Abstract
The theory of island biogeography has revolutionised the study of island biology stimulating considerable debate and leading to the development of new advances in related areas. One criticism of the theory is that it is ‘true but trivial’, i.e., on the basis of analyses of annual turnovers of organisms on islands, it has been posited that stochastic turnover mainly comprises rare species, or repeated immigrations and extinctions thereof, and thereby contribute little to the overall ecological dynamics. Here, both the absolute and relative turnover of breeding land birds are analysed for populations on Skokholm, Wales, over census intervals of 1, 3, 6, 12, and 24 years. As expected, over short census intervals (≤6 years), much of the turnover comprised repeated colonisations and extinctions of rare species. However, at longer intervals (12 and 24 years), a sizeable minority of species (11% of the total recorded) showed evidence of colonisation and/or extinction events despite sizeable populations (some upwards of 50 pairs). These results suggest that a longer-term view is required to take into account turnover involving more common species.
1. Introduction
The MacArthur & Wilson (1967) theory of island biogeography [1] (hereafter ‘ETIB’) attempted to explain patterns of species richness in discrete areas such as oceanic islands (and fragments of habitat in landscapes) as a result of a dynamic equilibrium between extinction (extirpation) and immigration, in turn dependent on island size and the distance of islands from their source pools, respectively. A novel aspect of ETIB that set it apart from previous studies of islands was the dynamic nature of turnover involving a balance between the rates at which species arrive and become extirpated ([2], but see also [3]), and turnover is the subject of this paper. Paradoxically, ETIB remains both controversial but also highly influential, with a recent review highlighting several areas of research that have developed as a direct consequence of its influence [4].
A number of tests of turnover, immigration, and extinction rates have generated similar patterns with high rates of colonisation and extinction related to small population size or with small populations repeatedly recolonising after repeated extinctions. In experimental studies in Florida, the dynamics of arboreal arthropods were monitored on small mangrove islands and turnover was observed to comprise mainly of ‘transient’ species [5]. Similarly, in experimental tests of arthropods on small Spartina islands, only ‘ephemeral’ species showed evidence of turnover [6,7,8]. A similar pattern was found for ‘fugitive’ species of island orb spiders in the Caribbean [9]. Hence, although differing in their use of terminology, these studies came to similar conclusions with regard to turnover being dominated by rare species. All these studies were over timescales of no more than four years. A test of ETIB using long-term data [10] examined both breeding land bird data for Skokholm (47 years for 1928–1979 except the war years) and a 26-year dataset of breeding birds in Eastern Wood, Bookham Common, Surrey, UK [11]. General support was found for ETIB but it was noted that much of the turnover at equilibrium comprised ‘casual’ species. Subsequently, Williamson was more damning in his critique, suggesting that, if ETIB only applies to rare or transient species, then it is ‘true but trivial’ [12,13]. He did not go on further to explain what was meant by ‘trivial’ in this context but it could refer, for example, to the reduced role of small populations in food webs, their relatively small contribution to overall biomass and energy flow, or other similar ecological relationships.
In more detailed analyses of breeding land birds on several islands around the British coast, not only were the classic ETIB immigration and extinction curves found to be good fits for most islands [14], but, in analyses of turnover using both equilibrium and non-equilibrium models, contrasting results were found for short-term (<4 years intervals) and longer-term turnover (ca. 10–60 years). Whereas the latter was characterised by both ‘intrinsic’ (i.e., turnover, as defined in ETIB, due to stochastic processes) and ‘extrinsic’ turnover (that due to, for example, successional change or changes to the mainland species pool), the former was found to be inflated by ‘floaters’ (possibly individuals moving between islands and the mainland to breed for short periods) [15].
The aim of this investigation was to use long-term data to test turnover dynamics of land birds on Skokholm over differing census intervals in order to examine the contribution of population size on turnover, specifically whether turnover is exclusive to rare (or casual, transient, or floater) species in line with ETIB being ‘true but trivial’ [13], or, if not, to assess the implications for ETIB.
