, Madrid, Spain, 3–4 April 2019. [Google Scholar]
- Priyadarshana, K.; Manchanayaka, M.A.L.S.K.; Sudantha, B.H. IoT Based Greenhouse System for Tropical Countries. In Proceedings of the 2020 International Conference on Image Processing and Robotics (ICIP), Negombo, Srilanka, 6–8 March 2020; pp. 1–5. [Google Scholar]
- Widyawati, D.K.; Ambarwari, A.; Wahyudi, A. Design and Prototype Development of Internet of Things for Greenhouse Monitoring System. In Proceedings of the 2020 3rd International Seminar on Research of Information Technology and Intelligent Systems (ISRITI), Yogyakarta, Indonesia, 10 December 2020; pp. 389–393. [Google Scholar]
- Danita, M.; Mathew, B.; Shereen, N.; Sharon, N.; Paul, J.J. IoT Based Automated Greenhouse Monitoring System. In Proceedings of the 2018 Second International Conference on Intelligent Computing and Control Systems (ICICCS), Madurai, India, 14–15 June 2018; pp. 1933–1937. [Google Scholar]
- Saboya, N.G.F. Normas de Comunicación en Serie: RS-232, RS-422 y RS-485. Rev. Ingenio Libre 2012, 9, 86–94. [Google Scholar]
- Lorek, A.; Majewski, J. Humidity Measurement in Carbon Dioxide with Capacitive Humidity Sensors at Low Temperature and Pressure. Sensors 2018, 18, 2615. [Google Scholar] [CrossRef] [Green Version]
- Python Software Foundation. Welcome to Python.Org. Available online: https://www.python.org/ (accessed on 18 February 2022).
- Oracle Corporation and/or Its Affiliates. MySQL Workbench. Available online: https://www.mysql.com/products/workbench/ (accessed on 18 February 2022).
- Contributors PhpMyAdmin. Available online: https://www.phpmyadmin.net/ (accessed on 18 February 2022).
- Logroño, D.J.B.; Lara, O.O.E.; Rivera, A.D.P. Implementación del bootstrap como una metodología ágil en la web. Rev. Arbitr. Interdiscip. Koinonía 2020, 5, 268–286. [Google Scholar] [CrossRef] [Green Version]
- Sánchez, J.C.R.; Calderón, A.E.; Medina, J.A.P.; Carmona, J.J.D. Implementación en fpga de sensor de temperatura y humedad relativa para una futura plataforma de monitoreo ambiental. Pist. Educ. 2018, 38, 1296–1313. [Google Scholar]
- Venkateswaran, P.; Mukherjee, M.; Sanyal, A.; Das, S.; Nandi, R. Design and Implementation of FPGA Based Interface Model for Scale-Free Network Using I2C Bus Protocol on Quartus II 6.0. In Proceedings of the 2009 4th International Conference on Computers and Devices for Communication (CODEC), Kolkata, India, 14–16 December 2009; pp. 1–4. [Google Scholar]
- Shen, H.; Tang, Y.; Muraoka, H.; Washitani, I. Characteristics of Leaf Photosynthesis and Simulated Individual Carbon Budget in Primula Nutans under Contrasting Light and Temperature Conditions. J. Plant Res. 2008, 121, 191. [Google Scholar] [CrossRef]
- Zaks, J.; Amarnath, K.; Sylak-Glassman, E.J.; Fleming, G.R. Models and Measurements of Energy-Dependent Quenching. Photosynth. Res. 2013, 116, 389–409. [Google Scholar] [CrossRef] [Green Version]
- Van Rensen, J.J.S.; Vredenberg, W.J. Adaptation of Photosystem II to High and Low Light in Wild-Type and Triazine-Resistant Canola Plants: Analysis by a Fluorescence Induction Algorithm. Photosynth. Res. 2011, 108, 191. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kumar, J.L.G.; Zhao, Y.Q. A Review on Numerous Modeling Approaches for Effective, Economical and Ecological Treatment Wetlands. J. Environ. Manag. 2011, 92, 400–406. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, L.; Xing, D.; Wang, J. A Non-Invasive and Real-Time Monitoring of the Regulation of Photosynthetic Metabolism Biosensor Based on Measurement of Delayed Fluorescence in Vivo. Sensors 2007, 7, 52–66. [Google Scholar] [CrossRef] [Green Version]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Reference | Variables | Modeling Method |
