Wireless Sensing of Lower Lip and Thumb-Index Finger ‘Ramp-and-Hold’ Isometric Force Dynamics in a Small Cohort of Unilateral MCA Stroke: Discussion of Preliminary Findings

 , and
, and
Abstract
1. Introduction
2. Materials and Methods
2.1. Participants
2.2. Instrumentation
2.3. Protocol
2.4. Force Signal Processing
2.5. Statistical Analysis
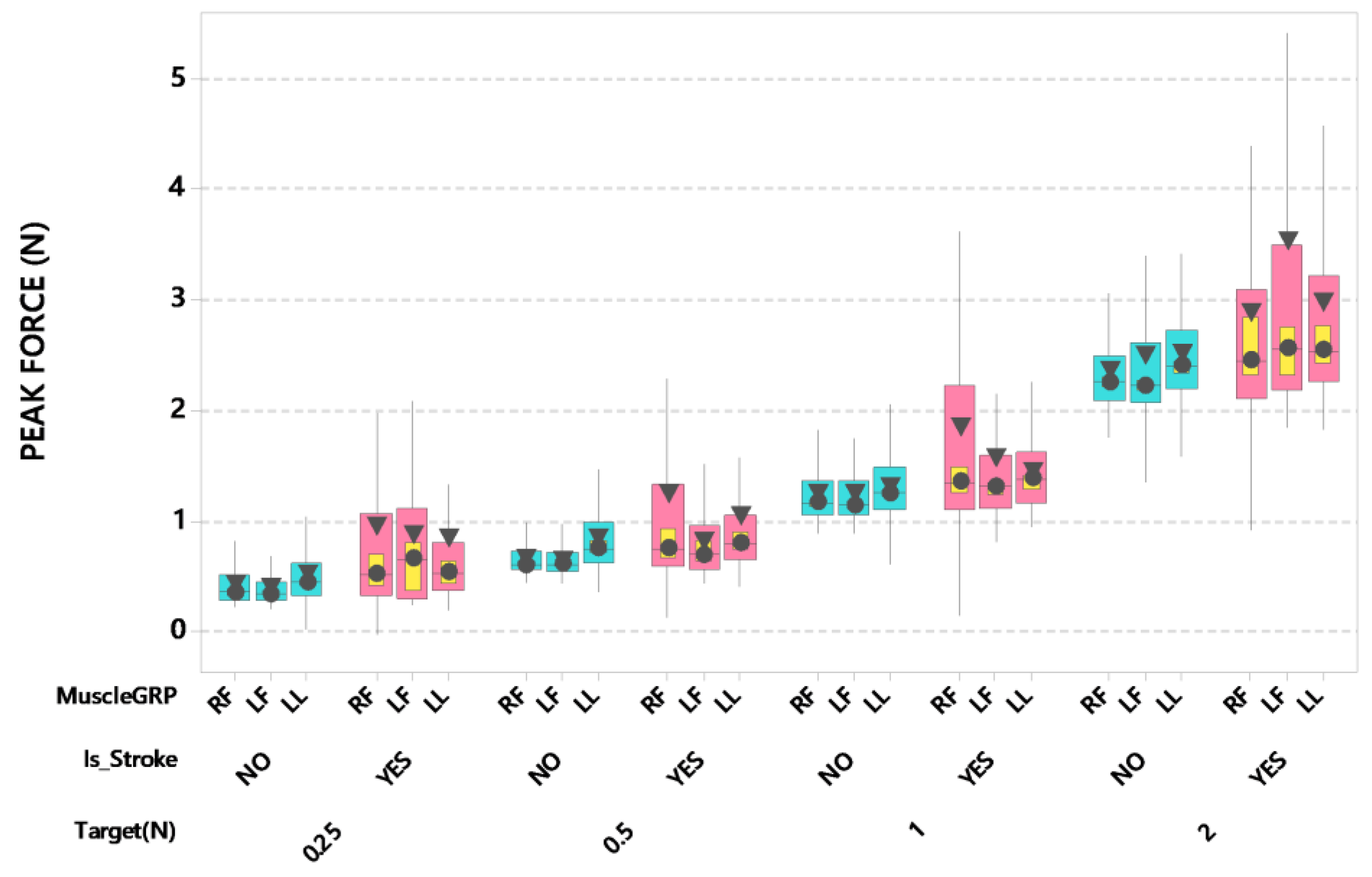
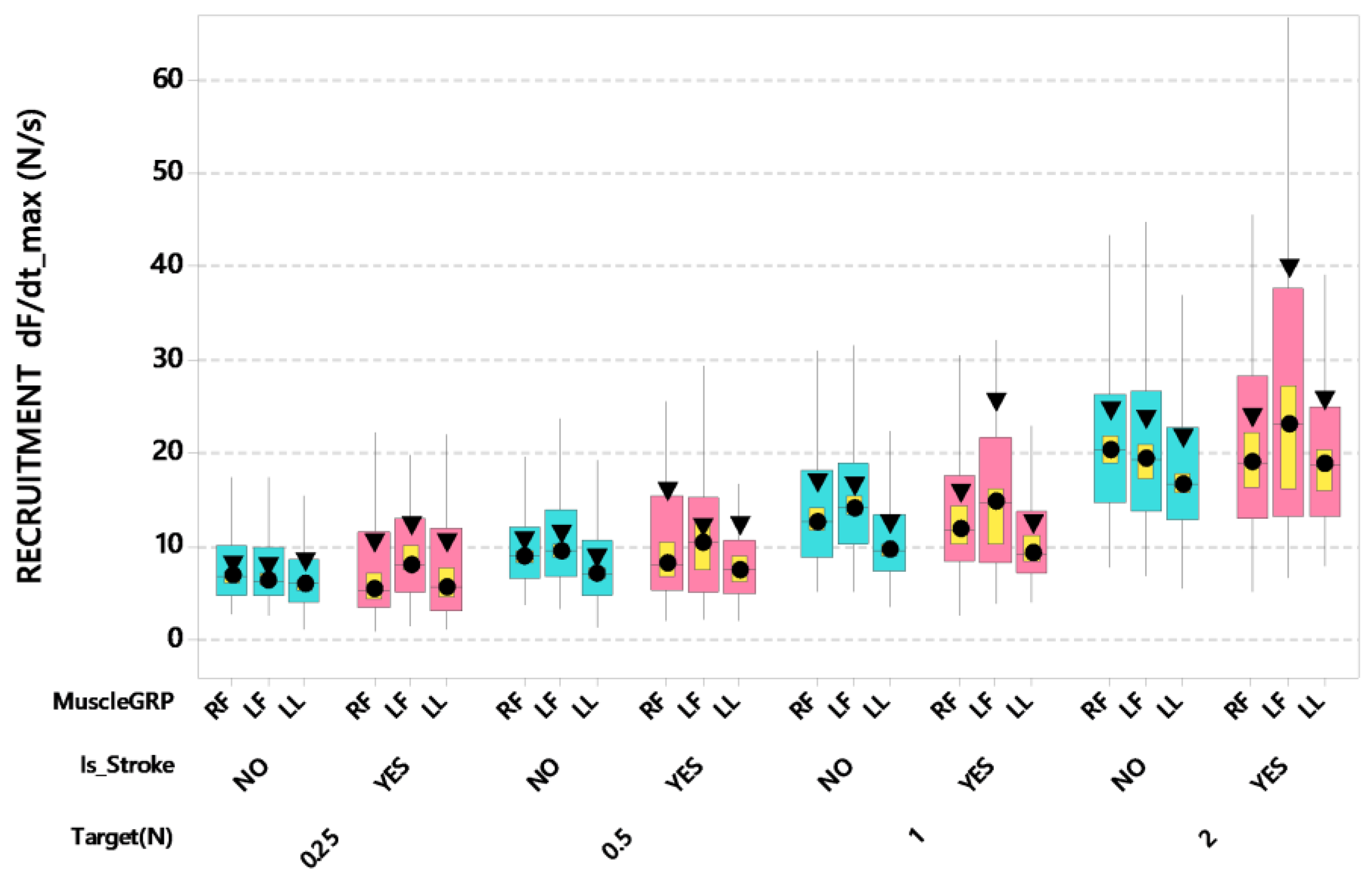
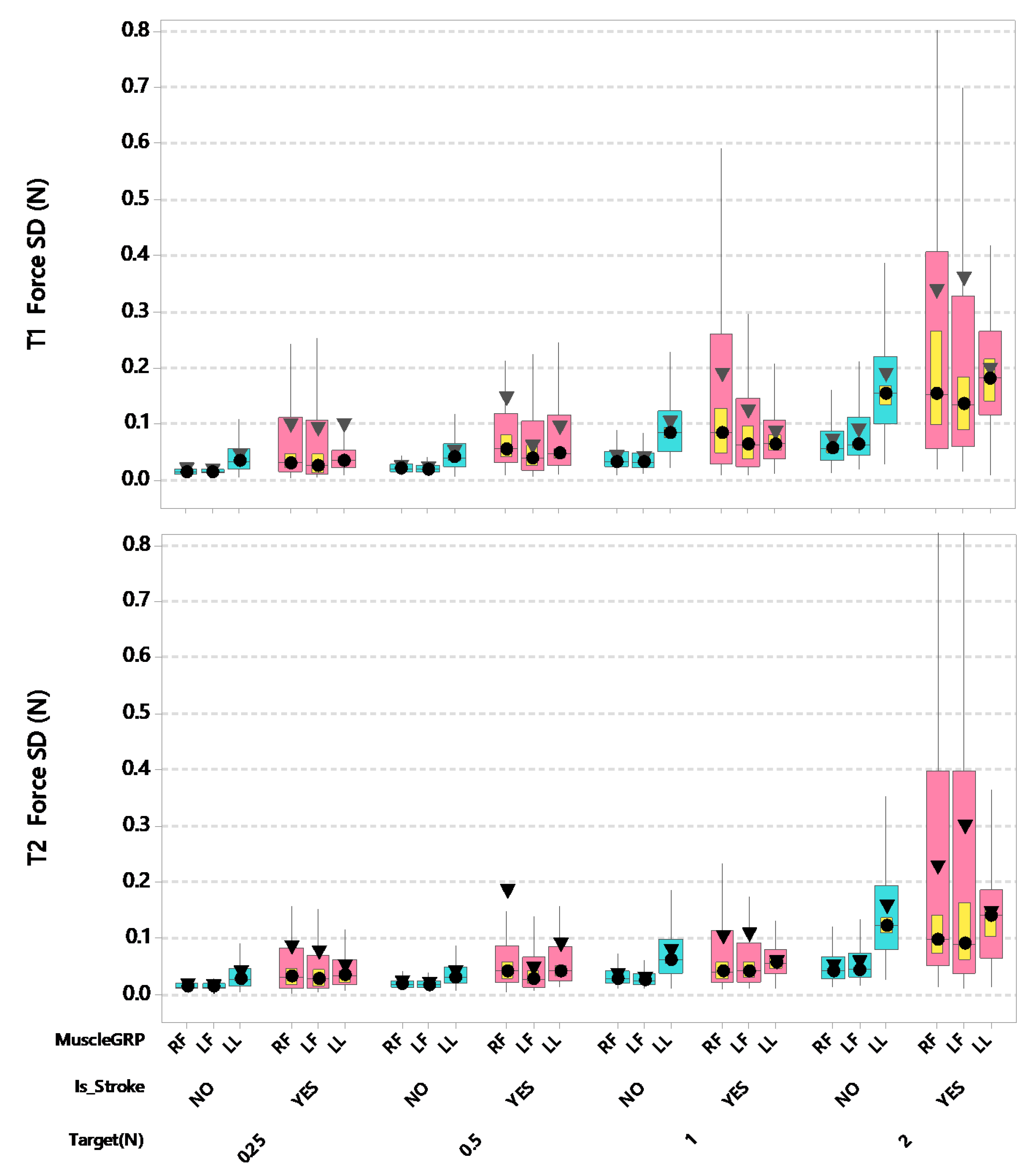
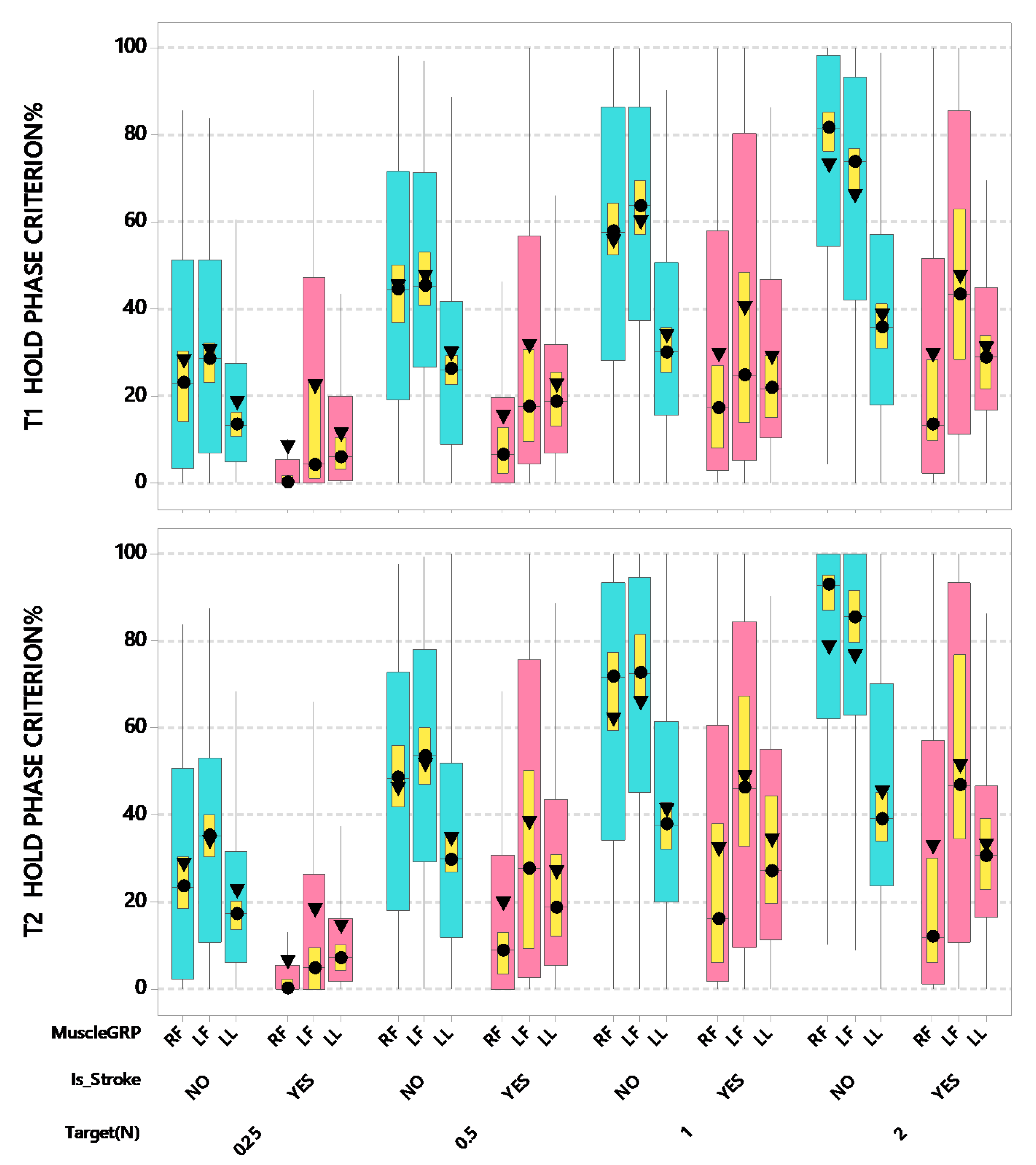
3. Results
4. Discussion
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Johnson, W.D.; Onuma, O.; Owolabi, M.; Sachdev, S. Stroke: A global response is needed. Bull. World Heal. Organ. 2016, 94, 634. [Google Scholar] [CrossRef]
- Dobkin, B.H. Clinical practice. Rehabilitation after stroke. N. Engl. J. Med. 2005, 352, 1677–1684. [Google Scholar] [CrossRef]
- Bustamante, A.; Giralt, D.; García-Berrocoso, T.; Rubiera, M.; Álvarez-Sabín, J.; Molina, C.; Serena, J.; Montaner, J. The impact of post-stroke complications on in-hospital mortality depends on stroke severity. Eur. Stroke J. 2016, 2, 54–63. [Google Scholar] [CrossRef]
- Kang, N.; Cauraugh, J.H. Force control in chronic stroke. Neurosci. Biobehav. Rev. 2015, 52, 38–48. [Google Scholar] [CrossRef] [PubMed]
- Dias, C.P.; Freire, B.; Goulart, N.B.A.; Onzi, E.S.; Becker, J.; Gomes, I.; Arampatzis, A.; Vaz, M.A. Muscle architecture and torque production in stroke survivors: An observational study. Top. Stroke Rehabil. 2016, 24, 206–213. [Google Scholar] [CrossRef] [PubMed]
- Gemperline, J.J.; Allen, S.; Walk, D.; Rymer, W.Z. Characteristics of motor unit discharge in subjects with hemiparesis. Muscle Nerve 1995, 18, 1101–1114. [Google Scholar] [CrossRef] [PubMed]
- Frontera, W.; Grimby, L.; Larsson, L. Firing rate of the lower motoneuron and contractile properties of its muscle fibers after upper motoneuron lesion in man. Muscle Nerve 1997, 20, 938–947. [Google Scholar] [CrossRef]
- Hara, Y.; Masakado, Y.; Chino, N. The physiological functional loss of single thenar motor units in the stroke patients: When does it occur? Does it progress? Clin. Neurophysiol. 2004, 115, 97–103. [Google Scholar] [CrossRef]
- Lukács, M. Electrophysiological signs of changes in motor units after ischaemic stroke. Clin. Neurophysiol. 2005, 116, 1566–1570. [Google Scholar] [CrossRef]
- Schimmel, M.; Leemann, B.; Schnider, A.; Herrmann, F.R.; Kiliaridis, S.; Müller, F. Changes in oro-facial function and hand-grip strength during a 2-year observation period after stroke. Clin. Oral Investig. 2012, 17, 867–876. [Google Scholar] [CrossRef]
- Hsu, H.-Y.; Lin, C.-F.; Su, F.-C.; Kuo, H.-T.; Chiu, H.-Y.; Kuo, L.-C. Clinical application of computerized evaluation and re-education biofeedback prototype for sensorimotor control of the hand in stroke patients. J. Neuroeng. Rehabil. 2012, 9, 26. [Google Scholar] [CrossRef] [PubMed]
- Kamper, D.G.; Fischer, H.C.; Conrad, M.O.; Towles, J.D.; Rymer, W.Z.; Triandafilou, K.M. Finger-thumb coupling contributes to exaggerated thumb flexion in stroke survivors. J. Neurophysiol. 2014, 111, 2665–2674. [Google Scholar] [CrossRef] [PubMed]
