Save Muscle Information–Unfiltered EEG Signal Helps Distinguish Sleep Stages

 ,
, 
 ,
,
Abstract
1. Introduction
2. Method and Material
2.1. Automatic Sleep Stage Annotation Algorithm
2.2. Statistics
2.3. Database
3. Results
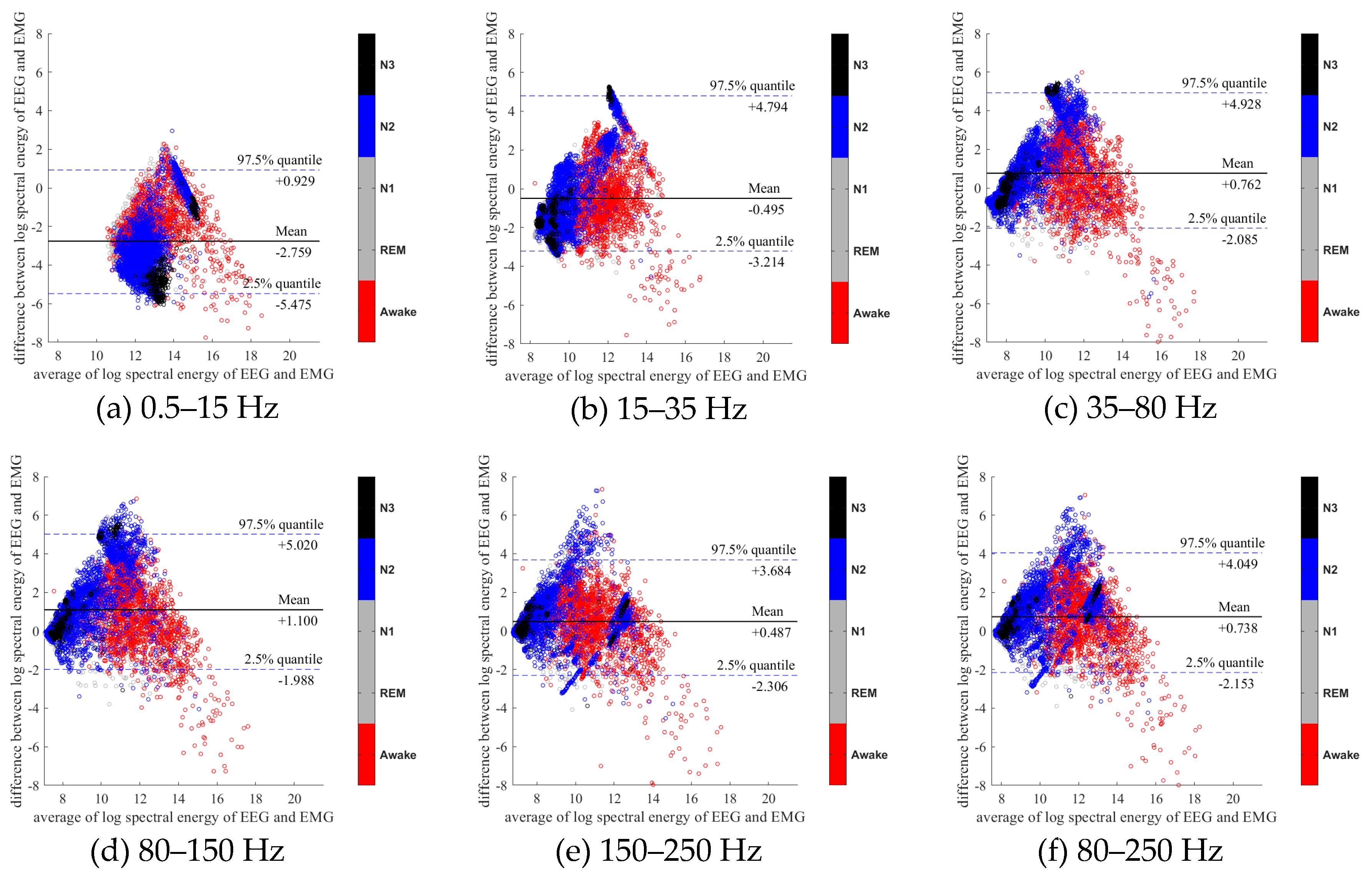
3.1. Relationship with EMG
3.2. Automatic Sleep Stage Annotation Results
- For the REM stage, Table 4 shows that 8% and 23% REM epochs were misclassified as N1 and N2 stages when the features extracted from low-pass-filtered EEG signals are used for classification. Table 5 shows that after taking high-frequency features extracted from the unfiltered raw data into account, the misclassification rate of REM epochs decreases (7% REM epochs were misclassified as N1 and 10% REM epochs were misclassified as N2).