|---|---|---|
| Farquhar et al. [3] | Temperature, CO2 concentration, light intensity, humidity, and oxygen concentration. | Mechanistic model |
| Chen et al. [6] | CO2, light, Rubisco, and air temperature. | Mechanistic model Non-linear regression |
| Zufferey et al. [7] | Light, leaf temperature, age of the leaves, CO2 gas exchange, and air temperature. | Non-linear regression Non-rectangular hyperbola |
| Boonen et al. [8] | Maximal photosynthetic rate, quantum efficiency and respiration rate at leaf level, and microclimatic data as spatial distribution of leaf area index, leaf angle (or extinction coefficient), air temperature, and photosynthetically active radiation (PAR). | Multi-layer model 3D scaling |
| Ye [9] | Irradiance, CO2 concentration, temperature, humidity, and oxygen concentration. | Non-rectangular hyperbolic, rectangular hyperbolic, binomial regression |
| Bernacchi et al. [10] | Rubisco-catalyzed carboxylation, rate of ribulose 1,5-bisphosphate (RuBP) regeneration via electron transport, or the rate of RuBP regeneration via triose phosphate utilization. | Mechanistic model |
| LI-COR [11] | CO2, H2O, air temperature, leaf temperature, airflow, pressure, and light. | Mechanistic model based on Farquhar et al., 1980 |
| Müller et al. [12] | CO2 and H2O gas exchange, leaf nitrogen content, growth temperature, among others. | Mechanistic |
| Johnson et al. [13] | Direct and diffuse light, temperature, nitrogen availability and CO2 concentration, protein distribution, leaf area index, and respiration. | White-box model using derivatives and integrals of nonlinear and non-exponential approximations |
| Lombardozzi et al. [14] | Stomatal conductance for CO2 diffusion, light compensation point, CO2 assimilation rate of the leaf, vapor pressure deficit, leaf-surface CO2 concentration, and CO2 compensation point. | Mechanistic |
| García-Camacho et al. [5] | Irradiance, nitrate, phosphate, chlorophyll, carbon, concentration of PSU, and dissolved O2 concentration. | Mechanistic model Steady state equations |
| Caemmerer [15] | CO2 assimilation and diffusion, light intensity, temperature. CO2 and O2 partial pressures, Rubisco, intercellular and chloroplast CO2 pressure. | Steady state models Kinetic constants of Rubisco are usually assumed to be similar among different species |
| Serbin et al. [16] | Visible and shortwave infrared spectra imaging (414–2447 nm). | Partial least-squares regression in pixel level variation |
| Janka et al. [17] | Stomatal conductance and leaf energy balance. | Dynamic mechanistic model evaluated by a linear regression of predicted values |
| Methods Used for Photosynthesis Estimation | Description |
|---|---|
| Invasive methods | |
| Destructive | Involves cutting a whole plant or a portion of it to estimate the photosynthetic activity based on the accumulation of dry matter in the plant, from the stage of germination until it is cut [20]. |
| Manometric | Directly measures oxygen (O2) pressure or carbon dioxide (CO2) in an isolated chamber with photosynthetic organisms [21]. |
| Electrochemical | Uses electrochemical electrodes to measure O2, CO2, or pH in aqueous solutions of the sample to detect variations that depend on photosynthetic activity [21]. |