2. Materials and Methods
The analysis here is based on an extension of data used by Williamson [10], recently updated to form a 69-year dataset of breeding land birds on the island of Skokholm [16,17]. The island is ca. 96 ha in size lying 3.5 km off the coast of Pembrokeshire, Southwest Wales, and is largely treeless with open communities of maritime grassland, bracken, heath, and bog. Bird records date back to 1928 with only short gaps. The efficacy of the data was tested in terms of potential observer bias (of which none could be detected [16]) and analysed in relation to successional turnover and equilibrium [18]. The time-series was found to have two distinct periods in which species richness was decreasing or increasing in 1928–1947 and 1980–2002, respectively, probably in response to habitat change. In between these two periods there were two periods of relative stability from 1948–1962 and 1963–1979, and these possibly represent equilibria as defined in ETIB [18].
A graphical approach is used here to analyse turnover using five census intervals: 1, 3, 6, 12, and 24 years. Absolute and relative numbers of immigrations and extinctions are graphically presented in relation to the population sizes of birds (1, 2, 3, 4, and ≥5 breeding pairs) over these census intervals. Absolute turnovers are the total number of immigrations and extinctions by different species over the census period, whereas relative turnovers are the absolute turnovers over n, the number of cases for each census interval. For example, for census intervals of 1, 3, 6, 12, and 24 years, respectively, the sum turnovers of single breeding pairs of bird species were 144 comprising 70 immigrations and 74 extinctions; 89 comprising 46 immigrations and 43 extinctions; 43 comprising 21 immigrations and 22 extinctions; 27 comprising 11 immigrations and 16 extinctions; and 12 comprising three immigrations and 9 extinctions (Figure 1a). The number of cases, n, for these census intervals were 64, 22, 12, 6 and 3, respectively, hence the relative turnovers are 144/64 (2.3 turnovers per census interval), 89/22 (4.0), 43/12 (3.6), 27/6 (4.5), and 12/3 (4.0), respectively (Figure 1b). As n for the longest census intervals (12 and 24 years) was just 6 and 3, respectively, confidence intervals were not calculated for any of these data.
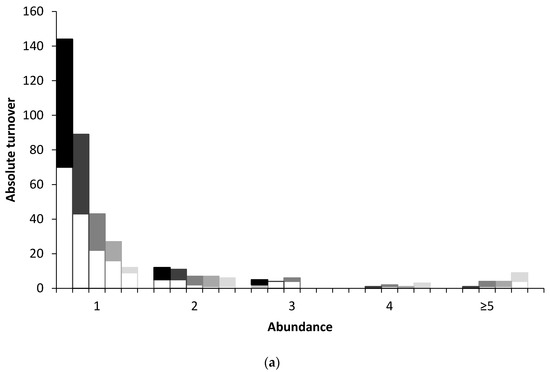
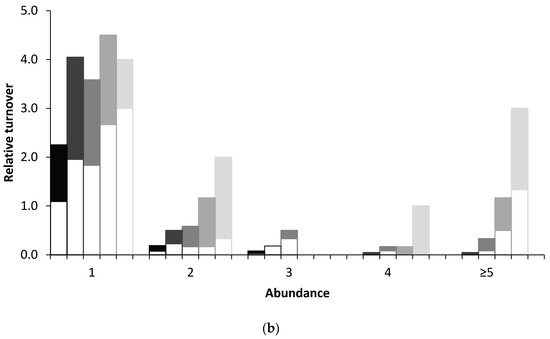
Figure 1.
Turnover showing Immigration (shaded bars) and Extinctions (open bars) in relation to abundance (numbers of breeding pairs) for (a) absolute and (b) relative data. In each abundance category, from left to right, data represent intervals of 1, 3, 6, 12, and 24 years (each having different shading), respectively. For example, in (a) for a population size of two the sum of immigrations and extinctions were 12, 11, 7, 7, and 6 for the respective census intervals. Relative turnovers are the absolute turnovers over n.
No bird species were excluded on the basis of migratory status. In total, five obligate migratory land bird species have bred on Skokholm, but three of these have not bred for several years (Corncrake Crex crex, Cuckoo Cuculus canorus, Whitethroat Sylvia communis). Just two migrant species are regular breeders: Wheatear Oenanthe oenanthe and Sedge Warbler Acrocephalus schoenobaenus [18,19]. It could be argued that, as migratory species depart from their breeding grounds, these species go extinct each year and thus have to recolonise again the following year [20]. However, some migratory species exhibit strong philopatry and return to the same localities each year after migration [20,21,22], and it would be arbitrary to exclude migrants [23], especially when we do not know whether resident species overwinter on islands or whether they undertake local migrations over to the mainland or other islands. Accordingly, I follow Lynch and Johnson (1974, p. 371) in defining the extinction as ‘the total disappearance from an island, for at least one reproductive cycle, of a species which had formerly bred there’—a definition that does not preclude recolonisation later [23]. In terms of the application of ETIB, this means that migratory species can be treated the same as resident species.
