
Influence of Culture Media on Microbial Fingerprints Using Raman Spectroscopy

 , ,
, ,
Abstract
:1. Introduction
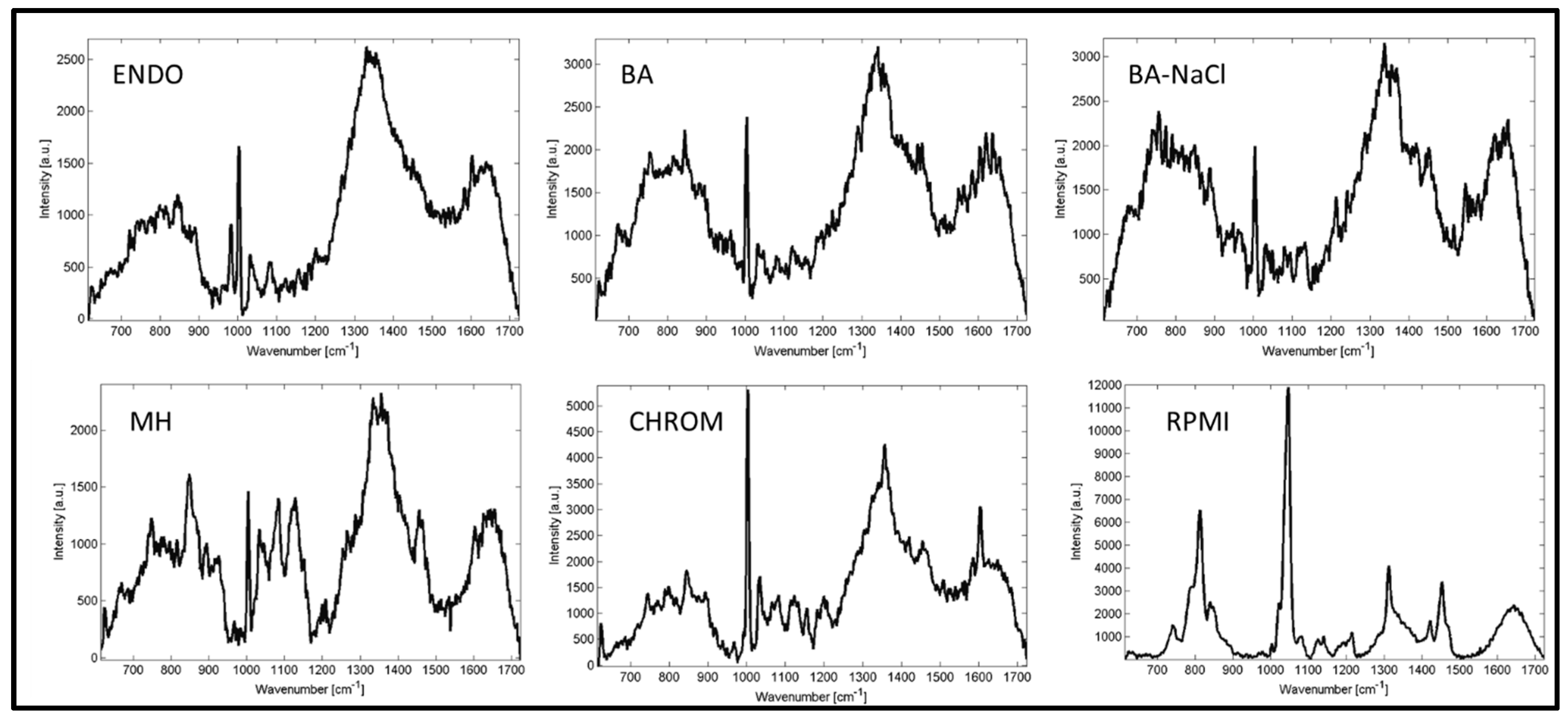
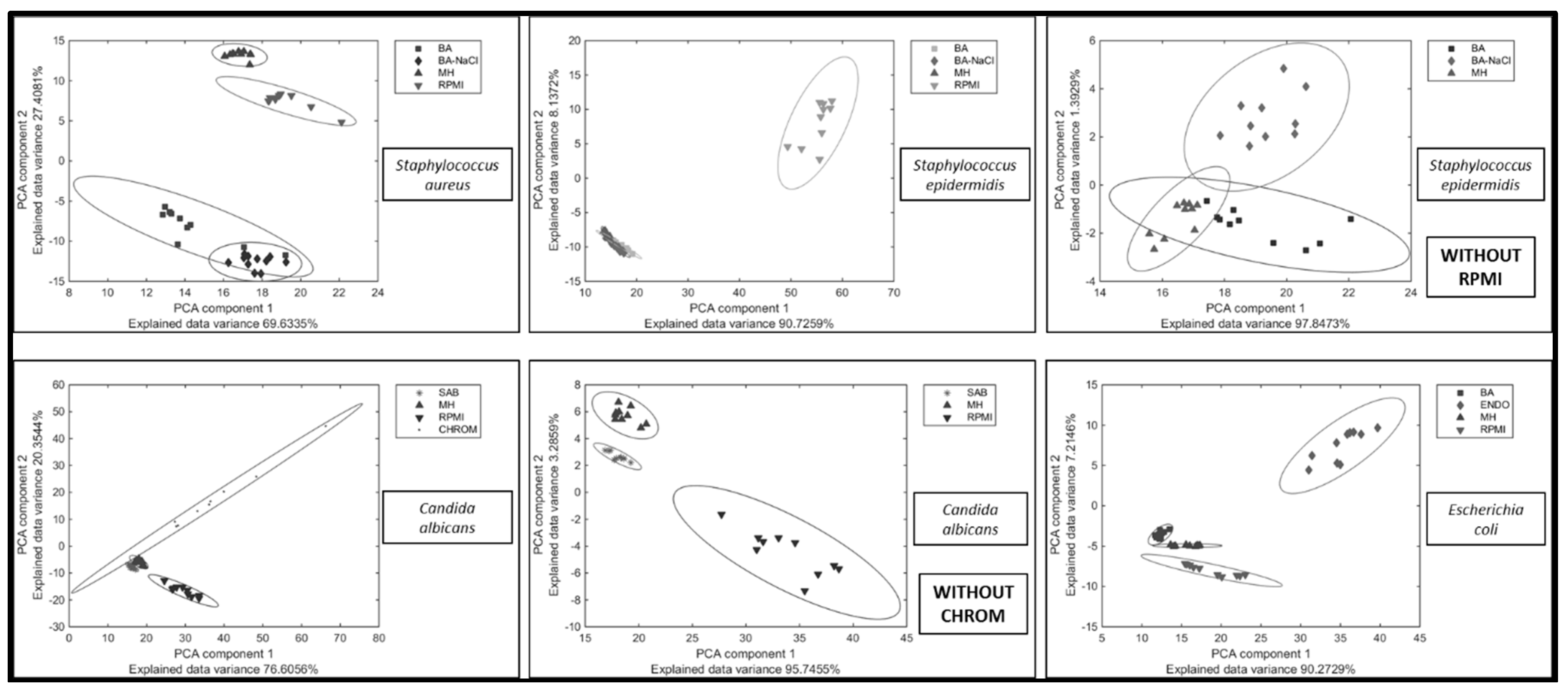
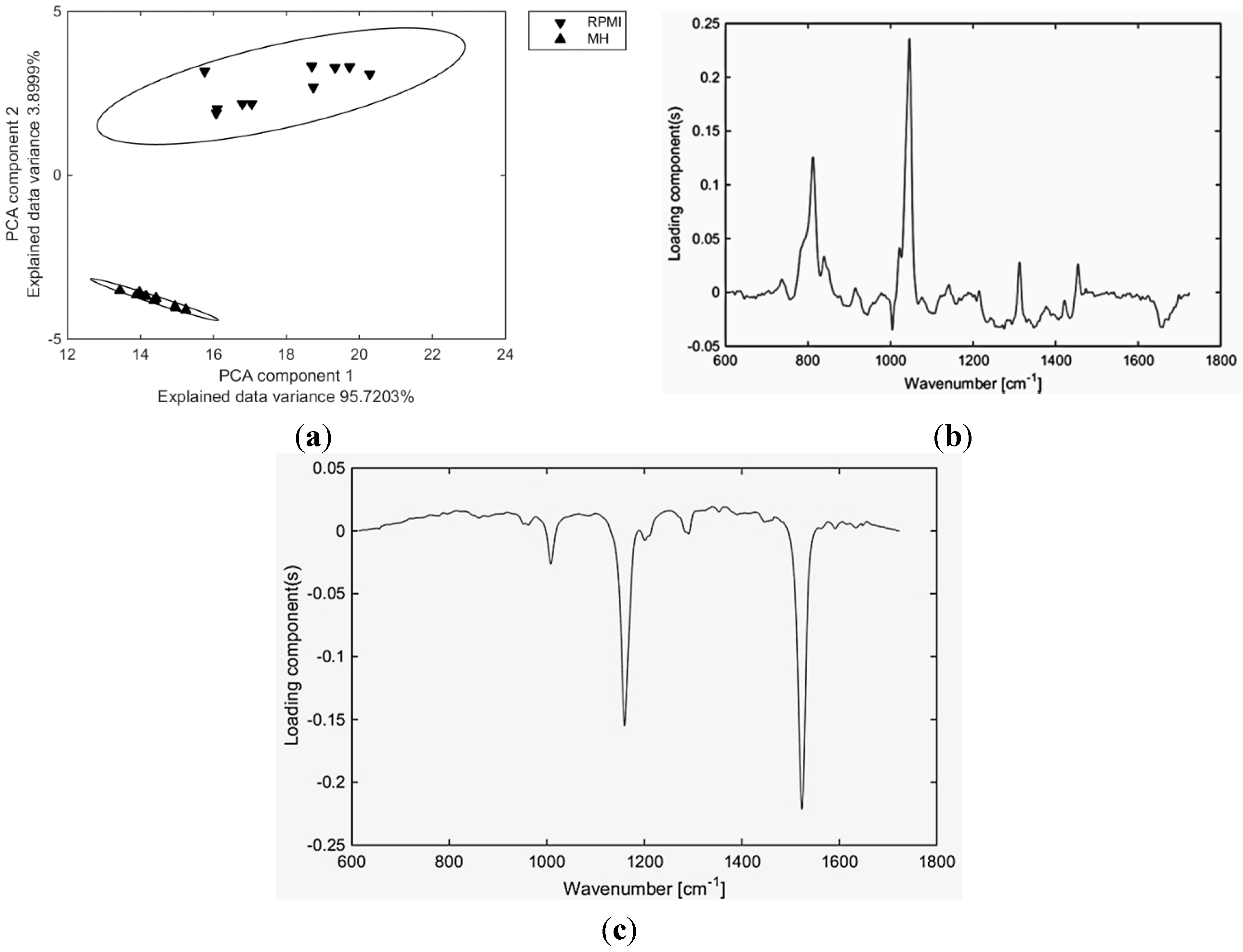
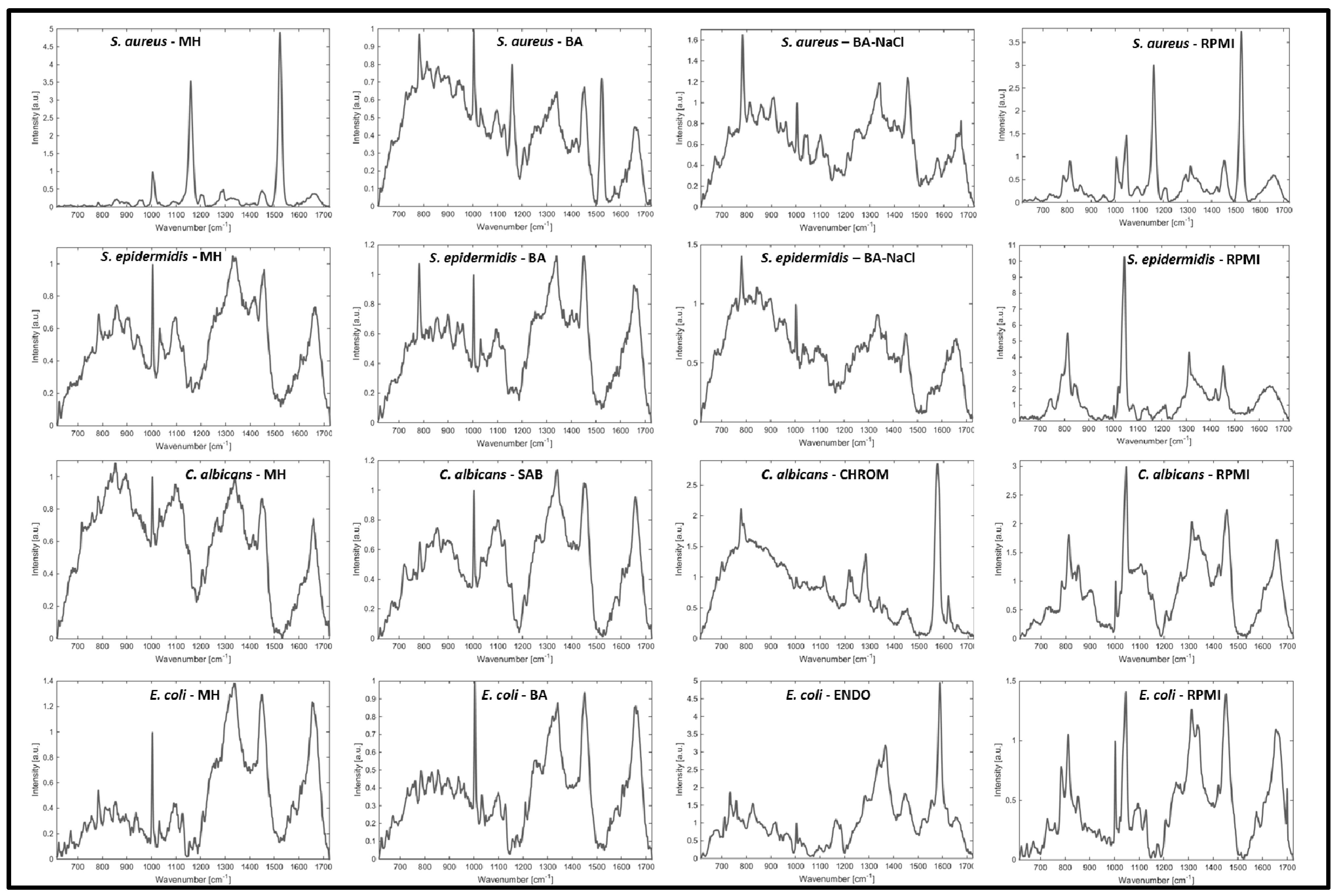
2. Results and Discussion
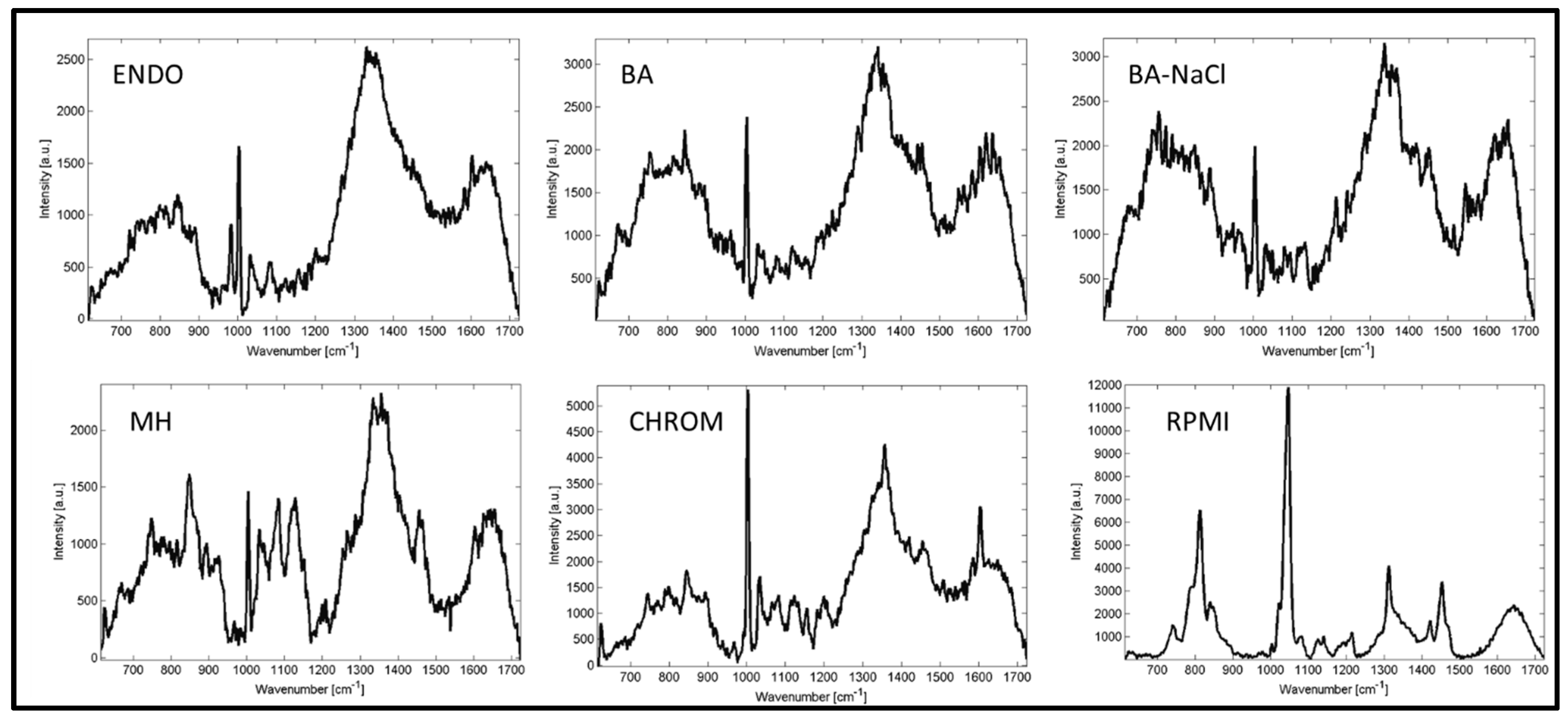
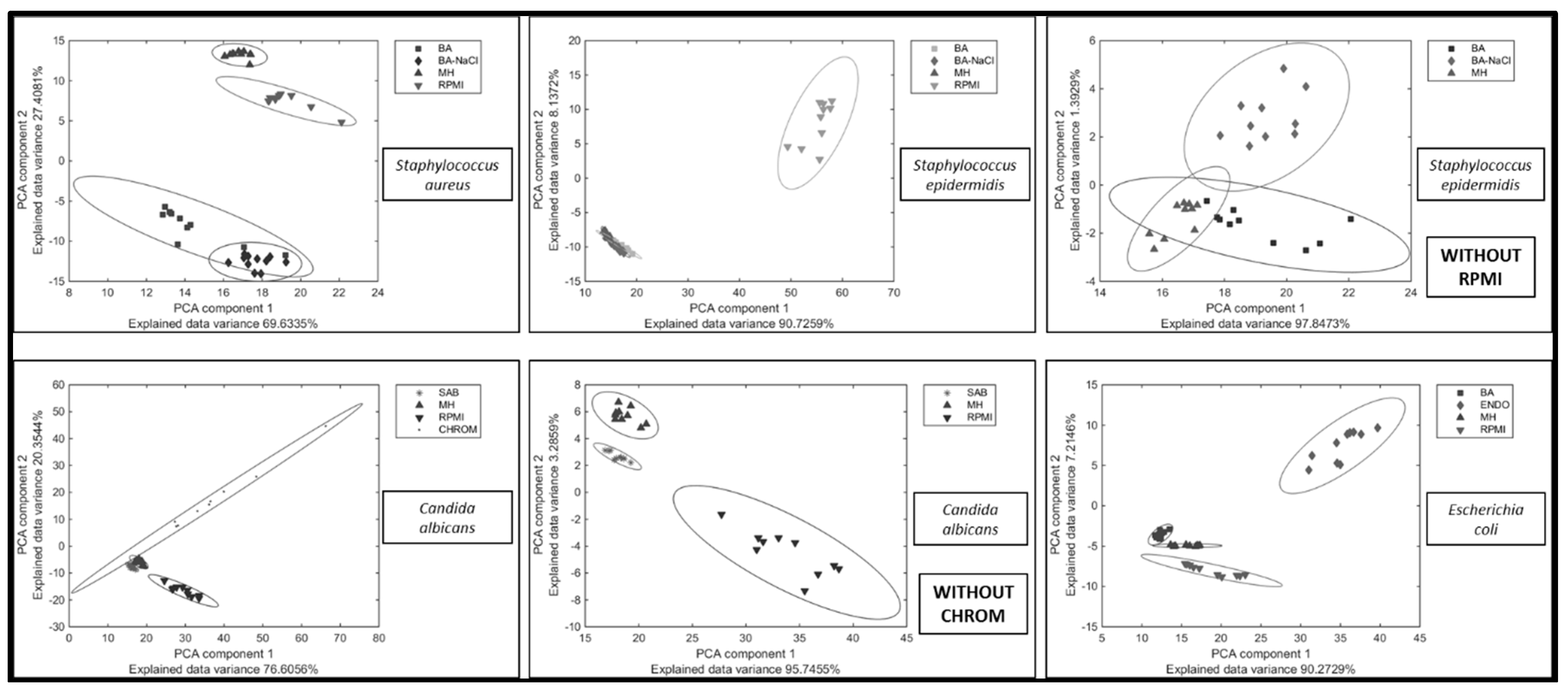
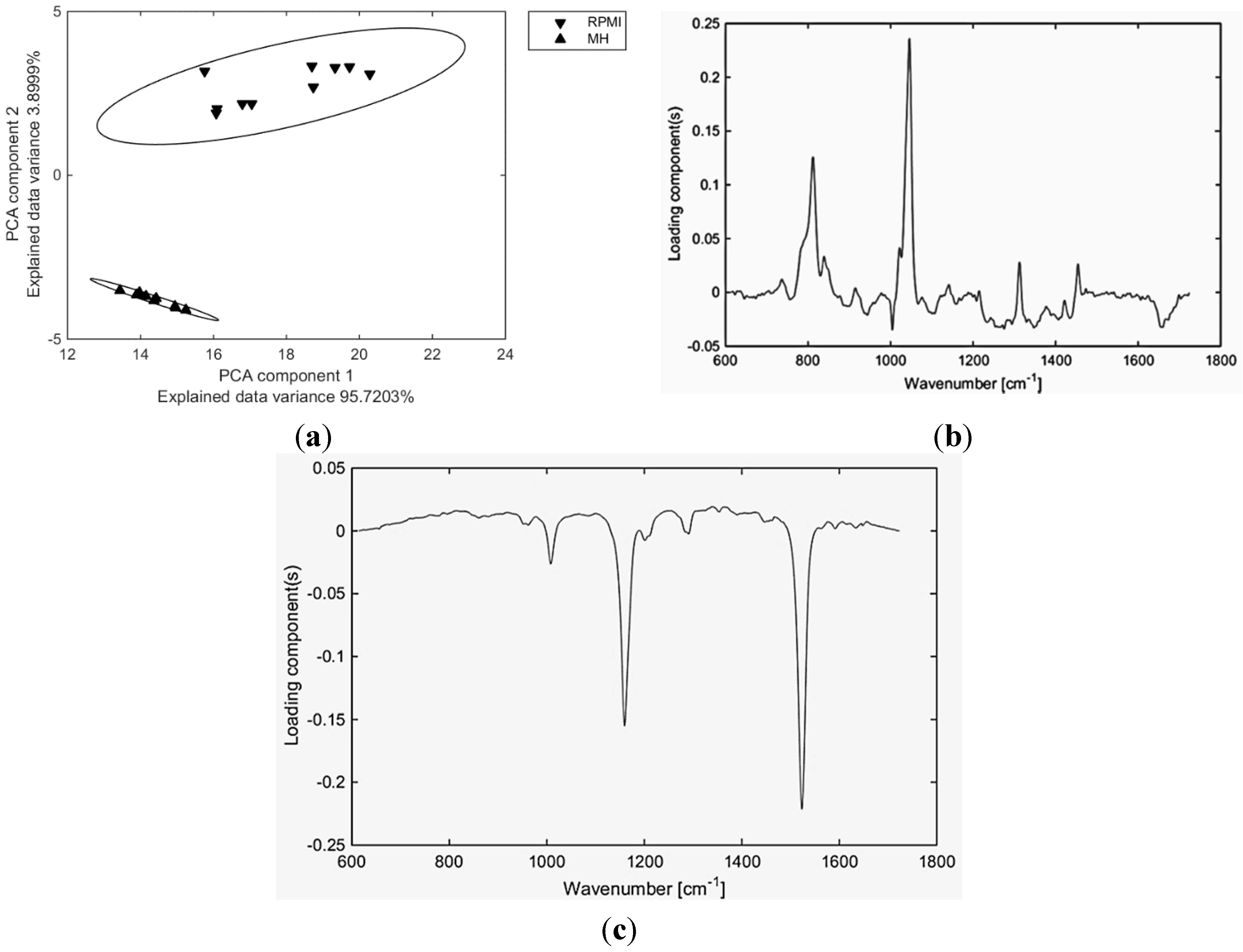
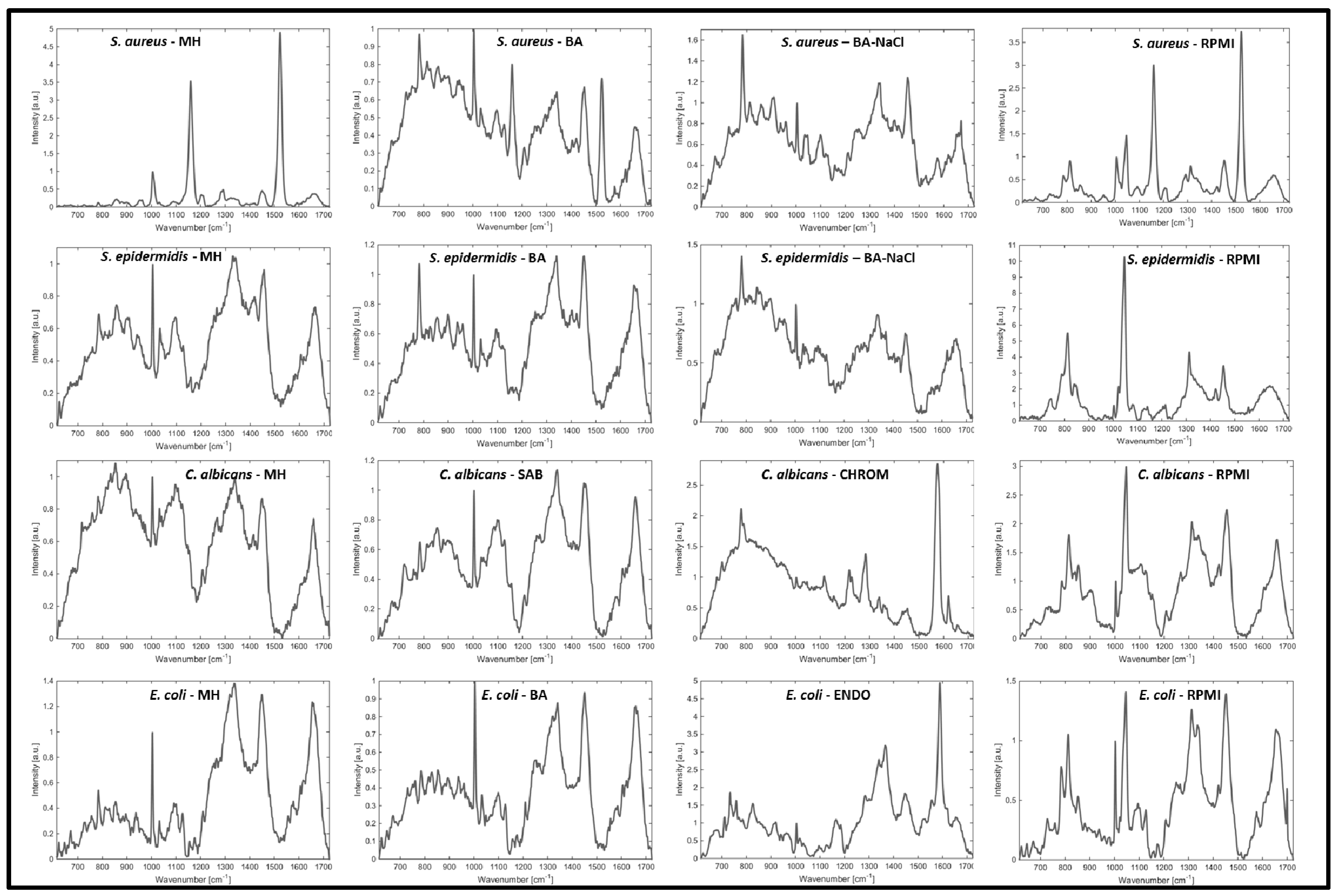
3. Materials and Methods
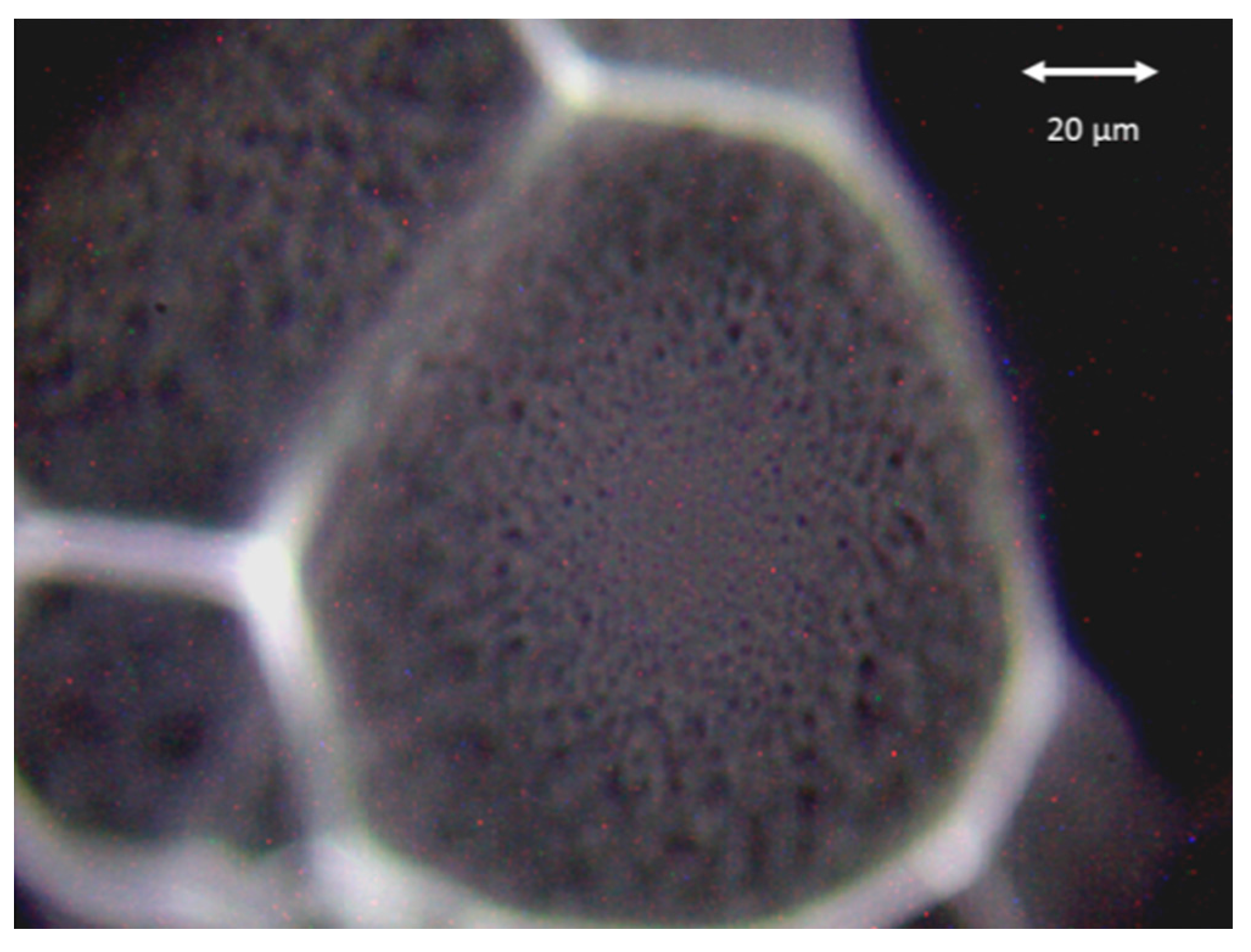
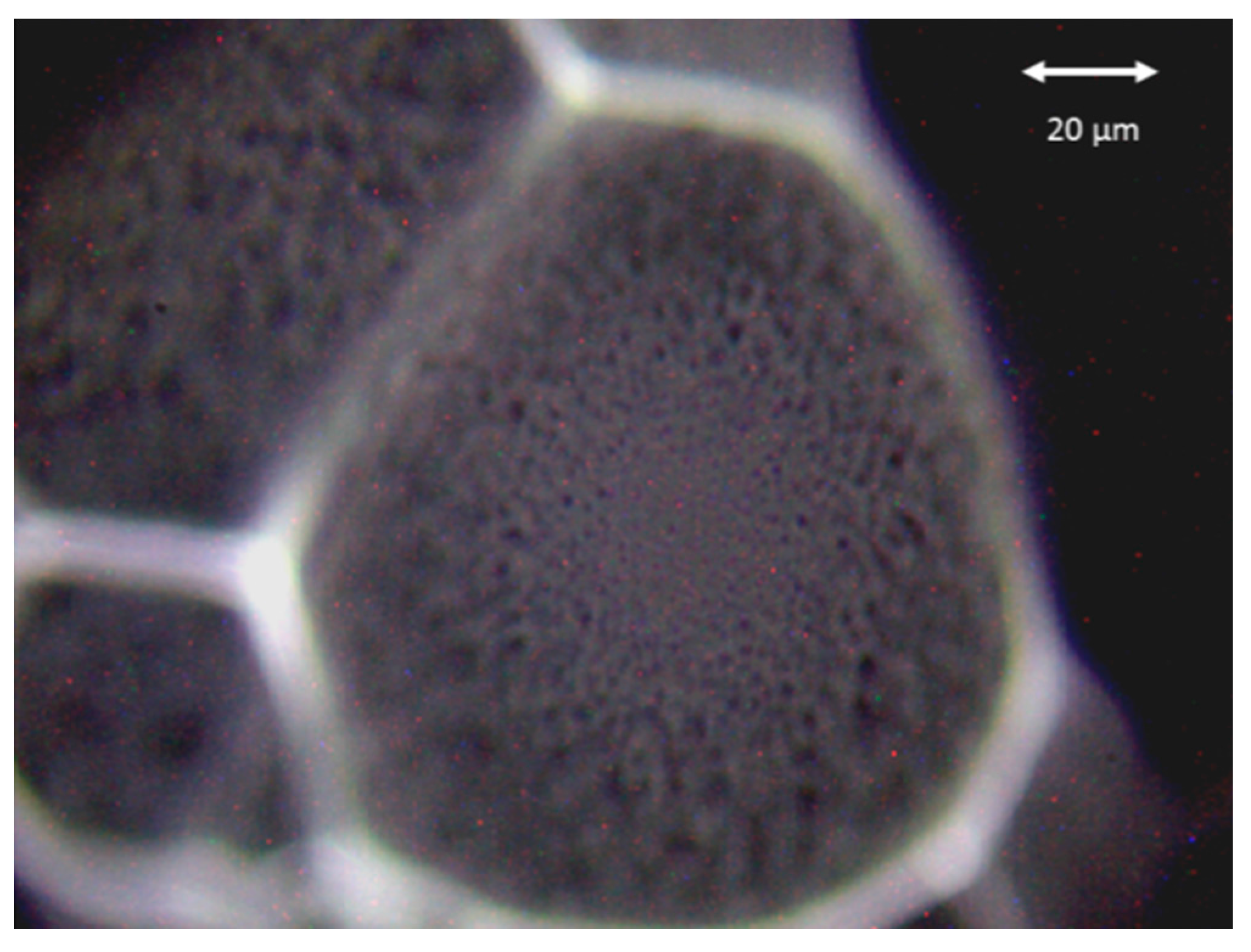
3.1. Microorganisms and Sample Preparation

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Medium/Organism | Staphylococcus aureus | Staphylococcus epidermidis | Escherichia coli | Candida albicans |
|---|---|---|---|---|
| Mueller-Hinton agar | x | x | x | x |
| Blood agar | x | x | x | |
| Blood agar-NaCl | x | x | ||
| CHROMagar Candida | x | |||
| Roosvelt-Park Institute Medium 1640 with L-glutamine | x | x | x | x |
| Sabouraud agar | x | |||
| Endo agar | x |
3.2. Experimental Setup
3.3. Data Analysis
4. Conclusions
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Schie, I.W.; Huser, T. Methods and applications of Raman microspectroscopy to single-cell analysis. Appl. Spectrosc. 2013, 67, 813–828. [Google Scholar] [PubMed]
- Read, D.S.; Whiteley, A.S. Chemical fixation methods for Raman spectroscopy-based analysis of bacteria. J. Microbiol. Methods 2015, 109, 79–83. [Google Scholar] [CrossRef] [PubMed]