- Seo, N.J.; Enders, L.; Motawar, B.; Kosmopoulos, M.L.; Fathi-Firoozabad, M. The extent of altered digit force direction correlates with clinical upper extremity impairment in chronic stroke survivors. J. Biomech. 2014, 48, 383–387. [Google Scholar] [CrossRef] [PubMed]
- Liang, L.; Chen, J.; Wang, Y.; Jakubiec, M.; Mierzejewska, J.; Piotrkiewicz, M. Changes in spinal motoneuron “fastness” in post-stroke spastic patients. J. Med. Biol. Eng. 2010, 30, 17–22. [Google Scholar]
- Chou, L.-W.; Palmer, J.; Binder-Macleod, S.; Josephson, M.D. Motor unit rate coding is severely impaired during forceful and fast muscular contractions in individuals post stroke. J. Neurophysiol. 2013, 109, 2947–2954. [Google Scholar] [CrossRef] [PubMed]
- Ni, J.; Li, M.-L.; Yao, M.; Cui, L. Early corticospinal tract Wallerian degeneration on diffusion-weighted MR imaging after adult stroke: Three cases report. Clin. Neurol. Neurosurg. 2013, 115, 1164–1166. [Google Scholar] [CrossRef] [PubMed]
- Ye, Y.; Ma, L.; Yan, T.; Liu, H.; Wei, X.; Song, R. Kinetic measurements of hand motor impairments after mild to moderate stroke using grip control tasks. J. Neuroeng. Rehabil. 2014, 11, 84. [Google Scholar] [CrossRef]
- Rosenfalck, A.; Andreassen, S. Impaired regulation of force and firing pattern of single motor units in patients with spasticity. J. Neurol. Neurosurg. Psychiatry 1980, 43, 907–916. [Google Scholar] [CrossRef][Green Version]
- Kinoshita, T.; Moritani, T.; Shrier, D.A.; Wang, H.Z.; Hiwatashi, A.; Numaguchi, Y.; Westesson, P.-L.A. Secondary degeneration of the substantia nigra and corticospinal tract after hemorrhagic middle cerebral artery infarction: Diffusion-weighted MR findings. Magn. Reson. Med Sci. 2002, 1, 175–178. [Google Scholar] [CrossRef]
- Lukacs, M.; Vécsei, L.; Beniczky, S. Large motor units are selectively affected following a stroke. Clin. Neurophysiol. 2008, 119, 2555–2558. [Google Scholar] [CrossRef]
- Mottram, C.J.; Heckman, C.J.; Powers, R.K.; Rymer, W.Z.; Suresh, N.L. Disturbances of motor unit rate modulation are prevalent in muscles of spastic-paretic stroke survivors. J. Neurophysiol. 2014, 111, 2017–2028. [Google Scholar] [CrossRef] [PubMed]
- Weeks, K.; Dzielak, D.; Hamadain, E.; Bailey, J. Examining the Relationship Between Stroke and Labial Strength. Contemp. Issues Commun. Sci. Disord. CICSD 2013, 40, 160–169. [Google Scholar] [CrossRef]
- Arce-McShane, F.; Hatsopoulos, N.G.; Lee, J.-C.; Ross, C.F.; Sessle, B.J. Modulation Dynamics in the Orofacial Sensorimotor Cortex during Motor Skill Acquisition. J. Neurosci. 2014, 34, 5985–5997. [Google Scholar] [CrossRef] [PubMed]
- Dai, R.; Lam, O.L.; Lo, E.C.; Li, L.S.; Wen, Y.; McGrath, C. Orofacial functional impairments among patients following stroke: A systematic review. Oral Dis. 2014, 21, 836–849. [Google Scholar] [CrossRef]
- Schimmel, M.; Ono, T.; Lam, O.L.T.; Müller, F. Oro-facial impairment in stroke patients. J. Oral Rehabil. 2017, 44, 313–326. [Google Scholar] [CrossRef]