Turnover is posited to be stochastic in ETIB, but turnover may also be due to habitat change, movements between islands, pseudo-turnover (apparent extinctions due to insufficient sampling), and ‘topping-up’ from mainland populations [5,23,24]. Whilst different types of turnover are difficult to distinguish, as the central question here concerns the timescales over which turnover operates, this is not seen to be a problem for the analysis undertaken. Repeated extinctions and re-immigrations of species (or ‘in and out’ effects’ [25]) are tested for using simple correlation.
3. Results
Both absolute turnover and turnover rates were inversely related to population size at all census intervals although relative turnover was more equitable (Figure 1a,b). However, whilst the overall trend was as expected, the abundance category ≥5 masks a very long tail and includes some populations which either declined to zero after previously having had large numbers or, conversely, were not present earlier in the time series but which, after colonisation, attained large numbers (see Discussion). Further, a plot of repeated immigrations by species shows a marginal statistically significant relationship with population size (Figure 2: Spearman’s rho = −0.58, p = 0.053), i.e., much of the turnover consisted of species with low numbers repeatedly colonising and going extinct again; more than half of all immigrations were of species that immigrated three of more times.
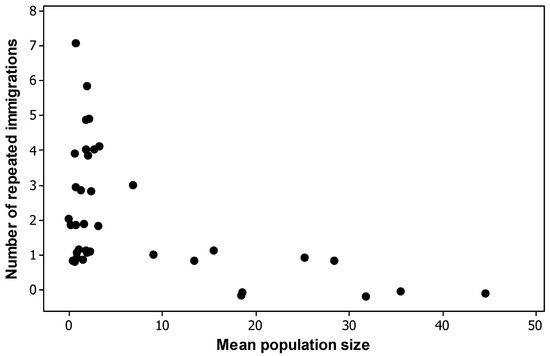
Figure 2.
Number of repeated immigrations per species versus mean population size. Most species involved in turnovers have small population sizes. Data have been jittered to display multiple species at the same points.
4. Discussion
This investigation supports the idea that most turnover consists of repeated immigrations and extinctions of rare, casual, or transient species when measured over short census intervals. However, over longer census intervals, measured up to 24 years in this study, a sizeable minority of species either colonise afresh and increase to large population sizes and/or decline from high numbers to extinction. This not only demonstrates that long time series are required in order to identify such long-term fluctuations in populations and how these might contribute to turnover, but that turnover is not exclusive to rare species. For example, Stock Dove Columba oenas first colonised Skokholm in 1967, establishing a single breeding pair. Within eight years, its population had rapidly increased to reach a peak of 62 pairs; however, after six years of relative stability at this level, it then proceeded to decline at a rate equivalent to its previous growth, and by 1984 it had gone extinct. Similarly, Jackdaw Corvus monedula colonised in 1965 establishing three pairs and had 60 breeding pairs within 10 years. It then underwent a period of decline, and, for the last 10 years of the available data, its abundance fluctuated at around 13 pairs. Further, Starling Sturnus vulgaris colonised in 1940 and its numbers remained below 11 pairs until 1963 when it began to increase. Between 1965 and 1982, its numbers increased to a peak of 50 but subsequently declined to about six pairs [17,19]. In summary, these three species comprise those in the ≥5 category for a census interval of 24 years. In addition, for a census interval of 12 years, Wren Troglodytes troglodytes colonised in 1988 and rapidly increased to reach levels of up to 21 pairs. In total, these four species built up relatively large numbers after colonisation and represent 11% of a total of 37 species of land birds that bred on the island between 1928 and 2002. Whilst these species were discussed previously [12], the analysis here demonstrates not just that such turnovers are only likely to be detected in datasets covering several decades, but that turnover is not exclusive to small populations.
