Does Stream Size Really Explain Biodiversity Patterns in Lotic Systems? A Call for Mechanistic Explanations

 ,
,
Abstract
:1. Introduction
- Quantify how many published articles determined stream size is a predictor of biodiversity.
- Identify the different measures of stream size used in biodiversity studies.
- Identify the types of biodiversity that are being compared across stream size gradients.
- Determine if biodiversity–stream size relationships may consistently differ among studies, systems, and organismal groups, and whether there is consensus regarding the relationships in each of these categories.
- Catalog the mechanistic explanations proposed for biodiversity–stream size relationships, and how often these explanations are invoked.
- Were patterns in our data analysis consistent with those identified in our literature review?
- Did biodiversity–stream size relationships change appreciably with different measures of stream size?
- Did biodiversity–stream size relationships differ among organismal groups?
2. Materials and Methods
2.1. A Systematic Review of Literature on Stream Size and Biodiversity
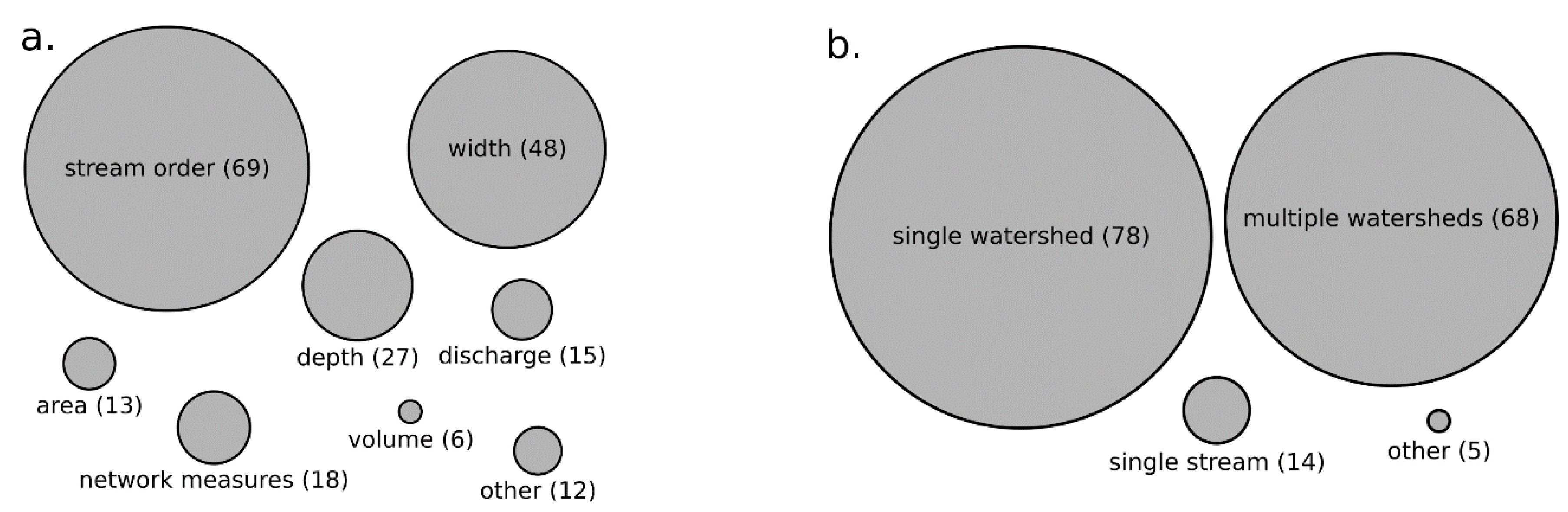
2.2. An Analysis of Stream Size Measures That Influence Biodiversity
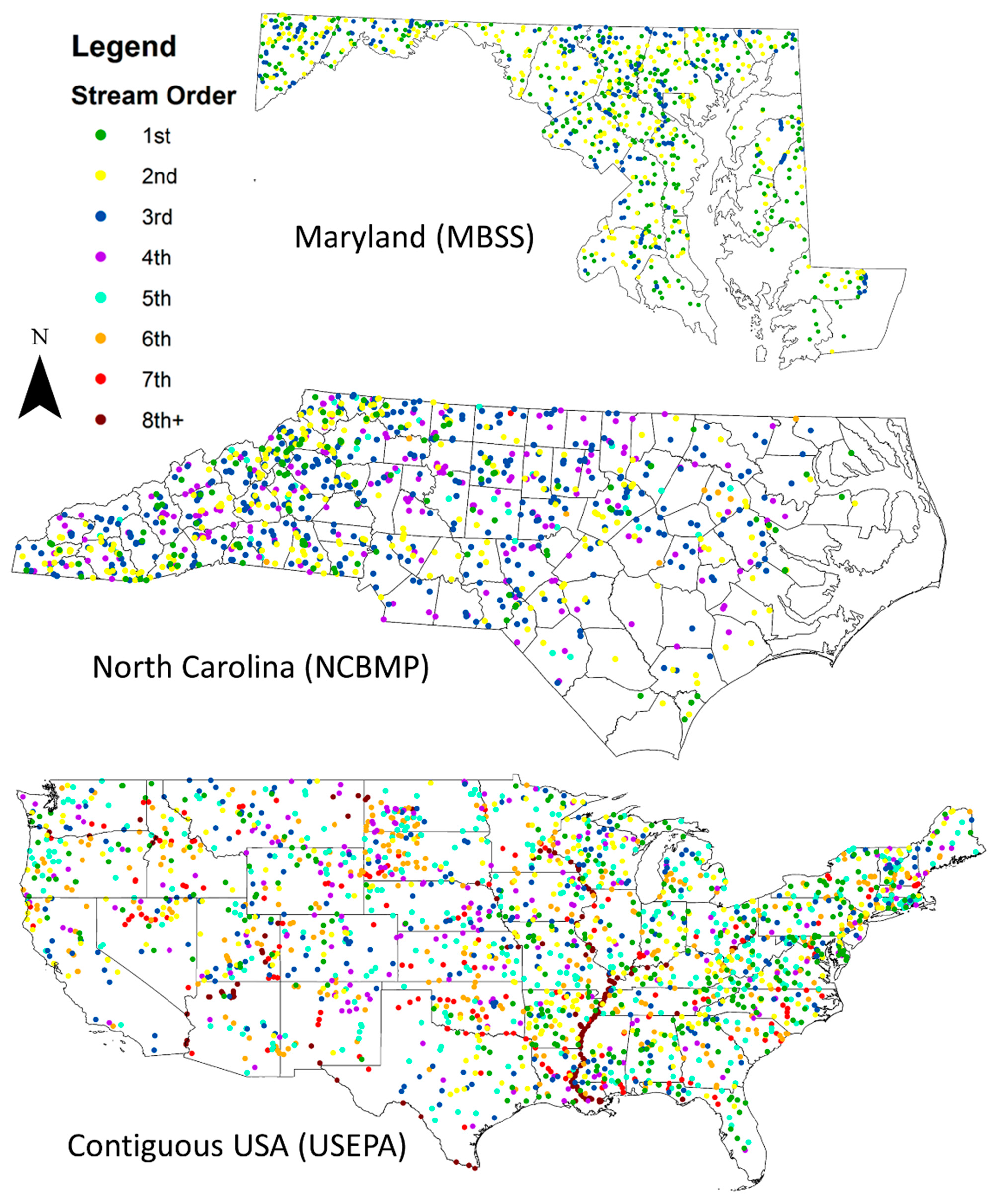
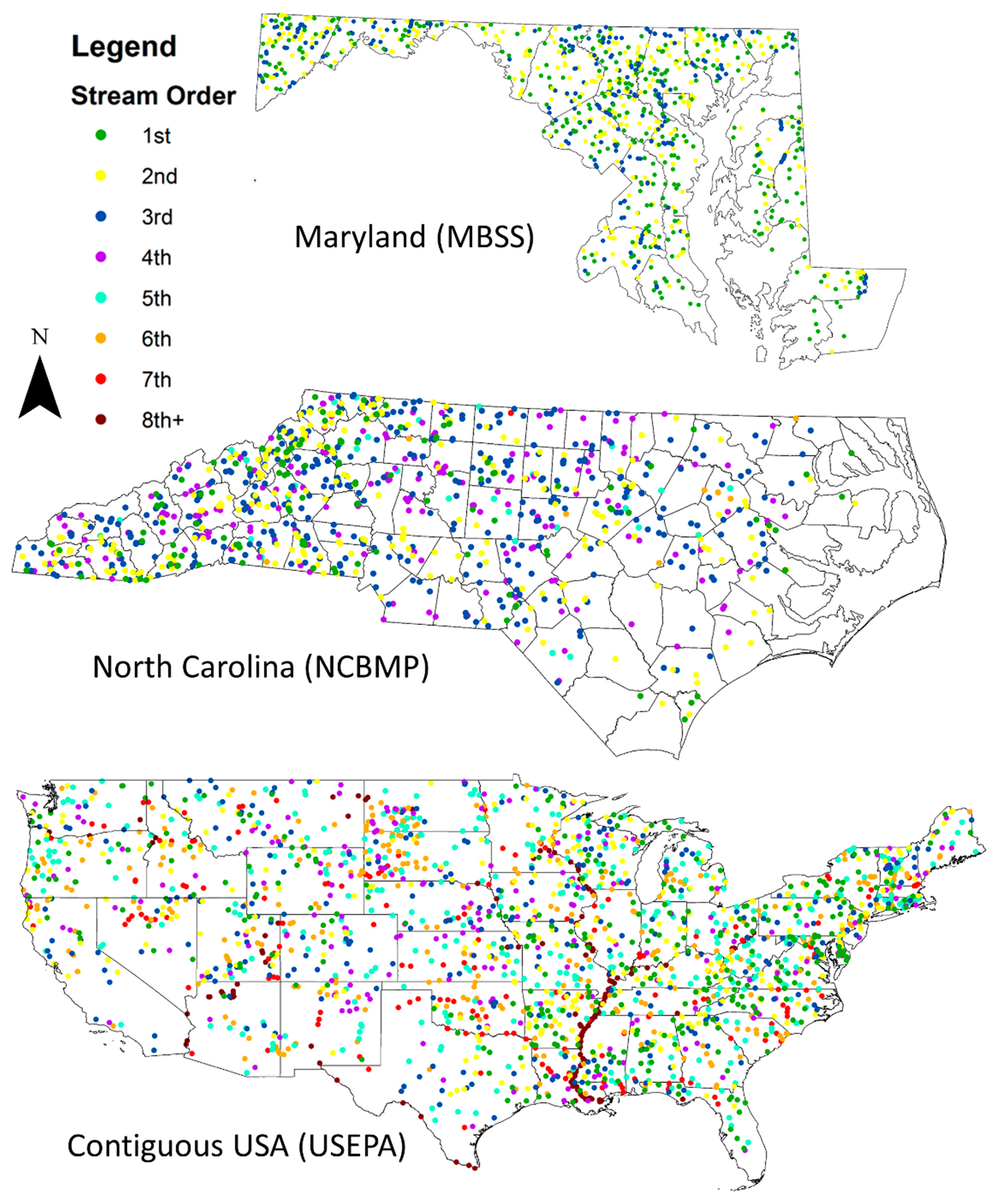
2.2.1. Datasets
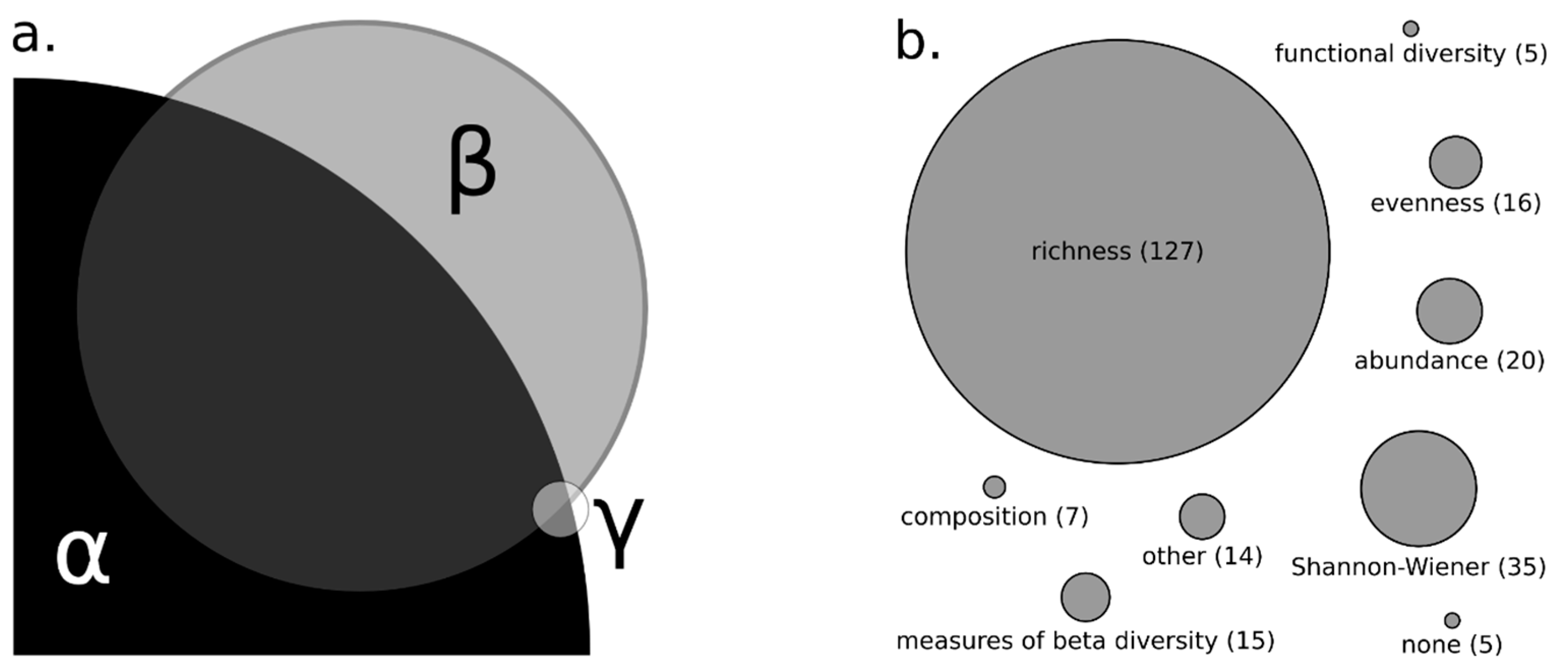
2.2.2. Trait Data and Measures of Taxonomic, Functional and Beta Diversity
2.2.3. Data Analysis
3. Results
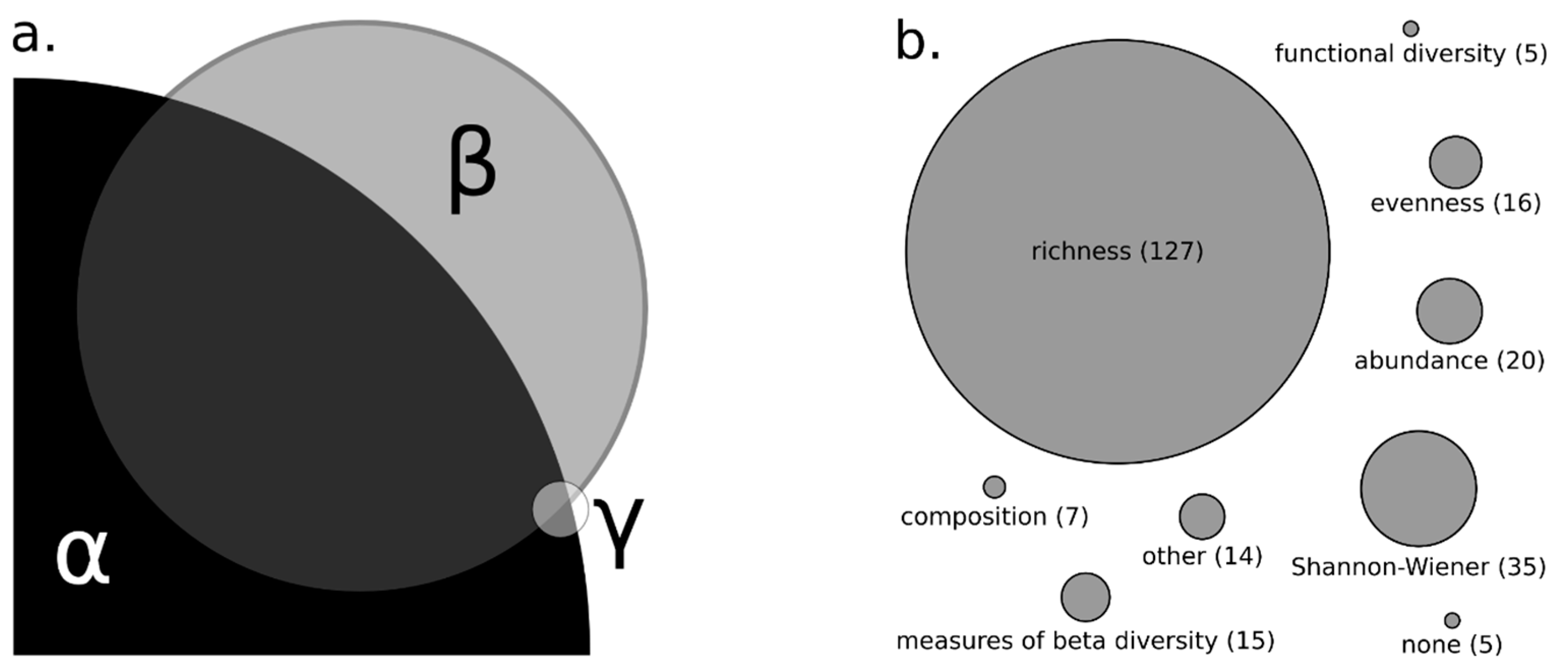
3.1. Measures of Biodiversity and Stream Size in Reviewed Literature
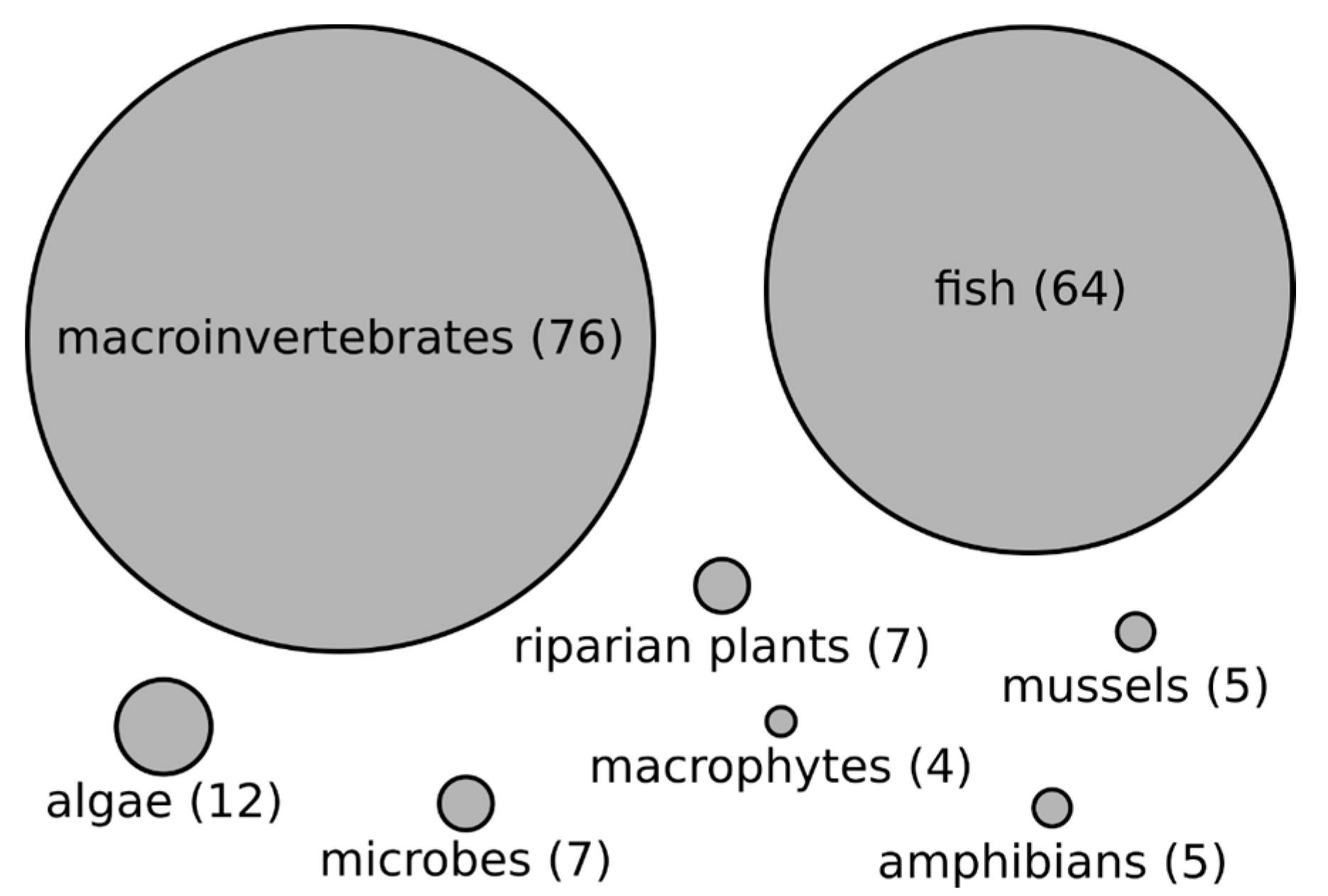
