Diversity and Spatial Distribution of Extant Freshwater Ostracodes (Crustacea) in Ancient Lake Ohrid (Macedonia/Albania)

Abstract
:1. Introduction
Study Region
2. Sampling Survey
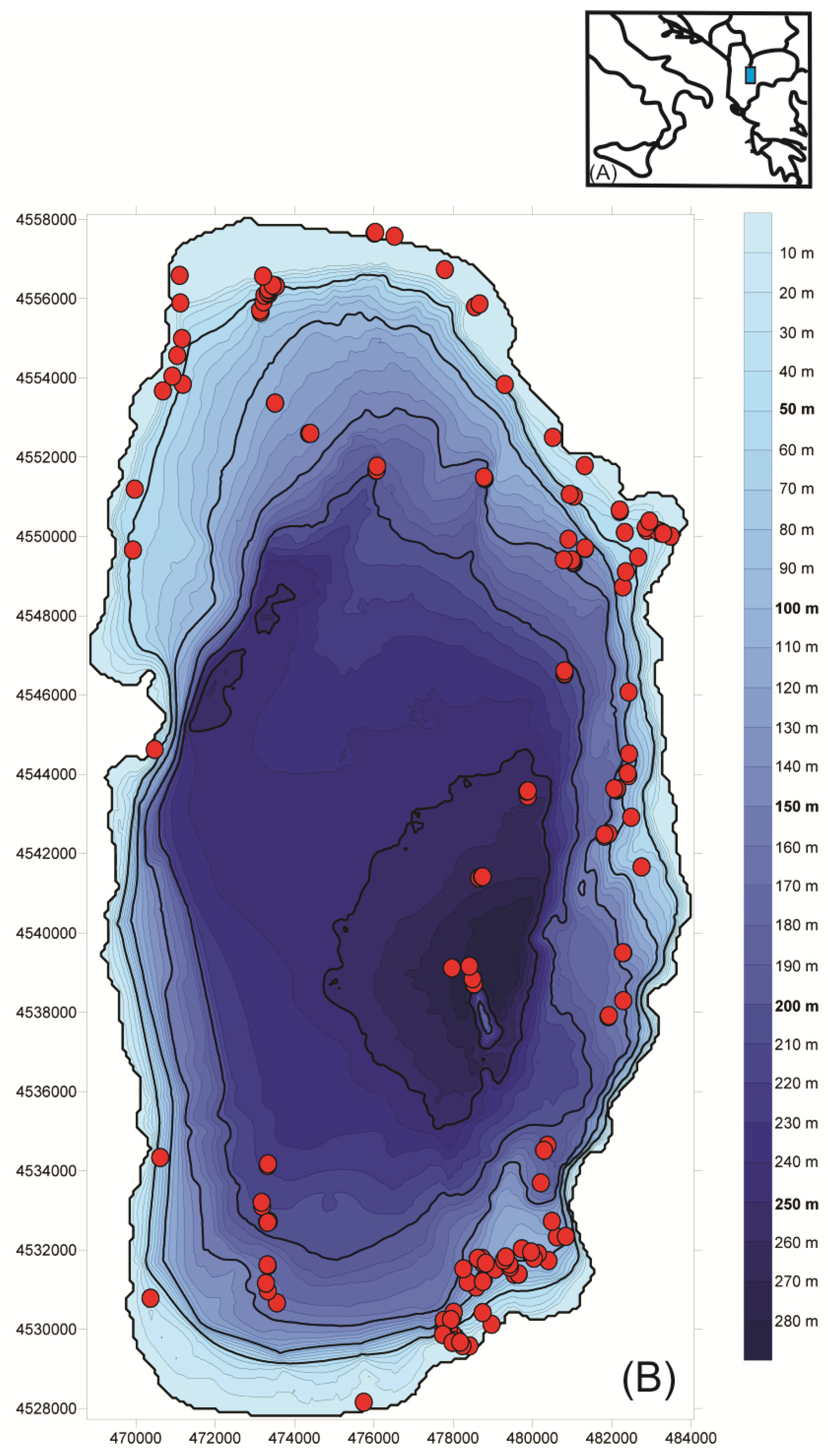
3. Results and Discussion
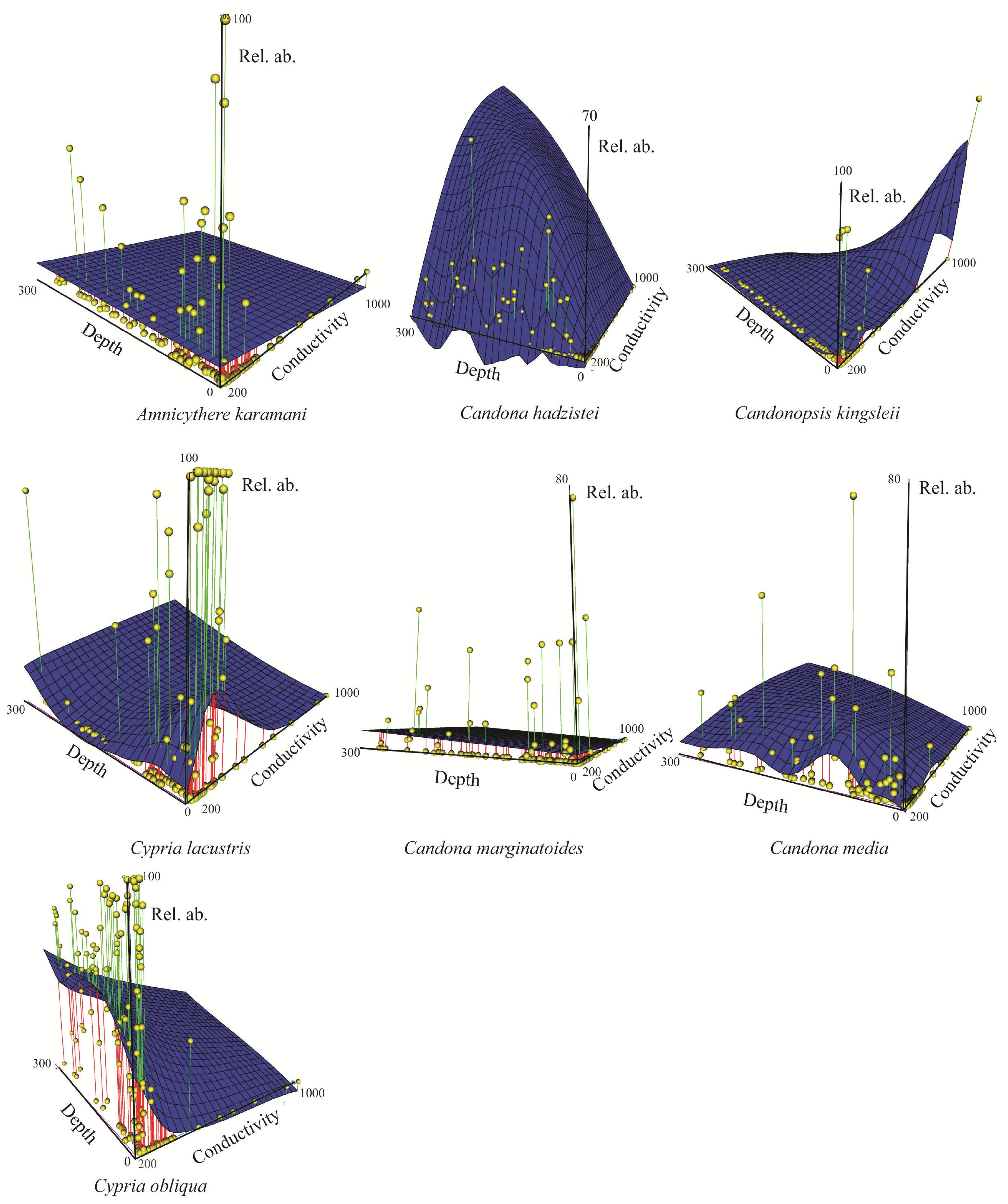
3.1. Ostracode Diversity
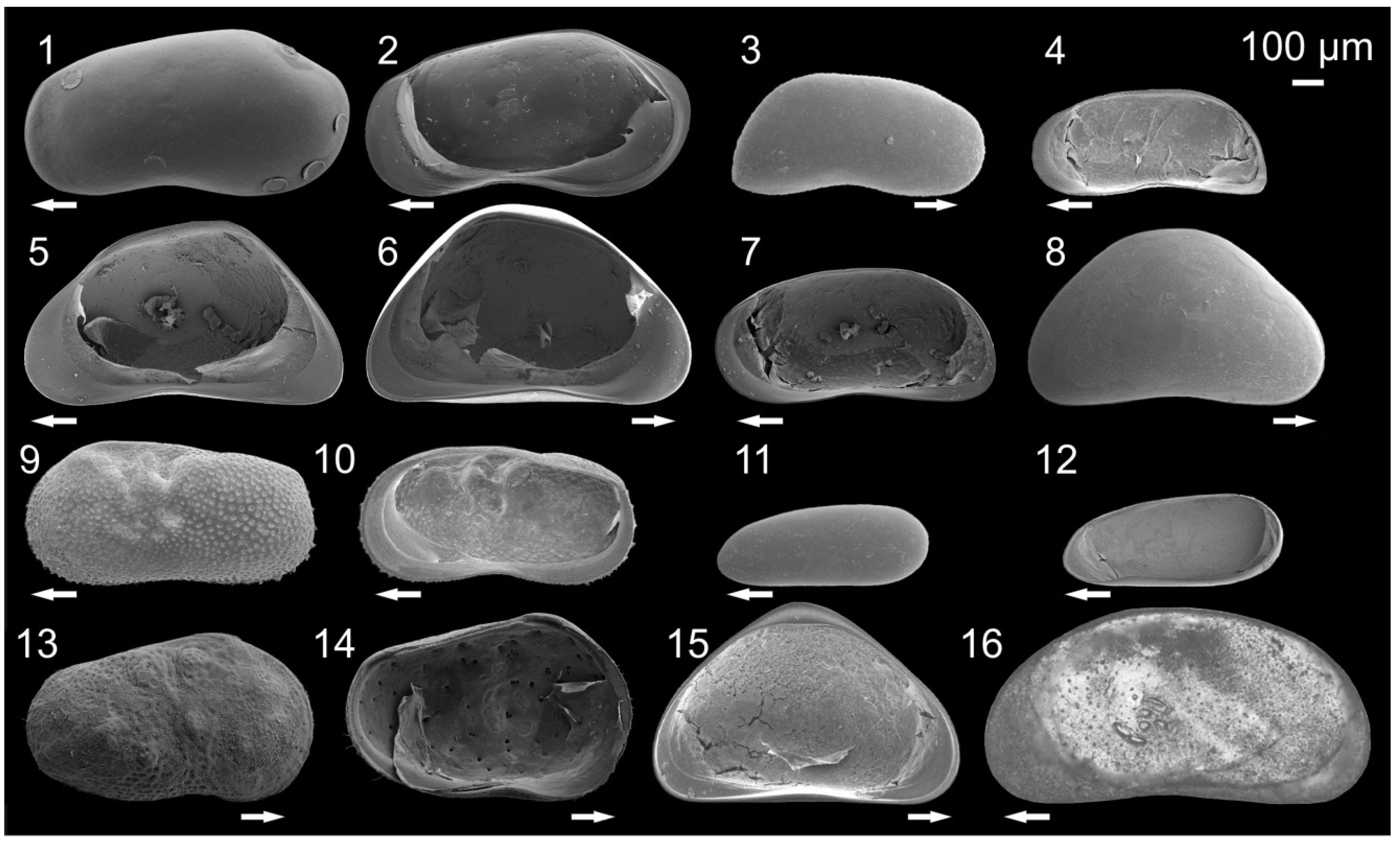
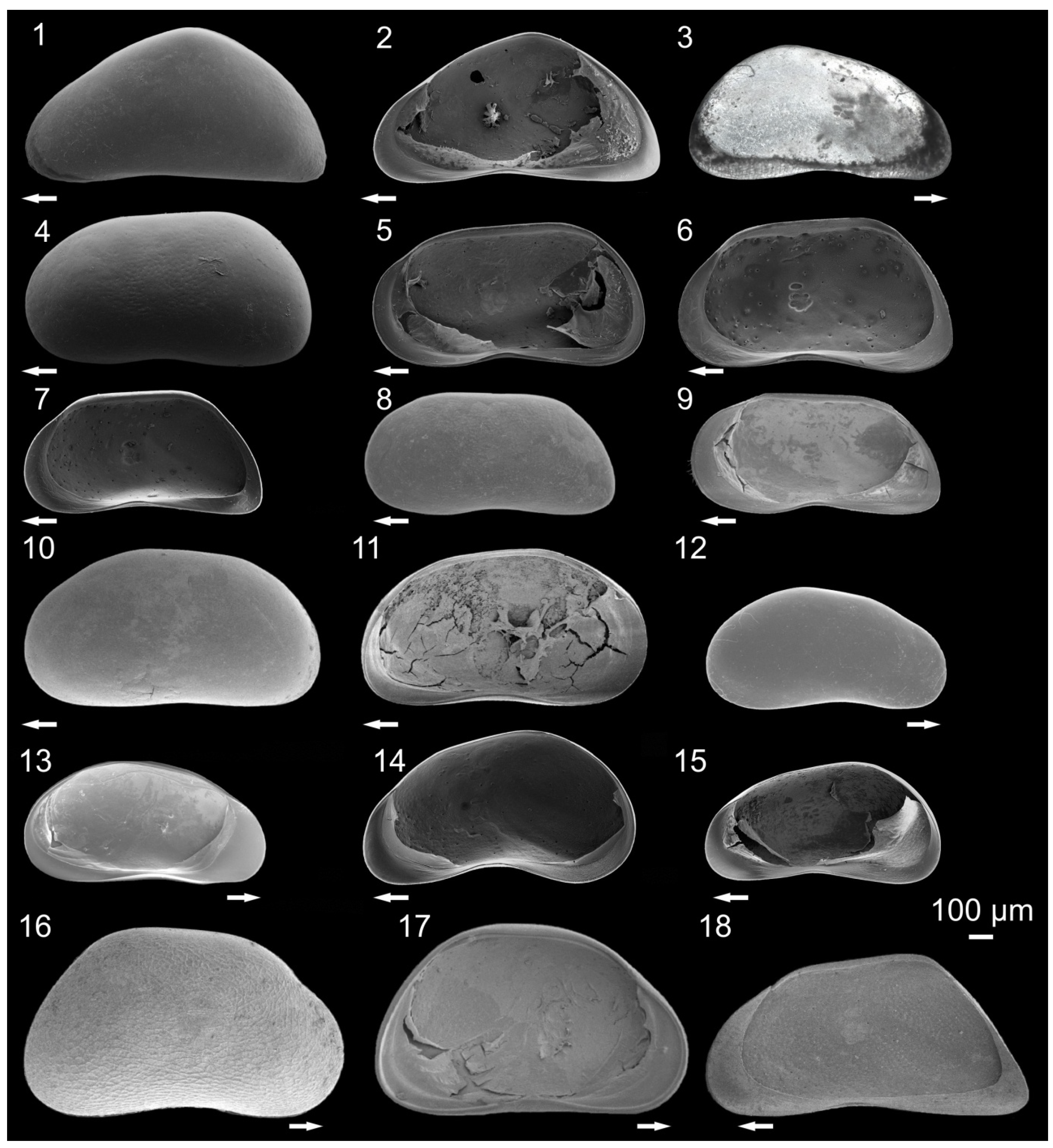
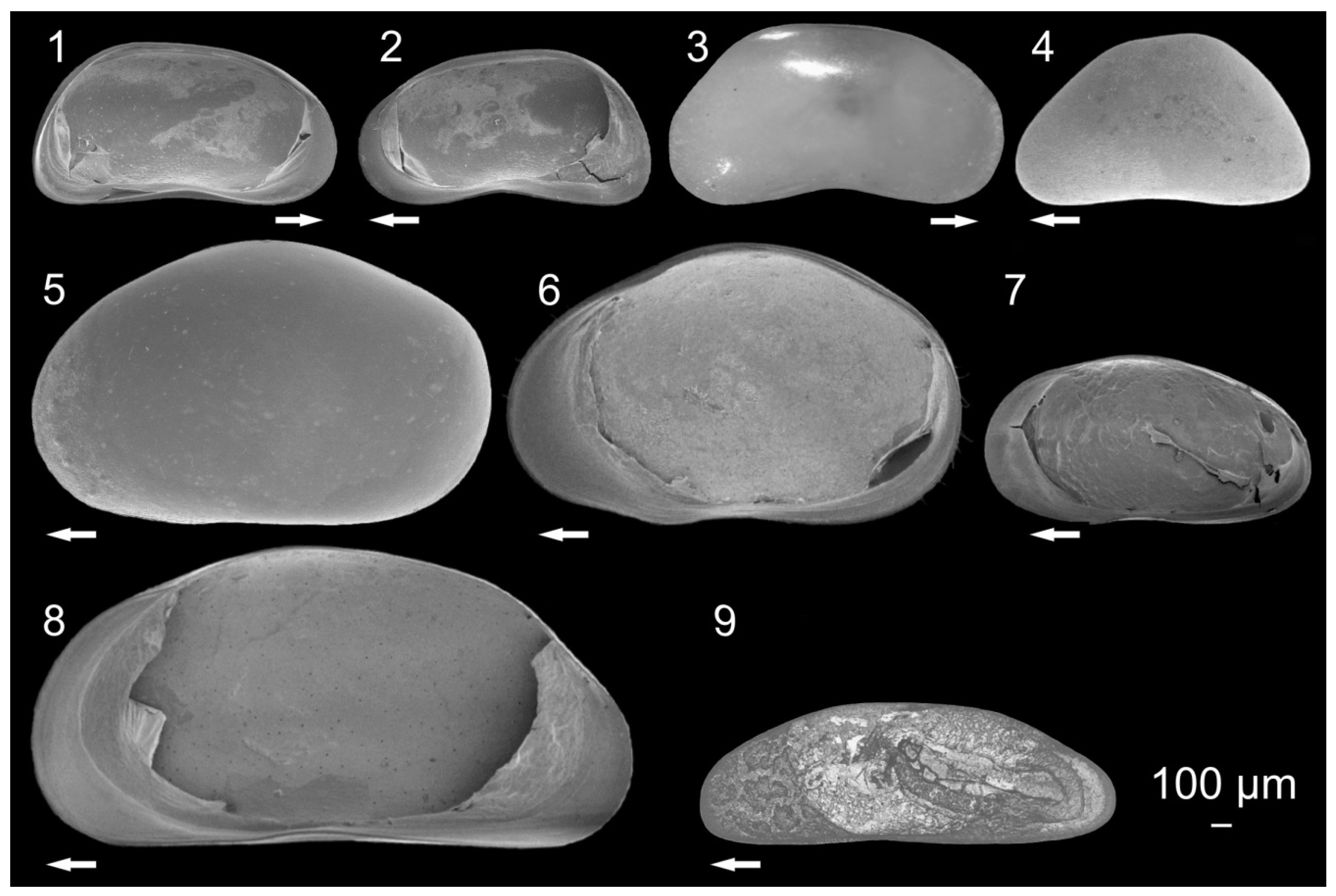
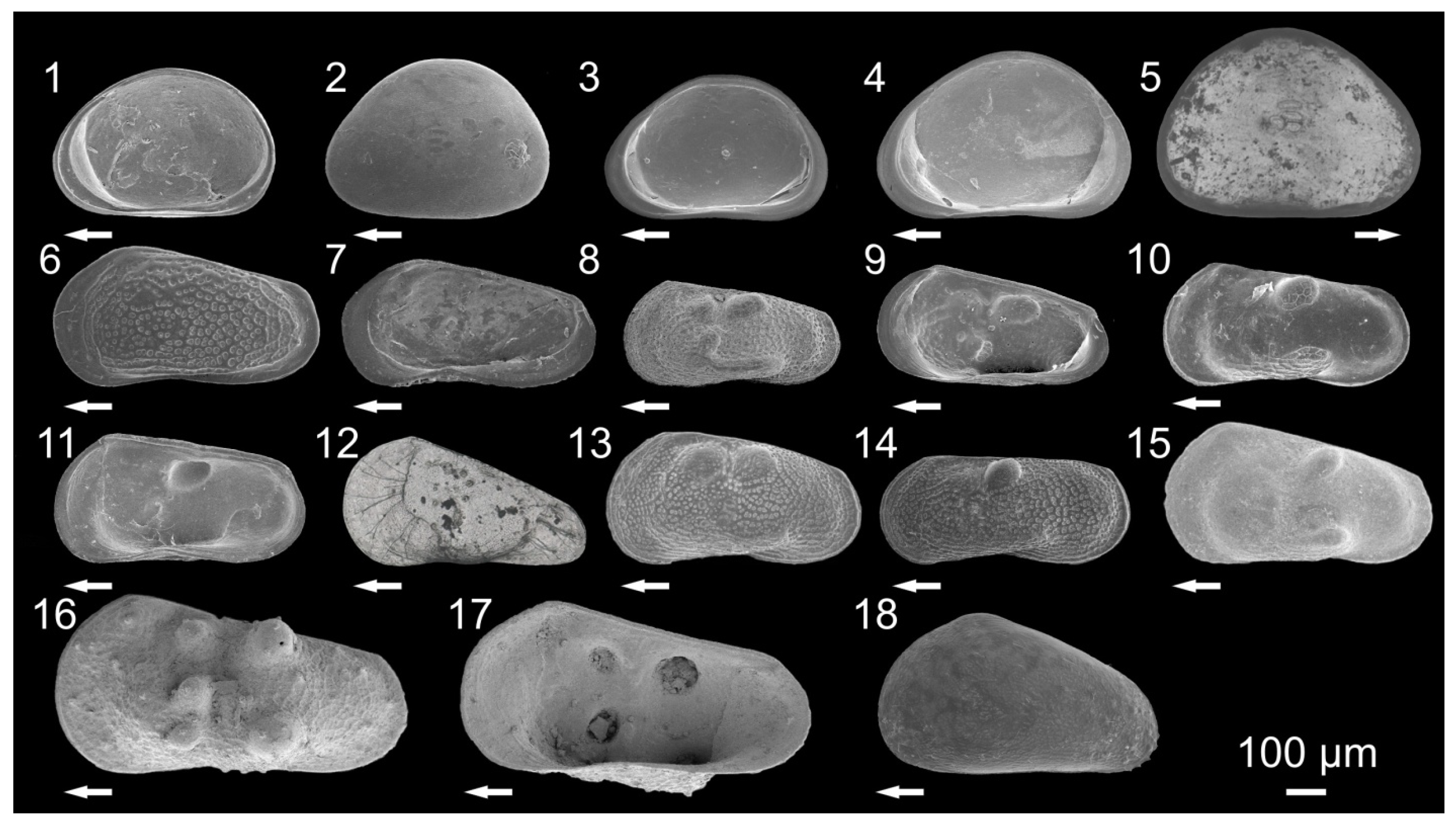

{kind=link}
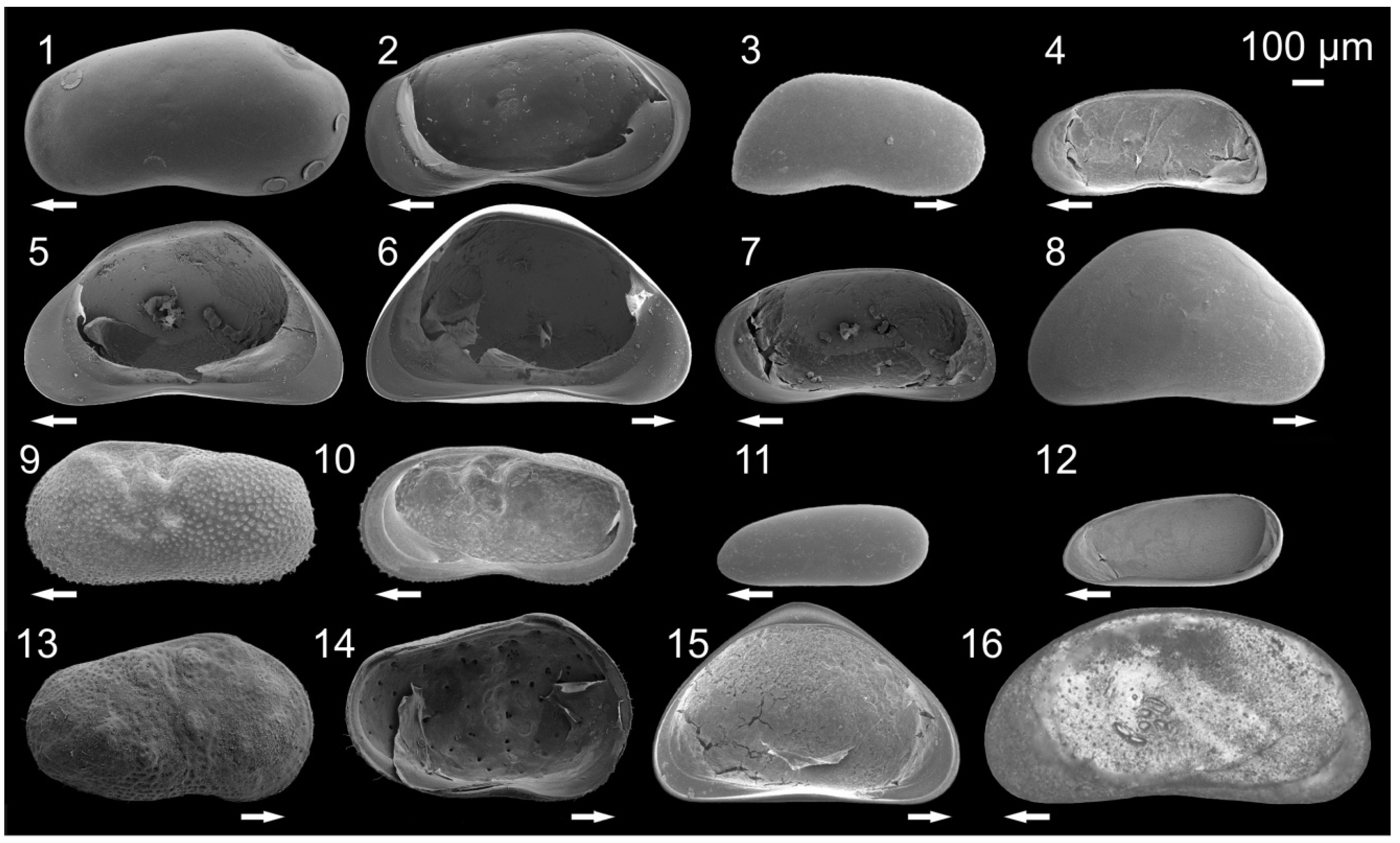{kind=link}
