Recent Advances in the Development of Biosensors for Malaria Diagnosis


Abstract
1. Introduction
2. Parasite Development in Humans, Biomarkers, and Diagnosis
3. Biosensors for the Detection of Malaria Biomarkers in Clinical Samples
3.1. Detection of PfHRP-2 in Clinical Samples
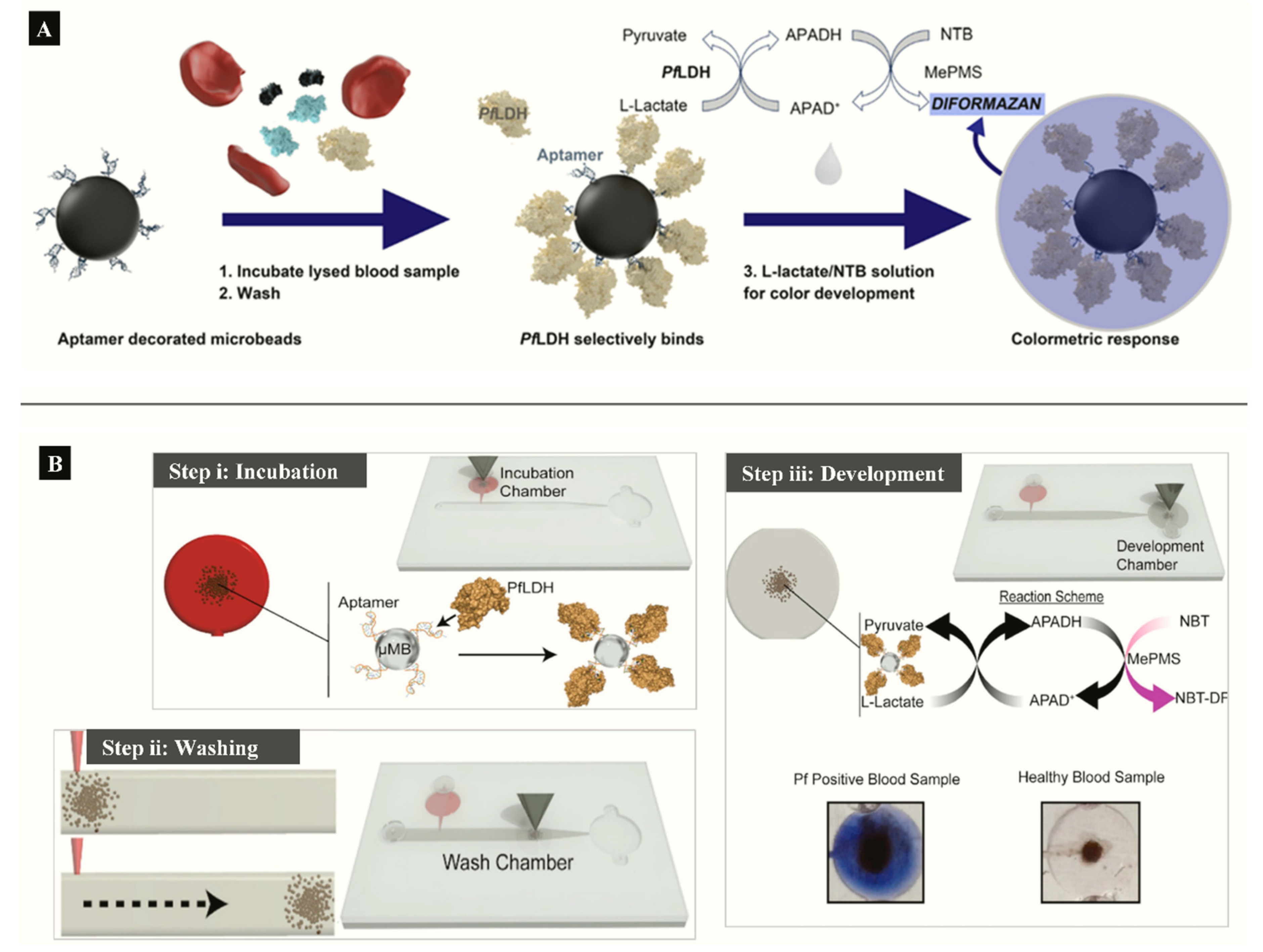
3.2. Detection of pLDH in Clinical Samples
3.3. Detection of GDH in Clinical Samples
3.4. Detection of Aldolase
3.5. Detection of Hemozoin in Clinical Samples
3.6. Detection of Other Relevant Malaria Biomarkers
3.7. Multi-Panel Biomarker Arrays for Malaria Detection
4. Conclusions and Future Perspective
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Jain, P.; Chakma, B.; Patra, S.; Goswami, P. Potential Biomarkers and Their Applications for Rapid and Reliable Detection of Malaria. Biomed Res. Int. 2014, 2014, 1–20. [Google Scholar] [CrossRef] [PubMed]
- WHO. The World Malaria Report 2018; World Health Organization: Geneva, Switzerland, 2018. [Google Scholar]
- Feachem, R.G.A.; Chen, I.; Akbari, O.; Bertozzi-Villa, A.; Bhatt, S.; Binka, F.; Boni, M.F.; Buckee, C.; Dieleman, J.; Dondorp, A.; et al. Malaria eradication within a generation: Ambitious, achievable, and necessary. Lancet 2019, 394, 1056–1112. [Google Scholar] [CrossRef]
- WHO. World Malaria Report 2019; World Health Organization: Geneva, Switzerland, 2019. [Google Scholar]
- White, N.J. Sulfadoxine-pyrimethamine for the treatment of malaria. Trans. R. Soc. Trop. Med. Hyg. 1991, 85, 556–557. [Google Scholar] [CrossRef]
- Scherf, A.; Lopez-Rubio, J.J.; Riviere, L. Antigenic Variation in Plasmodium falciparum. Annu. Rev. Microbiol. 2008, 62, 445–470. [Google Scholar] [CrossRef] [PubMed]
- Mueller, I.; Galinski, M.R.; Baird, J.K.; Carlton, J.M.; Kochar, D.K.; Alonso, P.L.; del Portillo, H.A. Key gaps in the knowledge of Plasmodium vivax, a neglected human malaria parasite. Lancet Infect. Dis. 2009, 9, 555–566. [Google Scholar] [CrossRef]
- Sinnis, P.; Willnow, T.E.; Briones, M.R.S.; Herz, J.; Nussenzweig, V. Remnant lipoproteins inhibit malaria sporozoite invasion of hepatocytes. J. Exp. Med. 1996, 184, 945–954. [Google Scholar] [CrossRef]
- Cerami, C.; Frevert, U.; Sinnis, P.; Takacs, B.; Clavijo, P.; Santos, M.J.; Nussenzweig, V. The basolateral domain of the hepatocyte plasma membrane bears receptors for the circumsporozoite protein of plasmodium falciparum sporozoites. Cell 1992, 70, 1021–1033. [Google Scholar] [CrossRef]
- Orito, Y.; Ishino, T.; Iwanaga, S.; Kaneko, I.; Kato, T.; Menard, R.; Chinzei, Y.; Yuda, M. Liver-specific protein 2: A Plasmodium protein exported to the hepatocyte cytoplasm and required for merozoite formation. Mol. Microbiol. 2013, 87, 66–79. [Google Scholar] [CrossRef]
- Gupta, D.K.; Dembele, L.; Voorberg-van der Wel, A.; Roma, G.; Yip, A.; Chuenchob, V.; Kangwanrangsan, N.; Ishino, T.; Vaughan, A.M.; Kappe, S.H.; et al. The Plasmodium liver-specific protein 2 (LISP2) is an early marker of liver stage development. Elife 2019, 8. [Google Scholar] [CrossRef]
- Nam, H.-W.; Song, K.J.; Ahn, H.J.; Yang, Z.; Chong, C.-K.; Cho, P.Y.; Ahn, S.K.; Kim, T.-S. Probability of Antibody Formation against Circumsporozoite Protein of Plasmodium vivax among Korean Malaria Patients. Korean J. Parasitol. 2014, 52, 143–149. [Google Scholar] [CrossRef]
- Reyburn, H. New WHO guidelines for the treatment of malaria. BMJ 2010, 340, c2637. [Google Scholar] [CrossRef] [PubMed]
- Amexo, M.; Tolhurst, R.; Barnish, G.; Bates, I. Malaria misdiagnosis: Effects on the poor and vulnerable. Lancet 2004, 364, 1896–1898. [Google Scholar] [CrossRef]
- Krampa, F.D.; Aniweh, Y.; Awandare, G.; Kanyong, P. Recent Progress in the Development of Diagnostic Tests for Malaria. Diagnostics 2017, 7, 54. [Google Scholar] [CrossRef] [PubMed]
- Britton, S.; Cheng, Q.; McCarthy, J.S. Novel molecular diagnostic tools for malaria elimination: A review of options from the point of view of high-throughput and applicability in resource limited settings. Malar. J. 2016, 15, 1–8. [Google Scholar] [CrossRef]
