Induced Gamma-Band Activity during Actual and Imaginary Movements: EEG Analysis


Abstract
1. Introduction
2. Material and Methods
2.1. Participants and Experiment Description
- Basal recording. Participants keep their eyes fixed on the center of the screen (to prevent eye movement; they also try not to blink) and refrain from performing any motor or specific mental activity. A total of 18 min of basal activity are recorded, divided into 3 parts (each 6 min long) with a rest of approximately 1 min between each. This step is the first performed by the participants.
- Imaginary motor task. An on-screen cue triggers the imaginary motor task, thereby obtaining in the EEG trace the motor GBA induced by that imaginary movement. The imaginary task consists of simulating, without muscle activation, rapid extension of the wrist followed by brief relaxation. This phase lasts approximately 40 min.
- Actual motor task. The subjects perform an actual motor task with the same characteristics and duration of imaginary motor task.
2.2. Data Acquisition
2.3. Data Analysis
2.4. Calculation of the GBA
- -
- GBA during the basal experiment: GBAb.
- -
- GBA during actual motor tasks: GBAac.
- -
- GBA during imaginary motor tasks: GBAim.
2.5. Calculation of ERS for the GBA
2.6. Statistical Analysis
3. Results
4. Discussion
5. Conclusions
Author Contributions
Funding
Conflicts of Interest
References
- Uhlhaas, P.J.; Pipa, G.; Neuenschwander, S.; Wibral, M.; Singer, W. A new look at gamma? High-(>60 Hz) γ-band activity in cortical networks: Function, mechanisms and impairment. Prog. Biophys. Mol. Biol. 2011, 105, 14–28. [Google Scholar] [CrossRef]
- Combrisson, E.; Perrone-Bertolotti, M.; Soto, J.L.; Alamian, G.; Kahane, P.; Lachaux, J.-P.; Guillot, A.; Jerbi, K. From intentions to actions: Neural oscillations encode motor processes through phase, amplitude and phase-amplitude coupling. Neuroimage 2017, 147, 473–487. [Google Scholar] [CrossRef] [PubMed]
- Uhlhaas, P.J.; Haenschel, C.; Nikolić, D.; Singer, W. The role of oscillations and synchrony in cortical networks and their putative relevance for the pathophysiology of schizophrenia. Schizophr. Bull. 2008, 34, 927–943. [Google Scholar] [CrossRef] [PubMed]