- Maquelin, K.; Kirschner, C.; Choo-Smith, L.P.; Ngo-Thi, N.A.; van Vreeswijk, T.; Stammler, M.; Endtz, H.P.; Bruining, H.A.; Naumann, D.; Puppels, G.J. Prospective study of the performance of vibrational spectroscopies for rapid identification of bacterial and fungal pathogens recovered from blood cultures. J. Clin. Microbiol. 2003, 41, 324–329. [Google Scholar] [CrossRef] [PubMed]
- Afseth, N.K.; Bloomfield, M.; Wold, J.P.; Matousek, P.A. Novel approach for subsurface through-skin analysis of salmon using spatially offset raman spectroscopy (SORS). Appl. Spectrosc. 2014, 68, 255–262. [Google Scholar] [CrossRef] [PubMed]
- Notingher, I. Raman spectroscopy cell-based biosensors. Sensors 2007, 7, 1343–1358. [Google Scholar] [CrossRef]
- Almarashi, J.F.M.; Kapel, N.; Wilkinson, T.S.; Telle, H.H. Raman spectroscopy of bacterial species and strains cultivated under reproducible conditions. Spectrosc. Int. J. 2012, 27, 361–365. [Google Scholar] [CrossRef]
- De Gelder, J.; de Gussem, K.; Vandenabeele, P.; Moens, L. Reference database of Raman spectra of biological molecules. J. Raman Spectrosc. 2007, 38, 1133–1147. [Google Scholar] [CrossRef]
- Martinelli, A. Effects of a protic ionic liquid on the reaction pathway during non-aqueous sol-gel synthesis of silica: A Raman spectroscopic investigation. Int. J. Mol. Sci. 2014, 15, 6488–6503. [Google Scholar] [CrossRef] [PubMed]
- Brauchle, E.; Schenke-Leyland, K. Raman spectroscopy in biomedicine—Non-invasive in vitro analysis of cells and extracellular matrix components in tissues. Biotechnol. J. 2013, 8, 288–297. [Google Scholar] [CrossRef] [PubMed]
- Samek, O.; Al-Marashi, J.F.M.; Telle, H.H. The potential of Raman spectroscopy for the identification of biofilm formation by Staphylococcus epidermidis. Laser Phys. Lett. 2010, 7, 378–383. [Google Scholar] [CrossRef]