- When the features extracted from the low-pass filtered EEG signals are used for classification, the F1 scores for REM and N1 stages are 66% and 29%, respectively. After taking high-frequency features extracted from the unfiltered raw data into account, the F1 scores for REM and N1 stages increase to 78% and 34%, respectively.
4. Discussion and Conclusion
Author Contributions
Funding
Conflicts of Interest
References
- Frølich, L.; Dowding, I. Removal of muscular artifacts in EEG signals: A comparison of linear decomposition methods. Brain Inform. 2018, 5, 13–22. [Google Scholar] [CrossRef] [PubMed]
- Larsen, L.H.; Moe, K.E.; Vitiello, M.V.; Prinz, P.N. Age trends in the sleep EEG of healthy older men and women. J. Sleep Res. 1995, 4, 160–172. [Google Scholar] [CrossRef] [PubMed]
- Barlow, J.S. Muscle spike artifact minimization in EEGs by time-domain filtering. Electroencephalogr. Clin. Neurophysiol. 1983, 55, 487–491. [Google Scholar] [CrossRef]
- Berry, R.B.; Brooks, R.; Gamaldo, C.E.; Harding, S.M.; Marcus, C.; Vaughn, B.V. The AASM manual for the scoring of sleep and associated events. In Rules, Terminology and Technical Specifications; American Academy of Sleep Medicine: Darien, IL, USA, 2012. [Google Scholar]
- Berry, R.B.; Budhiraja, R.; Gottlieb, D.J.; Gozal, D.; Iber, C.; Kapur, V.K.; Marcus, C.L.; Mehra, R.; Parthasarathy, S.; Quan, S.F.; et al. Rules for scoring respiratory events in sleep: Update of the 2007 AASM manual for the scoring of sleep and associated events. J. Clin. Sleep Med. 2012, 8, 597–619. [Google Scholar] [CrossRef] [PubMed]
- Mikkelsen, K.B.; Tabar, Y.R.; Kappel, S.L.; Christensen, C.B.; Toft, H.O.; Hemmsen, M.C.; Rank, M.L.; Otto, M.; Kidmose, P. Accurate whole-night sleep monitoring with dry-contact ear-EEG. Sci. Rep. 2019, 9, 16824. [Google Scholar] [CrossRef] [PubMed]
- Ferster, M.L.; Lustenberger, C.; Karlen, W. Configurable mobile system for autonomous high-quality sleep monitoring and closed-loop acoustic stimulation. IEEE Sens. Lett. 2019, 3, 1–4. [Google Scholar] [CrossRef]
- Nakamura, T.; Alqurashi, Y.D.; Morrell, M.J.; Mandic, D.P. Hearables: Automatic Overnight Sleep Monitoring With Standardized In-Ear EEG Sensor. IEEE Trans. Biomed. Eng. 2019, 67, 203–212. [Google Scholar] [CrossRef]
- Debener, S.; Emkes, R.; De Vos, M.; Bleichner, M. Unobtrusive ambulatory EEG using a smartphone and flexible printed electrodes around the ear. Sci. Rep. 2015, 5, 16743. [Google Scholar] [CrossRef]
- Yazicioglu, R.F.; Van Hoof, C.; Puers, R. Biopotential Readout Circuits for Portable Acquisition Systems; Springer Science & Business Media, Springer: Dordrecht, The Netherlands, 2008. [Google Scholar]
- Tuncel, D.; Dizibuyuk, A.; Kiymik, M.K. Time frequency based coherence analysis between EEG and EMG activities in fatigue duration. J. Med. Syst. 2010, 34, 131–138. [Google Scholar] [CrossRef]
- Pope, K.J.; Fitzgibbon, S.P.; Lewis, T.W.; Whitham, E.M.; Willoughby, J.O. Relation of gamma oscillations in scalp recordings to muscular activity. Brain Topogr. 2009, 22, 13–17. [Google Scholar] [CrossRef]