- Cheyne, D.; Ferrari, P. MEG studies of motor cortex gamma oscillations: Evidence for a gamma “fingerprint” in the brain? Front. Hum. Neurosci. 2013, 7. [Google Scholar] [CrossRef]
- Tallon-Baudry, C.; Bertrand, O. Oscillatory gamma activity in humans and its role in object representation. Trends Cogn. Sci. 1999, 3, 151–162. [Google Scholar] [CrossRef]
- Szurhaj, W.; Bourriez, J.-L.; Kahane, P.; Chauvel, P.; Mauguière, F.; Derambure, P. Intracerebral study of gamma rhythm reactivity in the sensorimotor cortex. Eur. J. Neurosci. 2005, 21, 1223–1235. [Google Scholar] [CrossRef]
- Crone, N.E.; Miglioretti, D.L.; Gordon, B.; Lesser, R.P. Functional mapping of human sensorimotor cortex with electrocorticographic spectral analysis. II. Event-related synchronization in the gamma band. Brain 1998, 121, 2301–2315. [Google Scholar] [CrossRef]
- Aoki, F.; Fetz, E.E.; Shupe, L.; Lettich, E.; Ojemann, G.A. Increased gamma-range activity in human sensorimotor cortex during performance of visuomotor tasks. Clin. Neurophysiol. 1999, 110, 524–537. [Google Scholar] [CrossRef]
- Ohara, S.; Ikeda, A.; Kunieda, T.; Yazawa, S.; Baba, K.; Nagamine, T.; Taki, W.; Hashimoto, N.; Mihara, T.; Shibasaki, H. Movement-related change of electrocorticographic activity in human supplementary motor area proper. Brain 2000, 123, 1203–1215. [Google Scholar] [CrossRef]
- Pfurtscheller, G.; Graimann, B.; Huggins, J.E.; Levine, S.P.; Schuh, L.A. Spatiotemporal patterns of beta desynchronization and gamma synchronization in corticographic data during self-paced movement. Clin. Neurophysiol. 2003, 114, 1226–1236. [Google Scholar] [CrossRef]
- Miller, K.J.; Schalk, G.; Fetz, E.E.; den Nijs, M.; Ojemann, J.G.; Rao, R.P.N. Cortical activity during motor execution, motor imagery, and imagery-based online feedback. Proc. Natl. Acad. Sci. USA 2010, 107, 4430–4435. [Google Scholar] [CrossRef]
- Pfurtscheller, G.; Neuper, C. Simultaneous EEG 10 Hz desynchronization and 40 Hz synchronization during finger movements. Neuroreport 1992, 3, 1057–1060. [Google Scholar] [CrossRef] [PubMed]
- Shibata, T.; Shimoyama, I.; Ito, T.; Abla, D.; Iwasa, H.; Koseki, K.; Yamanouchi, N.; Sato, T.; Nakajima, Y. Event-related dynamics of the gamma-band oscillation in the human brain: information processing during a GO/NOGO hand movement task. Neurosci. Res. 1999, 33, 215–222. [Google Scholar] [CrossRef]