- Samek, O.; Telle, H.H.; Harris, L.G.; Bloomfield, M.; Mack, D. Raman spectroscopy for rapid discrimination of Staphylococcus epidermidis clones related to medical device-associated infections. Laser Phys. Lett. 2008, 5, 465–470. [Google Scholar] [CrossRef]
- Bernatová, S.; Samek, O.; Pilát, Z.; Šerý, M.; Ježek, J.; Jákl, P.; Šiler, M.; Krzyžánek, V.; Zemánek, P.; Holá, V.; et al. Following the mechanisms of bacteriostatic versus bactericidal action using Raman spectroscopy. Molecules 2013, 18, 13188–13199. [Google Scholar] [CrossRef] [PubMed]
- Samek, O.; Jonáš, A.; Pilát, Z.; Zemánek, P.; Nedbal, L.; Tříska, J.; Kotas, P.; Trtílek, M. Raman microspectroscopy of individual algal cells: Sensing unsaturation of storage lipids in vivo. Sensors 2010, 10, 8635–8651. [Google Scholar] [CrossRef] [PubMed]
- Sandt, C.; Smith-Palmer, T.; Pink, J.; Brennan, L.; Pink, D. Confocal Raman microspectroscopy as a tool for studying the chemical heterogeneities of biofilms in situ. J. Appl. Microbiol. 2007, 103, 1808–1820. [Google Scholar] [CrossRef] [PubMed]
- Choo-Smith, L.P.; Marquelin, K.; van Vreeswijk, T.; Bruining, H.A.; Puppels, G.J.; Ngo Thi, N.A.; Kirchner, C.; Naumann, D.; Ami, D.; Villa, A.M.; et al. Investigating microbial (Micro)colony heterogeneity by vibrational spectroscopy. Appl. Environ. Microbiol. 2001, 67, 1461–1469. [Google Scholar] [CrossRef] [PubMed]
- Samek, O.; Mlynariková, K.; Bernatová, S.; Ježek, J.; Krzyžánek, V.; Šiler, M.; Zemánek, P.; Růžička, F.; Holá, V.; Mahelová, M. Candida parapsilosis Biofilm Identification by Raman Spectroscopy. Int. J. Mol. Sci. 2014, 15, 23924–23935. [Google Scholar] [CrossRef] [PubMed]