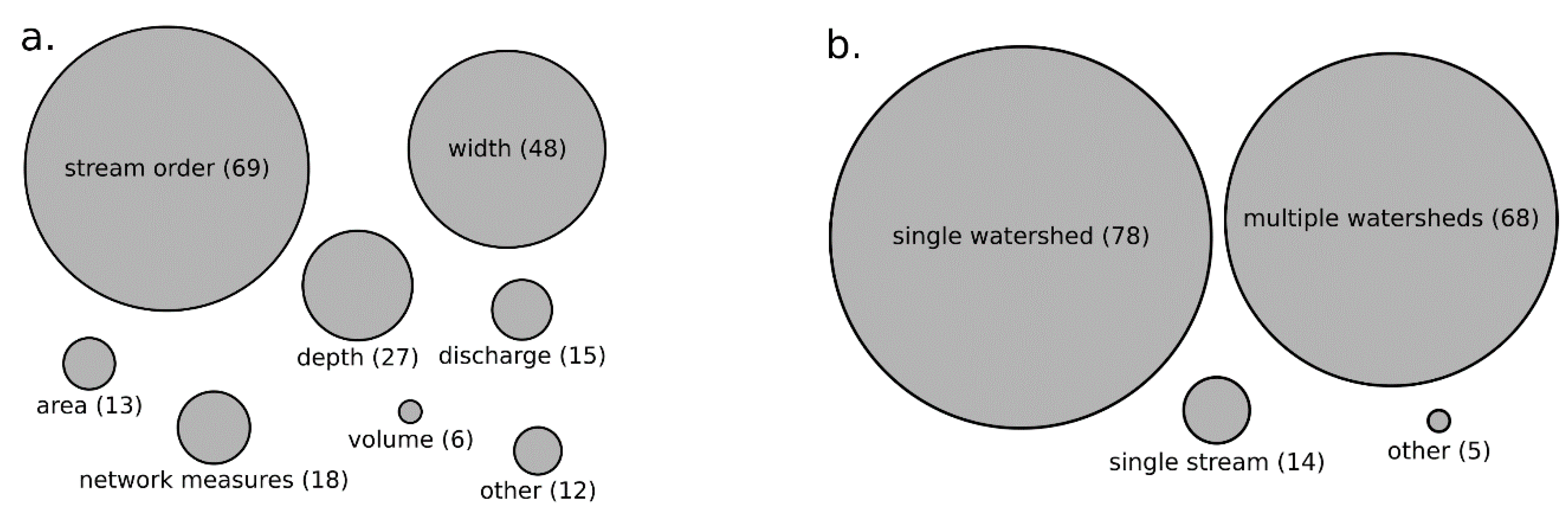
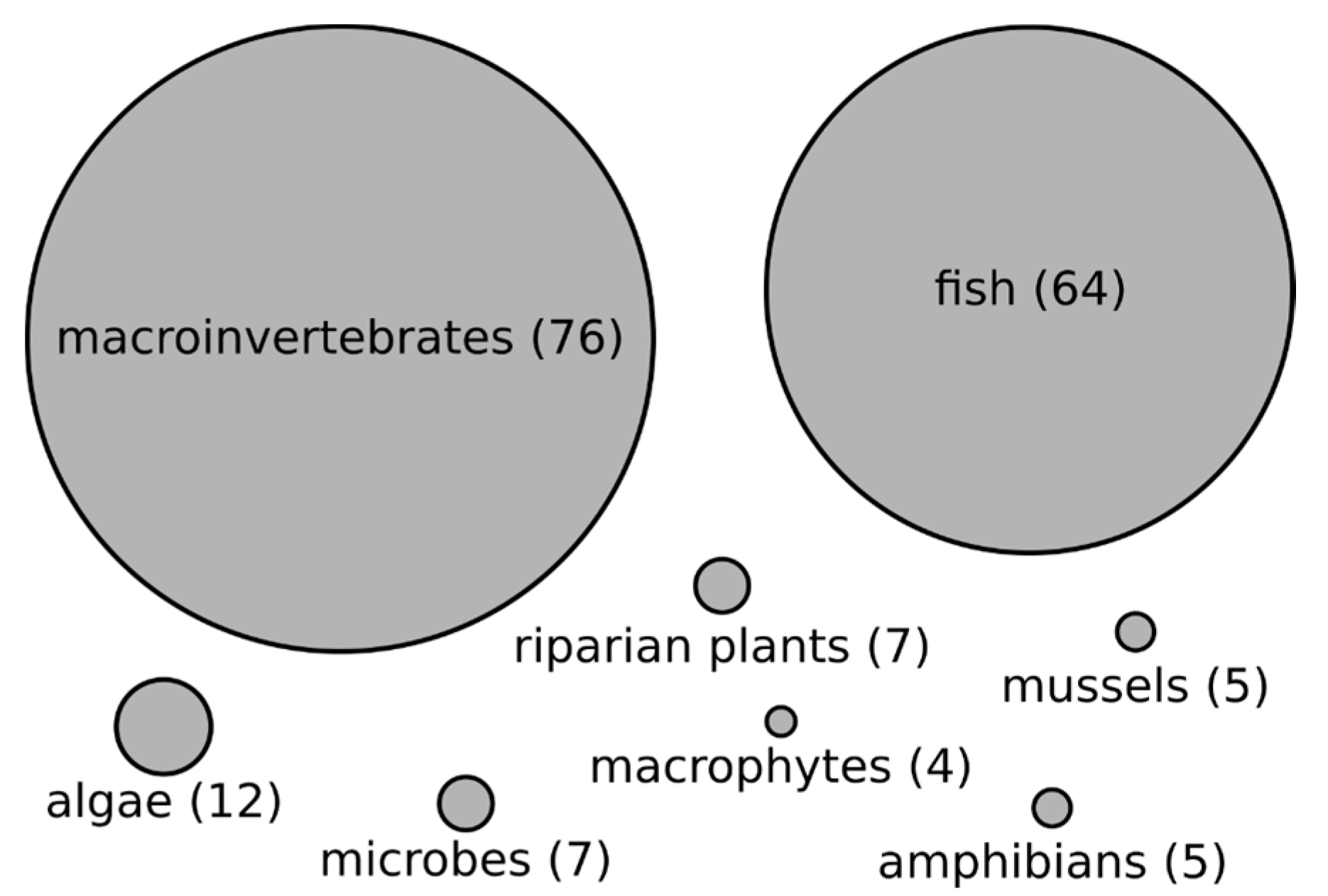
3.2. Organisms Studied and Their Relationship with Stream Size in Reviewed Literature
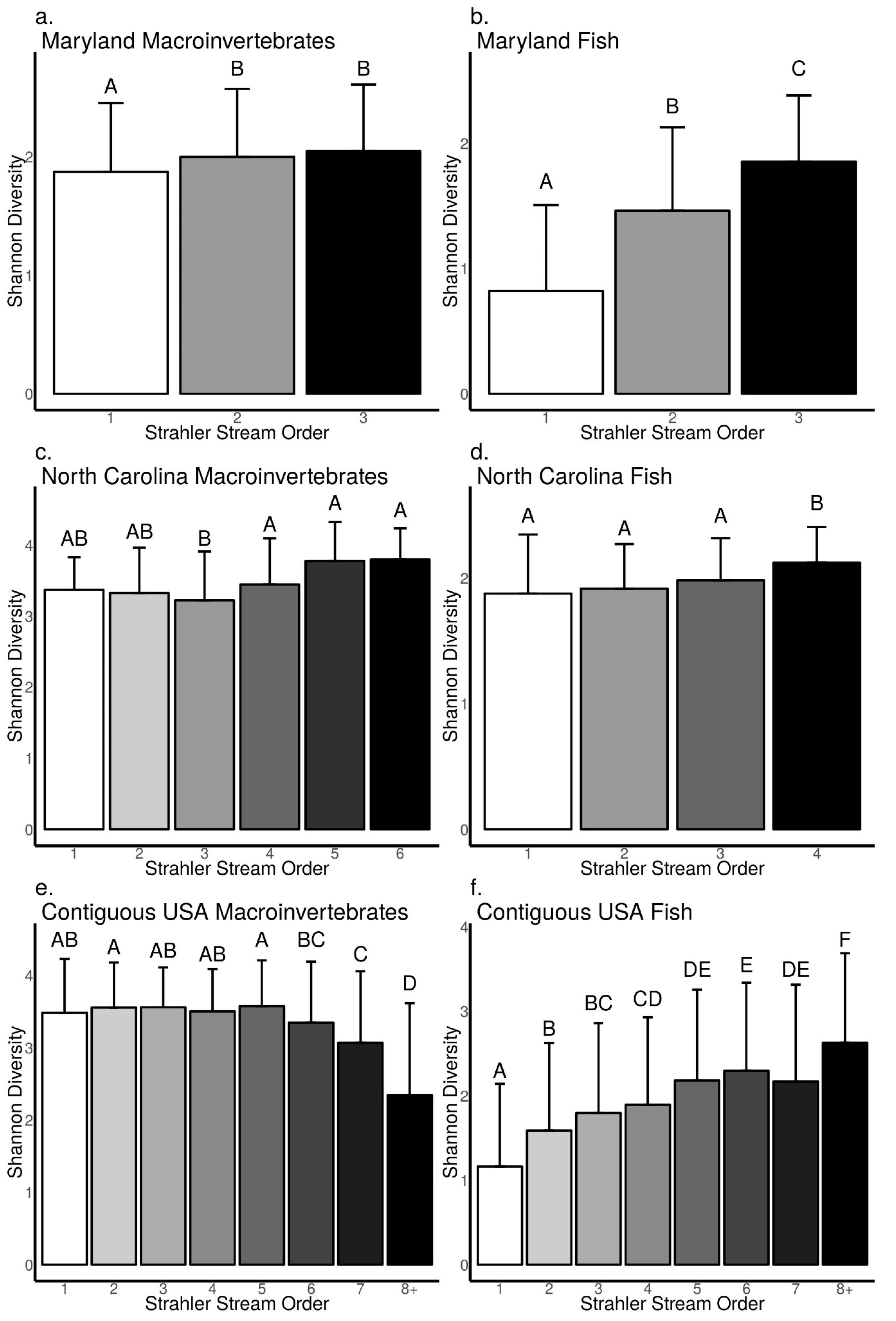
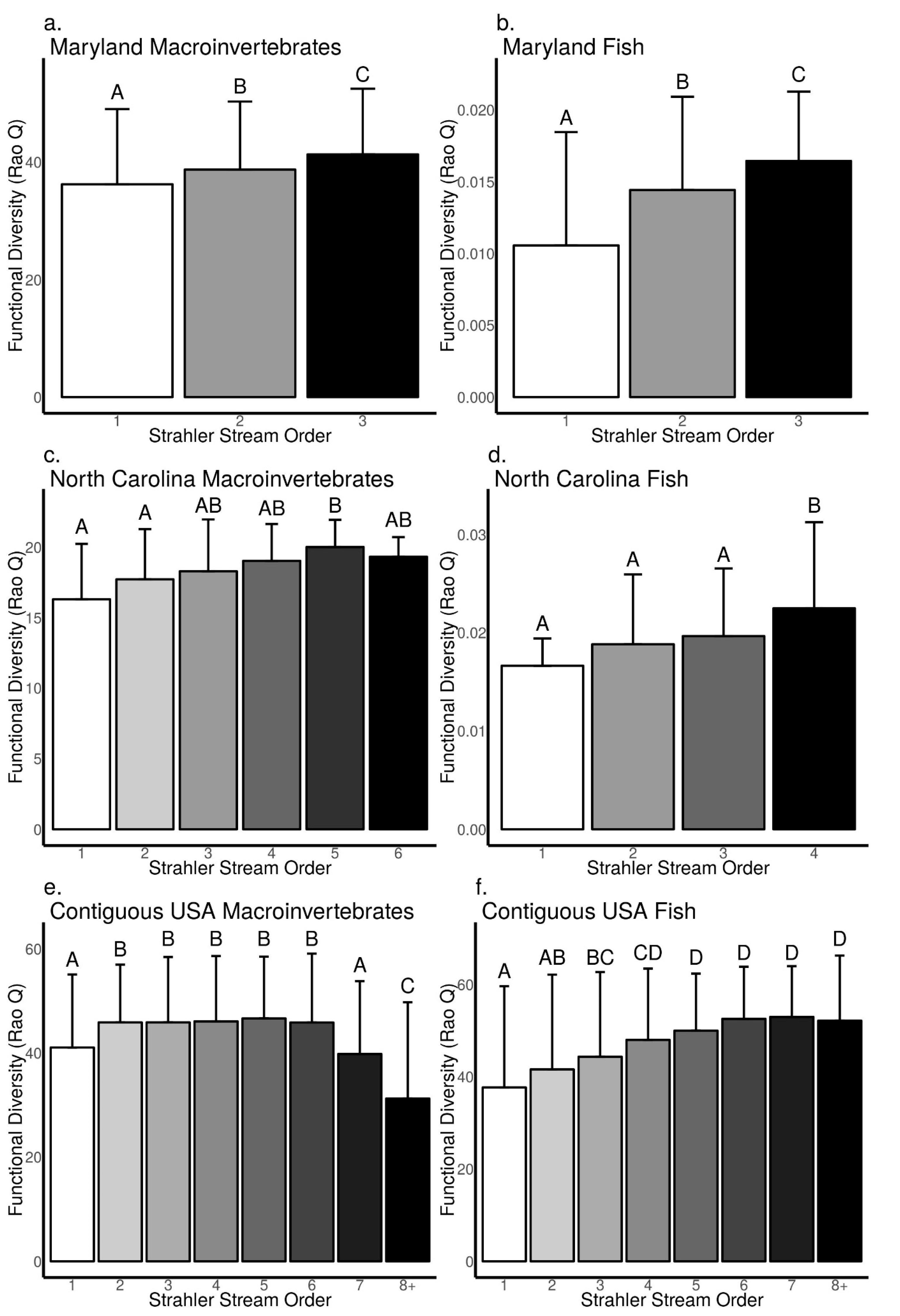
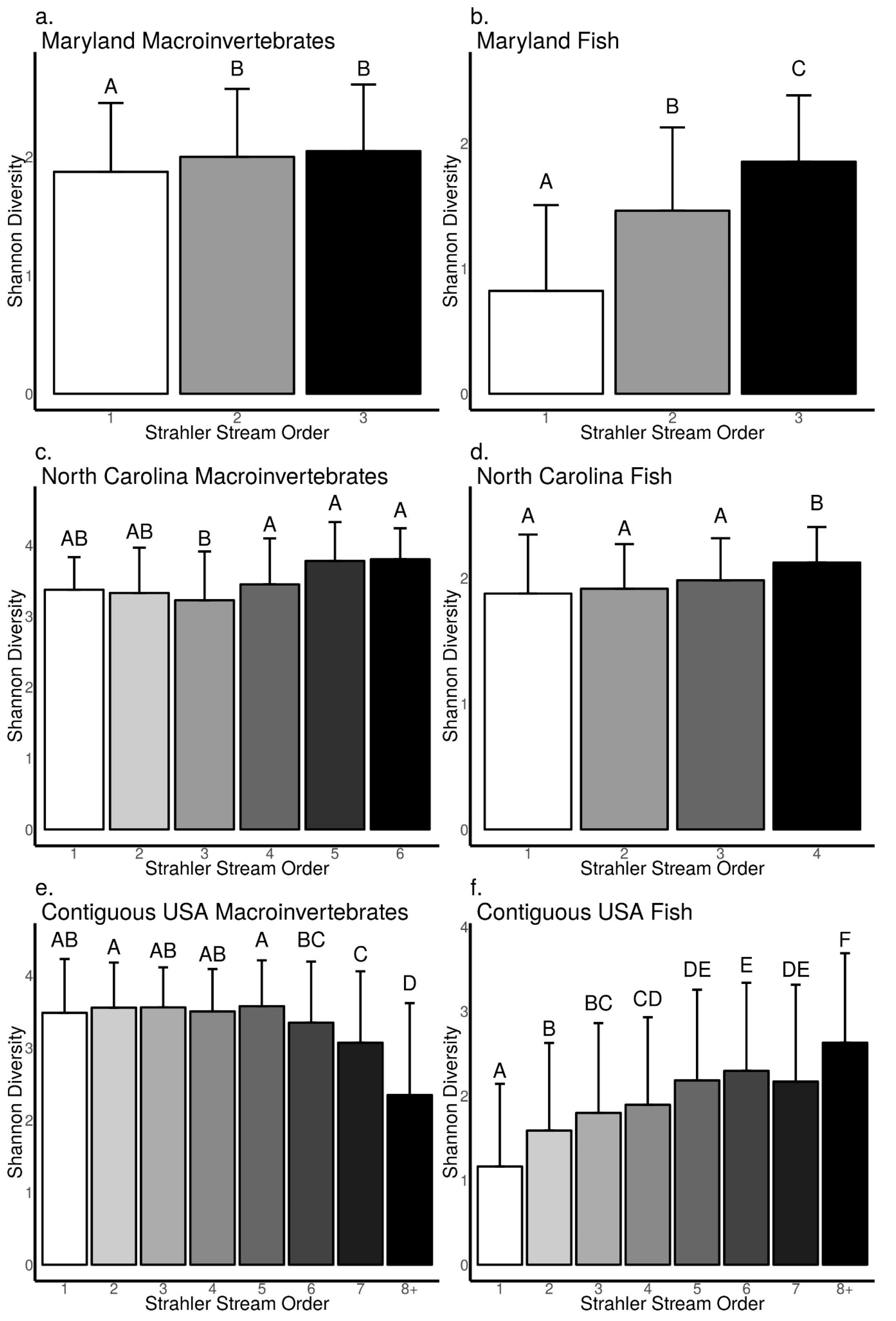
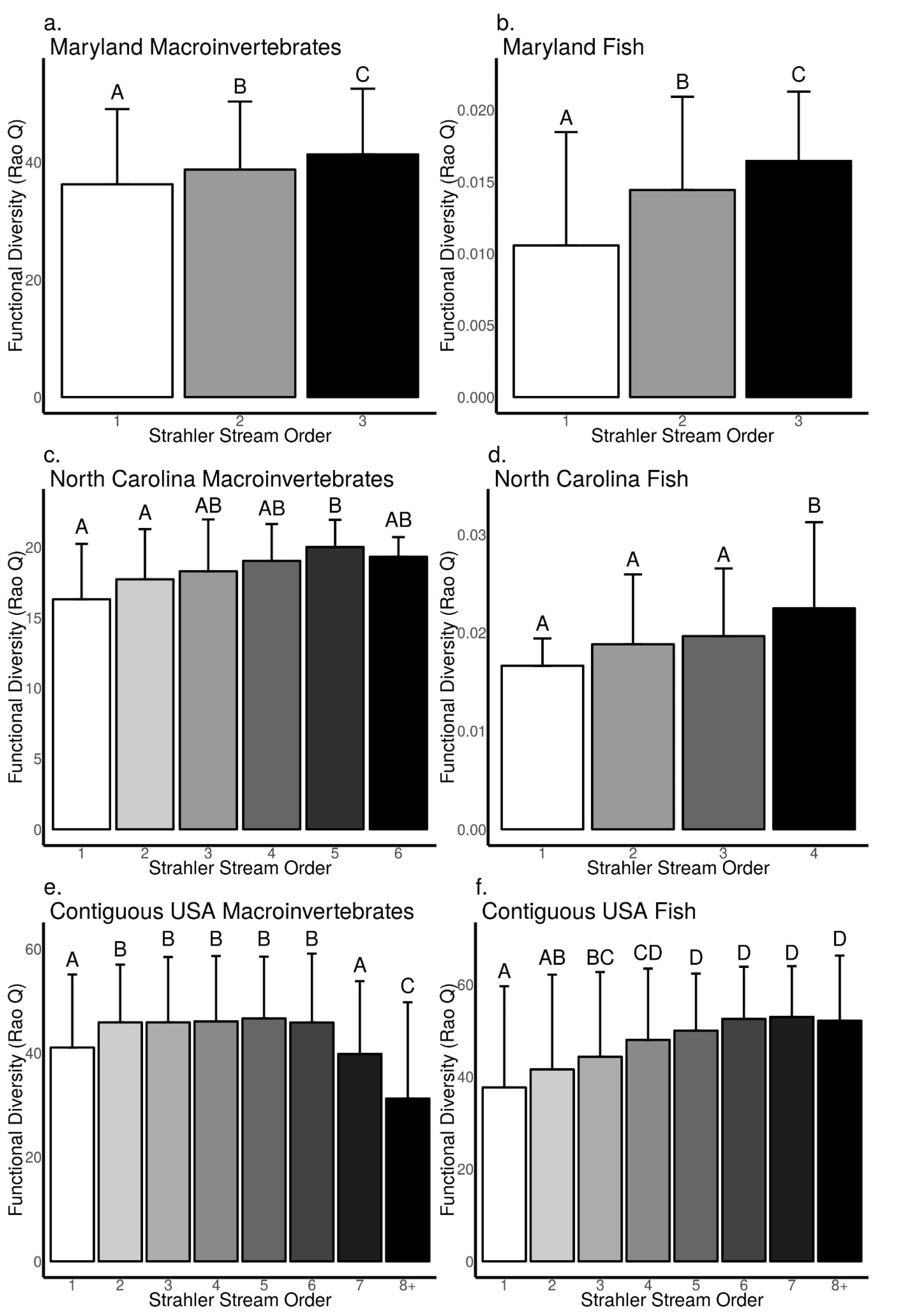
3.3. Relationships between Biodiversity and Strahler Stream Order across Sites in Maryland, North Carolina, and the Contiguous USA
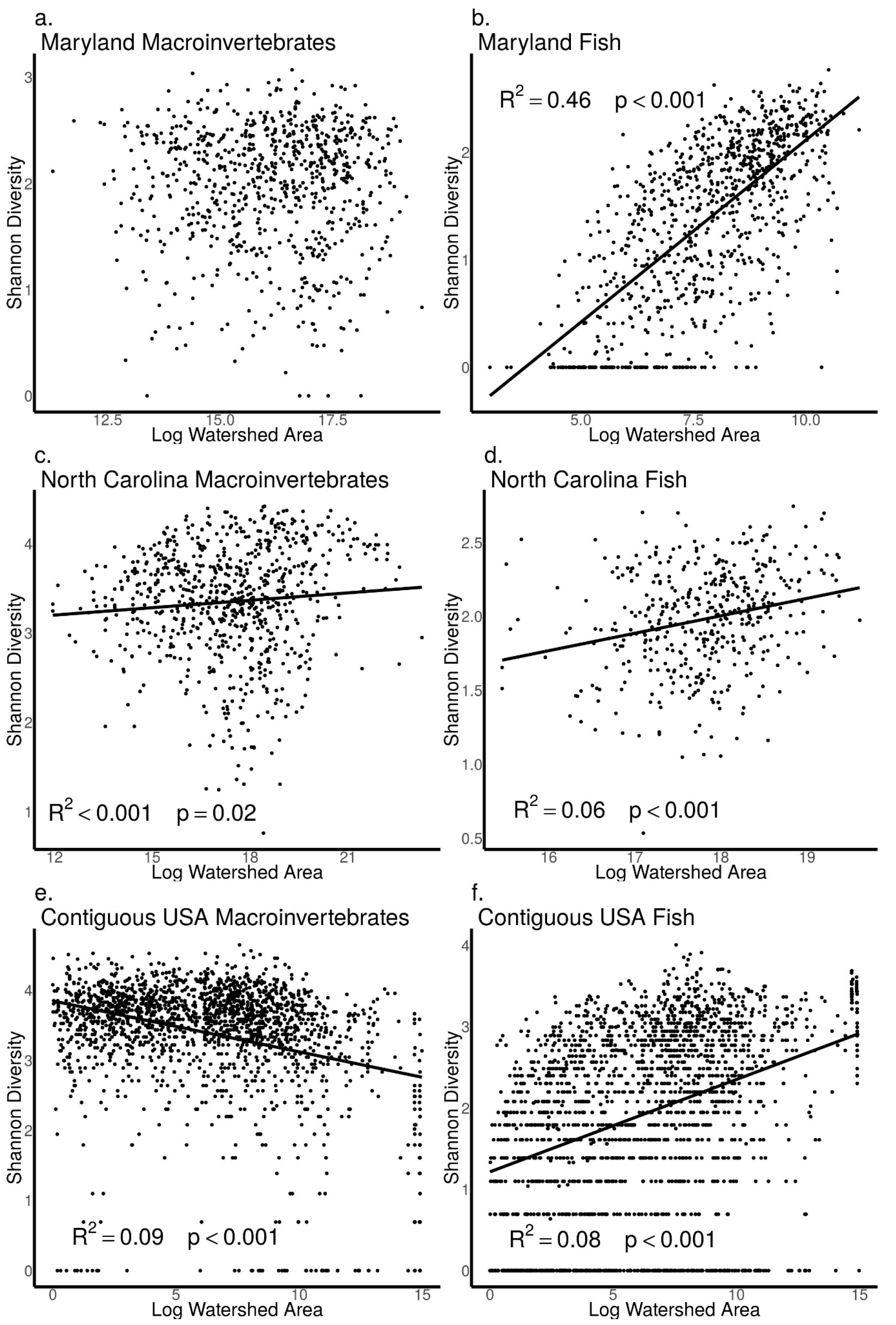
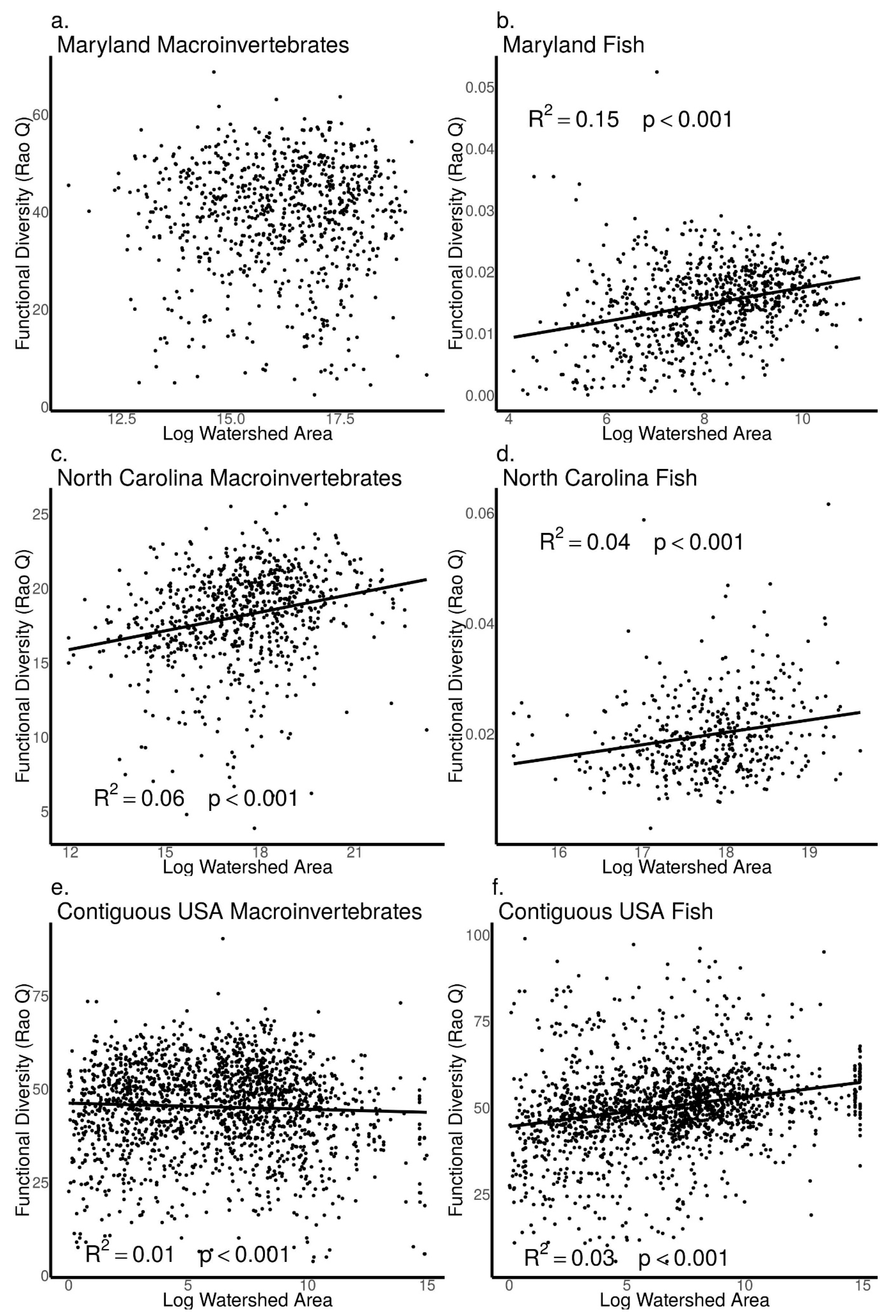
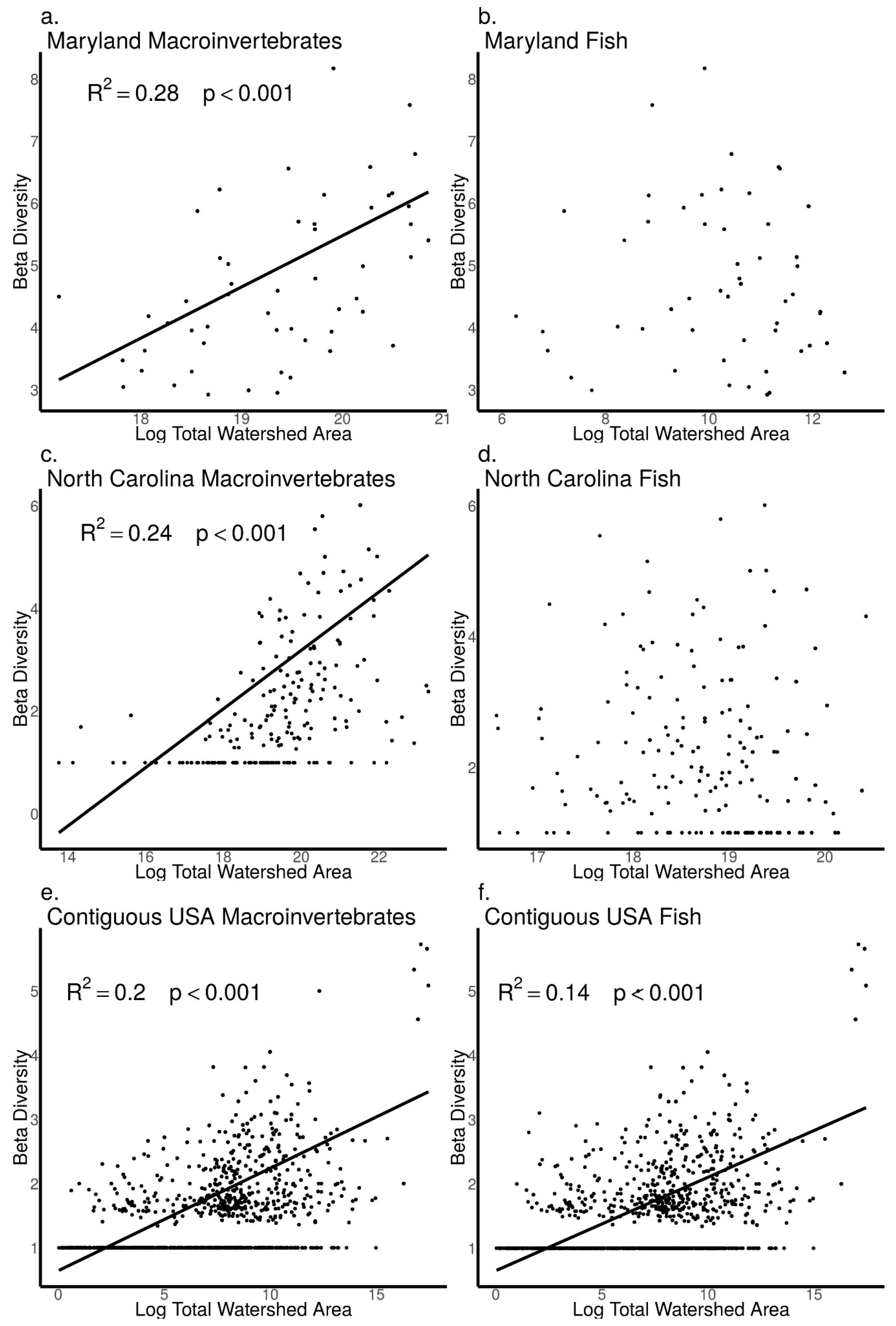
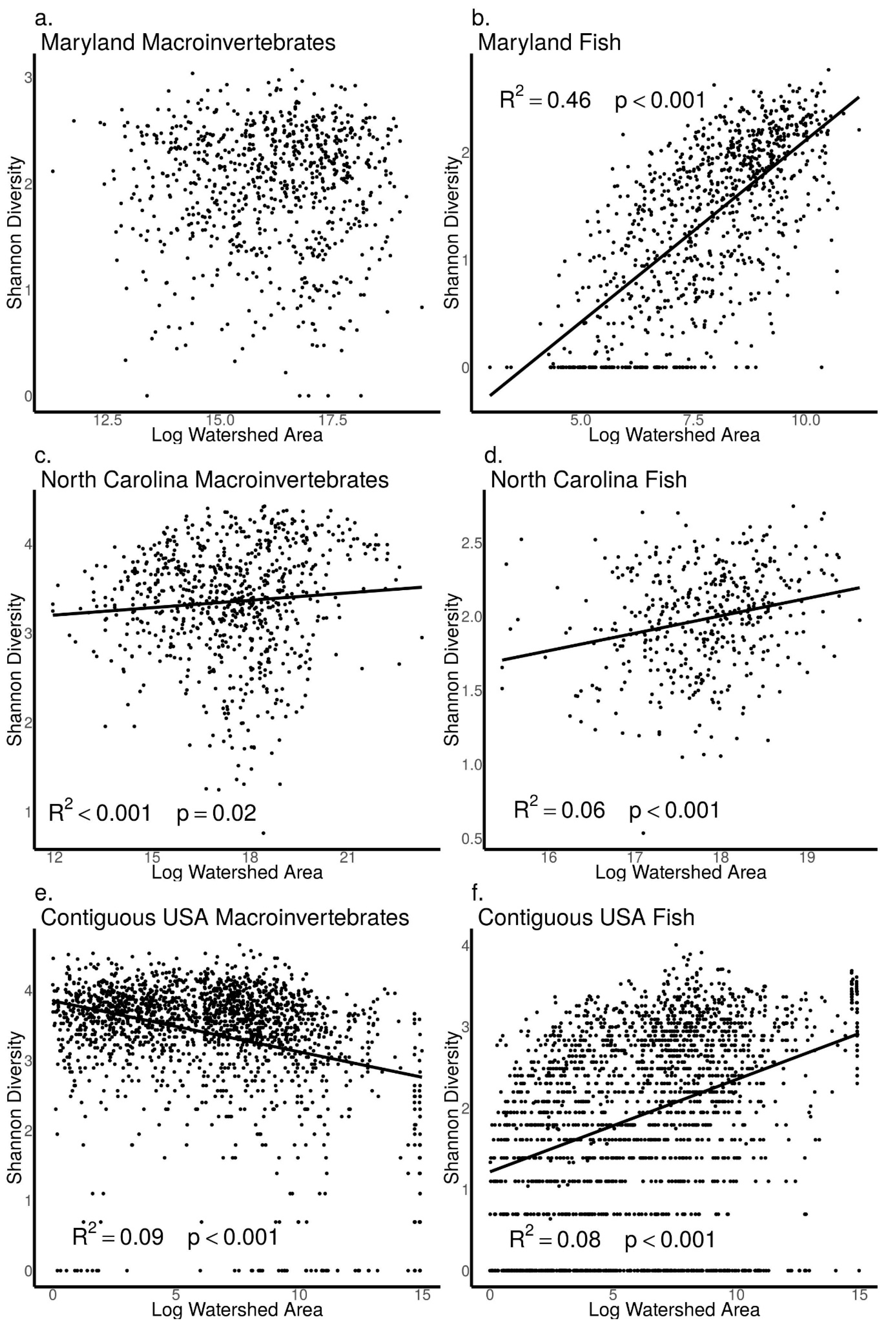
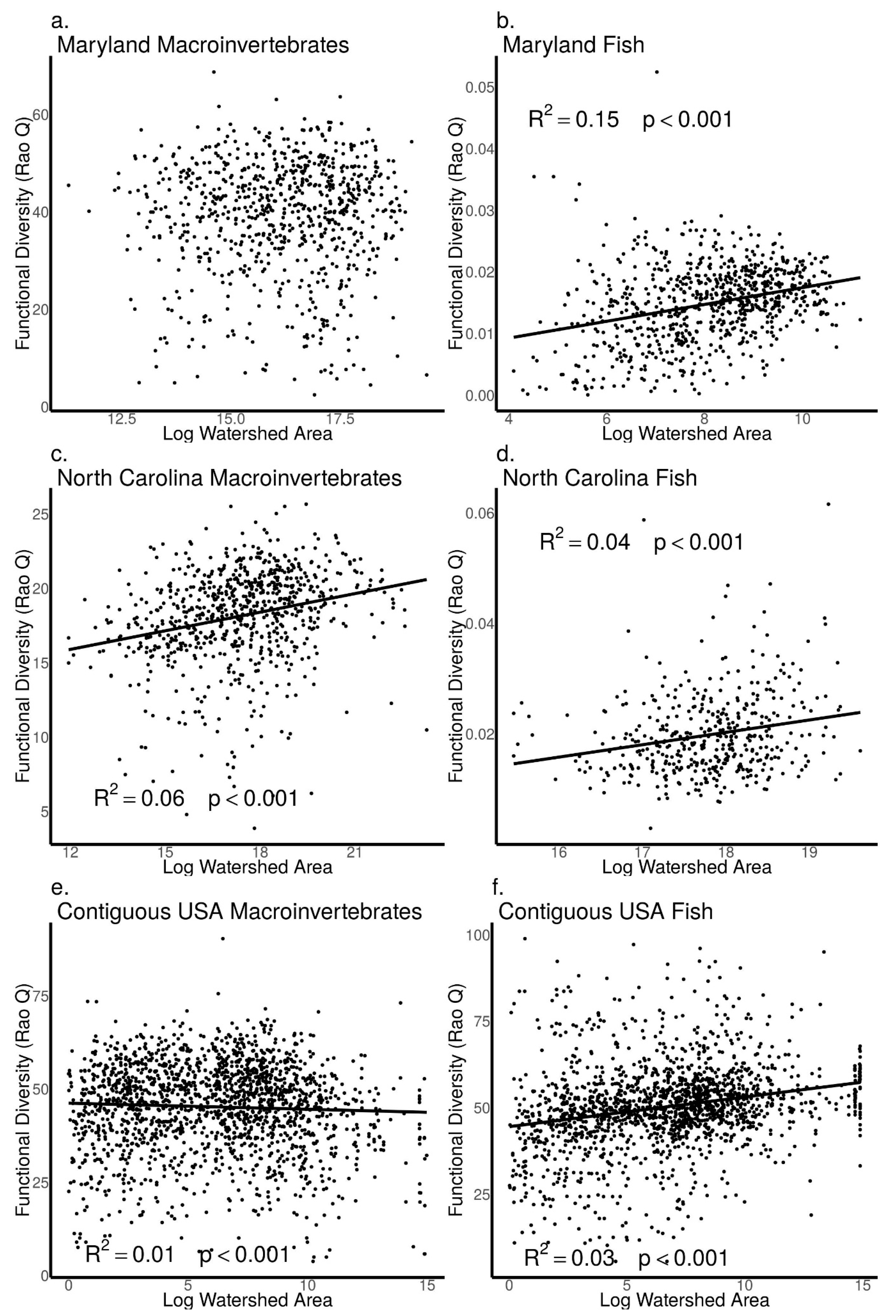
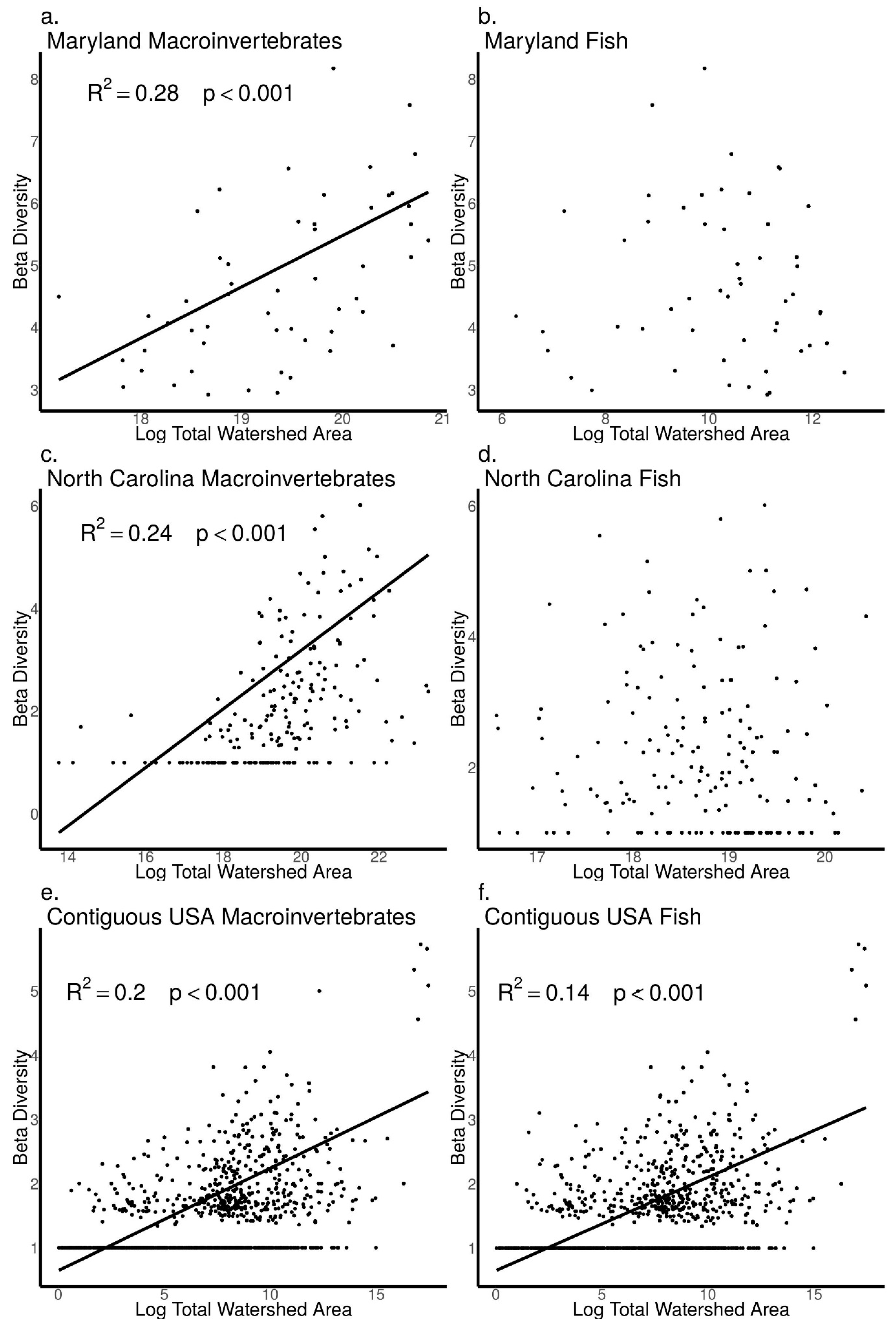
3.4. Relationships between Biodiversity and Watershed Area across Sites in Maryland, North Carolina, and the Contiguous USA
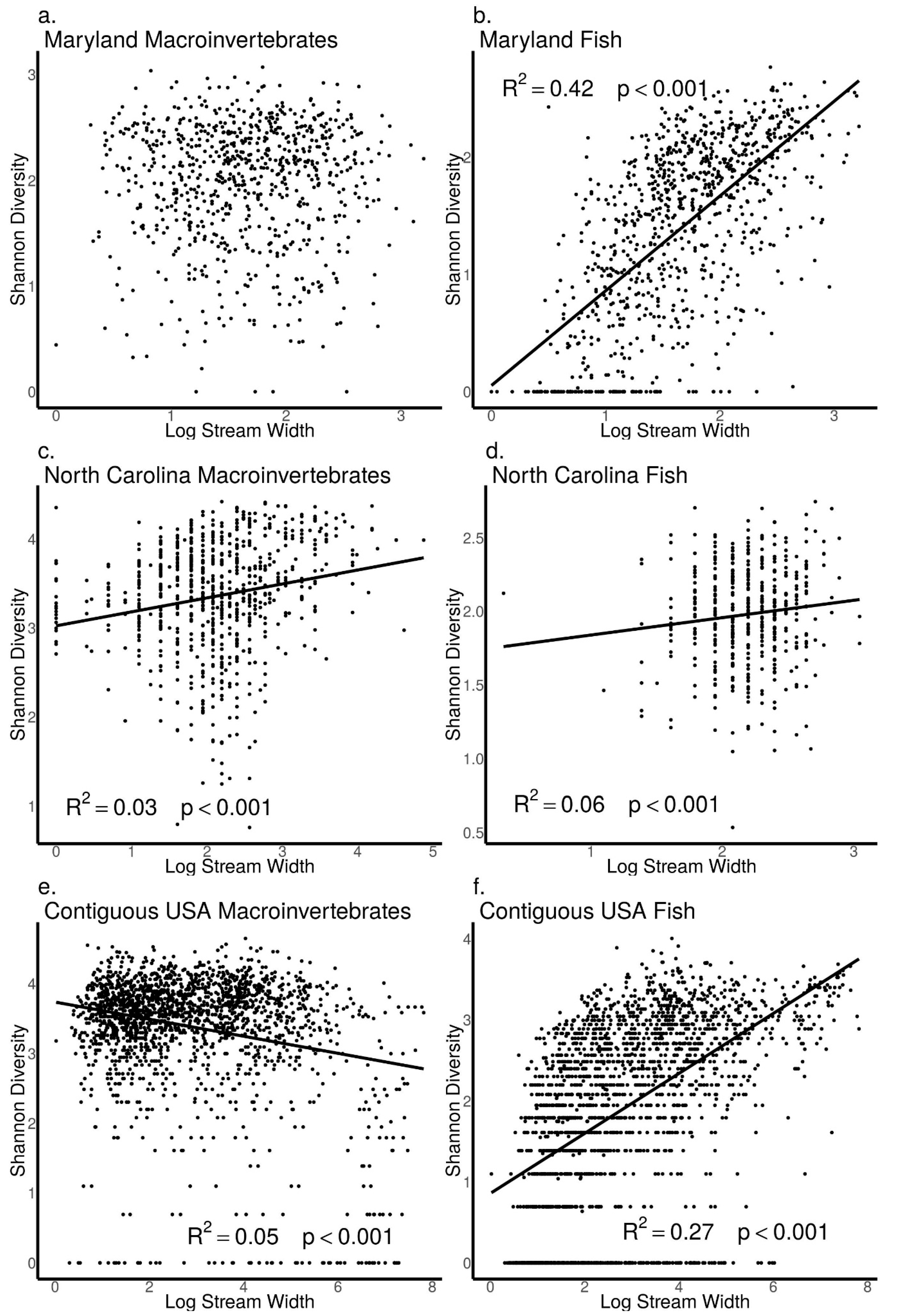
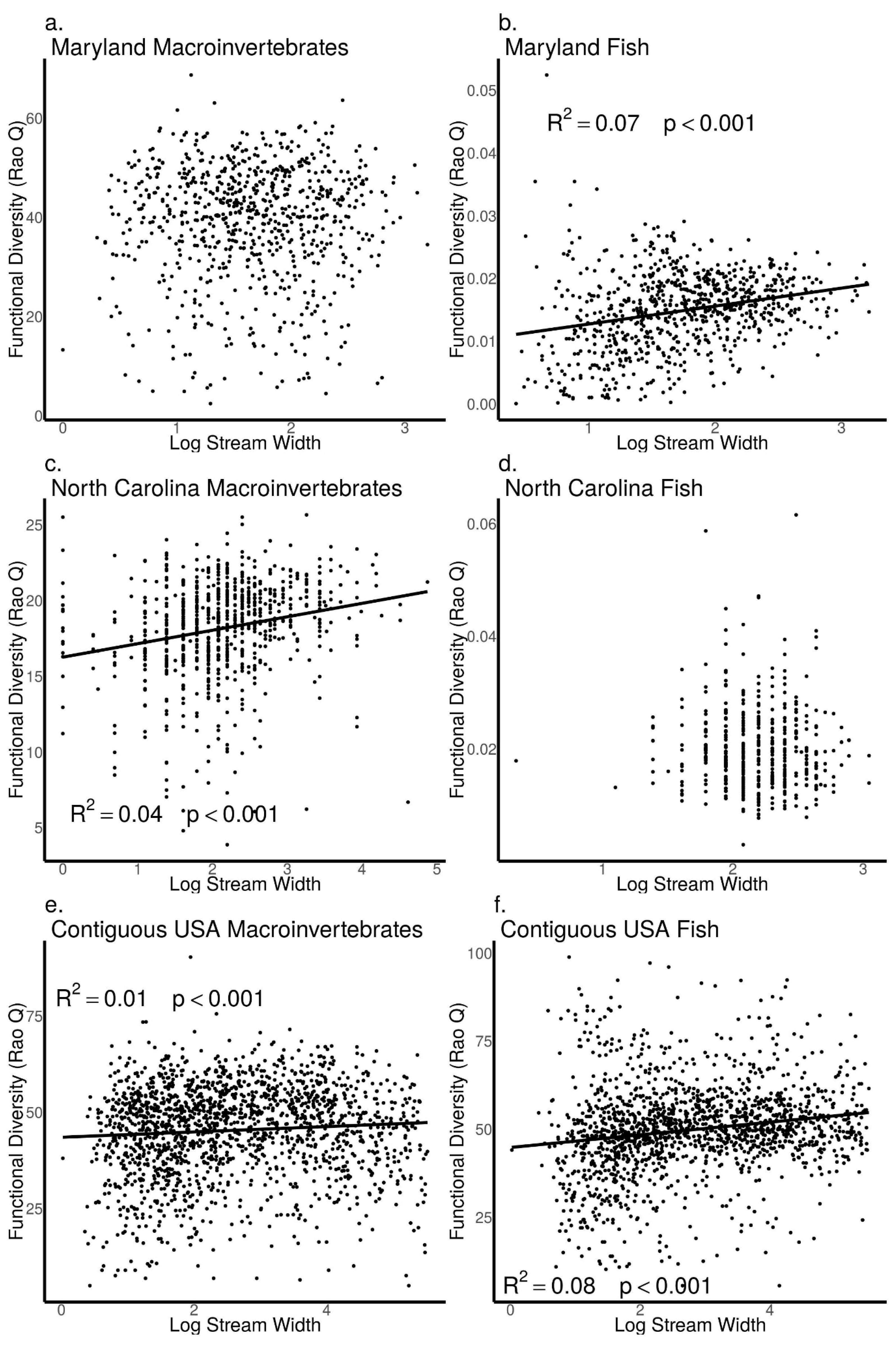
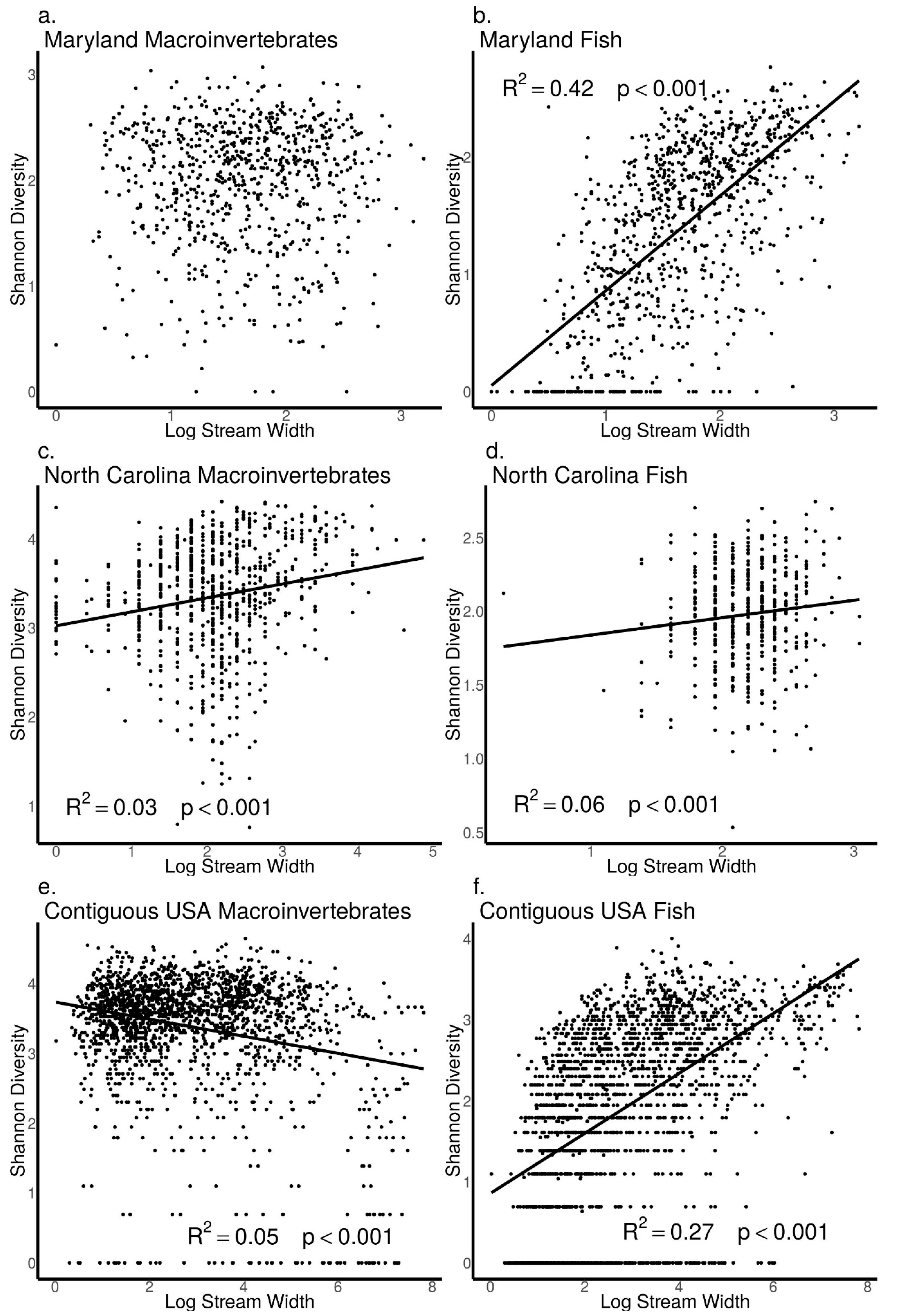
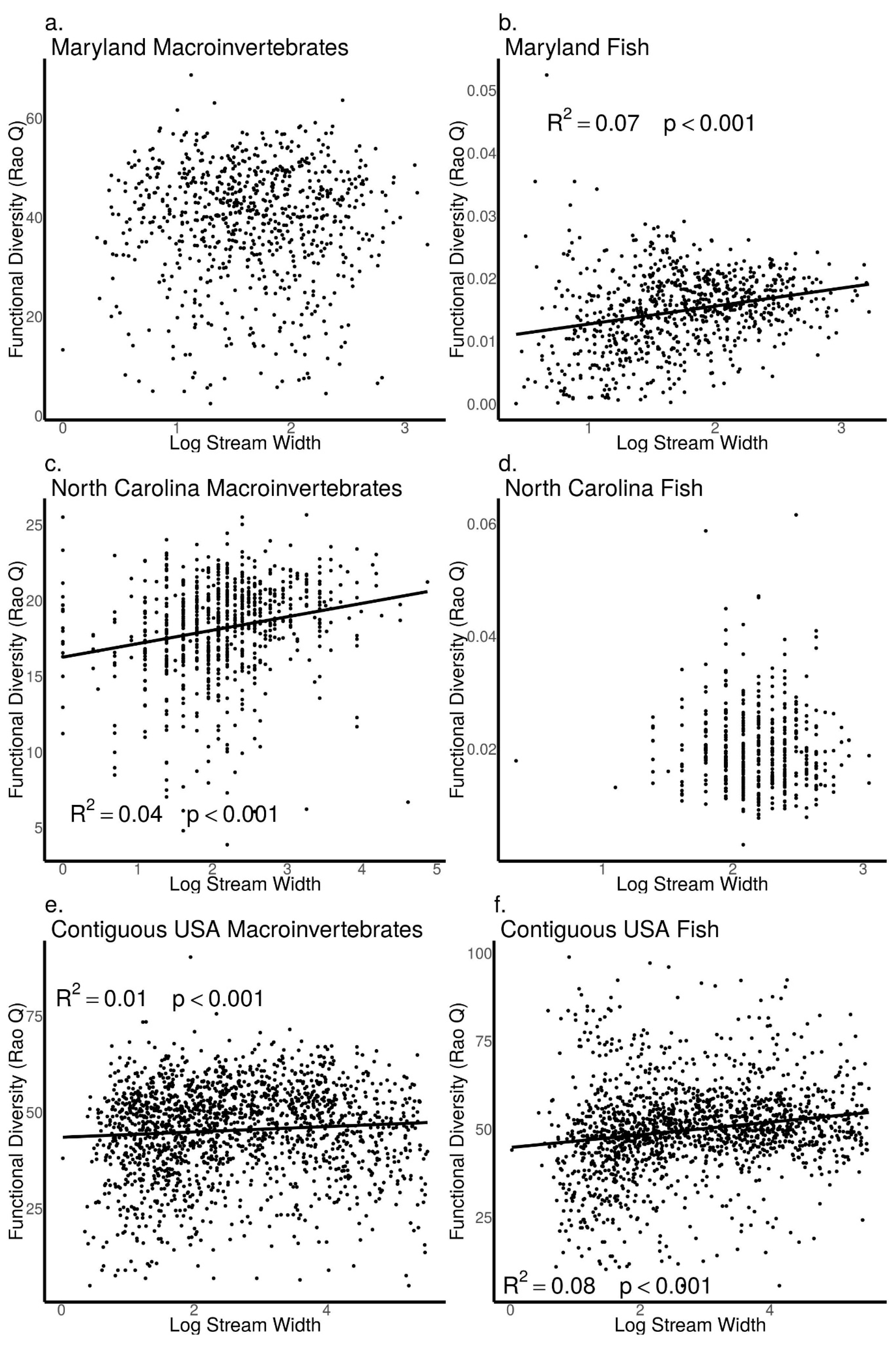
3.5. Relationships between Biodiversity and Stream Width across Sites in Maryland, North Carolina, and the Contiguous USA
4. Discussion
4.1. Patterns from Our Literature Review and Data Analysis Support Key Concepts in Riverine Ecology
4.2. Difference between Macroinvertebrates and Fish in the Frequency and Strength of Biodiversity–Stream Size Relationships
4.3. Important Lack of Explanatory Mechanisms across Behind Biodiversity–Stream Size Relationships
4.4. Management Implications
5. Conclusions
Supplementary Materials
Acknowledgments
Author Contributions
Conflicts of Interest
References