- Roth, J.M.; Korevaar, D.A.; Leeflang, M.M.G.; Mens, P.F. Molecular malaria diagnostics: A systematic review and meta-analysis. Crit. Rev. Clin. Lab. Sci. 2016, 53, 87–105. [Google Scholar] [CrossRef]
- Yanow, S.K. Molecular Diagnosis of Malaria in Low-Resource Settings. Point Care J. Near-Patient Test. Technol. 2016, 15, 41–42. [Google Scholar] [CrossRef]
- The malERA Consultative Group on Diagnoses A Research Agenda for Malaria Eradication: Diagnoses and Diagnostics. PLoS Med. 2011, 8, e1000396.
- Pologe, L.G.; Pavlovec, A.; Shio, H.; Ravetch, J.V. Primary structure and subcellular localization of the knob-associated histidine-rich protein of Plasmodium falciparum. Proc. Natl. Acad. Sci. USA 1987, 84, 7139–7143. [Google Scholar] [CrossRef]
- Rock, E.P.; Marsh, K.; Saul, A.J.; Wellems, T.E.; Taylor, D.W.; Maloy, W.L.; Howard, R.J. Comparative analysis of the Plasmodium falciparum histidine-rich proteins HRP-I, HRP-II and HRP-III in malaria parasites of diverse origin. Parasitology 1987, 95 Pt 2, 209–227. [Google Scholar] [CrossRef]
- Li, B.; Sun, Z.; Li, X.; Li, X.; Wang, H.; Chen, W.; Chen, P.; Qiao, M.; Mao, Y. Performance of pfHRP2 versus pLDH antigen rapid diagnostic tests for the detection of Plasmodium falciparum: A systematic review and meta-analysis. Arch. Med. Sci. 2017, 13, 541–549. [Google Scholar] [CrossRef]
- Ahmad, A.; Verma, A.K.; Krishna, S.; Sharma, A.; Singh, N.; Bharti, P.K. Plasmodium falciparum glutamate dehydrogenase is genetically conserved across eight malaria endemic states of India: Exploring new avenues of malaria elimination. PLoS ONE 2019, 14, e0218210. [Google Scholar] [CrossRef] [PubMed]
- Wilson, M.L. Malaria Rapid Diagnostic Tests. Clin. Infect. Dis. 2012, 54, 1637–1641. [Google Scholar] [CrossRef] [PubMed]
- Luppa, P.B.; Müller, C.; Schlichtiger, A.; Schlebusch, H. Point-of-care testing (POCT): Current techniques and future perspectives. TrAC Trends Anal. Chem. 2011, 30, 887–898. [Google Scholar] [CrossRef]
- Gubala, V.; Harris, L.F.; Ricco, A.J.; Tan, M.X.; Williams, D.E. Point of Care Diagnostics: Status and Future. Anal. Chem. 2012, 84, 487–515. [Google Scholar] [CrossRef] [PubMed]
- Derda, R.; Gitaka, J.; Klapperich, C.M.; Mace, C.R.; Kumar, A.A.; Lieberman, M.; Linnes, J.C.; Jores, J.; Nasimolo, J.; Ndung’u, J.; et al. Enabling the Development and Deployment of Next Generation Point-of-Care Diagnostics. PLoS Negl. Trop. Dis. 2015, 9, e0003676. [Google Scholar] [CrossRef]
- Hu, J.; Wang, S.; Wang, L.; Li, F.; Pingguan-Murphy, B.; Lu, T.J.; Xu, F. Advances in paper-based point-of-care diagnostics. Biosens. Bioelectron. 2014, 54, 585–597. [Google Scholar] [CrossRef]
- Posthuma-Trumpie, G.A.; Korf, J.; van Amerongen, A. Lateral flow (immuno)assay: its strengths, weaknesses, opportunities and threats. A literature survey. Anal. Bioanal. Chem. 2009, 393, 569–582. [Google Scholar] [CrossRef]
- Zhan, L.; Guo, S.Z.; Song, F.; Gong, Y.; Xu, F.; Boulware, D.R.; McAlpine, M.C.; Chan, W.C.W.; Bischof, J.C. The Role of Nanoparticle Design in Determining Analytical Performance of Lateral Flow Immunoassays. Nano Lett. 2017, 17, 7207–7212. [Google Scholar] [CrossRef]
- Le Roch, K.G. Global analysis of transcript and protein levels across the Plasmodium falciparum life cycle. Genome Res. 2004, 14, 2308–2318. [Google Scholar] [CrossRef]
- Barber, B.E.; William, T.; Grigg, M.J.; Piera, K.; Yeo, T.W.; Anstey, N.M. Evaluation of the Sensitivity of a pLDH-Based and an Aldolase-Based Rapid Diagnostic Test for Diagnosis of Uncomplicated and Severe Malaria Caused by PCR-Confirmed Plasmodium knowlesi, Plasmodium falciparum, and Plasmodium vivax. J. Clin. Microbiol. 2013, 51, 1118–1123. [Google Scholar] [CrossRef]
- Hendriksen, I.C.E.; Mtove, G.; Pedro, A.J.; Gomes, E.; Silamut, K.; Lee, S.J.; Mwambuli, A.; Gesase, S.; Reyburn, H.; Day, N.P.J.; et al. Evaluation of a PfHRP2 and a pLDH-based rapid diagnostic test for the diagnosis of severe malaria in 2 populations of african children. Clin. Infect. Dis. 2011, 52, 1100–1107. [Google Scholar] [CrossRef] [PubMed]