- Maquelin, K.; Choo-Smith, L.P.; van Vreeswijk, T.; Endtz, H.P.; Smith, B.; Bennett, R.; Bruining, H.A.; Puppels, G.J. Raman Spectroscopic Method for Identification of Clinically Relevant Microorganisms Growing on Solid Culture Medium. Anal. Chem. 2000, 72, 12–19. [Google Scholar] [CrossRef] [PubMed]
- Maquelin, K.; Choo-Smith, L.P.; Endtz, H.P.; Bruining, H.A.; Puppels, G.J. Rapid identification of Candida species by confocal Raman microspectroscopy. J. Clin. Microbiol. 2002, 40, 594–600. [Google Scholar] [CrossRef] [PubMed]
- Almarashi, J.F.M.; Kapel, N.; Wilkinson, T.S.; Telle, H.H. Raman spectroscopy of bacterial species and strains cultivated under reproducible conditions. In Advances in Biomedical Spectroscopy; IOS Press: Amsterdam, The Netherlands, 2013. [Google Scholar]
- Espagnon, I.; Ostrovskii, D.; Mathey, R.; Dupoy, M.; Joly, P.L.; Novelli-Rousseau, A.; Pinston, F.; Gal, O.; Mallard, F.; Leroux, D.F. Direct identification of clinically relevant bacterial and yeast microcolonies and macrocolonies on solid culture media by Raman spectroscopy. J. Biomed. Opt. 2014, 19. [Google Scholar] [CrossRef] [PubMed]
- Wulf, M.W.H.; Willemse-Erix, D.; Verduin, C.M.; Puppels, G.; van Belkum, A.; Maquelin, K. The use of Raman spectroscopy in the epidemiology of methicillin-resistant Staphylococcus aureus of human- and animal-related clonal lineages. Clin. Microbiol. Infect. 2012, 18, 147–152. [Google Scholar] [CrossRef] [PubMed]
- Mathey, R.; Dupoy, M.; Espagnon, I.; Leroux, D.; Mallard, F.; Novelli-Rousseau, A. Viability of 3 h grown bacterial micro-colonies after direct Raman identification. J. Microbiol. Methods 2015, 109, 67–73. [Google Scholar] [CrossRef] [PubMed]
