Dispersal, Isolation, and Interaction in the Islands of Polynesia: A Critical Review of Archaeological and Genetic Evidence


Abstract
1. Introduction
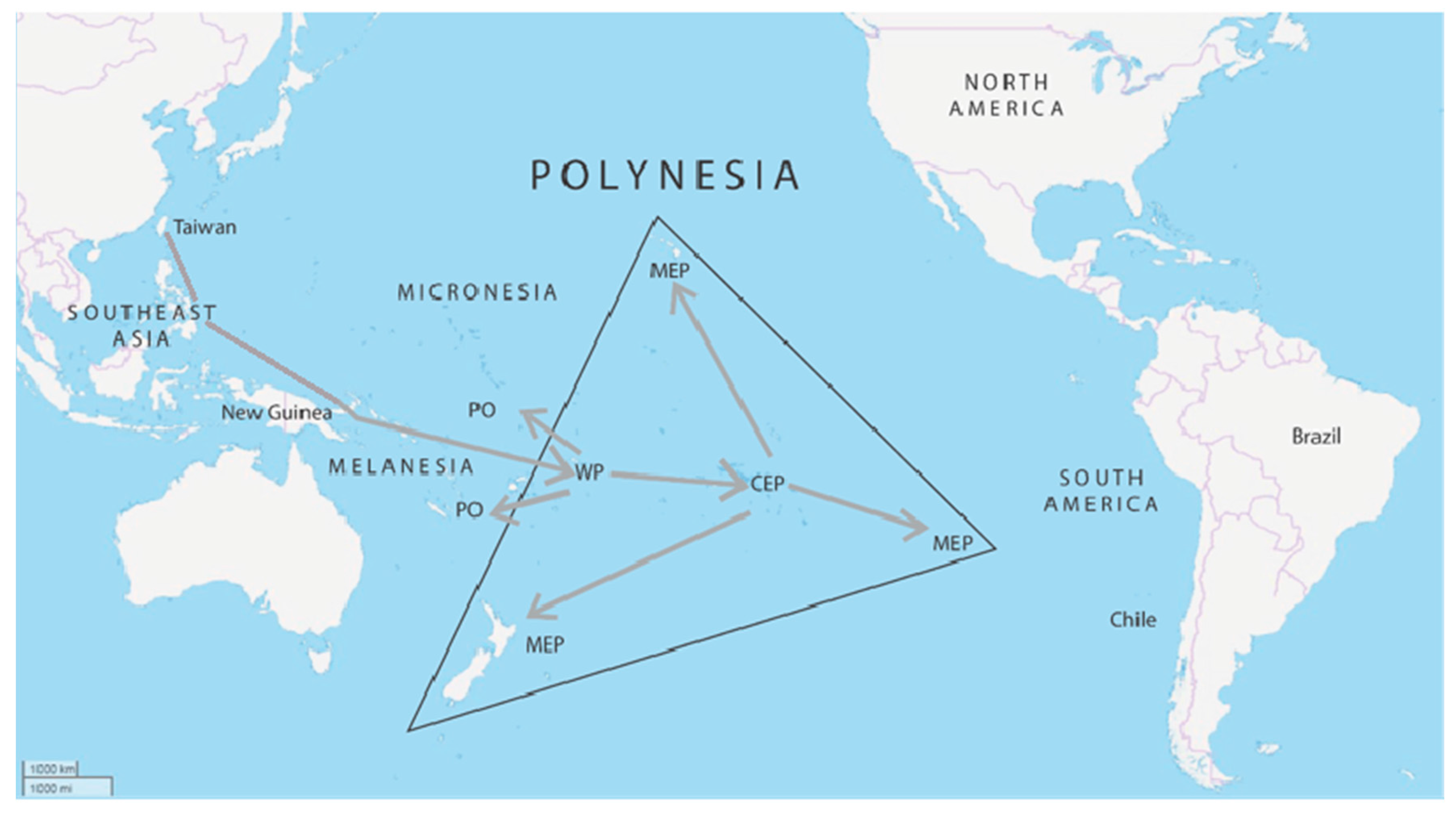
2. Origins of the Polynesians
2.1. Archaeology
2.1.1. The New Chronometric Hygiene
2.1.2. Chronology of Settlement of Western Polynesia
2.1.3. Chronology of Settlement of Eastern Polynesia
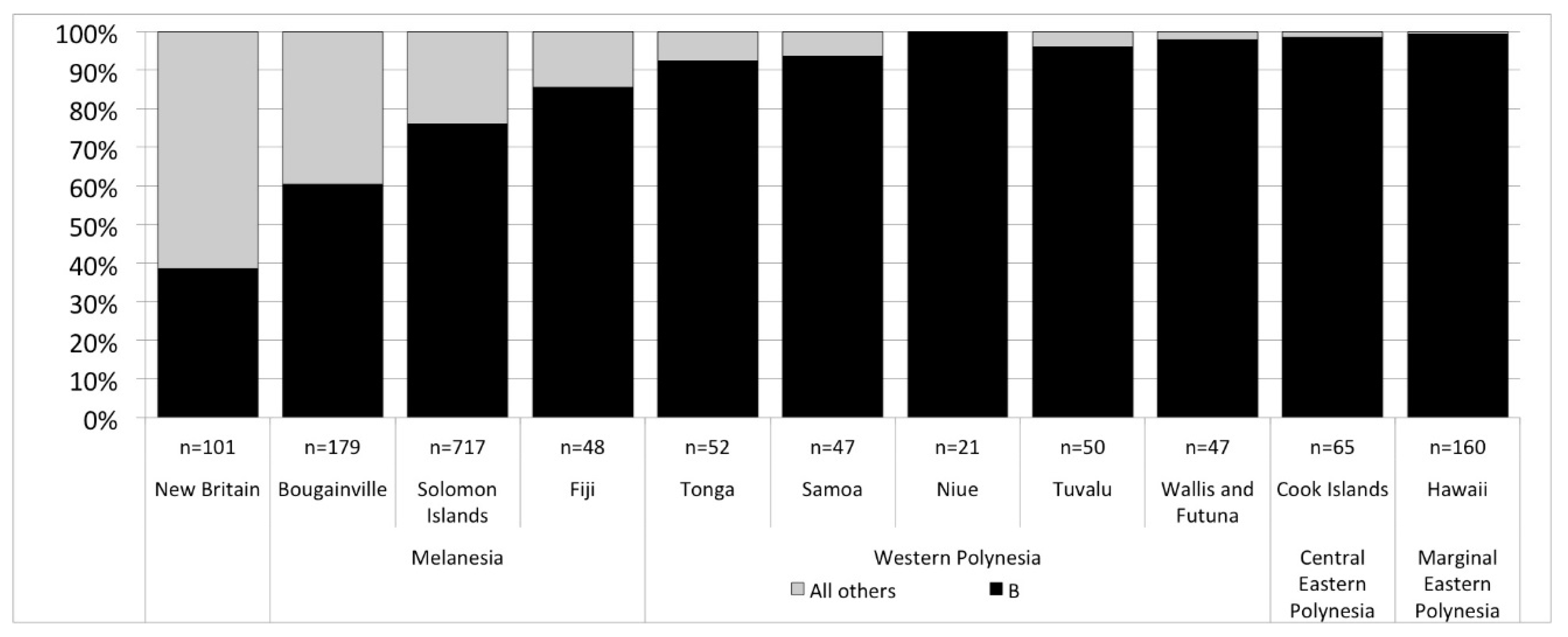
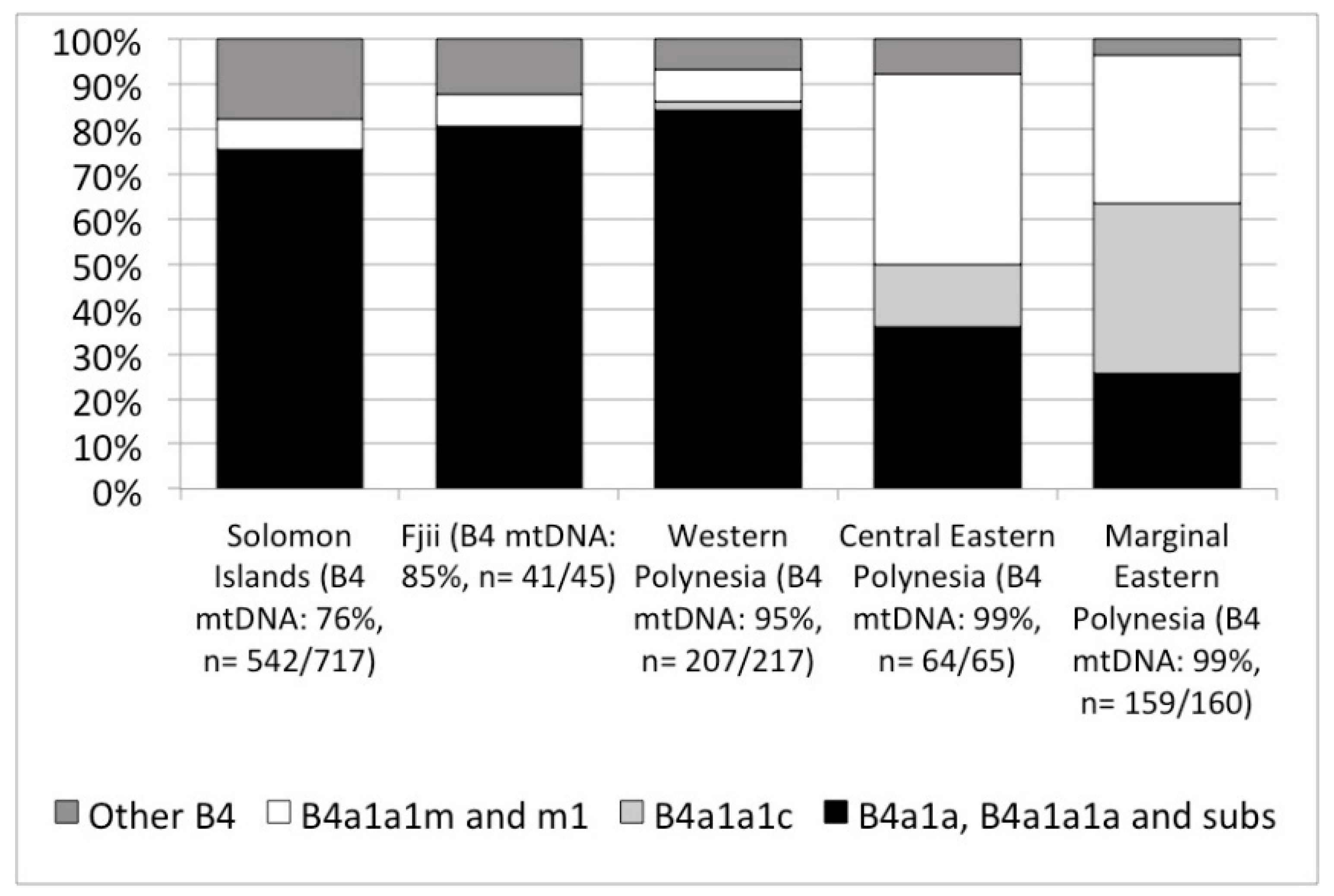
2.2. Modern DNA
2.3. Ancient DNA
3. Long-Distance Contacts between Polynesians and Native Americas
3.1. Archaeology
3.2. Modern DNA
3.3. Ancient DNA
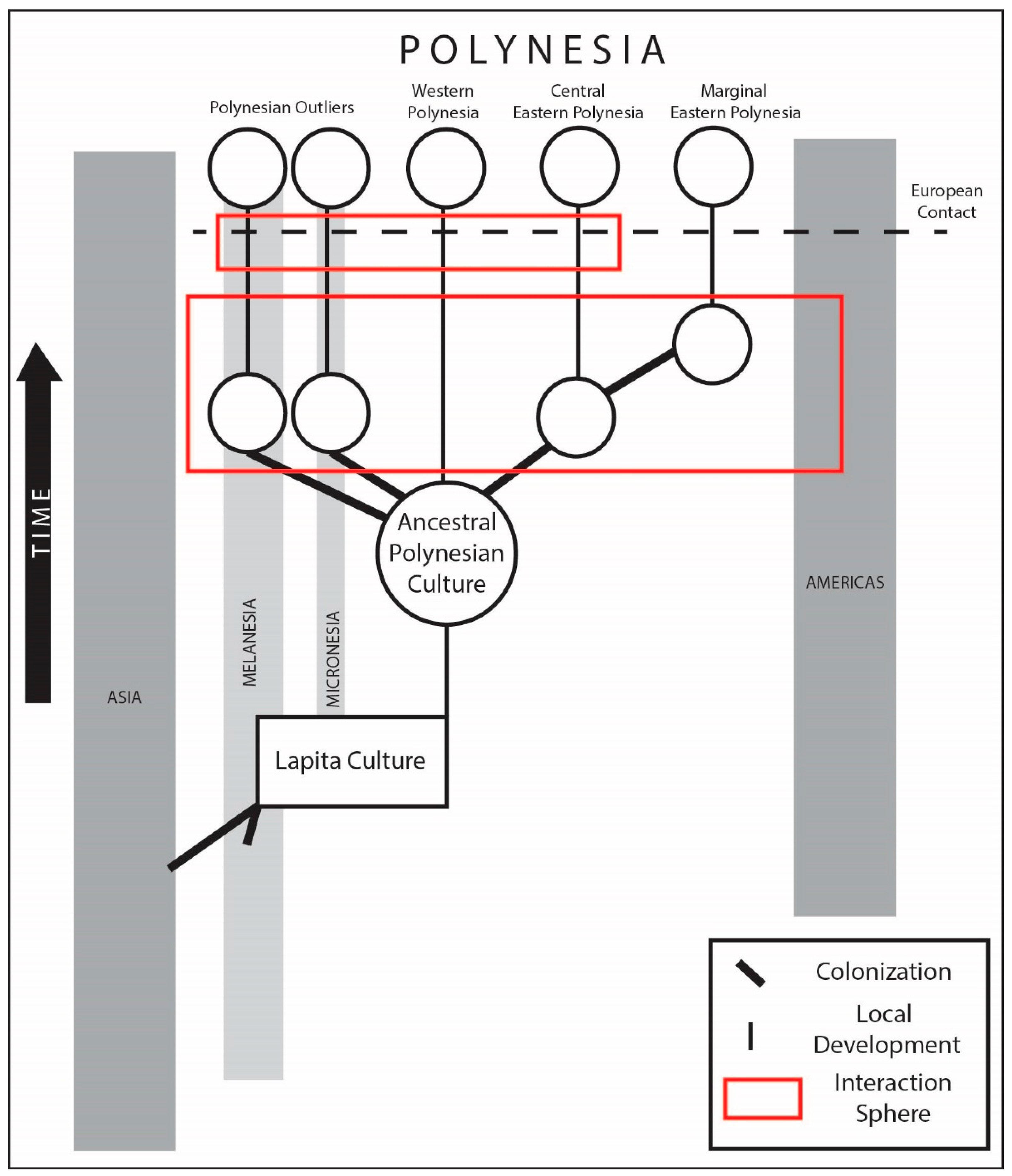
4. Patterns of Dispersal, Interaction, and Isolation
5. Conclusions
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Keegan, W.F.; Diamond, J.M. Colonization of Islands by Humans: A biogeographic perspective. Adv. Archaeol. Method Theory 1987, 10, 49–92. [Google Scholar]
- MacArthur, R.H.; Wilson, E.O. The Theory of Island Biogeography; Princeton University Press: Princeton, NJ, USA, 1967. [Google Scholar]
- Mead, M. Introduction to Polynesia as a laboratory for the development of models in the study of cultural evolution. J. Polyn. Soc. 1957, 66, 145. [Google Scholar]
- Kirch, P.V. Hawaii as a model system for human ecodynamics. Am. Anthropol. 2007, 109, 8–26. [Google Scholar] [CrossRef]
- Vitousek, P.M. Oceanic islands as model systems for ecological studies. J. Biogeogr. 2002, 29, 573–582. [Google Scholar] [CrossRef]
- Slatkin, M.; Racimo, F. Ancient DNA and human history. Proc. Natl. Acad. Sci. USA 2016, 113, 6380–6387. [Google Scholar] [CrossRef] [PubMed]
- Haber, M.; Mezzavilla, M.; Xue, Y.; Tyler-Smith, C. Ancient DNA and the rewriting of human history: Be sparing with Occam’s razor. Genome Biol. 2016, 17. [Google Scholar] [CrossRef] [PubMed]
- Horsburgh, K.A. Molecular anthropology: The judicial use of genetic data in archaeology. J. Archaeol. Sci. 2015, 56, 141–145. [Google Scholar] [CrossRef]
- Horsburgh, K.A.; Orton, J.; Klein, R.G. Beware the springbok in sheep’s clothing: How secure are the faunal identifications upon which we build our models? Afr. Archaeol. Rev. 2016, 33, 353–361. [Google Scholar] [CrossRef]
- Fitzpatrick, S.M.; Anderson, A. Islands of isolation: Archaeology and the power of aquatic perimeters. J. Isl. Coast. Archaeol. 2008, 3, 4–16. [Google Scholar] [CrossRef]
- D’Urville, J.S.C.D. Sur les iles du Grand Ocean. Bulletin de la Societe de Geographie 1832, 17, 1–21. [Google Scholar]
- Bellwood, P. Man’s Conquest of the Pacific: The Prehistory of Southeast Asia and Oceania; Collins Publishers Ltd.: Auckland, New Zealand, 1978. [Google Scholar]
- Kirch, P.V.; Green, R.C. Hawaiki, Ancestral Polynesia: An Essay in Historical Anthropology; Cambridge University Press: Cambridge, UK, 2001. [Google Scholar]
- Boyd, R.; Mulder, M.B.; Durham, W.H.; Richerson, P.J. Are cultural phylogenies possible? In Human by Nature: Between Biology and the Social Sciences; Weingart, P., Mitchell, S.D., Richerson, P.J., Maasen, S., Eds.; Lawrence Erlbaum Associates: London, UK, 1997; pp. 355–386. [Google Scholar]
- Addison, D.J.; Matisoo-Smith, E. Rethinking Polynesians origins: A West-Polynesia Triple-I Model. Archaeol. Ocean. 2010, 45, 1–12. [Google Scholar] [CrossRef]
- Burley, D.V.; Addison, D.J. Tonga and Sāmoa in Oceanic prehistory: Contemporary debates and personal perspectives. In The Oxford Handbook of Prehistoric Oceania; Cochrane, E., Hunt, T., Eds.; Oxford University Press: Oxford, UK, 2014. [Google Scholar]
- Cochrane, E.E.; Kane, H.; Fletcher, C., III; Horrocks, M.; Mills, J.; Barbee, M.; Morrison, A.E.; Tautunu, M.M. Lack of suitable coastal plains likely influenced Lapita (~2800 cal. BP) settlement of Sāmoa: Evidence from south-eastern ′Upolu. Holocene 2015, 26, 126–135. [Google Scholar] [CrossRef]