- Bell, D.; Wongsrichanalai, C.; Barnwell, J.W. Ensuring quality and access for malaria diagnosis: how can it be achieved? Nat. Rev. Microbiol. 2006, 4, 682–695. [Google Scholar] [CrossRef] [PubMed]
- Perkins, M.D.; Bell, D.R. Working without a blindfold: the critical role of diagnostics in malaria control. Malar. J. 2008, 7, S5. [Google Scholar] [CrossRef] [PubMed]
- Muzyka, K. Biosensors and Bioelectronics Current trends in the development of the electrochemiluminescent immunosensors. Biosens. Bioelectron. 2014, 54, 393–407. [Google Scholar] [CrossRef] [PubMed]
- Turner, A.P.F. Biosensors: sense and sensibility. Chem. Soc. Rev. 2013, 42, 3184. [Google Scholar] [CrossRef] [PubMed]
- Mascini, M. A Brief Story of Biosensor Technology. In Biotechnological Applications of Photosynthetic Proteins: Biochips, Biosensors and Biodevices; Springer US: Boston, MA, USA, 2007; ISBN 978-0-387-33009-9. [Google Scholar]
- Lee, T.M.H. Over-the-counter biosensors: Past, present, and future. Sensors 2008, 8, 5535–5559. [Google Scholar] [CrossRef]
- Belluzo, M.; Ribone, M.; Lagier, C. Assembling Amperometric Biosensors for Clinical Diagnostics. Sensors 2008, 8, 1366–1399. [Google Scholar] [CrossRef]
- Wang, J. Electrochemical glucose biosensors. In Electrochemical Sensors, Biosensors and their Biomedical Applications; Elsevier: Amsterdam, The Netherlands, 2008; pp. 57–69. ISBN 9780123737380. [Google Scholar]
- Bojdi, M.K.; Mashhadizadeh, M.H.; Behbahani, M.; Farahani, A.; Davarani, S.S.H.; Bagheri, A. Synthesis, characterization and application of novel lead imprinted polymer nanoparticles as a high selective electrochemical sensor for ultra-trace determination of lead ions in complex matrixes. Electrochim. Acta 2014, 136, 59–65. [Google Scholar] [CrossRef]
- Hosseini, H.; Behbahani, M.; Mahyari, M.; Kazerooni, H.; Bagheri, A.; Shaabani, A. Ordered carbohydrate-derived porous carbons immobilized gold nanoparticles as a new electrode material for electrocatalytical oxidation and determination of nicotinamide adenine dinucleotide. Biosens. Bioelectron. 2014, 59, 412–417. [Google Scholar] [CrossRef]
- Paul, B.; Panigrahi, A.K.; Singh, V.; Singh, S.G. A multi-walled carbon nanotube-zinc oxide nanofiber based flexible chemiresistive biosensor for malaria biomarker detection. Analyst 2017, 142, 2128–2135. [Google Scholar] [CrossRef]
- Solanki, P.R.; Kaushik, A.; Agrawal, V.V.; Malhotra, B.D. Nanostructured metal oxide-based biosensors. NPG Asia Mater. 2011, 3, 17–24. [Google Scholar] [CrossRef]
- Faridbod, F.; Gupta, V.K.; Zamani, H.A. Electrochemical Sensors and Biosensors. Int. J. Electrochem. 2011, 2011, 1–2. [Google Scholar] [CrossRef]
- Kumar, B.; Bhalla, V.; Singh Bhadoriya, R.P.; Suri, C.R.; Varshney, G.C. Label-free electrochemical detection of malaria-infected red blood cells. RSC Adv. 2016, 6, 75862–75869. [Google Scholar] [CrossRef]
- Ribaut, C.; Reybier, K.; Reynes, O.; Launay, J.; Valentin, A.; Fabre, P.L.; Nepveu, F. Electrochemical impedance spectroscopy to study physiological changes affecting the red blood cell after invasion by malaria parasites. Biosens. Bioelectron. 2009, 24, 2721–2725. [Google Scholar] [CrossRef]
- Singh, N.K.; Thungon, P.D.; Estrela, P.; Goswami, P. Development of an aptamer-based field effect transistor biosensor for quantitative detection of Plasmodium falciparum glutamate dehydrogenase in serum samples. Biosens. Bioelectron. 2019, 123, 30–35. [Google Scholar] [CrossRef]
- Noedl, H.; Wongsrichanalai, C.; Scott Miller, R.; Saw Aye Myint, K.; Looareesuwan, S.; Sukthana, Y.; Wongchotigul, V.; Kollaritsch, H.; Wiedermann, G.; Wernsdorfer, W.H. Plasmodium falciparum: effect of anti-malarial drugs on the production and secretion characteristics of histidine-rich protein II. Exp. Parasitol. 2002, 102, 157–163. [Google Scholar] [CrossRef]
- Koita, O.A.; Doumbo, O.K.; Ouattara, A.; Tall, L.K.; Konaré, A.; Diakité, M.; Diallo, M.; Sagara, I.; Masinde, G.L.; Doumbo, S.N.; et al. False-negative rapid diagnostic tests for malaria and deletion of the histidine-rich repeat region of the hrp2 gene. Am. J. Trop. Med. Hyg. 2012, 86, 194–198. [Google Scholar] [CrossRef]