- Schuster, K.C.; Urlaub, E.; Gapes, J.R. Single-cell analysis of bacteria by Raman microscopy: Spectral information on the chemical composition of cells and on the heterogeneity in a culture. J. Microbiol. Methods 2000, 42, 29–38. [Google Scholar] [CrossRef]
- Vandenbergh, M.F.; Verbrugh, H.A. Carriage of Staphylococcus aureus: Epidemiology and clinical relevance. J. Lab. Clin. Med. 1999, 133, 525–534. [Google Scholar] [CrossRef]
- Piette, A.; Verschraegen, G. Role of coagulase-negative staphylococci in human disease. Vet. Microbiol. 2009, 134, 45–54. [Google Scholar] [CrossRef] [PubMed]
- Kocianova, S.; Vuong, C.; Yao, Y.; Voyich, J.M.; Fischer, E.R.; DeLeo, F.R.; Otto, M. Key role of poly-g-DL-glutamic acid in immune evasion and virulence of Staphylococcus epidermidis. J. Clin. Investig. 2005, 115, 688–694. [Google Scholar] [CrossRef] [PubMed]
- Lindberg, E.; Adlerberth, I.; Matricardi, P.; Bonanno, C.; Tripodi, S.; Panetta, V.; Hesselmar, B.; Saalman, R.; Åberg, N.; Wold, A.E. Effect of lifestyle factors on Staphylococcus aureus gut colonization in Swedish and Italian infants. Clin. Microbiol. Infect. 2011, 17, 1209–1215. [Google Scholar] [CrossRef] [PubMed]
- Van den Berg, S.; Bonarius, H.P.J.; van Kessel, K.P.M.; Elsinga, G.S.; Kooi, N.; Westra, H.; Bosma, T.; van der Kooi-Pol, M.M.; Koedijk, D.G.A.M.; Groen, H.; et al. A human monoclonal antibody targeting the conserved staphylococcal antigen IsaA protects mice against Staphylococcus aureus bacteremia. Int. J. Med. Microbiol. 2015, 305, 55–64. [Google Scholar] [CrossRef] [PubMed]