- Rieth, T.M.; Cochrane, E.E. The Chronology of Colonization in Remote Oceania. In The Oxford Handbook of Prehistoric Oceania; Cochrane, E., Hunt, T., Eds.; Oxford University Press: Oxford, UK, 2014. [Google Scholar]
- Wilmshurst, J.M.; Hunt, T.L.; Lipo, C.P.; Anderson, A.J. High-precision radiocarbon dating shows recent and rapid initial human colonization of East Polynesia. Proc. Natl. Acad. Sci. USA 2011, 108, 1815–1820. [Google Scholar] [CrossRef] [PubMed]
- Spriggs, M.; Anderson, A. Late colonization of East Polynesia. Antiquity 1993, 67, 200–217. [Google Scholar] [CrossRef]
- Allen, M.S.; Huebert, J.M. Short-Lived Plant Materials, Long-Lived Trees, and Polynesian 14C Dating: Considerations for 14C Sample Selection and Documentation. Radiocarbon 2014, 56, 257–276. [Google Scholar] [CrossRef]
- Petchey, F.; Allen, M.A.; Addison, D.J.; Anderson, A. Stability in the South Pacific surface marine 14C reservoir over the last 750 years. Evidence from American Samoa, the southern Cook Islands and the Marquesas. J. Archaeol. Sci. 2009, 36, 2234–2243. [Google Scholar] [CrossRef]
- Weisler, M.I.; Hua, Q.; Zhao, J. Late Holocene C-14 marine reservoir ages in Hawaii from U-series dated corals. Radiocarbon 2009, 51, 955–968. [Google Scholar] [CrossRef]
- Rieth, T.M.; Athens, J.S. Suggested best practices for the application of radiocarbon dating to Hawaiian archaeology. Hawaii. Arch. 2013, 13, 3–29. [Google Scholar]
- Smith, I. Protocols for organising radiocarbon dated assemblages from New Zealand archaeological sites for comparative analysis. J. Pac. Archaeol. 2010, 1, 184–187. [Google Scholar]
- Carson, M. Chronology of Kaua’i: Colonisation, land use demography. J. Polyn. Soc. 2006, 115, 173–186. [Google Scholar]
- McCoy, M.D. A revised late Holocene culture history for Moloka’i Island. Hawai’i Radiocarb. 2007, 49, 1273–1322. [Google Scholar] [CrossRef]
- Rieth, T.M.; Hunt, T.L.; Lipo, C.; Wilmshurst, J.M. The 13th century Polynesian colonization of Hawai’i Island. J. Archaeol. Sci. 2011, 38, 2740–2749. [Google Scholar] [CrossRef]
- Rieth, T.M.; Hunt, T.L. A radiocarbon chronology for Sāmoan prehistory. J. Archaeol. Sci. 2008, 35, 1901–1927. [Google Scholar] [CrossRef]
- Mulrooney, M.A. An island-wide assessment of the chronology of settlement and land use on Rapa Nui (Easter Island) based on radiocarbon data. J. Archaeol. Sci. 2013, 40, 4377–4399. [Google Scholar] [CrossRef]
- Athens, J.S.; Rieth, T.M.; Dye, T.S. A paleoenvironmental and archaeological model-based age estimate for the colonization of Hawai’i. Am. Antiq. 2014, 79, 144–155. [Google Scholar] [CrossRef]
- Mulrooney, M.A.; Bickler, S.H.; Allen, M.S.; Ladefoged, T.N. High-precision dating of colonization and settlement in East Polynesia. Proc. Natl. Acad. Sci. USA 2011, 108, E192–E194. [Google Scholar] [CrossRef] [PubMed]
- Wilmshurst, J.M.; Hunt, T.L.; Lipo, C.P.; Anderson, A.J. Reply to Mulrooney et al.: Accepting lower precision radiocarbon dates results in longer colonization chronologies for East Polynesia. Proc. Natl. Acad. Sci. USA 2011, 108, E195. [Google Scholar] [CrossRef]
- Kirch, P.V. When did the Polynesians settle Hawaii? A review of 150 years of scholarly inquiry and a tentative answer. Hawaii. Arch. 2011, 12, 3–26. [Google Scholar]
- Dye, T.S. Dating human dispersal in Remote Oceania: A Bayesian view from Hawai‘i. World Archaeol. 2015, 47, 661–676. [Google Scholar] [CrossRef]
- Burley, D.; Weisler, M.I.; Zhao, J. High precision U/Th dating of first Polynesian settlement. PLoS ONE 2012, 7, e48769. [Google Scholar] [CrossRef] [PubMed]
- Burley, D.V.; Barton, A.; Dickinson, W.R.; Connaughton, S.P.; Taché, K. Nukuleka as a founder colony for West Polynesian settlement: New insights from recent excavations. J. Pac. Archaeol. 2010, 1, 128–144. [Google Scholar]