- Gamboa, D.; Ho, M.-F.F.; Bendezu, J.; Torres, K.; Chiodini, P.L.; Barnwell, J.W.; Incardona, S.; Perkins, M.; Bell, D.; McCarthy, J.; et al. A large proportion of P. falciparum isolates in the Amazon region of Peru lack pfhrp2 and pfhrp3: Implications for malaria rapid diagnostic tests. PLoS ONE 2010, 5, e8091. [Google Scholar] [CrossRef]
- Bharti, P.K.; Chandel, H.S.; Ahmad, A.; Krishna, S.; Udhayakumar, V.; Singh, N. Prevalence of pfhrp2 and/or pfhrp3 gene deletion in plasmodium falciparum population in eight highly endemic states in India. PLoS ONE 2016, 11, e0157949. [Google Scholar] [CrossRef]
- Jeon, W.; Lee, S.; Dh, M.; Ban, C. A colorimetric aptasensor for the diagnosis of malaria based on cationic polymers and gold nanoparticles. Anal. Biochem. 2013, 439, 11–16. [Google Scholar] [CrossRef]
- Lee, S.; Song, K.M.; Jeon, W.; Jo, H.; Shim, Y.B.; Ban, C. A highly sensitive aptasensor towards Plasmodium lactate dehydrogenase for the diagnosis of malaria. Biosens. Bioelectron. 2012, 35, 291–296. [Google Scholar] [CrossRef] [PubMed]
- Jain, P.; Das, S.; Chakma, B.; Goswami, P. Aptamer-graphene oxide for highly sensitive dual electrochemical detection of Plasmodium lactate dehydrogenase. Anal. Biochem. 2016, 514, 32–37. [Google Scholar] [CrossRef] [PubMed]
- Brince Paul, K.; Kumar, S.; Tripathy, S.; Vanjari, S.R.K.; Singh, V.; Singh, S.G. A highly sensitive self assembled monolayer modified copper doped zinc oxide nanofiber interface for detection of Plasmodium falciparum histidine-rich protein-2: Targeted towards rapid, early diagnosis of malaria. Biosens. Bioelectron. 2016, 80, 39–46. [Google Scholar] [CrossRef] [PubMed]
- Gikunoo, E.; Abera, A.; Woldesenbet, E. A novel carbon Nanofibers grown on glass microballoons immunosensor: A tool for early diagnosis of Malaria. Sensors (Switzerland) 2014, 14, 14686–14699. [Google Scholar] [CrossRef] [PubMed]
- Singh, N.K.; Arya, S.K.; Estrela, P.; Goswami, P. Capacitive malaria aptasensor using Plasmodium falciparum glutamate dehydrogenase as target antigen in undiluted human serum. Biosens. Bioelectron. 2018, 117, 246–252. [Google Scholar] [CrossRef] [PubMed]
- Hemben, A.; Ashley, J.; Tothill, I. Development of an Immunosensor for PfHRP 2 as a Biomarker for Malaria Detection. Biosensors 2017, 7, 28. [Google Scholar] [CrossRef]
- Hemben, A.; Ashley, J.; Tothill, I.E. An immunosensor for parasite lactate dehydrogenase detection as a malaria biomarker – Comparison with commercial test kit. Talanta 2018, 187, 321–329. [Google Scholar] [CrossRef]
- Chakma, B.; Jain, P.; Singh, N.K.; Goswami, P. Development of an Indicator Displacement Based Detection of Malaria Targeting HRP-II as Biomarker for Application in Point-of-Care Settings. Anal. Chem. 2016, 88, 10316–10321. [Google Scholar] [CrossRef]
- Dirkzwager, R.M.; Liang, S.; Tanner, J.A. Development of Aptamer-Based Point-of-Care Diagnostic Devices for Malaria Using Three-Dimensional Printing Rapid Prototyping. ACS Sensors 2016, 1, 420–426. [Google Scholar] [CrossRef]
- Fraser, L.A.; Kinghorn, A.B.; Dirkzwager, R.M.; Liang, S.; Cheung, Y.-W.; Lim, B.; Shiu, S.C.-C.; Tang, M.S.L.; Andrew, D.; Manitta, J.; et al. A portable microfluidic Aptamer-Tethered Enzyme Capture (APTEC) biosensor for malaria diagnosis. Biosens. Bioelectron. 2018, 100, 591–596. [Google Scholar] [CrossRef]
- Sharma, M.K.; Rao, V.K.; Agarwal, G.S.; Rai, G.P.; Gopalan, N.; Prakash, S.; Sharma, S.K.; Vijayaraghavan, R. Highly sensitive amperometric immunosensor for detection of plasmodium falciparum histidine-rich protein 2 in serum of humans with malaria: Comparison with a commercial kit. J. Clin. Microbiol. 2008, 46, 3759–3765. [Google Scholar] [CrossRef] [PubMed]
- Kenry; Geldert, A.; Zhang, X.; Zhang, H.; Lim, C.T. Highly Sensitive and Selective Aptamer-Based Fluorescence Detection of a Malarial Biomarker Using Single-Layer MoS 2 Nanosheets. ACS Sensors 2016, 1, 1315–1321. [Google Scholar] [CrossRef]