| Gas exchange | Isolates the sample for analysis in a closed chamber to quantify the CO2 concentration [22,23]. Concentrated CO2 gas is detected by an infrared gas sensor (called IRGA for Infra-Red Gas Analysis sensors) [11]. |
| Carbon isotopes | Uses carbon isotopes such as 11C, 12C, and 14C to produce incorporated CO2 with radioactivity. This methodology is applied to analyze samples in isolated and illuminated chambers to produce a maximum fixation of radioactive CO2 during photosynthesis [24,25]. The main disadvantage is that it is destructive as it fixes a radioactive compound onto the sample; and furthermore, precision depends on lighting conditions. |
| Acoustic waves | Based on the principle of sound wave distortion in the medium in which waves propagate. The technique involves placing an acoustic transmitter on the seabed of the intended area to monitor photosynthetic activity. The disadvantage is that it dependent on water conditions and is sensitive to environmental disturbances [26]. |
| Fluorescence | Way in which a certain amount of light energy absorbed by chlorophylls is dissipated. The fluorescence emission can be analyzed and quantified, providing information on the electron transport rate, the quantum yield, and the existence of photoinhibition of photosynthesis. Indeed, fluorescence is used in various ways, and it has different applications. For further details, see reference [27]. |
| Non-invasive methods (Optical techniques) | |
| Spectroscopy | Allows to determine the qualitative and quantitative composition of a sample, using known patterns or spectra; thus, detecting the absorption or emission in wavelengths of electromagnetic radiation, by means of spectrum analyzers [28]. |
| Thermography | Measures the electromagnetic radiation emitted by the plant through its temperature. To infer a body’s temperature based on the amount of infrared light it radiates enables us to avoid any physical contact with it. This procedure uses an infrared thermography camera for the measurement (Therma CAM FLIR E25, with range 7–13.5 μm) [28]. |
| Chlorophyll fluorescence | Based on the fact that chlorophyll, when excited by solar radiation, has the ability to re-emit photons at approximately 685 and 740 nm. After fluorescing, chlorophyll returns to its stable state. The relationship between fluorescence and the amount of active chlorophyll is directly proportional. Fluorescence measurement has been proposed through a Phase Amplitude Modulator (PAM) type fluorimeter in conjunction with a lock-in amplifier [28]. |
| Gas analysis | Consists of a gas analysis, where the subject’s O2 and CO2 gas changes are measured in closed or open chambers using infrared gas sensors; thus, measuring the decrease or change in the quantum flux density [28]. |
| Photoacoustics | The absorption of light in the leaf generates a change in molecular volume and in photoreaction enthalpy. These changes produce pressure, heat, and oxygen signals at the same frequency as the light beam and are sensed by a piezoelectric transducer for analysis [28]. |
| Optical microscopy | Allows for the examination biological structures at the molecular detection level and to carry out investigations of functional dynamics in living cells for prolonged periods of time [28]. |