- Barlow, S.M.; Burton, M.K. Ramp-and-hold force control in the upper and lower lips: Developing new neuromotor assessment applications in traumatically brain injured adults. J. Speech Hear. Res. 1990, 33, 660–675. [Google Scholar] [CrossRef]
- Andreatta, R.D.; Barlow, S.M. Somatosensory Gating Is Dependent on the Rate of Force Recruitment in the Human Orofacial System. J. Speech Lang. Hear. Res. 2009, 52, 1566–1578. [Google Scholar] [CrossRef]
- Barlow, S.M.; Abbs, J.H. Orofacial fine motor control impairments in congenital spasticity: Evidence against hypertonus-related performance deficits. Neurology 1984, 34, 145–150. [Google Scholar] [CrossRef]
- Barlow, S.M.; Abbs, J.H. Fine force and position control of select orofacial structures in the upper motor neuron syndrome. Exp. Neurol. 1986, 94, 699–713. [Google Scholar] [CrossRef]
- Andreatta, R.D.; Barlow, S.M.; Biswas, A.; Finan, D.S. Mechanosensory Modulation of Perioral Neuronal Groups During Active Force Dynamics. J. Speech, Lang. Hear. Res. 1996, 39, 1006–1017. [Google Scholar] [CrossRef]
- Estep, M.; Barlow, S.M. Modulation of the trigeminofacial pathway during syllabic speech. Brain Res. 2007, 1171, 67–74. [Google Scholar] [CrossRef] [PubMed]
- Greenwood, J.; Hozan, M.; Barlow, S.M. Wireless Isometric Force Sensor Platform for Bedside Automated Evaluation and Therapeutics of Orofacial and Hand Muscle Systems Across the Lifespan. Biomed. J. Sci. Tech. Res. 2019, 21, 15824–15829. [Google Scholar] [CrossRef]
- Barlow, S.; Hozan, M.; Lee, J.; Greenwood, J.; Custead, R.; Wardyn, B.; Tippin, K. Orofacial and thumb-index finger ramp-and-hold isometric force dynamics in young neurotypical adults. J. Biomech. 2018, 72, 81–89. [Google Scholar] [CrossRef] [PubMed]
- SAS Institute. SAS/STAT 9.2 User’s Guide; SAS Institute Inc.: Cary, NC, USA, 2002–2012.
- Barlow, S.M. Real time modulation of speech-orofacial motor performance by means of motion sense. J. Commun. Disord. 1998, 31, 511–534. [Google Scholar] [CrossRef]
- Cole, K.J. Hand Motor Control: Maturing an Immature Science. Mot. Control. 2015, 19, 131–134. [Google Scholar] [CrossRef]
- Latash, M.L.; Friedman, J.; Kim, S.W.; Feldman, A.G.; Zatsiorsky, V.M. Prehension synergies and control with referent hand configurations. Exp. Brain Res. 2009, 202, 213–229. [Google Scholar] [CrossRef]
- Gracco, V.L.; Abbs, J.H. Dynamic control of the perioral system during speech: Kinematic analyses of autogenic and nonautogenic sensorimotor processes. J. Neurophysiol. 1985, 54, 418–432. [Google Scholar] [CrossRef]
- Barlow, S.M.; Muller, E.M. The Relation Between Interangle Span and In Vivo Resultant Force in the Perioral Musculature. J. Speech, Lang. Hear. Res. 1991, 34, 252–259. [Google Scholar] [CrossRef]
- Oluwatosin, O.M. Orofacial indices: A study in 240 Nigerian children. Afr. J. Med. Med Sci. 1999, 27, 39–42. [Google Scholar]
- Shiller, D.M.; LaBoissière, R.; Ostry, D.J. Relationship Between Jaw Stiffness and Kinematic Variability in Speech. J. Neurophysiol. 2002, 88, 2329–2340. [Google Scholar] [CrossRef]
- Chu, S.-Y.; Barlow, S.; Kieweg, D.; Lee, J. OroSTIFF: Face-referenced measurement of perioral stiffness in health and disease. J. Biomech. 2010, 43, 1476–1482. [Google Scholar] [CrossRef] [PubMed][Green Version]