- Cosgrove, S.E. The relationship between antimicrobial resistance and patient outcomes: Mortality, length of hospital stay, and health care costs. Clin. Infect. Dis. 2006, 42 (Suppl. S2), S82–S89. [Google Scholar] [CrossRef] [PubMed]
- McCann, M.T.; Gilmore, B.F.; Gorman, S.P. Staphylococcus epidermidis device-related infections: Pathogenesis and clinical management. J. Pharm. Pharmacol. 2008, 60, 1551–1571. [Google Scholar] [CrossRef] [PubMed]
- Verhoef, J.; Fleer, A. Staphylococcus epidermidis endocarditis and Staphylococcus epidermidis infection in an intensive care unit. Scand. J. Infect. Dis. Suppl. 1983, 41, 56–64. [Google Scholar] [PubMed]
- Jansen, B.; Hartmann, C.; Schaumacher-Pedreau, F.; Peters, G. Late onset endopthalmitis associated with intraocular lens: A case of molecularly proved S. epidermidis aetiology. Br. J. Ophthalmol. 1991, 75, 440–441. [Google Scholar] [CrossRef] [PubMed]
- Warren, J.W. Catheter-associated urinary tract infection. Int. J. Antimicrob. Agents 2001, 17, 299–303. [Google Scholar] [CrossRef]
- Rupp, M.E.; Archer, G.L. Coagulase-negative staphylococci: Pathogens associated with medical progress. Clin. Infect. Dis. 1994, 19, 231–245. [Google Scholar] [CrossRef] [PubMed]
- Rupp, M.E.; Hamer, K.E. Effect of subinhibitory concentrations of vancomycin, cefazolin, ofloxacin, L-ofloxacin and D-ofloxacin on adherence to intravascular catheters and biofilm formation by Staphylococcus epidermidis. J. Antimicrob. Chemother. 1998, 41, 155–161. [Google Scholar] [CrossRef] [PubMed]
- Gallo, J.; Kolar, M.; Novotny, R.; Rihakova, P.; Ticha, V.V. Pathogenesis of prosthesis-related infection. Biomed. Pap. Med. Fac. Univ. Palacky Olomouc Czech. Repub. 2003, 147, 27–35. [Google Scholar] [CrossRef] [PubMed]