- McGill, B.J.; Enquist, B.J.; Weiher, E.; Westoby, M. Rebuilding community ecology from functional traits. Trends Ecol. Evol. 2006, 21, 178–185. [Google Scholar] [CrossRef] [PubMed]
- Prevedello, J.A.; Gotelli, N.J.; Metzger, J.P. A stochastic model for landscape patterns of biodiversity. Ecol. Monogr. 2016, 86, 462–479. [Google Scholar] [CrossRef]
- Cabello, J.; Fernández, N.; Alcaraz-Segura, D.; Oyonarte, C.; Piñeiro, G.; Altesor, A.; Delibes, M.; Paruelo, J. The ecosystem functioning dimension in conservation: Insights from remote sensing. Biodivers. Conserv. 2012, 21, 3287–3305. [Google Scholar] [CrossRef]
- Palmer, M.A.; Menninger, H.L.; Bernhardt, E. River restoration, habitat heterogeneity and biodiversity: A failure of theory or practice? Freshwater Biol. 2010, 55, 205–222. [Google Scholar] [CrossRef]
- Vorosmarty, C.J.; McIntyre, P.B.; Gessner, M.O.; Dudgeon, D.; Prusevich, A.; Green, P.; Glidden, S.; Bunn, S.E.; Sullivan, C.A.; Liermann, C.R.; et al. Global threats to human water security and river biodiversity. Nature 2010, 467, 555–561. [Google Scholar] [CrossRef] [PubMed]
- Gomi, T.; Sidle, R.C.; Richardson, J.S. Understanding processes and downstream linkages of headwater systems. BioScience 2002, 52, 905–916. [Google Scholar] [CrossRef]
- Strayer, D.L.; Dudgeon, D. Freshwater biodiversity conservation: Recent progress and future challenges. J. N. Am. Benthol. Soc. 2010, 29, 344–358. [Google Scholar] [CrossRef]
- Walther, G.-R.; Post, E.; Convey, P.; Menzel, A.; Parmesan, C.; Beebee, T.J.C.; Fromentin, J.-M.; Hoegh-Guldberg, O.; Bairlein, F. Ecological responses to recent climate change. Nature 2002, 416, 389–395. [Google Scholar] [CrossRef] [PubMed]