| Intracellular oxygen concentrations | Allows for the measurement of intracellular concentrations of O2 in plants. It consists of injecting oxygen cells that are sensitive to phosphorescence (encapsulated in polystyrene microbeads), an excitation signal of a modulated optical multifrequency is then applied. This allows a precise determination of any changes in the life of the phosphorescent characteristics that are due to oxygen. The measurement of the internal oxygen concentration of plant tissue proves to be a direct quantifier of its photosynthetic activity [28]. |
| Irradiance | Consists of the measurement of photons available in the radiation of photosynthesis (PAR), which are measured in a wavelength that ranges from 400 to 700 nm [28]. |
| Variable to Measure | Proposed Sensor |
|---|---|
| Leaf temperature | Thermopile TMP006 |
| Relative humidity | SHT75 sensor |
| Solar radiation | Light to Frequency Converter TSL230RD |
| Parameter | M23 | M33 |
|---|---|---|
| a1 | 0.20590 | 0.20590 |
| a2 | −0.50650 | −0.08284 |
| a3 | 0.45090 | 0.17126 |
| a4 | 0.30280 | 0.30280 |
| a5 | −0.11820 | −0.11820 |
| Oa1 | 1.51698 | 0.46231 |
| Oa1 | 14.8369482 | 6.93875017 |
| Plant | Model | Rho/CI | p-Value | Cohen’s d | Average Error (%) |
|---|---|---|---|---|---|
| C. annuum L. | OM23 | 0.98 [0.99, 1.0] | <0.05 | 0.37 | 43.79 |
| C. annuum L. | AM23 | 0.98 [0.99, 1.0] | <0.05 | 0 | 3.1 |
| C. annuum L. | OM33 | 0.98 [0.94, 1.0] | <0.05 | 0.35 | 10.07 |
| C. annuum L. | AM33 | 0.98 [0.94, 1.0] | <0.05 | 0 | 8.21 |
| C. chinense Jacq. | OM23 | 0.98 [0.73, 1.0] | <0.05 | 5.92 | 165.21 |
| C. chinense Jacq. | AM23 | 0.98 [0.8, 1.0] | <0.05 | 0.61 | 21.72 |
| C. chinense Jacq. | OM33 | 0.99 [0.86, 1.0] | 0.05 | 2.73 | 73.53 |
| C. chinense Jacq. | AM33 | 0.99 [0.86, 1.0] | <0.05 | 0 | 18.45 |
| Plant | Model | Cohen’s d |
|---|---|---|
| C. chinense Jacq. | OM23 | 5.92 |
| C. chinense Jacq. | AM23 | 0.61 |
| C. chinense Jacq. | OM33 | 2.73 |
| C. chinense Jacq. | AM33 | 0 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
García-Rodríguez, L.d.C.; Prado-Olivarez, J.; Guzmán-Cruz, R.; Heil, M.; Guevara-González, R.G.; Diaz-Carmona, J.; López-Tapia, H.; Padierna-Arvizu, D.d.J.; Espinosa-Calderón, A. Black-Box Mathematical Model for Net Photosynthesis Estimation and Its Digital IoT Implementation Based on Non-Invasive Techniques: Capsicum annuum L. Study Case. Sensors 2022, 22, 5275. https://doi.org/10.3390/s22145275
García-Rodríguez LdC, Prado-Olivarez J, Guzmán-Cruz R, Heil M, Guevara-González RG, Diaz-Carmona J, López-Tapia H, Padierna-Arvizu DdJ, Espinosa-Calderón A. Black-Box Mathematical Model for Net Photosynthesis Estimation and Its Digital IoT Implementation Based on Non-Invasive Techniques: Capsicum annuum L. Study Case. Sensors. 2022; 22(14):5275. https://doi.org/10.3390/s22145275
Chicago/Turabian StyleGarcía-Rodríguez, Luz del Carmen, Juan Prado-Olivarez, Rosario Guzmán-Cruz, Martin Heil, Ramón Gerardo Guevara-González, Javier Diaz-Carmona, Héctor López-Tapia, Diego de Jesús Padierna-Arvizu, and Alejandro Espinosa-Calderón. 2022. "Black-Box Mathematical Model for Net Photosynthesis Estimation and Its Digital IoT Implementation Based on Non-Invasive Techniques: Capsicum annuum L. Study Case" Sensors 22, no. 14: 5275. https://doi.org/10.3390/s22145275
APA StyleGarcía-Rodríguez, L. d. C., Prado-Olivarez, J., Guzmán-Cruz, R., Heil, M., Guevara-González, R. G., Diaz-Carmona, J., López-Tapia, H., Padierna-Arvizu, D. d. J., & Espinosa-Calderón, A. (2022). Black-Box Mathematical Model for Net Photosynthesis Estimation and Its Digital IoT Implementation Based on Non-Invasive Techniques: Capsicum annuum L. Study Case. Sensors, 22(14), 5275. https://doi.org/10.3390/s22145275