{kind=link}
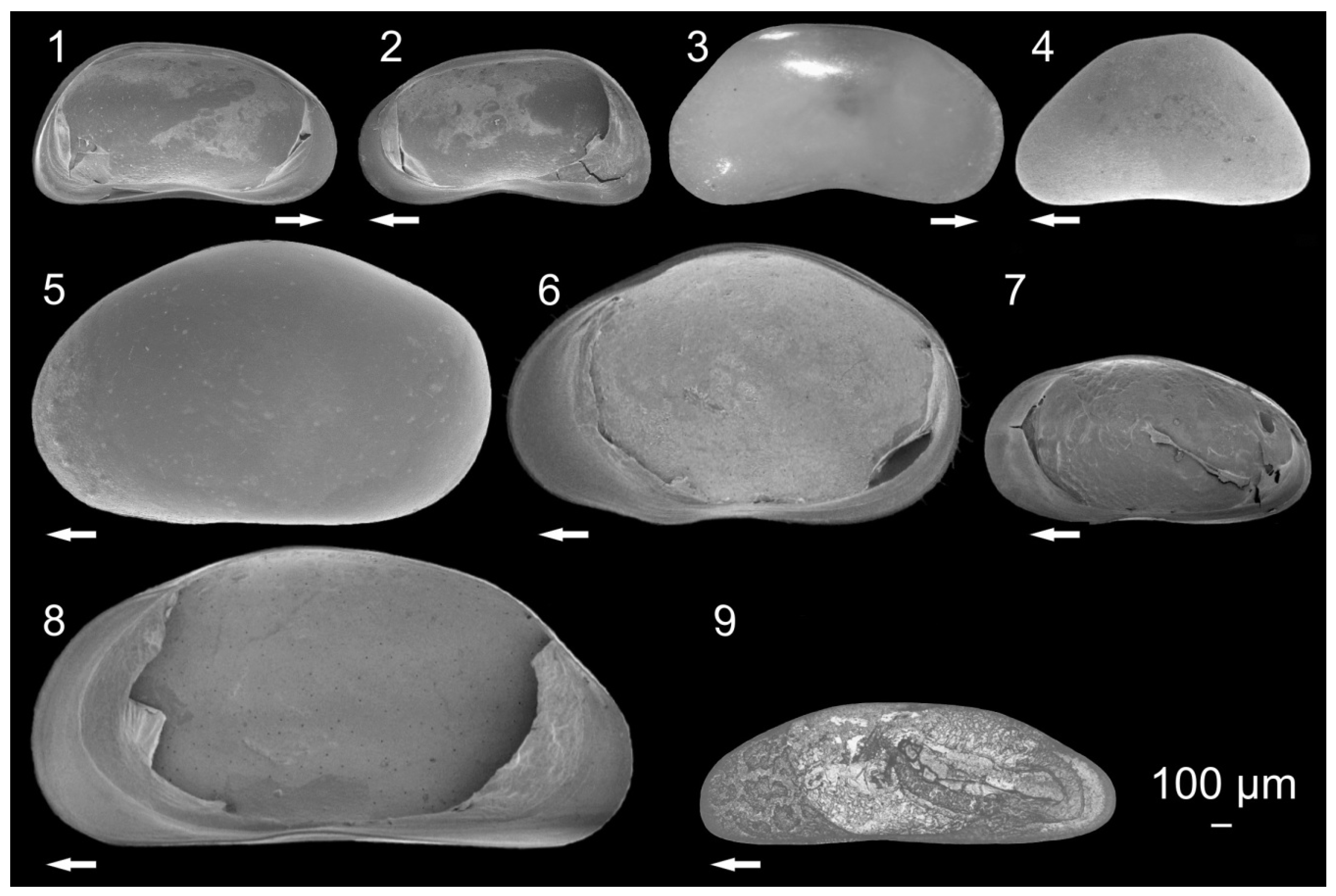{kind=link}
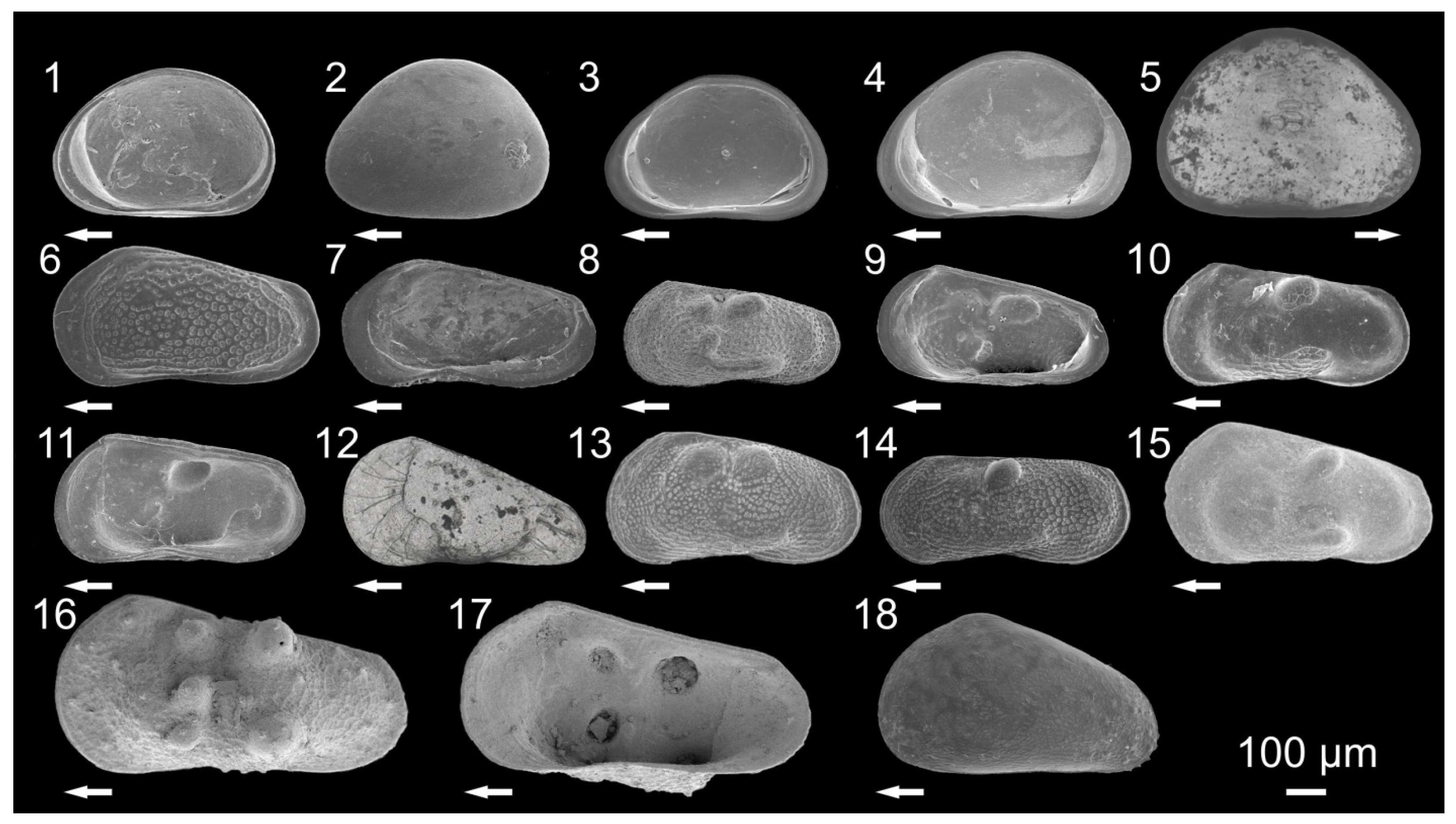{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Ostracode Species in Lake Ohrid | This Paper | Other Authors | Petkovski (2005, Unpubl. Data) |
|---|---|---|---|
| Amnicythere angulata | (Klie, 1939b) | + | |
| Amnicythere karamani | + | (Klie, 1939b) | + |
| Amnicythere prespensis | (Petkovski & Keyser, 1992) | + | |
| Amnicythere proboscidea | (Klie, 1939b) | + | |
| Bradleystrandesia reticulata | + | ||