- Ball, T.; Demandt, E.; Mutschler, I.; Neitzel, E.; Mehring, C.; Vogt, K.; Aertsen, A.; Schulze-Bonhage, A. Movement related activity in the high gamma range of the human EEG. Neuroimage 2008, 41, 302–310. [Google Scholar] [CrossRef] [PubMed]
- Darvas, F.; Scherer, R.; Ojemann, J.G.; Rao, R.P.; Miller, K.J.; Sorensen, L.B. High gamma mapping using EEG. Neuroimage 2010, 49, 930–938. [Google Scholar] [CrossRef] [PubMed]
- Demandt, E.; Mehring, C.; Vogt, K.; Schulze-Bonhage, A.; Aertsen, A.; Ball, T. Reaching movement onset- and end-related characteristics of EEG spectral power modulations. Front. Neurosci. 2012, 6. [Google Scholar] [CrossRef] [PubMed]
- Amo, C.; Del Castillo, M.O.; Barea, R.; de Santiago, L.; Martínez-Arribas, A.; Amo-López, P.; Boquete, L. Induced Gamma-Band Activity During Voluntary Movement: EEG Analysis for Clinical Purposes. Mot. Control 2016, 20, 409–428. [Google Scholar] [CrossRef]
- Decety, J. The neurophysiological basis of motor imagery. Behav. Brain Res. 1996, 77, 45–52. [Google Scholar] [CrossRef]
- Hardwick, R.M.; Caspers, S.; Eickhoff, S.B.; Swinnen, S.P. Neural correlates of action: Comparing meta-analyses of imagery, observation, and execution. Neurosci. Biobehav. Rev. 2018, 94, 31–44. [Google Scholar] [CrossRef]
- Leuthardt, E.C.; Schalk, G.; Wolpaw, J.R.; Ojemann, J.G.; Moran, D.W. A brain-computer interface using electrocorticographic signals in humans. J. Neural Eng. 2004, 1, 63–71. [Google Scholar] [CrossRef]
- Khan, Y.U.; Sepulveda, F. Brain–computer interface for single-trial EEG classification for wrist movement imagery using spatial filtering in the gamma band. IET Signal Process. 2010, 4, 510–517. [Google Scholar] [CrossRef]
- Kiroi, V.N.; Vladimirskii, B.M.; Aslanyan, E.V.; Bakhtin, O.M.; Minyaeva, N.R. Electrographic Correlates of Actual and Imagined Movements: Spectral Analysis. Neurosci. Behav. Physiol. 2012, 42, 21–27. [Google Scholar] [CrossRef]
- Smith, M.M.; Weaver, K.E.; Grabowski, T.J.; Rao, R.P.N.; Darvas, F. Non-invasive detection of high gamma band activity during motor imagery. Front. Hum. Neurosci. 2014, 8. [Google Scholar] [CrossRef] [PubMed]