- Mayo, N.E.; Wood-Dauphinee, S.; Ahmed, S.; Gordon, C.; Higgins, J.; McEwen, S.; Salbach, N.; Mayo, S.W.-D.N.E. Disablement following stroke. Disabil. Rehabil. 1999, 21, 258–268. [Google Scholar] [CrossRef] [PubMed]
- Vermillion, B.C.; Dromerick, A.; Lee, S.W. Toward Restoration of Normal Mechanics of Functional Hand Tasks Post-Stroke: Subject-Specific Approach to Reinforce Impaired Muscle Function. IEEE Trans. Neural Syst. Rehabil. Eng. 2019, 27, 1606–1616. [Google Scholar] [CrossRef] [PubMed]
- Schimmel, M.; Voegeli, G.; Duvernay, E.; Leemann, B.; Müller, F. Oral tactile sensitivity and masticatory performance are impaired in stroke patients. J. Oral Rehabil. 2017, 44, 163–171. [Google Scholar] [CrossRef]
- Barlow, S.M.; Custead, R.; Lee, J. Adaptive Vibrotactile Threshold Estimation of the Glabrous Hand And Perioral Face Following MCA Stroke. Biomed. J. Sci. Tech. Res. 2019, 23, 17399–17406. [Google Scholar] [CrossRef]
- Colebatch, J.G.; Gandevia, S.C. THE DISTRIBUTION OF MUSCULAR WEAKNESS IN UPPER MOTOR NEURON LESIONS AFFECTING THE ARM. Brain 1989, 112, 749–763. [Google Scholar] [CrossRef]
- Saladin, L.K. Cerebrovascular disease: Stroke. In Pathophysiology of the Motor Systems: Principles and Clinical Presentations; Saladin, L.K., Fredericks, C.M., Eds.; F.A. Davis Co.: Philadelphia, PA, USA, 1996; pp. 486–512. [Google Scholar]
- Mosier, K.; Lau, C.; Wang, Y.; Venkadesan, M.; Valero-Cuevas, F.J. Controlling instabilities in manipulation requires specific cortical-striatal-cerebellar networks. J. Neurophysiol. 2011, 105, 1295–1305. [Google Scholar] [CrossRef]
- Carod-Artal, J.; A Egido, J.; González, J.L.; De Seijas, E.V. Quality of life among stroke survivors evaluated 1 year after stroke: Experience of a stroke unit. Stroke 2000, 31, 2995–3000. [Google Scholar] [CrossRef]
- Clarke, P.; Black, S. Quality of Life Following Stroke: Negotiating Disability, Identity, and Resources. J. Appl. Gerontol. 2005, 24, 319–336. [Google Scholar] [CrossRef]
- Nichols-Larsen, D.; Clark, P.; Zeringue, A.; Greenspan, A.; Blanton, S. Factors Influencing Stroke Survivors’ Quality of Life During Subacute Recovery. Stroke 2005, 36, 1480–1484. [Google Scholar] [CrossRef]
- Li, K.; Nataraj, R.; Marquardt, T.L.; Li, Z.-M. Directional Coordination of Thumb and Finger Forces during Precision Pinch. PLOS ONE 2013, 8, e79400. [Google Scholar] [CrossRef] [PubMed]
- Westling, G.; Johansson, R.S. Responses in glabrous skin mechanoreceptors during precision grip in humans. Exp. Brain Res. 1987, 66, 128–140. [Google Scholar] [CrossRef] [PubMed]
- Müller, E.M.; Milenkovic, P.H.; MacLeod, G.E. Perioral tissue mechanics during speech production. In Mathematics and Computers in Biomedical Application; Eisenfeld, J., DeLisi, C., Eds.; Elsevier: Amsterdam, The Netherlands, 1985; pp. 363–371. [Google Scholar]
- Etter, N.M.; McKeon, P.; Dressler, E.; Andreatta, R.D. Effects of ageing on orofacial fine force control and its relationship with parallel change in sensory perception. Int. J. Speech-Language Pathol. 2017, 20, 502–515. [Google Scholar] [CrossRef] [PubMed]