- Cisotto, G.; Michieli, U.; Badia, L. A coherence study on EEG and EMG signals. arXiv 2017, arXiv:171201277. preprint. [Google Scholar]
- Levendowski, D.J.; Ferini-Strambi, L.; Gamaldo, C.; Cetel, M.; Rosenberg, R.; Westbrook, P.R. The accuracy, night-to-night variability, and stability of frontopolar sleep electroencephalography biomarkers. J. Clin. Sleep Med. 2017, 13, 791–803. [Google Scholar] [CrossRef] [PubMed]
- Levendowski, D.J.; Louis, E.K.S.; Strambi, L.F.; Galbiati, A.; Westbrook, P.; Berka, C. comparison of eMg power during sleep from the submental and frontalis muscles. Nat. Sci. Sleep 2018, 10, 431. [Google Scholar] [CrossRef] [PubMed]
- Frauscher, B.; Iranzo, A.; Gaig, C.; Gschliesser, V.; Guaita, M.; Raffelseder, V.; Ehrmann, L.; Sola, N.; Salamero, M.; Tolosa, E.; et al. Normative EMG values during REM sleep for the diagnosis of REM sleep behavior disorder. Sleep 2012, 35, 835–847. [Google Scholar] [CrossRef] [PubMed]
- Kim, H.; Choi, S. Automatic Sleep Stage Classification Using EEG and EMG Signal. In Proceedings of the 2018 Tenth International Conference on Ubiquitous and Future Networks (ICUFN), Prague, Czech Republic, 3–6 July 2018; pp. 207–212. [Google Scholar]
- Koley, B.; Dey, D. An ensemble system for automatic sleep stage classification using single channel EEG signal. Comput. Biol. Med. 2012, 42, 1186–1195. [Google Scholar] [CrossRef]
- Tsinalis, O.; Matthews, P.M.; Guo, Y. Automatic sleep stage scoring using time-frequency analysis and stacked sparse autoencoders. Ann. Biomed. Eng. 2016, 44, 1587–1597. [Google Scholar] [CrossRef]
- Supratak, A.; Dong, H.; Wu, C.; Guo, Y. DeepSleepNet: A model for automatic sleep stage scoring based on raw single-channel EEG. IEEE Trans. Neural Syst. Rehabil. Eng. 2017, 25, 1998–2008. [Google Scholar] [CrossRef]
- Vilamala, A.; Madsen, K.H.; Hansen, L.K. Deep convolutional neural networks for interpretable analysis of EEG sleep stage scoring. In Proceedings of the 2017 IEEE 27th International Workshop on Machine Learning for Signal Processing (MLSP), Tokyo, Japan, 25–28 September 2017; pp. 1–6. [Google Scholar]
- Liu, G.-R.; Lo, Y.-L.; Malik, J.; Sheu, Y.-C.; Wu, H.-T. Diffuse to fuse EEG spectra–Intrinsic geometry of sleep dynamics for classification. Biomed. Signal Process. Control 2020, 55, 101576. [Google Scholar] [CrossRef]
- Goldberger, A.L.; Amaral, L.A.; Glass, L.; Hausdorff, J.M.; Ivanov, P.C.; Mark, R.G.; Mietus, J.E.; Moody, G.B.; Peng, C.K.; Stanley, H.E. PhysioBank, PhysioToolkit, and PhysioNet: Components of a new research resource for complex physiologic signals. Circulation 2000, 101, e215–e220. [Google Scholar] [CrossRef]
- Mallat, S. Group invariant scattering. Commun. Pure Appl. Math. 2012, 65, 1331–1398. [Google Scholar] [CrossRef]
- Andén, J.; Mallat, S. Deep scattering spectrum. IEEE Trans. Signal Process. 2014, 62, 4114–4128. [Google Scholar] [CrossRef]