- Korik, A.; Sosnik, R.; Siddique, N.; Coyle, D. Decoding Imagined 3D Hand Movement Trajectories From EEG: Evidence to Support the Use of Mu, Beta, and Low Gamma Oscillations. Front. Neurosci. 2018, 12. [Google Scholar] [CrossRef]
- Lazurenko, D.M.; Kiroy, V.N.; Aslanyan, E.V.; Shepelev, I.E.; Bakhtin, O.M.; Minyaeva, N.R. Electrographic Properties of Movement-Related Potentials. Neurosci. Behav. Physiol. 2018, 48, 1078–1087. [Google Scholar] [CrossRef]
- Veslin, E.Y.; Dutra, M.S.; Bevilacqua, L.; Raptopoulos, L.S.C.; Andrade, W.S.; Pereira, A.S.; Fiorani, M.; Soares, J.G.M. Lower gamma band in the classification of left and right elbow movement in real and imaginary tasks. J. Braz. Soc. Mech. Sci. Eng. 2019, 41, 91. [Google Scholar] [CrossRef]
- Nottage, J.F.; Horder, J. State-of-the-Art Analysis of High-Frequency (Gamma Range) Electroencephalography in Humans. Neuropsychobiology 2015, 72, 219–228. [Google Scholar] [CrossRef]
- Allegrini, P.; Menicucci, D.; Bedini, R.; Fronzoni, L.; Gemignani, A.; Grigolini, P.; West, B.J.; Paradisi, P. Spontaneous brain activity as a source of ideal 1/f noise. Phys. Rev. E. Stat. Nonlin. Soft Matter Phys. 2009, 80. [Google Scholar] [CrossRef]
- Hyvärinen, A. Independent component analysis: recent advances. Philos. Trans. A. Math. Phys. Eng. Sci. 2013, 371. [Google Scholar] [CrossRef]
- Ogino, M.; Kanoga, S.; Muto, M.; Mitsukura, Y. Analysis of Prefrontal Single-Channel EEG Data for Portable Auditory ERP-Based Brain–Computer Interfaces. Front. Hum. Neurosci. 2019. [Google Scholar] [CrossRef]
- Ajami, S.; Mahnam, A.; Abootalebi, V. Development of a practical high frequency brain–computer interface based on steady-state visual evoked potentials using a single channel of EEG. Biocybern. Biomed. Eng. 2018, 38, 106–114. [Google Scholar] [CrossRef]
- Lucey, B.P.; Mcleland, J.S.; Toedebusch, C.D.; Boyd, J.; Morris, J.C.; Landsness, E.C.; Yamada, K.; Holtzman, D.M. Comparison of a single-channel EEG sleep study to polysomnography. J. Sleep Res. 2016, 25, 625–635. [Google Scholar] [CrossRef] [PubMed]
- Michielli, N.; Acharya, U.R.; Molinari, F. Cascaded LSTM recurrent neural network for automated sleep stage classification using single-channel EEG signals. Comput. Biol. Med. 2019, 106, 71–81. [Google Scholar] [CrossRef]