- Burley, D.; Edinborough, K.; Weisler, M.; Zhao, J. Bayesian modeling and chronological precision for Polynesian settlement of Tonga. PLoS ONE 2015, 10, e0120795. [Google Scholar] [CrossRef] [PubMed]
- Rieth, T.M.; Morrison, A.E.; Addison, D.J. The temporal and spatial patterning of the initial settlement of Sāmoa. J. Isl. Coast. Archaeol. 2008, 3, 214–239. [Google Scholar] [CrossRef]
- Smith, A. An Archaeology of West Polynesian Prehistory; Australian National University Press: Canberra, Australia, 2002. [Google Scholar]
- Davidson, J.M. Intrusion, integration and innovation on small and not-so-small islands with particular reference to Samoa. Archaeol. Ocean. 2012, 47, 1–13. [Google Scholar] [CrossRef]
- Kirch, P.V.; Yen, D.E. Tikopia: The prehistory and ecology of a Polynesian Outlier; Bishop Museum Press: Honolulu, HI, USA, 1982. [Google Scholar]
- Storey, A.A.; Clarke, A.C.; Ladefoged, T.; Robins, J.; Matisoo-Smith, E. DNA and Pacific Commensal Models: Construction, limitations, and future prospects. J. Isl. Coast. Archaeol. 2013, 8, 37–65. [Google Scholar] [CrossRef]
- Greig, K.; Walter, R.; Matisoo-Smith, E.A. Dogs and people in Southeast Asia and the Pacific. In Routledge Handbook of Bioarchaeology in Southeast Asia and the Pacific Islands; Oxenham, M., Buckley, H., Eds.; Routledge: Abingdon, UK, 2016; pp. 462–482. [Google Scholar]
- Larson, G.; Cucchi, T.; Fujita, M.; Matisoo-Smith, E.; Robins, J.; Anderson, A.; Rolett, B.; Spriggs, M.; Dolman, G.; Kim, T.; et al. Phylogeny and ancient DNA of Sus provides insights into neolithic expansion in Island Southeast Asia and Oceania. Proc. Natl. Acad. Sci. USA 2007, 104, 4834–4839. [Google Scholar] [CrossRef] [PubMed]
- Clarke, A. Origins and Dispersal of the Sweet Potato and Bottle Gourd in Oceania: Implications for Prehistoric Human Mobility. Ph.D. Thesis, Massey University, Palmerston North, New Zealand, 2009. [Google Scholar]
- Roullier, C.; Duputié, A.; Wennekes, P.; Benoit, L.; Fernández Bringas, V.M.; Rossel, G.; Tay, D.; McKey, D.; Lebot, V. Disentangling the Origins of Cultivated Sweet Potato (Ipomoea batatas (L.) Lam.). PLoS ONE 2013, 8, e62707. [Google Scholar] [CrossRef]
- Roullier, C.; Kambouo, R.; Paofa, J.; McKey, D.; Lebot, V. On the origin of sweet potato (Ipomoea batatas (L.) Lam.) genetic diversity in New Guinea, a secondary centre of diversity. Heredity 2013, 110, 594–604. [Google Scholar] [CrossRef] [PubMed]
- Burley, D.V. Fijian Polygenesis and the Melanesian/Polynesian divide. Curr. Anthropol. 2013, 54, 436–462. [Google Scholar] [CrossRef]
- Nunn, P. Comment on “Fijian Polygenesis and the Melanesian/Polynesian divide” by David V. Burley. Curr. Anthropol. 2013, 54, 453–454. [Google Scholar]
- Petersen, G. Micronesia’s breadfruit revolution and the evolution of a culture area. Archaeol. Ocean. 2006, 41, 82–92. [Google Scholar] [CrossRef]
- Allen, M.S.; McAlister, A. Early Marquesan settlement patterns of interaction: New insights from Hatiheu Valley, Nuku Hiva Island. J. Pac. Archaeol. 2013, 4, 90–109. [Google Scholar]
- Allen, M.S.; Wallace, R. New evidence from the East Polynesian gateway: Substantive and methodological results from Aitutaki, southern Cook Islands. Radiocarbon 2007, 49, 1163–1179. [Google Scholar] [CrossRef]
- Bollt, R. Peva: The Archaeology of an Austral Island Settlement. Bulletins in Anthropology; Bishop Museum Press: Honolulu, HI, USA, 2008. [Google Scholar]
- Conte, E.; Molle, G. Reinvestigating a key site for Polynesian prehistory: New results from the Hane dune site, Ua Huka (Marquesas). Archaeol. Ocean. 2014, 49, 121–136. [Google Scholar] [CrossRef]
- Molle, G. Ua Huka, une île dans l’Histoire. Histoire pré-et post-Européenne d’une Société Marquisienne. Ph.D. Thesis, University of French Polynesia, Tahiti, France, 2 December 2011. [Google Scholar]