- Xenopoulos, M.A.; Lodge, D.M.; Alcamo, J.; Marker, M.; Schulze, K.; Van Vuuren, D.P. Scenarios of freshwater fish extinctions from climate change and water withdrawal. Glob. Chang. Biol. 2005, 11, 1557–1564. [Google Scholar] [CrossRef]
- Heino, J. The importance of metacommunity ecology for environmental assessment research in the freshwater realm. Biol. Rev. 2013, 88, 166–178. [Google Scholar] [CrossRef] [PubMed]
- Hilsenhoff, W.L. Rapid field assessment of organic pollution with a family-level biotic index. J. N. Am. Benthol. Soc. 1988, 7, 65–68. [Google Scholar] [CrossRef]
- Hynes, H.B.N. The Biology of Polluted Waters; Liverpool University Press: Liverpool, UK, 1960. [Google Scholar]
- De Jalon, G.; Mayo, D.; Molles, M.C. Characterization of spanish pyrenean stream habitat: Relationships between fish communities and their habitat. Regul. Rivers Resour. Manag. 1996, 12, 305–316. [Google Scholar] [CrossRef]
- Heino, J.; Paavola, R.; Virtanen, R.; Muotka, T. Searching for biodiversity indicators in running waters: Do bryophytes, macroinvertebrates, and fish show congruent diversity patterns? Biodivers. Conserv. 2005, 14, 415–428. [Google Scholar] [CrossRef]
- Pease, A.A.; GonzÁLez-DÍAz, A.A.; Rodiles-HernÁNdez, R.; Winemiller, K.O. Functional diversity and trait-environment relationships of stream fish assemblages in a large tropical catchment. Freshwater Biol. 2012, 57, 1060–1075. [Google Scholar] [CrossRef]
- Pease, A.A.; Taylor, J.M.; Winemiller, K.O.; King, R.S. Ecoregional, catchment, and reach-scale environmental factors shape functional-trait structure of stream fish assemblages. Hydrobiologia 2015, 753, 265–283. [Google Scholar] [CrossRef]