- Han, C.-H.; Kim, Y.-W.; Kim, D.Y.; Kim, S.H.; Nenadic, Z.; Im, C.-H. Electroencephalography-based endogenous brain–computer interface for online communication with a completely locked-in patient. J. Neuroeng. Rehabil. 2019, 16, 18. [Google Scholar] [CrossRef]
- Diez, P.F.; Torres Müller, S.M.; Mut, V.A.; Laciar, E.; Avila, E.; Bastos-Filho, T.F.; Sarcinelli-Filho, M. Commanding a robotic wheelchair with a high-frequency steady-state visual evoked potential based brain–computer interface. Med. Eng. Phys. 2013, 35, 1155–1164. [Google Scholar] [CrossRef]
- Edelman, B.J.; Meng, J.; Suma, D.; Zurn, C.; Nagarajan, E.; Baxter, B.S.; Cline, C.C.; He, B. Noninvasive neuroimaging enhances continuous neural tracking for robotic device control. Sci. Robot. 2019, 4. [Google Scholar] [CrossRef]
- Xu, Y.; Ding, C.; Shu, X.; Gui, K.; Bezsudnova, Y.; Sheng, X.; Zhang, D. Shared control of a robotic arm using non-invasive brain–computer interface and computer vision guidance. Rob. Auton. Syst. 2019, 115, 121–129. [Google Scholar] [CrossRef]
- Padfield, N.; Zabalza, J.; Zhao, H.; Masero, V.; Ren, J. EEG-Based Brain-Computer Interfaces Using Motor-Imagery: Techniques and Challenges. Sensors 2019, 19, 1423. [Google Scholar] [CrossRef]
- Perdikis, S.; Tonin, L.; Saeedi, S.; Schneider, C.; del Millán, J.R. The Cybathlon BCI race: Successful longitudinal mutual learning with two tetraplegic users. PLoS Biol. 2018, 16, e2003787. [Google Scholar] [CrossRef]
- Ahn, M.; Ahn, S.; Hong, J.H.; Cho, H.; Kim, K.; Kim, B.S.; Chang, J.W.; Jun, S.C. Gamma band activity associated with BCI performance: simultaneous MEG/EEG study. Front. Hum. Neurosci. 2013, 7. [Google Scholar] [CrossRef]
- Szczuko, P. Real and imaginary motion classification based on rough set analysis of EEG signals for multimedia applications. Multimed. Tools Appl. 2017, 76, 25697–25711. [Google Scholar] [CrossRef]
- Van Benthem, K.D.; Cebulski, S.; Herdman, C.M.; Keillor, J. An EEG Brain–Computer Interface Approach for Classifying Vigilance States in Humans: A Gamma Band Focus Supports Low Misclassification Rates. Int. J. Hum.-Comput. Interact. 2018, 34, 226–237. [Google Scholar] [CrossRef]
- Herrmann, C.S.; Munk, M.H.J.; Engel, A.K. Cognitive functions of gamma-band activity: memory match and utilization. Trends Cogn. Sci. 2004, 8, 347–355. [Google Scholar] [CrossRef] [PubMed]