- Allen, M.S. Marquesan colonization chronologies and post-colonisation interaction: Implications for Hawaiian origins and the ‘Marquesan Homeland’ hypothesis. J. Pac. Archaeol. 2004, 5, 1–17. [Google Scholar]
- Brooks, E.; Jacomb, C.; Walter, R. Archaeological investigations at Wairau Bar. Archaeol. N. Z. 2009, 52, 259–268. [Google Scholar]
- Brooks, E.; Walter, R.; Jacomb, C. History of excavations at Wairau Bar. Rec. Canterb. Mus. 2011, 25, 13–58. [Google Scholar]
- Davidson, J.; Findlater, A.; Fyfe, R.; MacDonald, J.; Marshall, B. Connections with Hawaiki: The evidence of a shell tool from Wairau Bar, Marlborough, New Zealand. J. Pac. Archaeol. 2011, 2, 93–102. [Google Scholar]
- Jacomb, C.; Holdaway, R.; Allentoft, M.E.; Bunce, M.; Oskam, C.; Walter, R. High-precision dating and ancient DNA profiling of moa (Aves: Dinornithiformes) eggshell documents a complex feature at Wairau Bar and refines the chronology of New Zealand settlement by Polynesians. J. Archaeol. Sci. 2014, 50, 24–30. [Google Scholar] [CrossRef]
- Jacomb, C.; Walter, R.; Brooks, E. Living on Pipi (Paphies. australis): Specialised shellfish harvest in a marginal environment at Karamea, West Coast, New Zealand. J. Pac. Archaeol. 2010, 1, 36–52. [Google Scholar]
- Oskam, C.L.; Jacomb, C.; Allentoft, M.E.; Walter, R.; Scofield, R.P.; Haile, J.; Holdaway, R.N.; Bunce, M. Molecular and morphological analyses of avian eggshell excavated from a late thirteenth century earth oven. J. Archaeol. Sci. 2011, 38, 2589–2595. [Google Scholar] [CrossRef]
- Anderson, A. Subpolar settlement in South Polynesia. Antiquity 2005, 79, 791–800. [Google Scholar] [CrossRef]
- Dye, T.S. A model-based age estimate for Polynesian colonization of Hawai’i. Archaeol. Ocean. 2011, 46, 130–138. [Google Scholar] [CrossRef]
- Hunt, T.L.; Lipo, C.P. Late colonization of Easter Island. Science 2006, 311, 1603–1606. [Google Scholar] [CrossRef] [PubMed]
- Anderson, A. Slow boats from China: Issues in the prehistory of Indo-Pacific seafaring. In East of Wallace’s Line: Studies of Past and Present Maritime Cultures of the Indo-Pacific Region; O’Connor, S., Veth, P., Eds.; A Balkema: Rotterdam, The Netherlands, 2000; pp. 13–51. [Google Scholar]
- Anderson, A.; Chappell, J.; Gagan, M.; Grove, R. Prehistoric maritime migration in the Pacific Islands: An hypothesis of ENSO forcing. Holocene 2006, 16, 1–6. [Google Scholar] [CrossRef]
- Goodwin, I.D.; Browning, S.A.; Anderson, A.J. Climate windows for Polynesian voyaging to New Zealand and Easter Island. Proc. Natl. Acad. Sci. USA 2014, 111, 14716–14721. [Google Scholar] [CrossRef] [PubMed]
- Anderson, A. Polynesian seafaring and American horizons: A response to Jones and Klar. Am. Antiq. 2006, 71, 759–763. [Google Scholar] [CrossRef]
- Di Piazza, A.; Di Piazza, P.; Pearthree, E. Sailing virtual canoes across Oceania: Revisiting island accessibility. J. Archaeol. Sci. 2007, 34, 1219–1225. [Google Scholar] [CrossRef]
- Montenegro, Á.; Callaghan, R.T.; Fitzpatrick, S.M. Using seafaring simulations and shortest-hop trajectories to model the prehistoric colonization of Remote Oceania. Proc. Natl. Acad. Sci. USA 2016, 113, 12685–12690. [Google Scholar] [CrossRef] [PubMed]
- Montenegro, A.; Callaghan, R.T.; Fitzpatrick, S.M. From west to east: Environmental influences on the rate and pathways of Polynesian colonization. Holocene 2014, 24, 242–256. [Google Scholar] [CrossRef]
- Walter, R.; Jacomb, C.; Boron-Muth, S. Colonisation, mobility and exchange in New Zealand prehistory. Antiquity 2010, 84, 497–513. [Google Scholar] [CrossRef]
- McCoy, M.D.; Robles, H.R. The Geographic Range of Interaction Spheres during the Colonization of New Zealand (Aotearoa): New evidence for obsidian circulation in Southern New Zealand. J. Isl. Coast. Archaeol. 2015. [Google Scholar] [CrossRef]