- Johansson, R.S.; Hager-Ross, C.; Riso, R. Somatosensory control of precision grip during unpredictable pulling loads – II. Changes in load force rate. Exp. Brain Res. 1992, 89, 192–203. [Google Scholar] [CrossRef]
- Macefield, V.; Johansson, R.S. Control of grip force during restraint of an object held between finger and thumb: Responses of muscle and joint afferents from the digits. Exp. Brain Res. 1996, 108, 172–184. [Google Scholar] [CrossRef]
- Johansson, R.S.; Trulsson, M.; Olsson, K.Å.; Abbs, J.H. Mechanoreceptive afferent activity in the infraorbital nerve in man during speech and chewing movements. Exp. Brain Res. 1988, 72, 209–214. [Google Scholar] [CrossRef]
- Andreatta, R.D.; Barlow, S.; Finan, D.S. Modulation of the mechanically evoked perioral reflex during active force dynamics in young adults. Brain Res. 1994, 646, 175–179. [Google Scholar] [CrossRef]
- Trulsson, M.; Johansson, R.S. Orofacial mechanoreceptors in humans: Encoding characteristics and responses during natural orofacial behaviors. Behav. Brain Res. 2002, 135, 27–33. [Google Scholar] [CrossRef]
- Coronado, J.; Ulloa-Perez, A.; Garcia-Cordova, F.; Villalba-Fernandez, J. Experimental development for thumb-index pinch force measurements during object manipulation with application to robotic hands. In Proceedings of the 1999 IEEE International Conference on Systems, Man, and Cybernetics (Cat. No.99CH37028), Tokyo, Japan, 12–15 October 2003; Volume 6, pp. 786–791. [Google Scholar]
- McNeil, M.R.; Weismer, G.; Adams, S.; Mulligan, M. Oral structure nonspeech motor control in normal, dysarthric, aphasic and apraxic speakers: Isometric force and static position control. J. Speech Hear. Res. 1990, 33, 255–268. [Google Scholar] [CrossRef]
- Barlow, S.M.; Iacono, R.P.; Paseman, L.A.; Biswas, A.; D’Antonio, L.D. The effects of experimental posteroventral pallidotomy on force and speech aerodynamics in Parkinson’s disease. In Speech Motor Control; Cannito, M.P., Yorkston, K.M., Beukelman, D.R., Eds.; Paul H. Brookes Publishing Company: Baltimore, MD, USA, 1998; pp. 117–156. [Google Scholar]
- DePaul, R.; Abbs, J.H.; Caligiuri, M.; Gracco, V.L.; Brooks, B.R. Hypoglossal, trigeminal, and facial motoneuron involvement in amyotrophic lateral sclerosis. Neurology 1988, 38, 281. [Google Scholar] [CrossRef]
- Dworkin, J.P. Tongue strength measurement in patients with amyotrophic lateral sclerosis: Qualitative vs quantitative procedures. Arch. Phys. Med. Rehabil. 1980, 61, 422–424. [Google Scholar]
- Barlow, S.M.; Rath, E.M. Maximum Voluntary Closing Forces in the Upper and Lower Lips of Humans. J. Speech, Lang. Hear. Res. 1985, 28, 373–376. [Google Scholar] [CrossRef] [PubMed]
- An, K.N.; Askew, L.; Chao, E.Y. Biomechanics and Functional Assessment of Upper Extremities. In Trends in Ergonomics/Human Factors III; Karwowski, W., Ed.; Elsevier Science Ltd.: Amsterdam, The Netherlands, 1986; pp. 573–580. [Google Scholar]