| Candona acricauda | Mikulić, 1961 | synonym of Candona holmesi | |
| Candona alta | Klie, 1939a | Candona neglecta (review needed) | |
| Candona bimucronata | + | ||
| Candona candida | Holmes, 1937 | ||
| Candona compressa | Klie, 1942 | Pseudocandona compressa | |
| Candona cristatella | Klie, 1939a | synonym of Candona ohrida | |
| Candona depressa | Klie, 1939a | + | |
| Candona expansa | * | Mikulić,1961 | + |
| Candona fabaeformis | Klie, 1939a | Eucandona (Fabaeformiscandona) krstici | |
| Candona formosa | Mikulić,1961 | + | |
| Candona goricensis | + | Mikulić,1961 | + |
| Candona hadzistei | + | Petkovski et al. 2002 | + |
| Candona hartmanni | + | Petkovski, 1969a | + |
| Candona holmesi | * | Petkovski, 1960a | + |
| Candona jordeae | Petkovski et al. 2002 | + | |
| Candona litoralis | + | Mikulić,1961 | + |
| Candona lucida | Petkovski, 1969a | renamed in Candona jordeae | |
| Candona lychnitis | Petkovski, 1969a | + | |
| Candona macedonica | Mikulić,1961 | + | |
| Candona margaritana | + | Mikulić,1961 | |
| Candona marginata | * | Klie, 1942 | + |