- Bruna, J.; Mallat, S.; Bacry, E.; Muzy, J.-F. Intermittent process analysis with scattering moments. Ann. Stat. 2015, 43, 323–351. [Google Scholar] [CrossRef]
- Hardoon, D.R.; Szedmak, S.; Shawe-Taylor, J. Canonical correlation analysis: An overview with application to learning methods. Neural Comput. 2004, 16, 2639–2664. [Google Scholar] [CrossRef] [PubMed]
- Talmon, R.; Wu, H.-T. Latent common manifold learning with alternating diffusion: Analysis and applications. Appl. Comput. Harmon. Anal. 2019, 47, 848–892. [Google Scholar] [CrossRef]
- Scholkopf, B.; Smola, A.J. Learning with Kernels: Support Vector Machines, Regularization, Optimization, and Beyond; MIT Press: Cambridge, MA, USA, 2001. [Google Scholar]
- Efron, B.; Tibshirani, R.J. An Introduction to the Bootstrap; CRC Press: Boca Raton, FL, USA, 1994. [Google Scholar]
- Good, P. Permutation Tests: A Practical Guide to Resampling Methods for Testing Hypotheses; Springer-Verlag Series in Statistics: New York, NY, USA, 1994. [Google Scholar]
- Giavarina, D. Understanding bland altman analysis. Biochem. Med. Biochem. Med. 2015, 25, 141–151. [Google Scholar] [CrossRef] [PubMed]
- Sigl, J.C.; Chamoun, N.G. An introduction to bispectral analysis for the electroencephalogram. J. Clin. Monit. 1994, 10, 392–404. [Google Scholar] [CrossRef]
- Lo, Y.L.; Lin, T.Y.; Fang, Y.F.; Wang, T.Y.; Chen, H.C.; Chou, C.L.; Chung, F.T.; Kuo, C.H.; Feng, P.H.; Liu, C.Y.; et al. Feasibility of bispectral index-guided propofol infusion for flexible bronchoscopy sedation: A randomized controlled trial. PLoS ONE 2011, 6, e27769. [Google Scholar] [CrossRef]
- Kim, D.; Ahn, J.H.; Jung, H.; Choi, K.Y.; Jeong, J.S. Effects of neuromuscular blockade reversal on bispectral index and frontal electromyogram during steady-state desflurane anesthesia: A randomized trial. Sci. Rep. 2019, 9, 1–7. [Google Scholar] [CrossRef]
- Lawoyin, S. Novel Technologies for the Detection and Mitigation of Drowsy Driving. PhD Thesis, Biomedical Engineering Department, Virginia Commonwealth University, Richmond, VA, USA, 2014. [Google Scholar]
- Narayan, R.; Kamat, A.; Khanolkar, M.; Kamat, S.; Desai, S.R.; Dhume, R. Quantitative evaluation of muscle relaxation induced by Kundalini yoga with the help of EMG integrator. Indian J. Physiol. Pharm. 1990, 34, 279–281. [Google Scholar]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Frequency Bands | Sleep Stages | |||||
|---|---|---|---|---|---|---|
| Awake | REM | N1 | N2 | N3 | All | |
| 0.5–15 | 0.48 ** | −0.12 | 0.16 ** | 0.07 * | 0.23 | 0.21 ** |
| 15–35 | 0.25 ** | 0.20 ** | 0.45 ** | 0.24 ** | 0.00 | 0.42 ** |
| 35–80 | 0.34 ** | 0.28 ** | 0.60 ** | 0.44 ** | 0.08 | 0.56 ** |
| 80–150 | 0.42 ** | 0.35 ** | 0.65 ** | 0.46 ** | 0.22 * | 0.60 ** |
| 150–250 | 0.57 ** | 0.81 ** | 0.77 ** | 0.65 ** | 0.93 ** | 0.73 ** |
| 80–250 | 0.48 ** | 0.70 ** | 0.72 ** | 0.58 ** | 0.90 ** | 0.68 ** |
| Frequency Bands | Sleep Stages | |||||
|---|---|---|---|---|---|---|