- Chen, F.C.; Favetto, A.; Mousavi, M.; Ambrosio, E.; Appendino, S.; Manfredi, D.; Pescarmona, F.; Calafiore, G.; Bona, B. Human Hand: Kinematics, Statics, and Dynamics. In Proceedings of the 41st International Conference on Environmental Systems; American Institute of Aeronautics and Astronautics (AIAA), Portland, OR, USA,, 17–21 July 2011. [Google Scholar]
- Toda, T.; Taoka, M. Converging patterns of inputs from oral structures in the postcentral somatosensory cortex of conscious macaque monkeys. Exp. Brain Res. 2004, 158, 43–49. [Google Scholar] [CrossRef] [PubMed]
- Barlow, S.M.; Bashford, G.; Singh, S. Neuroprotection in Acute MCA Stroke by Somatosensory-Induced Collateral Blood Flow; R01 NS117741-01; NINDS: Bethesda, MD, USA, 2019. [Google Scholar]
- Evarts, E.V. Role of Motor Cortex in Voluntary Movements in Primates. In Handbook of Physiology: Motor Control, Part 2; Brooks, V.B., Ed.; American Physiological Society: Rockville, MD, USA, 2011; pp. 1083–1120. [Google Scholar]
- Smith, A.; Hepp-Reymond, M.-C.; Wyss, U. Relation of activity in precentral cortical neurons to force and rate of force change during isometric contractions of finger muscles. Exp. Brain Res. 1975, 23, 315–332. [Google Scholar] [CrossRef] [PubMed]
- Humphrey, D.R.; Schmidt, E.M.; Thompson, W.D. Predicting Measures of Motor Performance from Multiple Cortical Spike Trains. Science 1970, 170, 758–762. [Google Scholar] [CrossRef] [PubMed]
- Cheney, P.D.; Fetz, E.E. Functional classes of primate corticomotoneuronal cells and their relation to active force. J. Neurophysiol. 1980, 44, 773–791. [Google Scholar] [CrossRef] [PubMed]
- Hepp-Reymond, M.C.; Wyss, U.R.; Anner, R. Neuronal coding of static force in the primate motor cortex. J. Physiol. 1978, 74, 287–291. [Google Scholar]
- Sanes, J.; Evarts, E.V. Regulatory role of proprioceptive input in motor control of phasic or maintained voluntary contractions in man. Adv. Neurol. 1983, 39, 47–59. [Google Scholar]
- Rizzolatti, G.; Fadiga, L.; Matelli, M.; Bettinardi, V.; Paulesu, E.; Perani, D.; Fazio, G. Localization of grasp representations in humans by PET: 1. Observation vs. execution. Exp. Brain Res. 1996, 111, 246–252. [Google Scholar] [CrossRef]
- Jeannerod, M.; Arbib, M.; Rizzolatti, G.; Sakata, H. Grasping objects: The cortical mechanisms of visuomotor transformation. Trends Neurosci. 1995, 18, 314–320. [Google Scholar] [CrossRef]
- Binkofski, F.; Buccino, G.; Posse, S.; Seitz, R.J.; Rizzolatti, G.; Freund, H.-J. A fronto-parietal circuit for object manipulation in man: Evidence from an fMRI-study. Eur. J. Neurosci. 1999, 11, 3276–3286. [Google Scholar] [CrossRef] [PubMed]
- Kuhtz-Buschbeck, J.P.; Ehrsson, H.H.; Forssberg, H. Human brain activity in the control of fine static precision grip forces: An fMRI study. Eur. J. Neurosci. 2001, 14, 382–390. [Google Scholar] [CrossRef] [PubMed]
- Ehrsson, H.H.; Fagergren, A.; Jonsson, T.; Westling, G.; Johansson, R.S.; Forssberg, H. Cortical activity in precision- versus power-grip tasks: An fMRI study. J. Neurophysiol. 2000, 83, 528–536. [Google Scholar] [CrossRef] [PubMed]
- Ehrsson, H.H.; Fagergren, A.; Forssberg, H. Differential Fronto-Parietal Activation Depending on Force Used in a Precision Grip Task: An fMRI Study. J. Neurophysiol. 2001, 85, 2613–2623. [Google Scholar] [CrossRef]