- Lawrence, M.; McCoy, M.D.; Barber, I.; Walter, R. Geochemical Sourcing Obsidian from the Purakaunui Site, South Island, New Zealand. Archaeol. Ocean. 2014, 49, 158–163. [Google Scholar] [CrossRef]
- Kinaston, R.L.; Shaw, B.; Gray, A.R.; Walter, R.K.; Jacomb, C.; Brooks, E.; Halcrow, S.E.; Buckley, H.R. Critical Review of Brown and Thomas “The First New Zealanders? An Alternative Interpretation of the Stable Isotope Data from Wairau Bar”. PLoS ONE 2015, 10, e0137616. [Google Scholar] [CrossRef] [PubMed]
- Kinaston, R.L.; Walter, R.K.; Jacomb, C.; Brooks, E.; Tables, N.; Halcrow, S.E.; Stirling, C.; Reid, M.; Gray, A.R.; Spinks, J.; et al. The First New Zealanders: Patterns of Diet and Mobility Revealed through Isotope Analysis. PLoS ONE 2013, 8, e64580. [Google Scholar] [CrossRef]
- Brown, A.A.; Thomas, T. The First New Zealanders? An Alternative Interpretation of Stable Isotope Data from Wairau Bar, New Zeal. PLoS ONE 2015. [Google Scholar] [CrossRef] [PubMed]
- Moodley, Y.; Linz, B.; Yamaoka, Y.; Windsor, H.M.; Breurec, S.; Wu, Y.; Maady, A.; Bernhöft, S.; Thiberge, J.; Phuanukoonnon, S.; et al. The Peopling of the Pacific from a Bacterial Perspective. Science 2009, 323, 527–530. [Google Scholar] [CrossRef] [PubMed]
- Hagelberg, E.; Clegg, J.B. Genetic polymorphism in prehistoric Pacific islanders determined by analysis of ancient bone DNA. Proc. R. Soc. Lond. B Biol. Sci. 1993, 252, 163–170. [Google Scholar] [CrossRef] [PubMed]
- Friedlaender, J.; Schurr, T.; Gentz, F.; Koki, G.; Friedlaender, F.; Horvat, G.; Babb, P.; Cerchio, S.; Kaestle, F.; Schanfield, M.; et al. Expanding Southwest Pacific Mitochondrial Haplogroups P and Q. Mol. Biol. Evol. 2005, 22, 1506–1517. [Google Scholar] [CrossRef] [PubMed]
- Melton, T.; Clifford, S.; Martinson, E.; Batzer, M.; Stoneking, M. Genetic evidence for the proto-Austronesian homeland in Asia: MtDNA and nuclear DNA variation in Taiwanese aboriginal tribes. Am. J. Hum. Genet. 1998, 63, 1807–1823. [Google Scholar] [CrossRef] [PubMed]
- Redd, A.J.; Takezaki, N.; Sherry, S.T.; McGarvey, S.T.; Sofro, A.S.M.; Stoneking, M. Evolutionary history of the COII/tRHA Lys intergenic 9 base pair deletion in human mitochondrial DNAs from the Pacific. Mol. Biol. Evol. 1995, 12, 604–615. [Google Scholar] [PubMed]
- Kayser, M.; Brauer, S.; Cordaux, R.; Casto, A.; Lao, O.; Zhivotovsky, L.A.; Moyse-Faurie, C.; Rutledge, R.B.; Schiefenhoevel, W.; Gil, D.; et al. Melanesian and Asian origins of Polynesians: MtDNA and Y chromosome gradients across the Pacific. Mol. Biol. Evol. 2006, 23, 2234–2244. [Google Scholar] [CrossRef] [PubMed]
- Mirabal, S.; Herrera, K.J.; Gaydena, T.; Regueiro, M.; Underhill, P.A.; Garcia-Bertrand, R.L.; Herrera, R.J. Increased Y-chromosome resolution of haplogroup O suggests genetic ties between the Ami aborigines of Taiwan and the Polynesian Islands of Samoa and Tonga. Gene 2010, 494, 339–348. [Google Scholar] [CrossRef] [PubMed]
- Kayser, M.; van Oven, M. Updated comprehensive phylogenetic tree of global human mitochondrial DNA variation. Hum. Mutat. 2009, 30, E386–E394. [Google Scholar]
- Knapp, M.; Horsburgh, K.A.; Prost, S.; Stanton, J.A.; Buckley, H.R.; Walter, R.K.; Matisoo-Smith, E.A. Complete mitochondrial DNA genome sequences from the first New Zealanders. Proc. Natl. Acad. Sci. USA 2012, 109, 18350–18354. [Google Scholar] [CrossRef] [PubMed]
- Benton, M.; Macartney-Coxson, D.; Eccles, D.; Griffiths, L.; Chambers, G.; Lea, R. Complete Mitochondrial Genome Sequencing Reveals Novel Haplotypes in a Polynesian Population. PLoS ONE 2012, 7, e35026. [Google Scholar] [CrossRef]
- Hather, J.G.; Kirch, P.V. Prehistoric sweet potato (Ipomoea batatas) from Mangaia Island, central Polynesia. Antiquity 1991, 65, 887–893. [Google Scholar] [CrossRef]
- Ladefoged, T.N.; Graves, M.W.; Coil, J.H. The introduction of sweet potato in Polynesia: Early remains in Hawai’i. J. Polyn. Soc. 2005, 114, 359–374. [Google Scholar]