Skokholm is a bird observatory, and considerable numbers of birds are recorded each year on passage [19]. Being only 3.5 km from the mainland, populations may be subject to regular ‘topping up’ (sensu [24]), which may not only affect extinction rates but also contribute to repeat immigrations and extinctions. For example, Wren Troglodytes troglodytes was a winter visitor both before and after they colonised Skokholm and were also regularly found on passage in October including a peak of 200 in 1974. Jackdaw Corvus monedula are regular spring visitors and Starling Sturnus vulgaris were regularly visitors on passage in autumn before and after they established breeding populations; in addition, large numbers of juveniles and adults arrive each year in July. Further, the decline of both Starling and Stock Dove Columba oenas coincided with national trends [19]. These repeated observations of individuals on passage, visiting, or over-wintering suggest possible linkages between mainland and island populations. An analysis of trends of birds on Ouessant, a 15 km2 island ca. 20 km off the coast of France in the English Channel, has shown that recent changes in species richness and turnover of birds on the island are significantly statistically correlated with mainland trends [26]. Accordingly, further analysis is recommended to consider trends in colonisation and extinction relative to population trends in Great Britain, not just for Skokholm but for other offshore islands as well.
A further explanation for turnover is the change in populations in direct response to habitat availability. Studying land bird populations on Skye, Scotland, turnover was ascribed to small populations often restricted by habitat availability [27]. It is, therefore, important to distinguish between successional and stochastic turnover and here none of the turnovers over longer census intervals directly corresponded to periods of successional change (1928–1947 and 1980–2002 [18], see Materials and Methods) and hence may be stochastic. In the analysis here, none of colonisations of the four species considered in detail corresponded with periods identified as those previously characterised by habitat change [18]; except for Wren, the remaining three species primarily nest on rocky outcrops, and there was no net change of this habitat.
There are several reasons to discount the ‘true but trivial’ criticism, including the occurrence of equilibria, despite high turnovers, and the possibility, outlined above, of relationships to mainland populations. To conclude, turnover may appear to be ‘trivial’ when considered over short timescales, but it is demonstrated here that, over long timescales, it is possible to detect colonisation events and/or extinction events that are in all likelihood stochastic in nature and that ultimately lead to the establishment of notable populations or result from the decline of previously ‘non-trivial’ populations, respectively.
Acknowledgments
Funds for covering the costs to publish in open access were received from the University of Northampton’s Open Access Fund. I would like to thank Jeff Ollerton and Robin Crockett for valuable discussions and three anonymous referees for their valuable input.
Conflicts of Interest
The author declares no conflict of interest.
References
- MacArthur, R.H.; Wilson, E.O. The Theory of Island Biogeography; Princeton University Press: Princeton, NJ, USA, 1967. [Google Scholar]
- Simberloff, D. The dynamic equilibrium theory of island biogeography: The origin, the legacy and the future. In Island Biology 2016 International Conference on Island Evolution, Ecology, and Conservation, University of the Azores, Azores, Portugal, 18–22 July 2016; Gabriel, R., Elias, R.B., Amorin, I.R., Borges, P.A.V., Eds.; University of the Azores: Azores, Portugal, Arquipelago: Life and Marine Sciences; Suppl. 9: 565, p. 2.