- Ehrsson, H.H.; Fagergren, A.; Johansson, R.S.; Forssberg, H. Evidence for the Involvement of the Posterior Parietal Cortex in Coordination of Fingertip Forces for Grasp Stability in Manipulation. J. Neurophysiol. 2003, 90, 2978–2986. [Google Scholar] [CrossRef]
- Tunik, E.; Frey, S.H.; Grafton, S.T. Virtual lesions of the anterior intraparietal area disrupt goal-dependent on-line adjustments of grasp. Nat. Neurosci. 2005, 8, 505–511. [Google Scholar] [CrossRef]
- Herzfeld, D.J.; Pastor, D.; Haith, A.M.; Rossetti, Y.; Shadmehr, R.; O’Shea, J. Contributions of the cerebellum and the motor cortex to acquisition and retention of motor memories. NeuroImage 2014, 98, 147–158. [Google Scholar] [CrossRef]
- Trulsson, M.; Essick, G. Mechanosensation. In Clinical Oral Physiology; Miles, T.S., Nauntofte, B., Svensson, P., Eds.; Quintessence Books: Copenhagen, Denmark, 2004; pp. 165–197. [Google Scholar]
- Dubner, R.; Sessle, B.J.; Storey, A.T. The Neural Basis of Oral and Facial Function; Springer Science and Business Media LLC: Berlin/Heidelberg, Germany, 1978. [Google Scholar]
- Halata, Z.; Munger, B. The sensory innervation of primate facial skin. II. Vermilion border and mucosal lip. Brain Res. 1983, 286, 81–107. [Google Scholar] [CrossRef]
- Munger, B.; Halata, Z. The sensory innervation of primate facial skin. I. Hairy skin. Brain Res. 1983, 286, 45–80. [Google Scholar] [CrossRef]
- Verrillo, R.T. Age Related Changes in the Sensitivity to Vibration. J. Gerontol. 1980, 35, 185–193. [Google Scholar] [CrossRef]
- Barlow, S.M. Mechanical frequency detection thresholds in the human face. Exp. Neurol. 1987, 96, 253–261. [Google Scholar] [CrossRef]
- Barlow, S.M. Vibrography: Single-Interval Up/Down (SIUD) Adaptive Vibrotactile Threshold Estimation of the Glabrous Hand and Perioral Face in Neurotypical Adults. Biomed. J. Sci. Tech. Res. 2019, 22, 16837–16847. [Google Scholar] [CrossRef]
- Verrillo, R.T.; Ecker, A.D. Effects of root or nerve destruction on vibrotactile sensitivity in trigeminal neuralgia. Pain 1977, 3, 239–255. [Google Scholar] [CrossRef]
- Andreatta, R.D.; Davidow, J.H. Mechanical frequency and stimulation-site-related differences in vibrotactile detection capacity along the lip vermilion in young adults. Clin. Oral Investig. 2005, 10, 17–22. [Google Scholar] [CrossRef] [PubMed]
- Venkatesan, L.; Barlow, S.; Kieweg, D. Age- and sex-related changes in vibrotactile sensitivity of hand and face in neurotypical adults. Somatosens. Mot. Res. 2014, 32, 44–50. [Google Scholar] [CrossRef]
- Barlow, S.M.; Poore, M.; Chu, S. Biodynamics of Speech and Orofacial Movement. In Assessment of Motor Speech Disorders; Lowit, A., Kent, R.D., Eds.; Plural Publishing Group: San Diego, CA, USA, 2011; pp. 193–229. [Google Scholar]
- Stal, P.; Eriksson, P.-O.; Eriksson, A.; Thornell, L.-E. Enzyme-histochemical differences in fibre-type between the human major and minor zygomatic and the first dorsal interosseus muscles. Arch. Oral Boil. 1987, 32, 833–841. [Google Scholar] [CrossRef]