- De Souza Castilho, M.; Laube, T.; Yamanaka, H.; Alegret, S.; Pividori, M.I. Magneto immunoassays for plasmodium falciparum histidine-rich protein 2 related to malaria based on magnetic nanoparticles. Anal. Chem. 2011, 83, 5570–5577. [Google Scholar] [CrossRef] [PubMed]
- Sikarwar, B.; Sharma, P.K.; Srivastava, A.; Agarwal, G.S.; Boopathi, M.; Singh, B.; Jaiswal, Y.K. Surface plasmon resonance characterization of monoclonal and polyclonal antibodies of malaria for biosensor applications. Biosens. Bioelectron. 2014, 60, 201–209. [Google Scholar] [CrossRef]
- Potipitak, T.; Ngrenngarmlert, W.; Promptmas, C.; Chomean, S.; Ittarat, W. Diagnosis and genotyping of Plasmodium falciparum by a DNA biosensor based on quartz crystal microbalance (QCM). Clin. Chem. Lab. Med. 2011, 49, 1367–1373. [Google Scholar] [CrossRef]
- Hede, M.S.; Okorie, P.N.; Fruekilde, S.K.; Fjelstrup, S.; Thomsen, J.; Franch, O.; Tesauro, C.; Bugge, M.T.; Christiansen, M.; Picot, S.; et al. Refined method for droplet microfluidics-enabled detection of Plasmodium falciparum encoded topoisomerase i in blood from malaria patients. Micromachines 2015, 6, 1505–1513. [Google Scholar] [CrossRef]
- Ngo, H.T.; Gandra, N.; Fales, A.M.; Taylor, S.M.; Vo-Dinh, T. Sensitive DNA detection and SNP discrimination using ultrabright SERS nanorattles and magnetic beads for malaria diagnostics. Biosens. Bioelectron. 2016, 81, 8–14. [Google Scholar] [CrossRef]
- Wangmaung, N.; Chomean, S.; Promptmas, C.; Mas-oodi, S.; Tanyong, D.; Ittarat, W. Silver quartz crystal microbalance for differential diagnosis of Plasmodium falciparum and Plasmodium vivax in single and mixed infection. Biosens. Bioelectron. 2014, 62, 295–301. [Google Scholar] [CrossRef]
- Kumar, B.; Bhalla, V.; Suri, C.R.; Varshney, G.C.; Kumar, B.; Bhalla, V.; Singh Bhadoriya, R.P.; Suri, C.R.; Varshney, G.C.; Looareesuwan, S.; et al. Label-free electrochemical detection of malaria-infected red blood cells. RSC Adv. 2016, 6, 1–5. [Google Scholar] [CrossRef]
- Kong, T.F.; Ye, W.; Peng, W.K.; Hou, H.W.; Marcos; Preiser, P.R.; Nguyen, N.T.; Han, J. Enhancing malaria diagnosis through microfluidic cell enrichment and magnetic resonance relaxometry detection. Sci. Rep. 2015, 5, 1–12. [Google Scholar]
- Rodriguez-del Valle, M.; Quakyi, I.A.; Amuesi, J.; Quaye, J.T.; Nkrumah, F.K.; Taylor, D.W. Detection of antigens and antibodies in the urine of humans with Plasmodium falciparum malaria. J. Clin. Microbiol. 1991, 29, 1236–1242. [Google Scholar] [CrossRef] [PubMed]
- Parra, M.E.; Evans, C.B.; Taylor, D.W. Identification of Plasmodium falciparum histidine-rich protein 2 in the plasma of humans with malaria. J. Clin. Microbiol. 1991, 29, 1629–1634. [Google Scholar] [CrossRef] [PubMed]
- Sin, M.L.Y.; Mach, K.E.; Wong, P.K.; Liao, J.C. Advances and challenges in biosensor-based diagnosis of infectious diseases. Expert Rev. Mol. Diagn. 2014, 14, 225–244. [Google Scholar] [CrossRef] [PubMed]
- Castro-Sesquen, Y.E.; Kim, C.; Gilman, R.H.; Sullivan, D.J.; Searson, P.C. Nanoparticle-Based Histidine-Rich Protein-2 Assay for the Detection of the Malaria Parasite Plasmodium falciparum. Am. J. Trop. Med. Hyg. 2016, 95, 354–357. [Google Scholar] [CrossRef] [PubMed]
- Cao, X.; Ye, Y.; Liu, S. Gold nanoparticle-based signal amplification for biosensing. Anal. Biochem. 2011, 417, 1–16. [Google Scholar] [CrossRef] [PubMed]
- Lei, J.; Ju, H. Signal amplification using functional nanomaterials for biosensing. Chem. Soc. Rev. 2012, 41, 2122. [Google Scholar] [CrossRef]
- Ju, H.; Zhang, X.; Wang, J. Signal Amplification for Nanobiosensing. In Biological and Medical Physics, Biomedical Engineering; Springer: New York, NY, USA, 2011. [Google Scholar]
- Sharma, M.K.; Agarwal, G.S.; Rao, V.K.; Upadhyay, S.; Merwyn, S.; Gopalan, N.; Rai, G.P.; Vijayaraghavan, R.; Prakash, S. Amperometric immunosensor based on gold nanoparticles/alumina sol-gel modified screen-printed electrodes for antibodies to Plasmodium falciparum histidine rich protein-2. Analyst 2010, 135, 608–614. [Google Scholar] [CrossRef]