- Vannote, R.L.; Minshall, G.W.; Cummins, K.W.; Sedell, J.R.; Cushing, D.H. The river continuum concept. Can. J. Fish. Aquat. Sci. 1980, 37, 130–137. [Google Scholar] [CrossRef]
- Hitt, N.P.; Angermeier, P.L. Fish community and bioassessment responses to stream network position. J. N. Am. Benthol. Soc. 2011, 30, 296–309. [Google Scholar] [CrossRef]
- Rosi-Marshall, E.J.; Vallis, K.L.; Baxter, C.V.; Davis, J.M. Retesting a prediction of the river continuum concept: Autochthonous versus allochthonous resources in the diets of invertebrates. Freshwater Sci. 2016, 35, 534–543. [Google Scholar] [CrossRef]
- Thorp, J.H.; Thoms, M.C.; Delong, M.D. The riverine ecosystem synthesis: Biocomplexity in river networks across space and time. River Res. Appl. 2006, 22, 123–147. [Google Scholar] [CrossRef]
- Tornwall, B.; Sokol, E.R.; Skelton, J.; Brown, B.L. Trends in stream biodiversity research since the river continuum concept. Diversity 2015, 7, 16–35. [Google Scholar] [CrossRef]
- Webster, J.R. Spiraling down the river continuum: Stream ecology and the u-shaped curve. J. N. Am. Benthol. Soc. 2007, 26, 375–389. [Google Scholar] [CrossRef]
- Cao, Y.; Stodola, A.; Douglass, S.; Shasteen, D.; Cummings, K.; Holtrop, A. Modelling and mapping the distribution, diversity and abundance of freshwater mussels (family unionidae) in wadeable streams of illinois, USA. Freshwater Biol. 2015, 60, 1379–1397. [Google Scholar] [CrossRef]
- Heino, J.; Mykrä, H.; Muotka, T. Temporal variability of nestedness and idiosyncratic species in stream insect assemblages. Divers. Distrib. 2009, 15, 198–206. [Google Scholar] [CrossRef]