- Nowak, M.; Zich, C.; Stagg, C.J. Motor Cortical Gamma Oscillations: What Have We Learnt and Where Are We Headed? Curr. Behav. Neurosci. Rep. 2018, 5, 136–142. [Google Scholar] [CrossRef] [PubMed]
- van Deursen, J.A.; Vuurman, E.F.P.M.; Verhey, F.R.J.; van Kranen-Mastenbroek, V.H.J.M.; Riedel, W.J. Increased EEG gamma band activity in Alzheimer’s disease and mild cognitive impairment. J. Neural Transm. 2008, 115, 1301–1311. [Google Scholar] [CrossRef] [PubMed]
- Başar, E.; Femir, B.; Emek-Savaş, D.D.; Güntekin, B.; Yener, G.G. Increased long distance event-related gamma band connectivity in Alzheimer’s disease. NeuroImage Clin. 2017, 14, 580–590. [Google Scholar] [CrossRef]
- Yamamoto, T.; Sugaya, N.; Siegle, G.J.; Kumano, H.; Shimada, H.; Machado, S.; Murillo-Rodriguez, E.; Rocha, N.B.; Nardi, A.E.; Takamura, M.; et al. Altered Gamma-Band Activity as a Potential Biomarker for the Recurrence of Major Depressive Disorder. Front. Psychiatry 2018, 9, 691. [Google Scholar] [CrossRef]
- Baradits, M.; Kakuszi, B.; Bálint, S.; Fullajtár, M.; Mód, L.; Bitter, I.; Czobor, P. Alterations in resting-state gamma activity in patients with schizophrenia: a high-density EEG study. Eur. Arch. Psychiatry Clin. Neurosci. 2019, 269, 429–437. [Google Scholar] [CrossRef]
- Oldfield, R.C. The assessment and analysis of handedness: the Edinburgh inventory. Neuropsychologia 1971, 9, 97–113. [Google Scholar] [CrossRef]
- Herrmann, C.S.; Demiralp, T. Human EEG gamma oscillations in neuropsychiatric disorders. Clin. Neurophysiol. 2005, 116, 2719–2733. [Google Scholar] [CrossRef]
- Oostenveld, R.; Fries, P.; Maris, E.; Schoffelen, J.-M. FieldTrip: Open source software for advanced analysis of MEG, EEG, and invasive electrophysiological data. Comput. Intell. Neurosci. 2011, 2011, 156869. [Google Scholar] [CrossRef] [PubMed]
- Fitzgibbon, S.P.; Lewis, T.W.; Powers, D.M.W.; Whitham, E.W.; Willoughby, J.O.; Pope, K.J. Surface Laplacian of central scalp electrical signals is insensitive to muscle contamination. IEEE Trans. Biomed. Eng. 2013, 60, 4–9. [Google Scholar] [CrossRef] [PubMed]
- Willems, R.M.; Toni, I.; Hagoort, P.; Casasanto, D. Body-specific motor imagery of hand actions: neural evidence from right-and left-handers. Front. Hum. Neurosci. 2009, 3. [Google Scholar] [CrossRef] [PubMed]