- Montenegro, A.; Avis, C.; Weaver, A.J. Modeling the prehistoric arrival of the sweet potato in Polynesia. J. Archaeol. Sci. 2008, 35, 355–367. [Google Scholar] [CrossRef]
- Jones, T.L.; Klar, K.A. Diffusionism reconsidered: Linguistic and archaeological evidence for prehistoric Polynesian contact with southern California. Am. Antiq. 2005, 70, 457–484. [Google Scholar] [CrossRef]
- Arnold, J.E. Credit Where Credit is Due: The history of the Chumash oceangoing plank canoe. Am. Antiq. 2007, 72, 196–209. [Google Scholar] [CrossRef]
- Jones, T.L.; Storey, A.A.; Matisoo-Smith, E.A.; Ramírez-Aliaga, J.M. (Eds.) Polynesians in America: Pre-Columbian Contacts with the New World; AltaMira Press: New York, NY, USA, 2011. [Google Scholar]
- Storey, A.A.; Ramírez, J.M.; Quiroz, D.; Burley, D.V.; Addison, D.J.; Walter, R.; Anderson, A.J.; Hunt, T.L.; Athens, J.S.; Huynen, L.; et al. Radiocarbon and DNA evidence for a pre-Columbian introduction of Polynesian chickens to Chile. Proc. Natl. Acad. Sci. USA 2007, 104, 10335–10339. [Google Scholar] [CrossRef] [PubMed]
- Storey, A.A.; Ladefoged, T.; Matisoo-Smith, E.A. Counting your chickens: Density and distribution of chicken remains in archaeological sites of Oceania. Int. J. Osteoarchaeol. 2008, 18, 240–261. [Google Scholar] [CrossRef]
- Gongora, J.; Rawlence, N.J.; Mobegi, V.A.; Jianlin, H.; Alcalde, J.A.; Matus, J.T.; Hanotte, O.; Moran, C.; Austin, J.J.; Ulm, S.; et al. Indo-European and Asian origins for Chilean and Pacific chickens revealed by mtDNA. Proc. Natl. Acad. Sci. USA 2008, 105, 10308–10313. [Google Scholar] [CrossRef] [PubMed]
- Fitzpatrick, S.M.; Callaghan, R. Examining dispersal mechanisms for the translocation of chicken (Gallus gallus) from Polynesia to South America. J. Archaeol. Sci. 2009, 36, 214–223. [Google Scholar] [CrossRef]
- Storey, A.A.; Athens, J.S.; Bryant, D.; Carson, M.; Emery, K.; deFrance, S.; Higham, C.; Huynen, L.; Intoh, M.; Jones, S.; et al. Investigating the global dispersal of chickens in prehistory using ancient mitochondrial DNA signatures. PLoS ONE 2012, 7, e39171. [Google Scholar] [CrossRef] [PubMed]
- Thomson, V.A.; Lebrasseur, O.; Austin, J.J.; Hunt, T.L.; Burney, D.A.; Denham, T.; Rawlence, N.J.; Wood, J.R.; Gongora, J.; Flink, L.G.; et al. Reply to Beavan, Bryant, and Storey and Matisoo-Smith: Ancestral Polynesian “D” haplotypes reflect authentic Pacific chicken lineages. Proc. Natl. Acad. Sci. USA 2014, 111, E3585–E3586. [Google Scholar] [CrossRef] [PubMed]
- Beavan, N. No evidence for sample contamination or diet offset for pre-Columbian chicken dates from El Arena 1. Proc. Natl. Acad. Sci. USA 2014, 111, E3582. [Google Scholar] [CrossRef] [PubMed]
- Malaspinas, A.; Oscar, L.; Schroeder, H.; Rasmussen, R.; Raghavan, M.; Moltke, I.; Campos, P.F.; Sagredo, F.S.; Rasmussen, S.; Gonçalves, V.F.; et al. Two ancient human genomes reveal Polynesian ancestry among the indigenous Botocudos of Brazil. Curr. Biol. 2014, 24, R1035–R1037. [Google Scholar] [CrossRef] [PubMed]
- Moreno-Mayar, J.V.; Rasmussen, S.; Seguin-Orlando, A.; Rasmussen, M.; Liang, M.; Flåm, S.T.; Lie, B.A.; Gilfillan, G.D.; Nielsen, R.; Thorsby, E.; et al. Genome-wide Ancestry Patterns in Rapanui Suggest Pre-European Admixture with Native Americans. Curr. Biol. 2014, 24, 2518–2525. [Google Scholar] [CrossRef] [PubMed]
- Mills, P.R.; Lundblad, S.P. Current perspectives on Hawai‘i’s stone tool economies. J. Pac. Archaeol. 2014, 5, 30–39. [Google Scholar]
- Burley, D.V.; Dickinson, W.R. Among Polynesia’s first pots. J. Archaeol. Sci. 2010, 37, 1020–1026. [Google Scholar] [CrossRef]
- Allen, M.S.; Johnson, K.T.M. Tracking ancient patterns of interaction: Recent geochemical studies in the southern Cook Islands. In Prehistoric Long-Distance Interaction in Oceania: An Interdisciplinary Approach; Weisler, M.I., Ed.; NZAA: Auckland, New Zealand, 1997; pp. 111–113. [Google Scholar]