- Osborne, L.L.; Wiley, M.J. Influence of tibutary spatial position on the structure of warmwater fish communities. Can. J. Fish. Aquat.Sci. 1992, 49, 671–681. [Google Scholar] [CrossRef]
- Stenger-Kovács, C.; Tóth, L.; Tóth, F.; Hajnal, É.; Padisák, J. Stream order-dependent diversity metrics of epilithic diatom assemblages. Hydrobiologia 2014, 721, 67–75. [Google Scholar] [CrossRef]
- Fernandes, I.M.; Lourenço, L.S.; Ota, R.P.; Moreira, M.M.M.; Zawadzki, C.H. Effects of local and regional factors on the fish assemblage structure in meridional amazonian streams. Environ. Biol. Fish. 2013, 96, 837–848. [Google Scholar] [CrossRef]
- Rahel, F.J.; Hubert, W.A. Fish assemblages and habitat gradients in a rocky mountain—Great plains stream: Biotic zonation and additive patterns of community change. Trans. Am. Fish. Soc. 1991, 120, 319–332. [Google Scholar] [CrossRef]
- Angermeier, P.L.; Schlosser, I.J. Species-area relationship for stream fishes. Ecology 1989, 70, 1450–1462. [Google Scholar] [CrossRef]
- Anjos, M.B.D.; Zuanon, J. Sampling effort and fish species richness in small terra firme forest streams of central amazonia, Brazil. Neotrop. Ichthyol. 2007, 5, 45–52. [Google Scholar] [CrossRef]
- Couto, T.B.D.A.; Aquino, P.D.P.U.D. Structure and integrity of fish assemblages in streams associated to conservation units in central brazil. Neotrop. Ichthyol. 2011, 9, 445–454. [Google Scholar] [CrossRef]
- Heino, J.; Paasivirta, L. Unravelling the determinants of stream midge biodiversity in a boreal drainage basin. Freshwater Biol. 2008, 53, 884–896. [Google Scholar] [CrossRef]
- Paller, M.H. Relationships between fish assemblage structure and stream order in south carolina coastal plain streams. Trans. Am. Fish. Soc. 1994, 123, 150–161. [Google Scholar] [CrossRef]