- Ip, D.; Yam, S.K.; Chen, C.K. Implications of the changing pattern of bacterial infections following total joint replacements. J. Orthop. Surg. 2005, 13, 125–130. [Google Scholar]
- Riley, L.W. Pandemic lineages of extraintestinal pathogenic Escherichia coli. Clin. Microbiol. Infect. 2014, 20, 380–390. [Google Scholar] [CrossRef] [PubMed]
- Trofa, D.; Gácser, A.; Nosanchuk, J.D. Candida parapsilosis, an emerging fungal pathogen. Clin. Microbiol. Rev. 2008, 21, 606–625. [Google Scholar] [CrossRef] [PubMed]
- Hattori, H.; Iwataa, T.; Nakagawa, Y.; Kawamoto, F.; Tomitaa, Y.; Kikuchi, A.; Kanbe, T. Genotype analysis of Candida albicans isolates obtained from different body locations of patientswith superficial candidiasis using PCRs targeting 25S rDNA and ALT repeat sequences of the RPS. J. Dermatol. Sci. 2006, 42, 31–46. [Google Scholar] [CrossRef] [PubMed]
- Lim, C.S.Y.; Rosli, R.; Seow, H.F.; Chong, P.P. Candida and invasive candidiasis: Back to basics. Eur. J. Clin. Microbiol. Infect. Dis. 2012, 31, 21–31. [Google Scholar] [CrossRef] [PubMed]
- Machová, E.; Fiačanová, L.; Čížová, A.; Korcová, J. Mannoproteins from yeast and hyphal form of Candida albicans considerably differ in mannan and protein content. Carbohydr. Res. 2015, 408, 12–17. [Google Scholar] [CrossRef] [PubMed]
- Yan, L.; Yang, C.; Tang, J. Disruption of the intestinal mucosal barrier in Candida albicans infections. Microbiol. Res. 2013, 168, 389–395. [Google Scholar] [CrossRef] [PubMed]