- Sheppard, P.; Walter, R.; Parker, R. Basalt sourcing and the development of Cook Island exchange systems. In Prehistoric Long-distance Interaction in Oceania: An Interdisciplinary Approach; Weisler, M.I., Ed.; NZAA: Auckland, New Zealand, 1997; pp. 85–110. [Google Scholar]
- Weisler, M.I. Hard Evidence for Prehistoric Interaction in Polynesia. Curr. Anthropol. 1998, 39, 521–532. [Google Scholar] [CrossRef]
- Clark, G.R.; Reepmeyer, C.; Melekiola, N.; Woodhead, J.; Dickinson, W.R.; Martinsson-Wallin, H. Stone tools from the ancient Tongan state reveal prehistoric interaction centers in the Central Pacific. Proc. Natl. Acad. Sci. USA 2014, 111, 10491–10496. [Google Scholar] [CrossRef] [PubMed]
- Allen, M.S.; Dickinson, W.R.; Huebert, J.M. The anomaly of Marquesan ceramics: A fifty year retrospective. J. Pac. Archaeol. 2012, 3, 90–104. [Google Scholar]
- Cochrane, E.E.; Rieth, T.M. Sāmoan artefact provenance reveals limited artefact transfer within and beyond the archipelago. Archaeol. Ocean. 2016. [Google Scholar] [CrossRef]
- Burley, D.V.; Sheppard, P.J.; Simonin, M. Tongan and Samoan volcanic glass: PXRF analysis and implications for constructs of ancestral Polynesian society. J. Archaeol. Sci. 2011, 38, 2625–2632. [Google Scholar] [CrossRef]
- Eckert, S.L.; James, W.D. Investigating the production and distribution of plain ware pottery in the Samoan archipelago with laser ablation-inductively coupled plasma-mass spectrometry (LA-ICP-MS). J. Archaeol. Sci. 2011, 38, 2155–2170. [Google Scholar] [CrossRef]
- Collerson, K.D.; Weisler, M.I. Stone adze compositions and the extent of ancient Polynesian voyaging and trade. Science 2007, 307, 1907–1911. [Google Scholar] [CrossRef] [PubMed]
- Anderson, A. Traditionalism, Interaction, and Long-Distance Seafaring in Polynesia. J. Isl. Coast. Archaeol. 2008, 3, 240–250. [Google Scholar] [CrossRef]
- Kahn, J.G.; Sinton, J.; Mills, P.R.; Lundblad, S.P. X-ray fluorescence analysis and intra-island exchange in the Society Island archipelago (Central Eastern Polynesia). J. Archaeol. Sci. 2013, 40, 1194–1202. [Google Scholar] [CrossRef]
- Rolett, B.V.; West, E.W.; Sinton, J.M.; Iovita, R. Ancient East Polynesian voyaging spheres: New evidence from the Vitaria adze quarry (Rurutu, Austral Islands). J. Archaeol. Sci. 2015, 53, 459–471. [Google Scholar] [CrossRef]
- Ditchfield, K.; Holdaway, S.; Allen, M.S.; McAlister, A. Measuring stone artifact transport: The experimental demonstration and pilot application of a new method to a prehistoric adze workshop, southern Cook Islands. J. Archaeol. Sci. 2014, 50, 512–523. [Google Scholar] [CrossRef]
- Ramírez-Aliaga, J.M. The Mapuche Connection. In Polynesians in America: Pre-Columbian contacts with the New World; Jones, T.L., Storey, A.A., Matisoo-Smith, E.A., Ramírez-Aliaga, J.M., Eds.; Alta Mira Press: Lanham, MD, USA, 2011; pp. 95–110. [Google Scholar]
- Roullier, C.; Benoit, L.; McKey, B.D.; Lebot, V. Historical collections reveal patterns of diffusion of sweet potato in Oceania obscured by modern plant movements and recombination. Proc. Natl. Acad. Sci. USA 2013, 110, 2205–2210. [Google Scholar] [CrossRef] [PubMed]
- Clarke, A.C.; Burtenshaw, M.K.; McLenachan, P.A.; Erickson, D.L.; Penny, D. Reconstructing the Origins and Dispersal of the Polynesian Bottle Gourd (Lagenaria siceraria). Mol. Biol. Evol. 2006, 23, 893–900. [Google Scholar] [CrossRef] [PubMed]
- Erickson, D.L.; Smith, B.D.; Clarke, A.C.; Sandweiss, D.H.; Tuross, N. An Asian origin for a 10,000-year-old domesticated plant in the Americas. Proc. Natl. Acad. Sci. USA 2005, 102, 18315–18320. [Google Scholar] [CrossRef] [PubMed]