- Yan, Y.; He, S.; Chu, L.; Xiang, X.; Jia, Y.; Toa, J.; Chen, Y. Spatial and temporal variation of fish assemblages in a subtropical small stream of the huangshan mountain. Curr. Zool. 2010, 56, 670–677. [Google Scholar]
- Desmond, J.S.; Zedler, J.B.; Williams, G.D. Fish use of tidal creek habitats in two southern california salt marshes. Ecol. Eng. 2000, 14, 233–252. [Google Scholar] [CrossRef]
- Gustafson, M.P. Effects of thermal regime on mayfly assemblages in mountain streams. Hydrobiologia 2008, 605, 235–246. [Google Scholar] [CrossRef]
- Kalaninová, D.; Bulánková, E.; Šporka, F. Caddisflies (trichoptera) as good indicators of environmental stress in mountain lotic ecosystems. Biologia 2014, 69, 1030–1045. [Google Scholar] [CrossRef]
- Grossman, G.D.; Ratajczak, R.E.; Farr, M.D.; Wagner, C.M.; Petty, J.T. Why there are fewer fish upstream. Am. Fish. Soc. Symp. 2010, 73, 63–81. [Google Scholar]
- Horwitz, R.J. Temporal variability patterns and the distributional patterns of stream fishes. Ecol. Monogr. 1978, 48, 307–321. [Google Scholar] [CrossRef]
- McGarvey, D.J.; Hughes, R.M. Longitudinal zonation of pacific northwest (USA) fish assemblages and the species-discharge relationship. Copeia 2008, 2008, 311–321. [Google Scholar] [CrossRef]
- Taylor, C.M.; Warren, M.L. Dynamics in species composition of stream fish assemblages: Environmental variability and nested subsets. Ecology 2001, 82, 2320–2330. [Google Scholar] [CrossRef]
- Clarviate Analytics. Web of Science. Available online: http://apps.webofknowledge.com/ (accessed on 7 May 2017).