- Zapała, D.; Zabielska-Mendyk, E.; Augustynowicz, P.; Cudo, A.; Jaśkiewicz, M.; Szewczyk, M.; Kopiś, N.; Francuz, P. The effects of handedness on sensorimotor rhythm desynchronization and motor-imagery BCI control. Sci. Rep. 2020, 10. [Google Scholar] [CrossRef]
- Pfurtscheller, G.; Lopes da Silva, F.H. Event-related EEG/MEG synchronization and desynchronization: basic principles. Clin. Neurophysiol. 1999, 110, 1842–1857. [Google Scholar] [CrossRef]
- Jeannerod, M.; Decety, J. Mental motor imagery: a window into the representational stages of action. Curr. Opin. Neurobiol. 1995, 5, 727–732. [Google Scholar] [CrossRef]
- Mannan, M.M.N.; Kamran, M.A.; Jeong, M.Y. Identification and Removal of Physiological Artifacts From Electroencephalogram Signals: A Review. IEEE Access 2018, 6, 30630–30652. [Google Scholar] [CrossRef]
- Saini, M.; Satija, U. An Effective and Robust Framework for Ocular Artifact Removal From Single-Channel EEG Signal Based on Variational Mode Decomposition. IEEE Sens. J. 2020, 20, 369–376. [Google Scholar] [CrossRef]
- Chavez, M.; Grosselin, F.; Bussalb, A.; De Vico Fallani, F.; Navarro-Sune, X. Surrogate-Based Artifact Removal From Single-Channel EEG. IEEE Trans. Neural Syst. Rehabil. Eng. 2018, 26, 540–550. [Google Scholar] [CrossRef]



{kind=link}
{kind=link}
| Authors | Number of Subjects | Channels | Frequency Range | Imagined Movement | Main Conclusions |
|---|---|---|---|---|---|
| Khan and Sepulveda, (2010) [21] | 5 | 64 | 32–48 Hz | Wrist: extension, flexion, pronation, and supination. | An average recognition rate of approximately 89% was achieved in four movement types between the left and right wrists. |
| Kiroi et al., (2012) [22] | 8 | 14 | 31–45 Hz 55–70 Hz | Flexion or oscillatory movement of the arm at the elbow, clenching of the hand. | Increase in activation levels, particularly in the central areas of the cortex. |
| Smith et al., (2014) [23] | 10 | 54 | 70–150 Hz | Finger movement imagery. | Significant power increase was observed during motor imagery. |
| Korik et al., (2018) [24] | 12 | 41 | 28–40 Hz | Imagined 3D limb movement. | The power spectral density contributes to the encoding of movement-related information during arm movement. |