- Kistler, L.; Montenegro, A.; Smith, B.D.; Gifford, J.A.; Greene, R.E.; Newsom, L.A.; Shapiro, B. Transoceanic drift and the domestication of African bottle gourds in the Americas. Proc. Natl. Acad. Sci. USA 2014, 111, 2937–2941. [Google Scholar] [CrossRef] [PubMed]
- Moncada, X.; Payacan, C.; Arriaza, F.; Lobos, S.; Seelenfreund, D.; Seelenfreund, A. DNA extraction and amplification from contemporary Polynesian bark-cloth. PLoS ONE 2013, 8, e56549. [Google Scholar] [CrossRef] [PubMed]
- Seelenfreund, D.; Clarke, A.C.; Oyanedel, N.; Piña, R.; Lobos, S.; Matisoo-Smith, E.A.; Seelenfreund, A. Paper mulberry (Broussonetia papyrifera) as a commensal model for human mobility in Oceania: Anthropological, botanical and genetic considerations. N. Z. J. Bot. 2010, 48, 231–247. [Google Scholar] [CrossRef]
- Zerega, N.; Wiesner-Hanks, T.; Ragone, D.; Irish, B.; Scheffler, B.; Simpson, S.; Zee, F. Diversity in the breadfruit complex (Artocarpus, Moraceae): Genetic characterization of critical germplasm. Tree Genet. Genomes 2015, 11, 4. [Google Scholar] [CrossRef]
- Greig, K.; Boocock, J.; Prost, S.; Horsburgh, K.A.; Jacomb, C.; Walter, R.; Matisoo-Smith, E. Complete mitochondrial genomes of New Zealand’s first dogs. PLoS ONE 2015, 10, e0138536. [Google Scholar] [CrossRef] [PubMed]
- Storey, A.A.; Matisoo-Smith, E. No evidence against Polynesian dispersal of chickens to pre-Columbian South America. Proc. Natl. Acad. Sci. USA 2014, 111, E3583. [Google Scholar] [CrossRef] [PubMed]
- Anderson, A. Islands of exile: Ideological motivation in maritime migration. J. Isl. Coast. Archaeol. 2006, 1, 33–47. [Google Scholar] [CrossRef]
- Goncalves, V.F.; Stenderup, J.; Rodrigues-Carvalho, C.; Silva, H.P.; Goncalves-Dornelas, H.; Liryo, A.; Kivisild, T.; Malaspinas, A.-S.; Campus, P.F.; Rasmussen, M.; et al. Identification of Polynesian mtDNA haplogroups in remains of Botocudo Amerindians from Brazil. Proc. Natl. Acad. Sci. USA 2013, 110, 6465–6469. [Google Scholar] [CrossRef] [PubMed]
- Kirch, P.V. On the Road of the Winds: An Archaeological History of the Pacific Islands before Europe; University of California Press: Oakland, CA, USA, 2002. [Google Scholar]
- Kirch, P.V. The Evolution of Polynesian Chiefdoms; Cambridge University Press: Cambridge, UK, 1984. [Google Scholar]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Taxa | Melanesia | Micronesia | Western Polynesia | Polynesian Outliers | Eastern Polynesia |
|---|---|---|---|---|---|
| Rat (Rattus exulans) Modern: 266 Ancient: 115 Total: 381 | 86/16 | 3/3 | 2/3 | 0/5 | 175/88 |
| Pig (Sus scrofa) Modern: 88 Ancient: 6 Total: 94 | 47/0 | 5/5 | 0/1 | 0/1 | 36/4 |
| Chicken (Gallus gallus) Modern: 48 Ancient: 31 Total: 79 | 43/3 | 5/4 | 0/4 | 0/2 | 0/18 |
| Dog (Canis lupus fimiliaris) Modern: 6 Ancient: 36 Total: 40 | 6/1 | 0/0 | 0/0 | 0/0 | 0/33 |
| Modern: 408 Ancient: 188 | 182/20 | 13/12 | 2/8 | 0/8 | 211/143 |
© 2017 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Horsburgh, K.A.; McCoy, M.D. Dispersal, Isolation, and Interaction in the Islands of Polynesia: A Critical Review of Archaeological and Genetic Evidence. Diversity 2017, 9, 37. https://doi.org/10.3390/d9030037
Horsburgh KA, McCoy MD. Dispersal, Isolation, and Interaction in the Islands of Polynesia: A Critical Review of Archaeological and Genetic Evidence. Diversity. 2017; 9(3):37. https://doi.org/10.3390/d9030037
Chicago/Turabian StyleHorsburgh, K. Ann, and Mark D. McCoy. 2017. "Dispersal, Isolation, and Interaction in the Islands of Polynesia: A Critical Review of Archaeological and Genetic Evidence" Diversity 9, no. 3: 37. https://doi.org/10.3390/d9030037
APA StyleHorsburgh, K. A., & McCoy, M. D. (2017). Dispersal, Isolation, and Interaction in the Islands of Polynesia: A Critical Review of Archaeological and Genetic Evidence. Diversity, 9(3), 37. https://doi.org/10.3390/d9030037