| Candona marginatoides | + | Petkovski, 1960b | + |
| Candona media | + | Klie, 1939a | Candona neglecta (review needed) |
| Candona neglecta | Holmes, 1937 | + | |
| Candona ohrida | + | Holmes, 1937 | + |
| Candona ovalis | + | Mikulić,1961 | + |
| Candona parvula | Mikulić,1961 | renamed in Candona hadzistei | |
| Candona sp. | Petkovski, 2005 (unpub. data) | + | |
| Candona sublitoralis | Mikulić,1961 | synonym of Candona ohrida | |
| Candona trapeziformis | + | Klie, 1939a | + |
| Candona triangulata | * | Klie, 1939a | + |
| Candona vidua | + | Klie, 1942 | + |
| Candonopsis kingsleii | + | Klie, 1942 | + |
| Cyclocypris ovum | + | Klie, 1939a | + |
| Cypria lacustris | + | Petkovski, 1960b | + |
| Cypria obliqua | + | Klie, 1939a | + |
| Cypria ophtalmica | * | Holmes, 1937 | |
| Cytherissa lacustris | + | Klie, 1942 | + |
| Darwinula stevensoni | + | Holmes, 1937 | + |
| Dolerocypris sinensis | + | ||
| Eucypris virens | + | ||
| Eucypris sp. | + | ||
| Fabaeformiscandona krstici | + | Petkovski, 1969a | + as Eucandona (Fabaeformiscandona) krstici |