- Brown, J.H.; Lomolino, M.V. On the nature of scientific revolutions: Independent discovery of the equilibrium theory of island biogeography. Ecology 1989, 70, 1954–1957. [Google Scholar] [CrossRef]
- Warren, B.H.; Simberloff, D.; Ricklefs, R.E.; Aguilee, R.; Condamine, F.L.; Gravel, D.; Morlon, H.; Mouquet, N.; Rosindell, J.; Casquet, J.; et al. Islands as model systems in ecology and evolution: Prospects fifty years after MacArthur-Wilson. Ecol. Lett. 2015, 18, 200–217. [Google Scholar] [CrossRef] [PubMed]
- Simberloff, D. Turnover and equilibrium island biogeography. Science 1976, 194, 572–578. [Google Scholar] [CrossRef] [PubMed]
- Rey, J.R. Ecological biogeography of arthropods on Spartina islands in northwest Florida. Ecol. Monogr. 1981, 51, 237–265. [Google Scholar] [CrossRef]
- Strong, D.R.; Rey, J.R. Testing for MacArthur-Wilson equilibrium with the arthropods of the miniature Spartina archipelago at Oyster Bay, Florida. Am. Zool. 1982, 22, 355–360. [Google Scholar] [CrossRef]
- Rey, J.R.; Strong, D.R. Immigration and extinction of salt-marsh arthropods on islands—An experimental-study. Oikos 1983, 41, 396–401. [Google Scholar] [CrossRef]
- Schoener, T.W.; Spiller, D.A. High population persistence m a system with high turnover. Nature 1987, 330, 474–477. [Google Scholar] [CrossRef]
- Williamson, M. Island Populations; Oxford University Press: Oxford, UK, 1981. [Google Scholar]
- Beven, G. Changes in breeding bird populations of an oak-wood on Bookham Common, Surrey, over twenty-seven years. Lond. Nat. 1976, 55, 23–42. [Google Scholar]
- Williamson, M. The land-bird community of Skokholm: Ordination and turnover. Oikos 1983, 41, 378–384. [Google Scholar] [CrossRef]
- Williamson, M. The MacArthur-Wilson theory today: True but trivial. J. Biogeogr. 1989, 16, 3–4. [Google Scholar]
- Manne, L.L.; Pimm, S.L.; Diamond, J.M.; Reed, T.M. The form of the curves: A direct evaluation of MacArthur & Wilson’s classic theory. J. Anim. Ecol. 1998, 67, 784–794. [Google Scholar]
- Russell, G.J.; Diamond, J.M.; Pimm, S.L.; Reed, T.M. A century of turnover: Community dynamics at three timescales. J. Anim. Ecol. 1995, 64, 628–641. [Google Scholar] [CrossRef]
- McCollin, D. Reconstructing long-term ecological data from annual census returns: A test for observer bias in counts of bird populations on Skokholm 1928–2002. Ecol. Indic. 2014, 46, 336–339. [Google Scholar] [CrossRef]
- McCollin, D. Abundance estimates for landbirds and seabirds extracted and compiled from annual reports of the Skokholm bird observatory. Available online: https://doi.pangaea.de/10.1594/PANGAEA.833759 (accessed on 1 December 2016).
- McCollin, D. The curious case of Skokholm: Equilibrium, non-equilibrium and a phase shift in an island landbird assemblage. Ecography 2015, 38, 986–991. [Google Scholar] [CrossRef]
- Thompson, G.V.F. The Natural History of Skokholm; Trafford: Bloomington, IN, USA, 2007. [Google Scholar]
- Pimm, S.L.; Jones, H.L.; Diamond, J. On the Risk of Extinction. Amer. Natur. 1988, 132, 757–785. [Google Scholar] [CrossRef]
- Greenwood, P.J. Mating systems, philopatry and dispersal in birds and mammals. Anim. Behav. 1980, 28, 1140–1162. [Google Scholar] [CrossRef]
- Förschler, M.I.; del Val, E.; Bairlein, F. Extraordinary high natal philopatry in a migratory passerine. J. Ornith. 2010, 151, 745–748. [Google Scholar] [CrossRef]
- Lynch, J.F.; Johnson, N.K. Turnover and equilibria in insular avifaunas, with special reference to the California Channel Islands. Condor 1974, 76, 370–384. [Google Scholar] [CrossRef]
- Brown, J.H.; Kodric-Brown, A. Turnover rates in insular biogeography: Effect of immigration on extinction. Ecology 1977, 58, 445–449. [Google Scholar] [CrossRef]
- Diamond, J.M.; May, R.M. Species turnover rates on islands: Dependence on census interval. Science 1977, 197, 266–270. [Google Scholar] [CrossRef] [PubMed]
- Christian, K.; Isabelle, L.V.; Frédéric, J.; Vincent, D. More species, fewer specialists: 100 Years of changes in community composition in an island-biogeographical study. Divers. Distrib. 2009, 15, 641–648. [Google Scholar] [CrossRef]
- Reed, T.M. Turnover frequency in island birds. J. Biogeogr. 1980, 7, 329–335. [Google Scholar] [CrossRef]
© 2017 by the author. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license ( http://creativecommons.org/licenses/by/4.0/).