- Chandra, J.; Kuhn, D.M.; Mukherjee, P.K.; Hoyer, L.L.; McCormick, T.; Ghannoum, M.A. Biofilm formation by the fungal pathogen Candida albicans: Development, architecture, and drug resistance. J. Bacteriol. 2001, 183, 5385–5394. [Google Scholar] [CrossRef] [PubMed]
- Renishaws EasyConfocal Raman Method. Technology Note from the Spectroscopy Products Division; SPD/TN/076; Issue 1.2; Renishaw PLC: Wotton-under-Edge, UK, 2003.
- De Maesschalck, R.; Jouan-Rimbaud, D.; Massart, D.L. The Mahalanobis distance. Chemometr. Intell. Lab. 2000, 50, 1–18. [Google Scholar] [CrossRef]
© 2015 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Mlynáriková, K.; Samek, O.; Bernatová, S.; Růžička, F.; Ježek, J.; Hároniková, A.; Šiler, M.; Zemánek, P.; Holá, V. Influence of Culture Media on Microbial Fingerprints Using Raman Spectroscopy. Sensors 2015, 15, 29635-29647. https://doi.org/10.3390/s151129635
Mlynáriková K, Samek O, Bernatová S, Růžička F, Ježek J, Hároniková A, Šiler M, Zemánek P, Holá V. Influence of Culture Media on Microbial Fingerprints Using Raman Spectroscopy. Sensors. 2015; 15(11):29635-29647. https://doi.org/10.3390/s151129635
Chicago/Turabian StyleMlynáriková, Katarína, Ota Samek, Silvie Bernatová, Filip Růžička, Jan Ježek, Andrea Hároniková, Martin Šiler, Pavel Zemánek, and Veronika Holá. 2015. "Influence of Culture Media on Microbial Fingerprints Using Raman Spectroscopy" Sensors 15, no. 11: 29635-29647. https://doi.org/10.3390/s151129635
APA StyleMlynáriková, K., Samek, O., Bernatová, S., Růžička, F., Ježek, J., Hároniková, A., Šiler, M., Zemánek, P., & Holá, V. (2015). Influence of Culture Media on Microbial Fingerprints Using Raman Spectroscopy. Sensors, 15(11), 29635-29647. https://doi.org/10.3390/s151129635