| Lazurenko et al., (2018) [25] | 24 | 17 | 30–48 Hz and 52–70 Hz | Imaginary hand, leg, and tongue movements. | Sensorimotor and associative areas of both hemispheres were actively involved in imaginary and actual movements. |
| Veslin et al., (2019) [26] | 12 | 14 | 35–45 | Right and left elbow movements. | Similar activity was obtained in the gamma band during the preparation and execution of both actual and imaginary movements. |
| Action | GBA | μV2 | Comparison of Means |
|---|---|---|---|
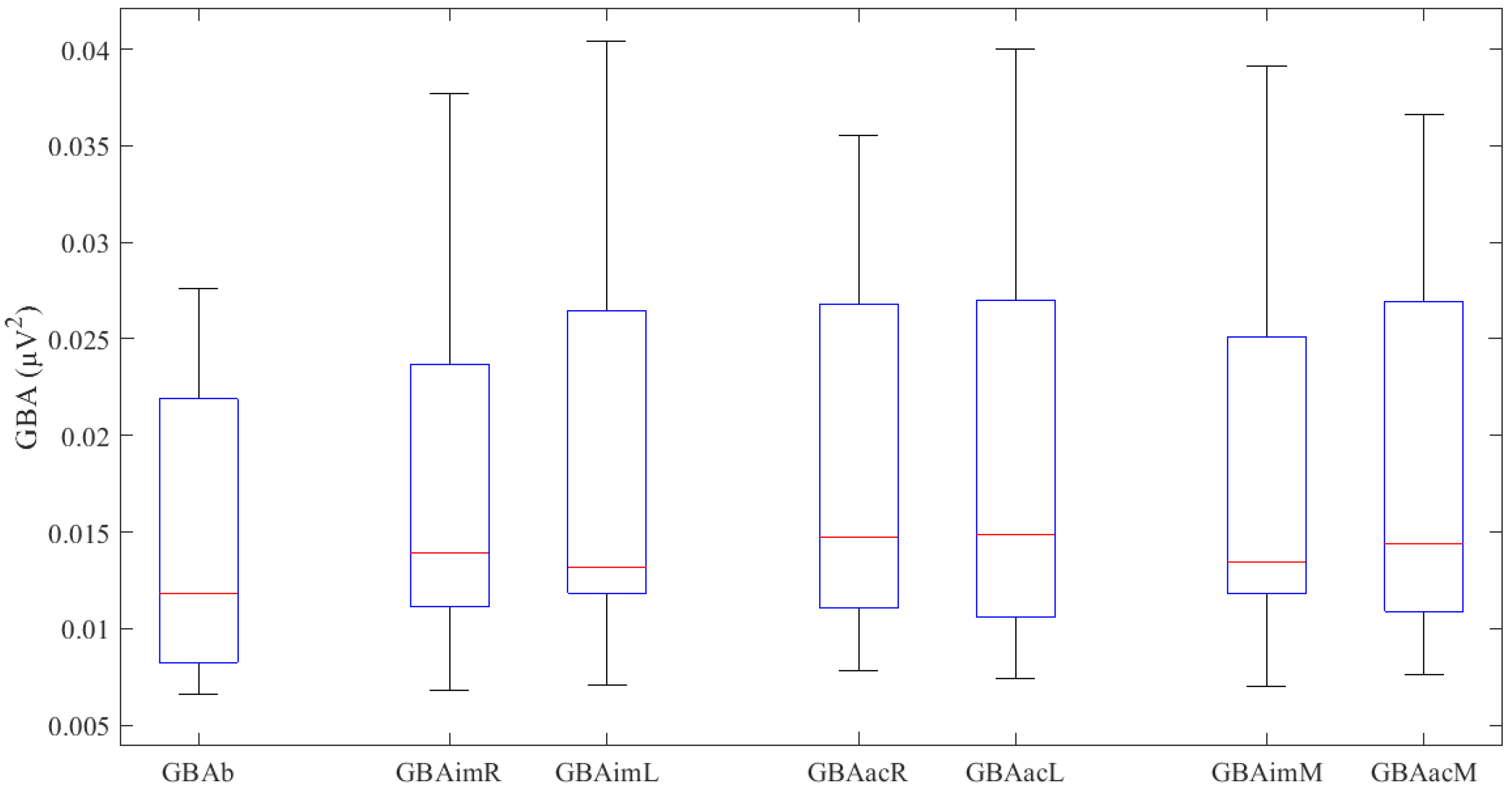
| Basal | GBAb * | 0.0145 ± 0.0076 | ----- |
| Right Hand | GBAimR * | 0.0175 ± 0.0098 | t(11) = −1.251, p = 0.237 |
| GBAacR * | 0.0185 ± 0.0097 | ||
| Left Hand | GBAimL | 0.0131 (0.0159) | Z = 0.275, p = 0.783 |
| GBAacL * | 0.0185 ± 0.0104 | ||
| Mean Values | GBAimM * | 0.0180 ± 0.0101 | t(11) = 1.236, p = 0.242 |
| GBAacM * | 0.0185 ± 0.0099 |
| ERS | ERS (%) | Comparison of Means Wilcoxon Signed-Rank Test | |
|---|---|---|---|
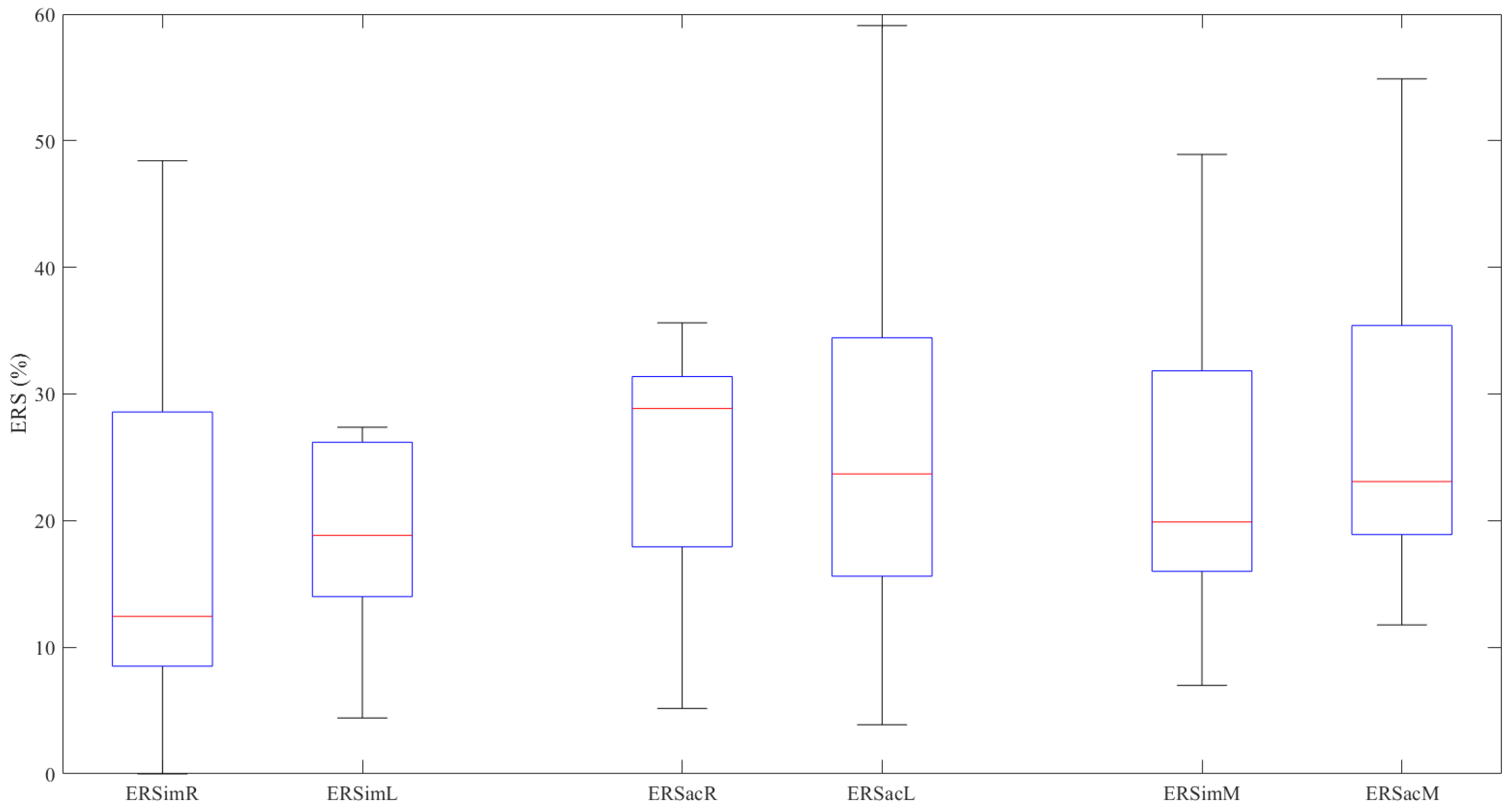
| Right Hand | ERSimR | 12.435 (21.124) | Z = −1.020, p = 0.308 |
| ERSacR | 28.850 (14.889) | ||
| Left Hand | ERSimL | 18.828 (13.578) | Z = −0.471, p = 0.638 |
| ERSacL * | 26.972 ± 17.447 | ||
| Mean Values | ERSimM | 15.983 (14.313) | Z = −1.569, p = 0.117 |
| ERSacM * | 27.479 ± 13.256 |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Amo Usanos, C.; Boquete, L.; de Santiago, L.; Barea Navarro, R.; Cavaliere, C. Induced Gamma-Band Activity during Actual and Imaginary Movements: EEG Analysis. Sensors 2020, 20, 1545. https://doi.org/10.3390/s20061545
Amo Usanos C, Boquete L, de Santiago L, Barea Navarro R, Cavaliere C. Induced Gamma-Band Activity during Actual and Imaginary Movements: EEG Analysis. Sensors. 2020; 20(6):1545. https://doi.org/10.3390/s20061545
Chicago/Turabian StyleAmo Usanos, Carlos, Luciano Boquete, Luis de Santiago, Rafael Barea Navarro, and Carlo Cavaliere. 2020. "Induced Gamma-Band Activity during Actual and Imaginary Movements: EEG Analysis" Sensors 20, no. 6: 1545. https://doi.org/10.3390/s20061545
APA StyleAmo Usanos, C., Boquete, L., de Santiago, L., Barea Navarro, R., & Cavaliere, C. (2020). Induced Gamma-Band Activity during Actual and Imaginary Movements: EEG Analysis. Sensors, 20(6), 1545. https://doi.org/10.3390/s20061545