| Awake | REM | N1 | N2 | N3 | All | |
| 0.5–15 | 0.43 ** | −0.13 | 0.27 ** | −0.14 | −0.36 | 0.18 ** |
| 15–35 | 0.20 ** | −0.07 | 0.40 ** | −0.08 | 0.07 | 0.32 ** |
| 35–80 | 0.32 ** | 0.19 * | 0.61 ** | 0.32 ** | 0.36 ** | 0.55 ** |
| 80–150 | 0.41 ** | 0.39 ** | 0.68 ** | 0.48 ** | 0.20 * | 0.63 ** |
| 150–250 | 0.61 ** | 0.81 ** | 0.82 ** | 0.73 ** | 0.94 ** | 0.79 ** |
| 80–250 | 0.49 ** | 0.81 ** | 0.78 ** | 0.66 ** | 0.91 ** | 0.75 ** |
| Frequency Bands | Sleep Stages | |||||
|---|---|---|---|---|---|---|
| Awake | REM | N1 | N2 | N3 | All | |
| 0.5–15 | 0.56 ** | 0.03 | 0.36 ** | 0.11 ** | −0.19 | 0.28 ** |
| 15–35 | 0.26 ** | 0.28 ** | 0.39 ** | 0.19 ** | −0.08 | 0.40 ** |
| 35–80 | 0.40 ** | 0.33 ** | 0.59 ** | 0.50 ** | 0.06 | 0.59 ** |
| 80–150 | 0.47 ** | 0.43 ** | 0.67 ** | 0.55 ** | 0.25 * | 0.65 ** |
| 150–250 | 0.61 ** | 0.86 ** | 0.79 ** | 0.73 ** | 0.95 ** | 0.78 ** |
| 80–250 | 0.53 ** | 0.80 ** | 0.76 ** | 0.71 ** | 0.93 ** | 0.75 ** |
| Predicted | Per-class Metrics | |||||||
|---|---|---|---|---|---|---|---|---|
| Awake | REM | N1 | N2 | N3 | PR | RE | F1 | |
| Awake (27%) | 1894 (91%) | 26 (1%) | 91 (4%) | 67 (3%) | 1 (0%) | 83 | 91 | 87 |
| REM (9%) | 27 (4%) | 470 (66%) | 54 (8%) | 162 (23%) | 0 (0%) | 66 | 66 | 66 |
| N1 (11%) | 258 (32%) | 137 (17%) | 181 (22%) | 238 (29%) | 2 (0%) | 41 | 22 | 29 |
| N2 (48%) | 97 (3%) | 83 (2%) | 116 (3%) | 3143 (88%) | 152 (4%) | 85 | 88 | 86 |
| N3 (5%) | 0 (0%) | 0 (0%) | 1 (0%) | 97 (27%) | 259 (73%) | 63 | 73 | 67 |
| Predicted | Per-class Metrics | |||||||
|---|---|---|---|---|---|---|---|---|
| Awake | REM | N1 | N2 | N3 | PR | RE | F1 | |
| Awake (27%) | 1817 (87%) | 36 (2%) | 164 (8%) | 62 (3%) | 0 (0%) | 83 | 87 | 85 |
| REM (9%) | 15 (2%) | 576 (81%) | 48 (7%) | 74 (10%) | 0 (0%) | 76 | 81 | 78 |
| N1 (11%) | 262 (32%) | 90 (11%) | 249 (31%) | 213 (26%) | 2 (0%) | 39 | 31 | 34 |
| N2 (48%) | 84 (2%) | 54 (2%) | 184 (5%) | 3106 (86%) | 163 (5%) | 88 | 86 | 87 |
| N3 (5%) | 0 (0%) | 0 (0%) | 1 (0%) | 95 (27%) | 261 (73%) | 61 | 73 | 67 |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Liu, G.-R.; Lustenberger, C.; Lo, Y.-L.; Liu, W.-T.; Sheu, Y.-C.; Wu, H.-T. Save Muscle Information–Unfiltered EEG Signal Helps Distinguish Sleep Stages. Sensors 2020, 20, 2024. https://doi.org/10.3390/s20072024
Liu G-R, Lustenberger C, Lo Y-L, Liu W-T, Sheu Y-C, Wu H-T. Save Muscle Information–Unfiltered EEG Signal Helps Distinguish Sleep Stages. Sensors. 2020; 20(7):2024. https://doi.org/10.3390/s20072024
Chicago/Turabian StyleLiu, Gi-Ren, Caroline Lustenberger, Yu-Lun Lo, Wen-Te Liu, Yuan-Chung Sheu, and Hau-Tieng Wu. 2020. "Save Muscle Information–Unfiltered EEG Signal Helps Distinguish Sleep Stages" Sensors 20, no. 7: 2024. https://doi.org/10.3390/s20072024
APA StyleLiu, G.-R., Lustenberger, C., Lo, Y.-L., Liu, W.-T., Sheu, Y.-C., & Wu, H.-T. (2020). Save Muscle Information–Unfiltered EEG Signal Helps Distinguish Sleep Stages. Sensors, 20(7), 2024. https://doi.org/10.3390/s20072024