- Ravaoarisoa, E.; Zamanka, H.; Fusai, T.; Bellalou, J.; Bedouelle, H.; Mercereau-Puijalon, O.; Fandeur, T. Recombinant antibodies specific for the Plasmodium falciparum histidine-rich protein 2. MAbs 2010, 2, 416–427. [Google Scholar] [CrossRef]
- Sharma, M.K.; Rao, V.K.; Merwyn, S.; Agarwal, G.S.; Upadhyay, S.; Vijayaraghavan, R. A novel piezoelectric immunosensor for the detection of malarial Plasmodium falciparum histidine rich protein-2 antigen. Talanta 2011, 85, 1812–1817. [Google Scholar] [CrossRef]
- Prodromidis, M.I. Impedimetric immunosensors—A review. Electrochim. Acta 2010, 55, 4227–4233. [Google Scholar] [CrossRef]
- Piper, R.; LeBras, J.; Wentworth, L.; Hunt-Cooke, A.; Houzé, S.; Chiodini, P.; Makler, M. Immunocapture diagnostic assays for malaria using Plasmodium lactate dehydrogenase (pLDH). Am. J. Trop. Med. Hyg. 1999, 60, 109–118. [Google Scholar] [CrossRef] [PubMed]
- Palmer, C.J.; Lindo, J.F.; Klaskala, W.I.; Quesada, J.A.; Kaminsky, R.; Baum, M.K.; Ager, A.L. Evaluation of the optimal test for rapid diagnosis of Plasmodium vivax and Plasmodium falciparurn malaria. J. Clin. Microbiol. 1998, 36, 203–206. [Google Scholar] [CrossRef]
- Makler, M.T.; Palmer, C.J.; Ager, A.L. A review of practical techniques for the diagnosis of malaria. Ann. Trop. Med. Parasitol. 1998, 92, 419–433. [Google Scholar] [PubMed]
- Figueroa-Miranda, G.; Feng, L.; Shiu, S.C.C.; Dirkzwager, R.M.; Cheung, Y.W.; Tanner, J.A.; Schöning, M.J.; Offenhäusser, A.; Mayer, D. Aptamer-based electrochemical biosensor for highly sensitive and selective malaria detection with adjustable dynamic response range and reusability. Sensors Actuators, B Chem. 2018, 255, 235–243. [Google Scholar] [CrossRef]
- Cohen, J.; Dupas, P.; Schaner, S. Price subsidies, diagnostic tests, and targeting of malaria treatment: Evidence from a randomized controlled trial. Am. Econ. Rev. 2015, 105, 609–645. [Google Scholar] [CrossRef]
- BRITTON, K.L.; BAKER, P.J.; RICE, D.W.; STILLMAN, T.J. Structural relationship between the hexameric and tetrameric family of glutamate dehydrogenases. Eur. J. Biochem. 1992, 209, 851–859. [Google Scholar] [CrossRef]
- Jagt, D.L.V.; Intress, C.; Heidrich, J.E.; Mrema, J.E.K.; Rieckmann, K.H.; Heidrich, H.-G. Marker Enzymes of Plasmodium falciparum and Human Erythrocytes as Indicators of Parasite Purity. J. Parasitol. 1982, 68, 1068. [Google Scholar] [CrossRef]
- Zocher, K.; Fritz-Wolf, K.; Kehr, S.; Fischer, M.; Rahlfs, S.; Becker, K. Biochemical and structural characterization of Plasmodium falciparum glutamate dehydrogenase 2. Mol. Biochem. Parasitol. 2012, 183, 52–62. [Google Scholar] [CrossRef]
- Li, Y.; Ning, Y.S.; Li, L.; Peng, D.D.; Dong, W.Q.; Li, M. Preparation of monoclonal antibodies against Plasmodium falciparum glutamate dehydrogenase and establishment of colloidal goldimmunochromatographic assay. Di Yi Jun Yi Da Xue Xue Bao 2005, 25, 435–438. [Google Scholar]
- Wagner, J.T.; Lüdemann, H.; Färber, P.M.; Lottspeich, F.; Krauth-Siegel, R.L. Glutamate dehydrogenase, the marker protein of Plasmodium falciparum cloning, expression and characterization of the malarial enzyme. Eur. J. Biochem. 1998, 258, 813–819. [Google Scholar] [CrossRef]
- Park, S.J.; Kwon, O.S.; Lee, S.H.; Song, H.S.; Park, T.H.; Jang, J. Ultrasensitive Flexible Graphene Based Field-Effect Transistor (FET)-Type Bioelectronic Nose. Nano Lett. 2012, 12, 5082–5090. [Google Scholar] [CrossRef] [PubMed]
- Srivastava, I.K.; Schmidt, M.; Certa, U.; Döbeli, H.; Perrin, L.H. Specificity and inhibitory activity of antibodies to Plasmodium falciparum aldolase. J. Immunol. 1990, 144, 1497–1503. [Google Scholar] [PubMed]
- Dzakah, E.E.; Kang, K.; Ni, C.; Tang, S.; Wang, J.; Wang, J. Comparative performance of aldolase and lactate dehydrogenase rapid diagnostic tests in Plasmodium vivax detection. Malar. J. 2014, 13, 272. [Google Scholar] [CrossRef]