- Seaber, P.R.; Kapinos, F.P.; Knapp, G.L. Hydrologic Unit Maps; United States Geological Survey Water-Supply Paper 2294; US Geological Survey: Anchorage, AK, USA, 1987.
- Poff, N.L.; Olden, J.D.; Vieira, N.K.M.; Finn, D.S.; Simmons, M.P.; Kondratieff, B.C. Functional trait niches of north american lotic insects: Traits-based ecological applications in light of phylogenetic relationships. J. N. Am. Benthol. Soc. 2006, 25, 730–755. [Google Scholar] [CrossRef]
- Frimpong, E.A.; Angermeier, P.L. Fish traits: A database of ecological and life-history traits of freshwater fishes of the united states. Fisheries 2009, 34, 487–495. [Google Scholar] [CrossRef]
- Botta-Dukát, Z. Rao’s quadratic entropy as a measure of functional diversity based on multiple traits. J. Veg. Sci. 2005, 16, 533–540. [Google Scholar] [CrossRef]
- Rao, C.R. Diversity and dissimilarity coefficients: A unified approach. Theor. Pop. Biol. 1982, 21, 24–43. [Google Scholar] [CrossRef]
- Jost, L. Partitioning diversity into independent alpha and beta components. Ecology 2007, 88, 2427–2439. [Google Scholar] [CrossRef] [PubMed]
- Laliberté, E.; Shipley, B. Fd: Measuring Functional Diversity (FD) from Multiple Traits, and Other Tools for Functional Ecology, R Package Version 1.0–11. Available online: https://rdrr.io/rforge/FD/ (accessed on 31 May 2017).
- Charney, N.; Record, S.; Charney, M.N. Package ‘Vegetarian’. Available online: https://cran.r-project.org/web/packages/vegetarian/vegetarian.pdf (accessed on 20 July 2009).
- De Mendiburu, F. Package ‘Agricolae’: Statistical Procedures for Agricultural Research, R Package Version 1.2–4. Available online: https://cran.r-project.org/web/packages/agricolae/index.html (accessed on 12 June 2016).
- Townsend, C.R. The patch dynamics concept of stream community ecology. J. N. Am. Benthol. Soc. 1989, 8, 36–50. [Google Scholar] [CrossRef]
- Minshall, G.W.; Brock, J.T.; Lapoint, T.W. Characterization and dynamics of benthic organic-matter and invertebrate functional feeding group relationships in the upper salmon river, idaho (USA). Int. Rev. Gesamten Hydrobiol. 1982, 67, 793–820. [Google Scholar]
- Hubackova, L.; Radkova, V.; Bojkova, J.; Syrovatka, V.; Polaskova, V.; Schenkova, J.; Horsak, M. Diversity patterns of aquatic specialists and generalists: Contrasts among two spring-fen mesohabitats and nearby streams. Biologia 2016, 71, 678–687. [Google Scholar] [CrossRef]
- Heino, J. Functional biodiversity of macroinvertebrate assemblages along major ecological gradients of boreal headwater streams. Freshwater Biol. 2005, 50, 1578–1587. [Google Scholar] [CrossRef]
- Arscott, D.B.; Tockner, K.; Ward, J. Thermal heterogeneity along a braided floodplain river (tagliamento river, northeastern italy). Can. J. Fish. Aquat.Sci. 2001, 58, 2359–2373. [Google Scholar] [CrossRef]
- Brown, B.L.; Swan, C.M.; Auerbach, D.A.; Grant, E.H.C.; Hitt, N.P.; Maloney, K.O.; Patrick, C. Metacommunity theory as a multispecies, multiscale framework for studying the influence of river network structure on riverine communities and ecosystems. J. N. Am. Benthol. Soc. 2011, 30, 310–327. [Google Scholar] [CrossRef]
- Grönroos, M.; Heino, J.; Siqueira, T.; Landeiro, V.L.; Kotanen, J.; Bini, L.M. Metacommunity structuring in stream networks: Roles of dispersal mode, distance type, and regional environmental context. Ecol. Evol. 2013, 3, 4473–4487. [Google Scholar] [CrossRef] [PubMed]
- Bilton, D.T.; Freeland, J.R.; Okamura, B. Dispersal in freshwater invertebrates. Annu. Rev. Ecol. Syst. 2001, 32, 159–181. [Google Scholar] [CrossRef]
- Mackay, R.J. Colonization by lotic macroinvertebrates: A review of processes and patterns. Can. J. Fish. Aquat. Sci. 1992, 49, 617–628. [Google Scholar] [CrossRef]
- Tonkin, J.D.; Stoll, S.; Sundermann, A.; Haase, P. Dispersal distance and the pool of taxa, but not barriers, determine the colonisation of restored river reaches by benthic invertebrates. Freshwater Biol. 2014, 59, 1843–1855. [Google Scholar] [CrossRef]
- Vander Vorste, R.; Malard, F.; Datry, T. Is drift the primary process promoting the resilience of river invertebrate communities? A manipulative field experiment in an intermittent alluvial river. Freshwater Biol. 2016, 61, 1276–1292. [Google Scholar] [CrossRef]
- Stoffels, R.J.; Clarke, K.R.; Linklater, D.S. Temporal dynamics of a local fish community are strongly affected by immigration from the surrounding metacommunity. Ecol. Evol. 2015, 5, 200–212. [Google Scholar] [CrossRef] [PubMed]
- Campbell, R.E.; McIntosh, A.R. Do isolation and local habitat jointly limit the structure of stream invertebrate assemblages? Freshwater Biol. 2013, 58, 128–141. [Google Scholar] [CrossRef]
- McHugh, P.A.; Thompson, R.M.; Greig, H.S.; Warburton, H.J.; McIntosh, A.R. Habitat size influences food web structure in drying streams. Ecography 2015, 38, 700–712. [Google Scholar] [CrossRef]
- Xenopoulos, M.A.; Lodge, D.M. Going with the flow: Using species-discharge relationships to forecast losses in fish biodiversity. Ecology 2006, 87, 1907–1914. [Google Scholar] [CrossRef]
- Al-Shami, S.A.; Heino, J.; Che Salmah, M.R.; Abu Hassan, A.; Suhaila, A.H.; Madrus, M.R. Drivers of beta diversity of macroinvertebrate communities in tropical forest streams. Freshwater Biol. 2013, 58, 1126–1137. [Google Scholar] [CrossRef]
- Kaelin, K.; Altermatt, F. Landscape-level predictions of diversity in river networks reveal opposing patterns for different groups of macroinvertebrates. Aquat. Ecol. 2016, 50, 283–295. [Google Scholar] [CrossRef]
- Fierer, N.; Leff, J.W.; Adams, B.J.; Nielsen, U.N.; Bates, S.T.; Lauber, C.L.; Owens, S.; Gilbert, J.A.; Wall, D.H.; Caporaso, J.G. Cross-biome metagenomic analyses of soil microbial communities and their functional attributes. Proc. Natl. Acad. Sci. USA 2012, 109, 21390–21395. [Google Scholar] [CrossRef] [PubMed]
- Louhi, P.; Mykrä, H.; Paavola, R.; Huusko, A.; Vehanen, T.; Mäki-Petäys, A.; Muotka, T. Twenty years of stream restoration in finland: Little response by benthic macroinvertebrate communities. Ecol. Appl. 2011, 21, 1950–1961. [Google Scholar] [CrossRef] [PubMed]
- Sundermann, A.; Stoll, S.; Haase, P. River restoration success depends on the species pool of the immediate surroundings. Ecol. Appl. 2011, 21, 1962–1971. [Google Scholar] [CrossRef] [PubMed]
- Lancaster, J.; Downes, B.J. A landscape-scale field experiment reveals the importance of dispersal in a resource-limited metacommunity. Ecology 2017, 98, 565–575. [Google Scholar] [CrossRef] [PubMed]

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Field | Search Term |
|---|---|
| Topic | “stream size” AND biodiversity |
| Topic | “stream order” AND diversity |
| Topic | diversity along the river continuum + “river continuum” AND diversity |
| Topic | “river size” AND diversity |
| Title | (network AND diversity) AND (river OR stream) |
| Title | headwater AND diversity |
© 2017 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Vander Vorste, R.; McElmurray, P.; Bell, S.; Eliason, K.M.; Brown, B.L. Does Stream Size Really Explain Biodiversity Patterns in Lotic Systems? A Call for Mechanistic Explanations. Diversity 2017, 9, 26. https://doi.org/10.3390/d9030026
Vander Vorste R, McElmurray P, Bell S, Eliason KM, Brown BL. Does Stream Size Really Explain Biodiversity Patterns in Lotic Systems? A Call for Mechanistic Explanations. Diversity. 2017; 9(3):26. https://doi.org/10.3390/d9030026
Chicago/Turabian StyleVander Vorste, Ross, Philip McElmurray, Spencer Bell, Kevin M. Eliason, and Bryan L. Brown. 2017. "Does Stream Size Really Explain Biodiversity Patterns in Lotic Systems? A Call for Mechanistic Explanations" Diversity 9, no. 3: 26. https://doi.org/10.3390/d9030026
APA StyleVander Vorste, R., McElmurray, P., Bell, S., Eliason, K. M., & Brown, B. L. (2017). Does Stream Size Really Explain Biodiversity Patterns in Lotic Systems? A Call for Mechanistic Explanations. Diversity, 9(3), 26. https://doi.org/10.3390/d9030026