- Stal, P.; Eriksson, P.-O.; Eriksson, A.; Thornell, L.-E. Enzyme-histochemical and morphological characteristics of muscle fibre types in the human buccinator and orbicularis oris. Arch. Oral Boil. 1990, 35, 449–458. [Google Scholar] [CrossRef]
- Nordin, M.; Hagbarth, K.-E. Mechanoreceptive units in the human infra-orbital nerve. Acta Physiol. Scand. 1989, 135, 149–161. [Google Scholar] [CrossRef]
- Desmyttere, G.; Mathieu, E.; Begon, M.; Simoneau-Buessinger, E.; Cremoux, S. Effect of the phase of force production on corticomuscular coherence with agonist and antagonist muscles. Eur. J. Neurosci. 2018, 48, 3288–3298. [Google Scholar] [CrossRef]
- Sullivan, M. Oral Angle Ramp-And-Hold Isometric Force Dynamics in Young Neurotypical Adults. Master’s Thesis, University of Nebraska, Lincoln, NE, USA, 2019. [Google Scholar]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| ID | Age (yrs) | Sex | Lesion Territory | Post-Stroke (mos) | Anatomic MRI (MPRAGE) | Fugl-Meyer Upper Ext Motor Score (66 Max) | |||
|---|---|---|---|---|---|---|---|---|---|
| Sagittal | Coronal | Transverse | Right | Left | |||||
| 1 | 67 | M | L-MCA | 70 |  | 59 | 66 | ||
| 2 | 31 | M | L-MCA | 37 |  | 60 | 66 | ||
| 3 | 23 | M | L-MCA | 143 |  | 34 | 65 | ||
| 4 | 47 | M | L-MCA | 38 |  | 38 | 66 | ||
| 5 | 66 | M | R-MCA | 87 |  | 66 | 24 | ||
| 6 | 26 | M | R-MCA | 30 |  | 60 | 33 | ||
| 7 | 67 | M | L-MCA | 86 |  | 66 | 66 | ||
| Mass(g) | 0 | 20 | 50 | 100 | 200 | 500 | R2 | |
|---|---|---|---|---|---|---|---|---|
| Mean | Finger | −0.0126 | 0.2639 | 0.5531 | 1.0377 | 1.9541 | 4.8345 | 0.9996 |
| Lip | −0.0001 | 0.1981 | 0.4966 | 0.9943 | 2.0075 | 5.1500 | 0.9999 | |
| Variance | Finger | 2.9 × 10−5 | 3.8 × 10−5 | 5.7 × 10−5 | 8.5 × 10−5 | 1.1 × 10−4 | 7.6 × 10−4 | |
| Lip | 2.8 × 10−6 | 2.8 × 10−4 | 1.8 × 10−3 | 2.3 × 10−5 | 3.4 × 10−4 | 2.1 × 10−3 |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Barlow, S.; Custead, R.; Lee, J.; Hozan, M.; Greenwood, J. Wireless Sensing of Lower Lip and Thumb-Index Finger ‘Ramp-and-Hold’ Isometric Force Dynamics in a Small Cohort of Unilateral MCA Stroke: Discussion of Preliminary Findings. Sensors 2020, 20, 1221. https://doi.org/10.3390/s20041221
Barlow S, Custead R, Lee J, Hozan M, Greenwood J. Wireless Sensing of Lower Lip and Thumb-Index Finger ‘Ramp-and-Hold’ Isometric Force Dynamics in a Small Cohort of Unilateral MCA Stroke: Discussion of Preliminary Findings. Sensors. 2020; 20(4):1221. https://doi.org/10.3390/s20041221
Chicago/Turabian StyleBarlow, Steven, Rebecca Custead, Jaehoon Lee, Mohsen Hozan, and Jacob Greenwood. 2020. "Wireless Sensing of Lower Lip and Thumb-Index Finger ‘Ramp-and-Hold’ Isometric Force Dynamics in a Small Cohort of Unilateral MCA Stroke: Discussion of Preliminary Findings" Sensors 20, no. 4: 1221. https://doi.org/10.3390/s20041221
APA StyleBarlow, S., Custead, R., Lee, J., Hozan, M., & Greenwood, J. (2020). Wireless Sensing of Lower Lip and Thumb-Index Finger ‘Ramp-and-Hold’ Isometric Force Dynamics in a Small Cohort of Unilateral MCA Stroke: Discussion of Preliminary Findings. Sensors, 20(4), 1221. https://doi.org/10.3390/s20041221