- Lee, N.; Baker, J.; Bell, D.; McCarthy, J.; Cheng, Q. Assessing the genetic diversity of the aldolase genes of Plasmodium falciparum and Plasmodium vivax and its potential effect on performance of aldolase-detecting rapid diagnostic tests. J. Clin. Microbiol. 2006, 44, 4547–4549. [Google Scholar] [CrossRef] [PubMed]
- Chugh, M.; Sundararaman, V.; Kumar, S.; Reddy, V.S.; Siddiqui, W.A.; Stuart, K.D.; Malhotra, P. Protein complex directs hemoglobin-to-hemozoin formation in Plasmodium falciparum. Proc. Natl. Acad. Sci. USA 2013, 110, 5392–5397. [Google Scholar] [CrossRef]
- Pagola, S.; Stephens, P.W.; Bohle, D.S.; Kosar, A.D.; Madsen, S.K. The structure of malaria pigment β-haematin. Nature 2000, 404, 307–310. [Google Scholar] [CrossRef]
- Chen, K.; Yuen, C.; Aniweh, Y.; Preiser, P.; Liu, Q. Towards ultrasensitive malaria diagnosis using surface enhanced Raman spectroscopy. Sci. Rep. 2016, 6, 20177. [Google Scholar] [CrossRef]
- Guirgis, B.S.S.; Sá E Cunha, C.; Gomes, I.; Cavadas, M.; Silva, I.; Doria, G.; Blatch, G.L.; Baptista, P.V.; Pereira, E.; Azzazy, H.M.E.; et al. Gold nanoparticle-based fluorescence immunoassay for malaria antigen detection. Anal. Bioanal. Chem. 2012, 402, 1019–1027. [Google Scholar] [CrossRef]
- Kattenberg, J.H.; Versteeg, I.; Migchelsen, S.J.; González, I.J.; Perkins, M.D.; Mens, P.F.; Schallig, H.D.F.H. New developments in malaria diagnostics: Monoclonal antibodies against plasmodium dihydrofolate reductase-thymidylate synthase, heme detoxification protein and glutamate rich protein. MAbs 2012, 4, 120–126. [Google Scholar] [CrossRef][Green Version]
- Mouatcho, J.C.; Dean Goldring, J.P.; Mouatcho, J.C. and Goldring, J.P. Malaria rapid diagnostic tests: Challenges and prospects. J. Med. Microbiol. 2013, 62, 1491–1505. [Google Scholar] [CrossRef]
- Garcia, L.S. Diagnostic Medical Parasitology, 5th ed.; ASM Press: Washington, DC, USA, 2007; pp. 130–140. [Google Scholar]
- Birch, C.M.; Hou, H.W.; Han, J.; Niles, J.C. Identification of malaria parasite-infected red blood cell surface aptamers by inertial microfluidic SELEX (I-SELEX). Sci. Rep. 2015, 5, 11347. [Google Scholar] [CrossRef] [PubMed]
- Cardoso, A.R.; Cabral-Miranda, G.; Reyes-Sandoval, A.; Bachmann, M.F.; Sales, M.G.F. Detecting circulating antibodies by controlled surface modification with specific target proteins: Application to malaria. Biosens. Bioelectron. 2017, 91, 833–841. [Google Scholar] [CrossRef] [PubMed]
- Cordray, M.S.; Richards-Kortum, R.R. Review: Emerging nucleic acid-based tests for point-of-care detection of malaria. Am. J. Trop. Med. Hyg. 2012, 87, 223–230. [Google Scholar] [CrossRef] [PubMed]
- Ittarat, W.; Chomean, S.; Sanchomphu, C.; Wangmaung, N.; Promptmas, C.; Ngrenngarmlert, W. Biosensor as a molecular malaria differential diagnosis. Clin. Chim. Acta 2013, 419, 47–51. [Google Scholar] [CrossRef] [PubMed]
- Nash, M.A.; Waitumbi, J.N.; Hoffman, A.S.; Yager, P.; Stayton, P.S. Multiplexed enrichment and detection of malarial biomarkers using a stimuli-responsive iron oxide and gold nanoparticle reagent system. ACS Nano 2012, 6, 6776–6785. [Google Scholar] [CrossRef] [PubMed]
- Markwalter, C.F.; Ricks, K.M.; Bitting, A.L.; Mudenda, L.; Wright, D.W. Simultaneous capture and sequential detection of two malarial biomarkers on magnetic microparticles. Talanta 2016, 161, 443–449. [Google Scholar] [CrossRef]
- Jepsen, M.P.G.; Röser, D.; Christiansen, M.; Larsen, S.O.; Cavanagh, D.R.; Dhanasarnsombut, K.; Bygbjerg, I.; Dodoo, D.; Remarque, E.J.; Dziegiel, M.; et al. Development and evaluation of a multiplex screening assay for Plasmodium falciparum exposure. J. Immunol. Methods 2012, 384, 62–70. [Google Scholar] [CrossRef]
- Iqbal, J.; Siddique, A.; Jameel, M.; Hira, P.R. Persistent histidine-rich protein 2, parasite lactate dehydrogenase, and panmalarial antigen reactivity after clearance of Plasmodium falciparum monoinfection. J. Clin. Microbiol. 2004, 42, 4237–4241. [Google Scholar] [CrossRef]
- Lafleur, L.; Stevens, D.; McKenzie, K.; Ramachandran, S.; Spicar-Mihalic, P.; Singhal, M.; Arjyal, A.; Osborn, J.; Kauffman, P.; Yager, P.; et al. Progress toward multiplexed sample-to-result detection in low resource settings using microfluidic immunoassay cards. Lab Chip 2012, 12, 1119–1127. [Google Scholar] [CrossRef]
- Deraney, R.N.; Mace, C.R.; Rolland, J.P.; Schonhorn, J.E. Multiplexed, Patterned-Paper Immunoassay for Detection of Malaria and Dengue Fever. Anal. Chem. 2016, 88, 6161–6165. [Google Scholar] [CrossRef]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Analytes | Sensing Technique/Response | Transducer | Biomarker | Receptor Molecule | LoD | Range | Response Time | Storage Stability | References |
|---|---|---|---|---|---|---|---|---|---|
| Antigens | Colorimetric | - | pLDH (PvLDH, PfLDH) | pL1 aptamer | 8.3–8.7 pM (PvLDH) 10.3–12.5 pM (PfLDH) | NA | NA | NA | [54] |
| EIS | Gold electrode | pLDH | pL1 aptamer | ** 108.5 fM for PvLDH ** 120.1 fM for PfLDH | NA | NA | NA | [55] | |
| EIS | GCE | pLDH | P38 aptamer (90 mer ssDNA) | 0.5 fM | NA | - | NA | [56] | |
| EIS | GCE | HRP-2 | Anti-HRP-2 antibody | ** 6.8 ag/mL. | 10 ag/mL–10 mg/mL | NA | 2 months (86.5%) | [57] | |
| Chemiresistive (electrical conductance) | - | PfHRP-2 | Anti-HRP-2 antibody | 0.97 fg/mL | 10 fg/mL–10 ng/mL | NA | 15 days (94.2%) | [44] | |
| - | - | PfHRP-2 | Anti-PfHRP-2 | 0.025 ng/mL | 0.01–10 ng/mL | - | - | [58] | |
| EIS | Gold disc electrodes | PfGDH | ssDNA aptamer (NG3) | * 0.77 pM | 100 fM–100 nM | NA | NA | [59] | |
| Potentiometric (FET) | Gold micro-electrodes | PfGDH | ssDNA aptamer (NG3) | ** 16.7 pM * 48.6 pM | 100 fM–10 nM | 5 s | [49] | ||
| Amperometric | Gold-SPE | PfHRP-2 | Anti-PfHRP 2 mAb | ** 36 pg/mL * 40 pg/mL | NA | NA | NA | [60] | |
| Amperometric | Gold-SPE | pLDH | pLDH capture antibody | ** 19 pg/mL * 23 pg/mL | - | - | - | [61] | |
| Spectrophotometric Indicator displacement medium | - | PfHRP-2 | NA | 30 ± 9.6 nM | 10–100 nM | 5 min | NA | [62] | |
| Colorimetric | - | PfLDH | 2008s-biotin DNA aptamer | ** 4.9 ng/mL | NA | <1h | 2 months | [63] | |
| Colorimetric | - | PfLDH | 2008s aptamer | - | NA | 20 min | - | [64] | |
| Amperometric | SPE | PfHRP-2 | Mouse anti-PfHRP-2 antibody | ** 8 ng/mL | NA | NA | NA | [65] | |
| FRET | - | pLDH | Fluorescently-labeled aptamer (36 mer ssDNA) | ** 550 pM | NA | NA | NA | [66] | |
| Amperometric magneto Immunosensor | - | PfHRP2 | Anti-HRP2 IgM Antibody | 0.36 ng/mL | 0.35–7.8 ng/mL | NA | NA | [67] | |
| Antibodies | SPR | Gold disc | Antibodies of Pf. | PfHRP2 | ** 5.6 pg for mAb | - | NA | NA | [68] |
| Nucleic acids | Quartz Crystal Microbalance | - | Pf msp2 gene | Biotinylated probe | ≥0.025 ng/mL of target DNA | NA | NA | 180 days | [69] |
| Droplet Microfluidic Platform | - | Pf topoisomerase I | ds DNA substrate | NA | NA | NA | NA | [70] | |
| SERS Nanoplatform | - | Pf DNA sequences | Magnetic bead and nanorattle | 100 attomoles | 10−11–10−10 M | NA | NA | [71] | |
| Quartz Crystal Microbalance | Silver electrode | 18s rRNA gene (Pf and Pv) | immobilized probe | - | - | [72] | |||
| Infected red blood cells | EIS | SPE | Pf infected RBCs | monoclonal antibody | - | 102–107 cells/mL | NA | NA | [73] |
| microfluidic separation and MRR | - | Infected RBCs | - | 0.0005% parasitemia | - | - | - | [74] |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Krampa, F.D.; Aniweh, Y.; Kanyong, P.; Awandare, G.A. Recent Advances in the Development of Biosensors for Malaria Diagnosis. Sensors 2020, 20, 799. https://doi.org/10.3390/s20030799
Krampa FD, Aniweh Y, Kanyong P, Awandare GA. Recent Advances in the Development of Biosensors for Malaria Diagnosis. Sensors. 2020; 20(3):799. https://doi.org/10.3390/s20030799
Chicago/Turabian StyleKrampa, Francis D., Yaw Aniweh, Prosper Kanyong, and Gordon A. Awandare. 2020. "Recent Advances in the Development of Biosensors for Malaria Diagnosis" Sensors 20, no. 3: 799. https://doi.org/10.3390/s20030799
APA StyleKrampa, F. D., Aniweh, Y., Kanyong, P., & Awandare, G. A. (2020). Recent Advances in the Development of Biosensors for Malaria Diagnosis. Sensors, 20(3), 799. https://doi.org/10.3390/s20030799