| Herpetocypris sp. 2 | + | ||
| Ilyocypris bradyi | + | ||
| Paralimnocythere alta | + | Klie, 1939b | + |
| Paralimnocythere georgevitschi | + | (Petkovski, 1960c) | + |
| Paralimnocythere karamani | + | (Petkovski, 1960c) | + |
| Paralimnocythere ochridense | + | (Klie, 1934) | + |
| Paralimnocythere slavei | + | (Petkovski, 1969b) | + |
| Paralimnocythere umbonata | * | (Klie, 1939b) | + |
| Physocypria sp. (Physocypria kraepelini?) | Holmes, 1937 | Physocypria kliei? (review needed) | |
| Prionocypris zenkeri | + | ||
| Pseudocandona compressa | + | ||
| Pseudocandona elongata | Holmes, 1937 | + | |
| Pseudocandona slavei | Petkovski, 1969a | + |
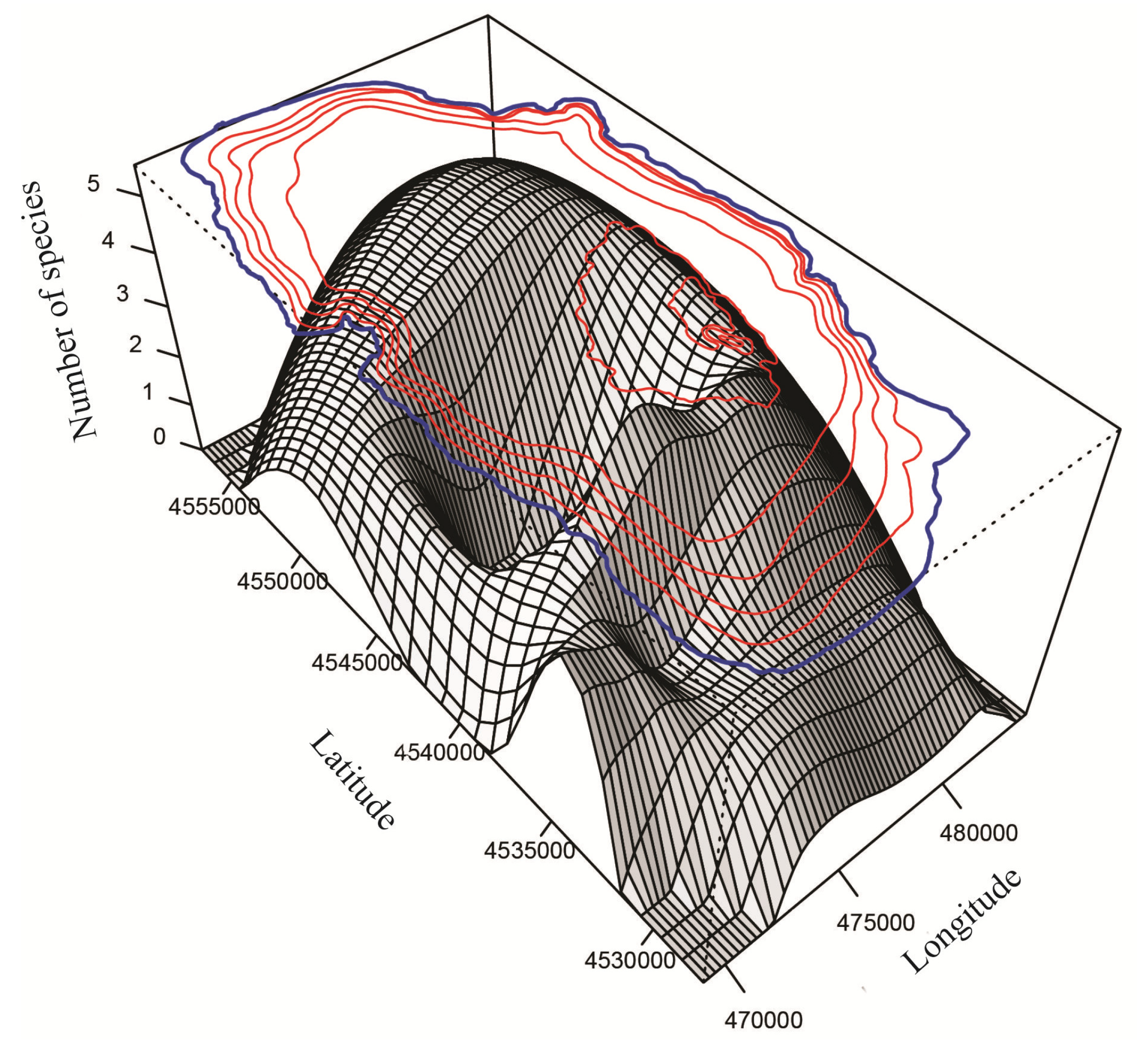
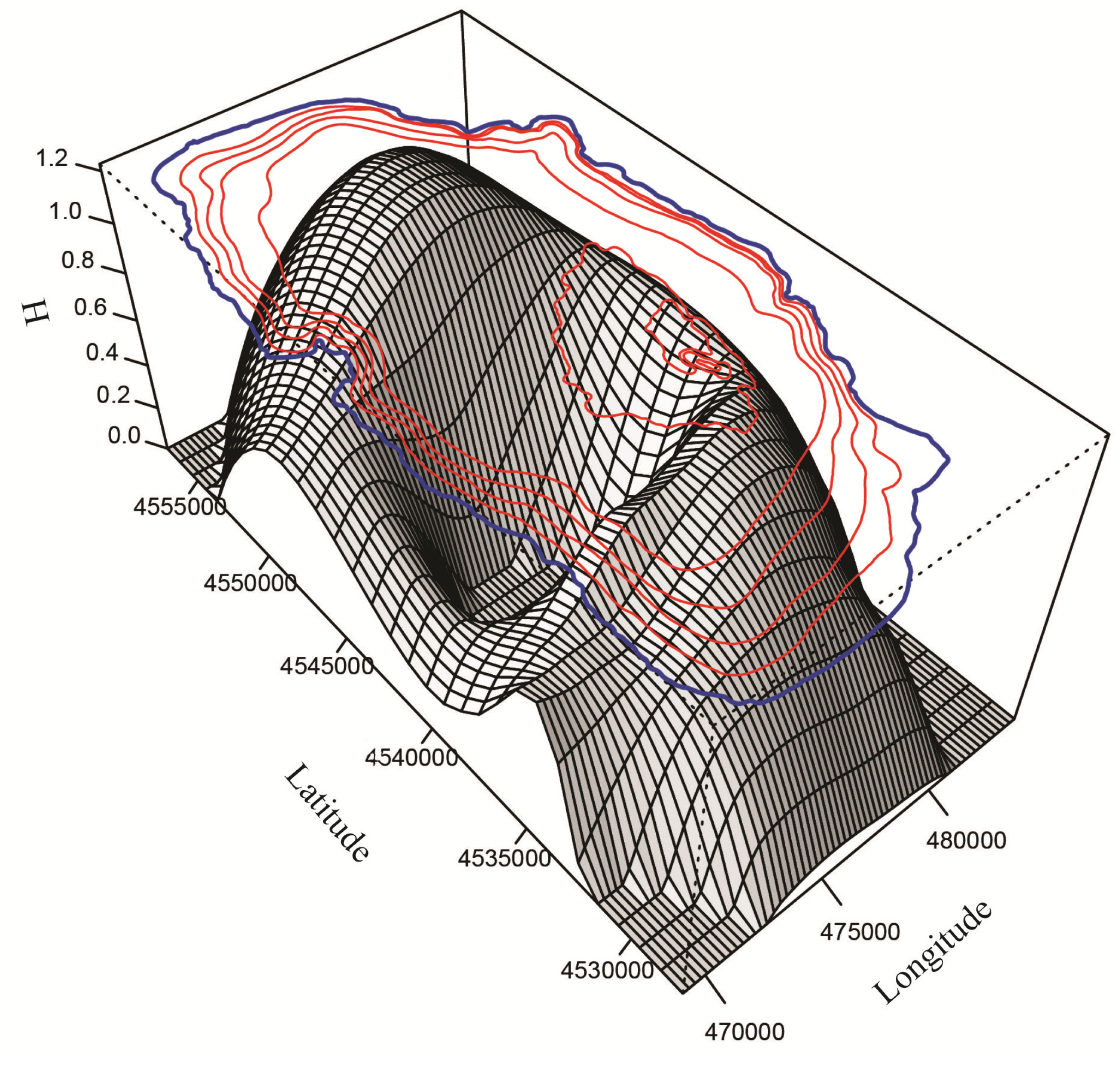
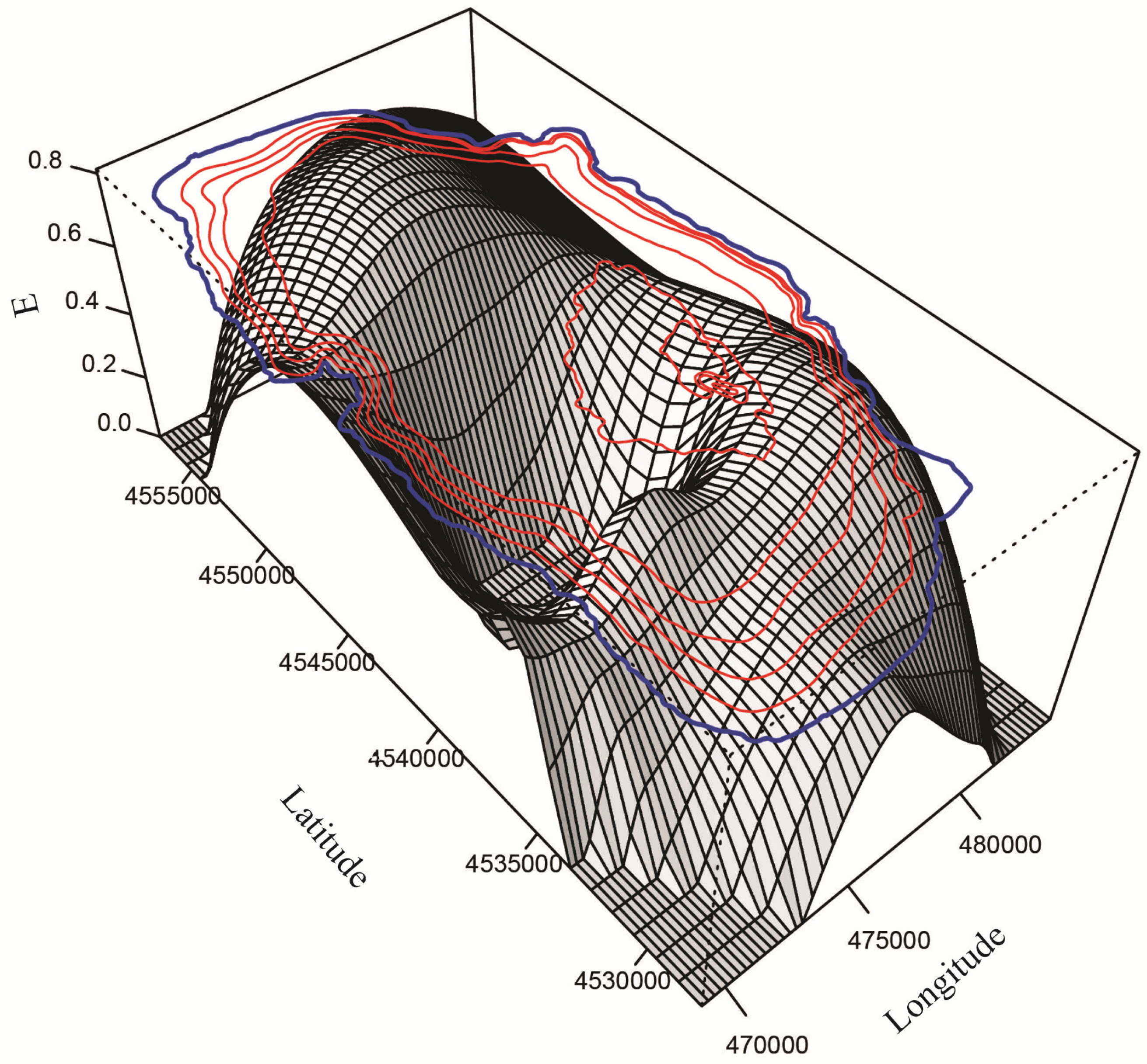
3.2. Spatial Distribution, Species Richness and Diversity of Ostracodes
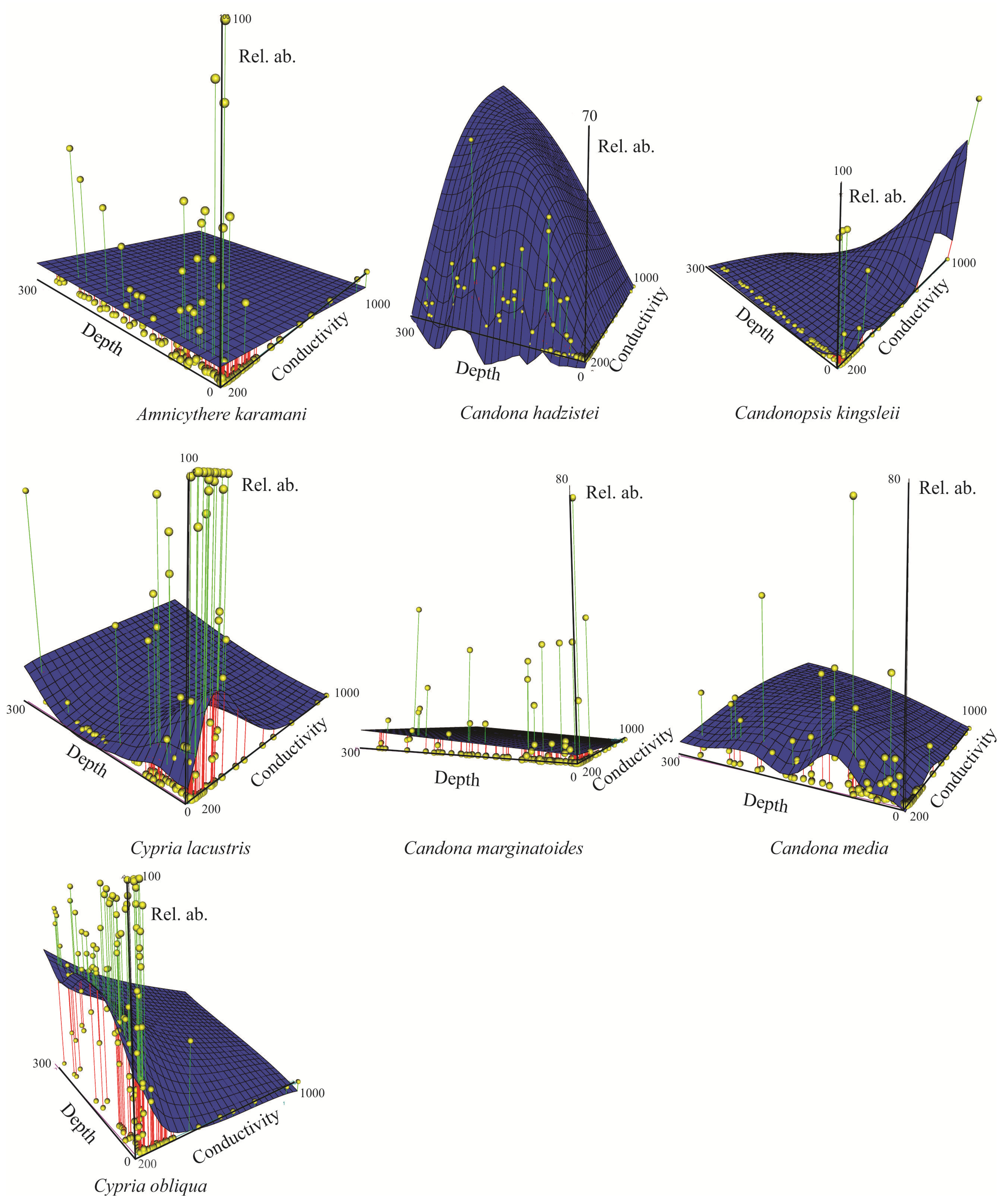
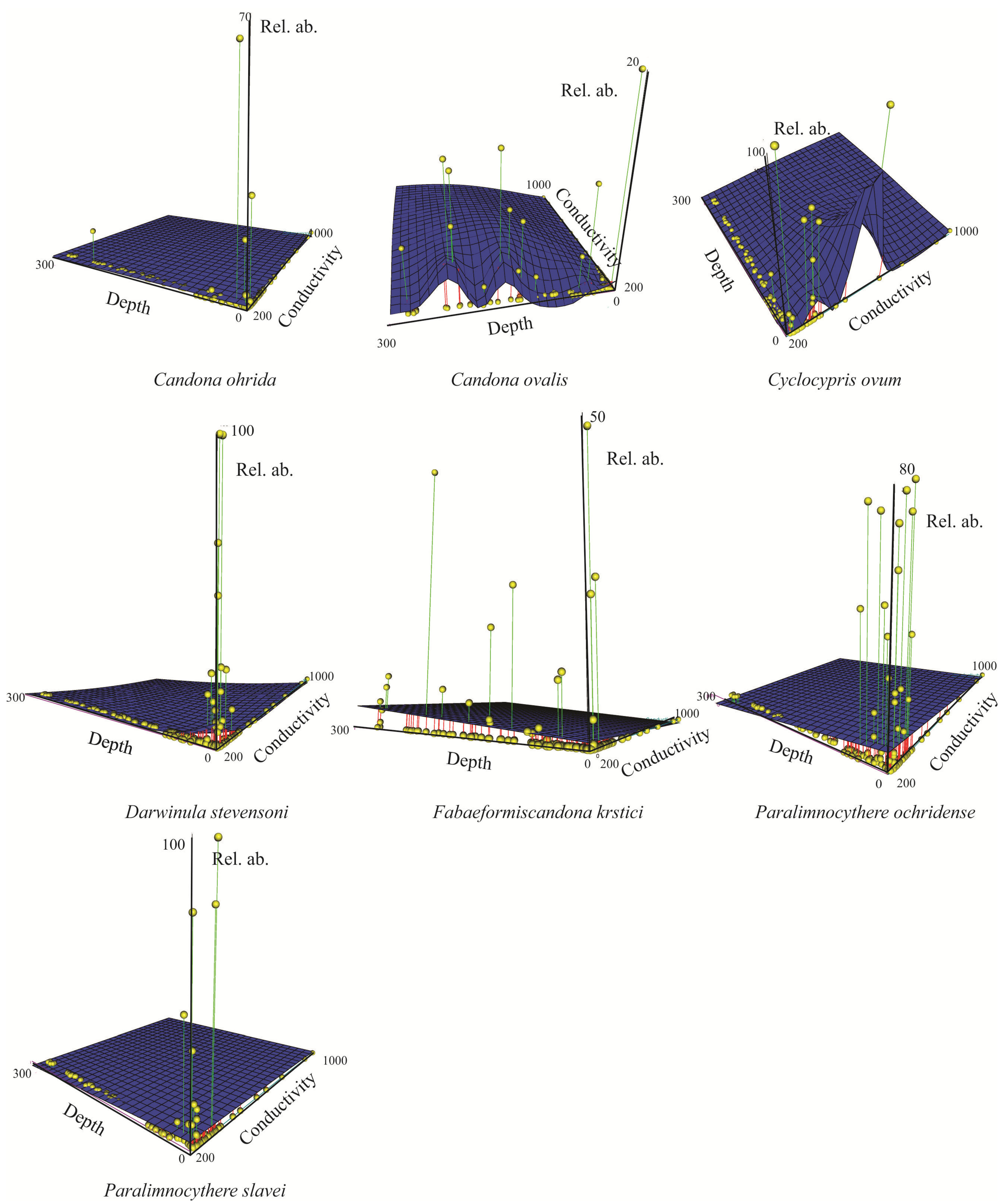
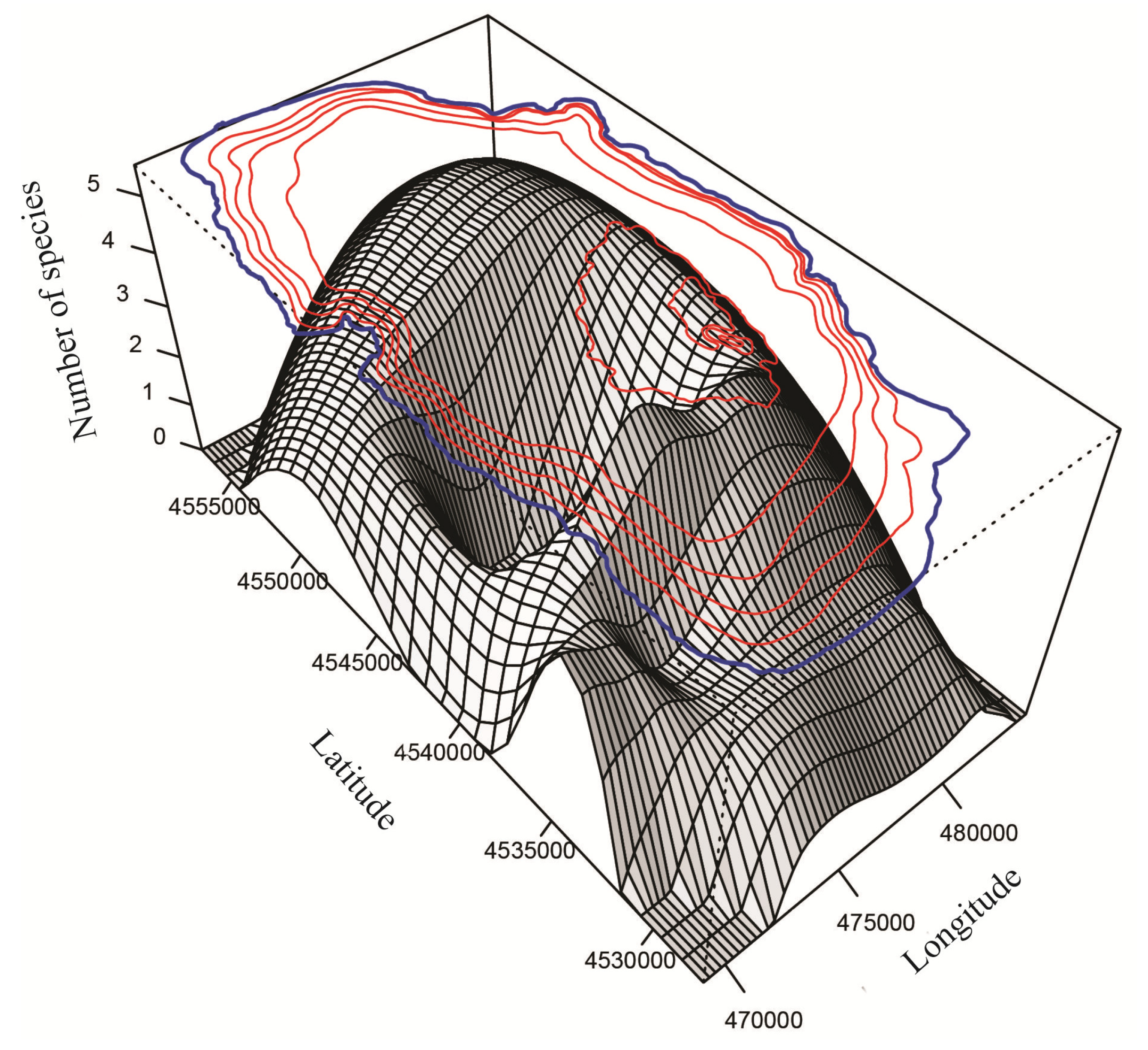
3.2.1. Spatial Distribution
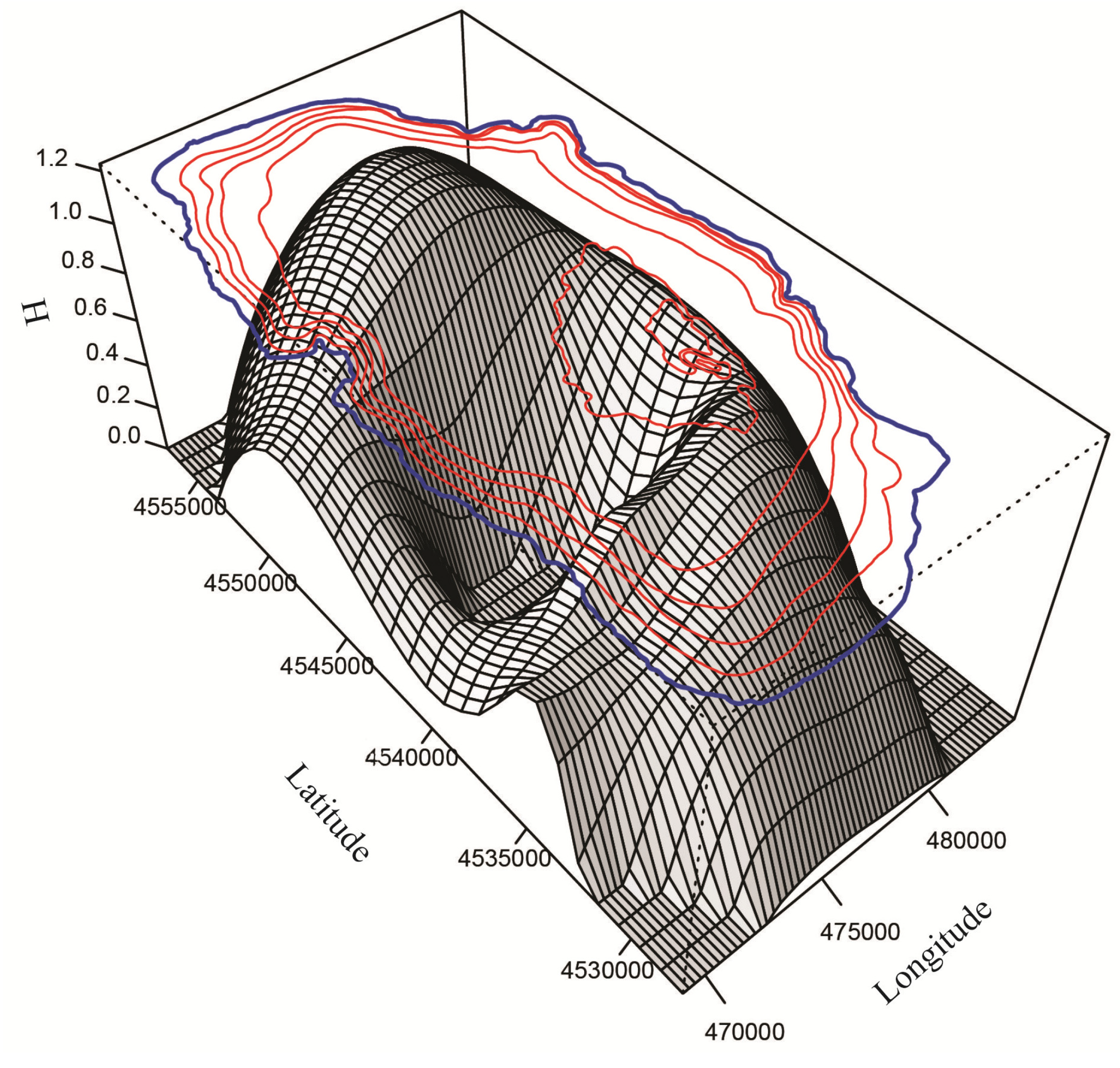
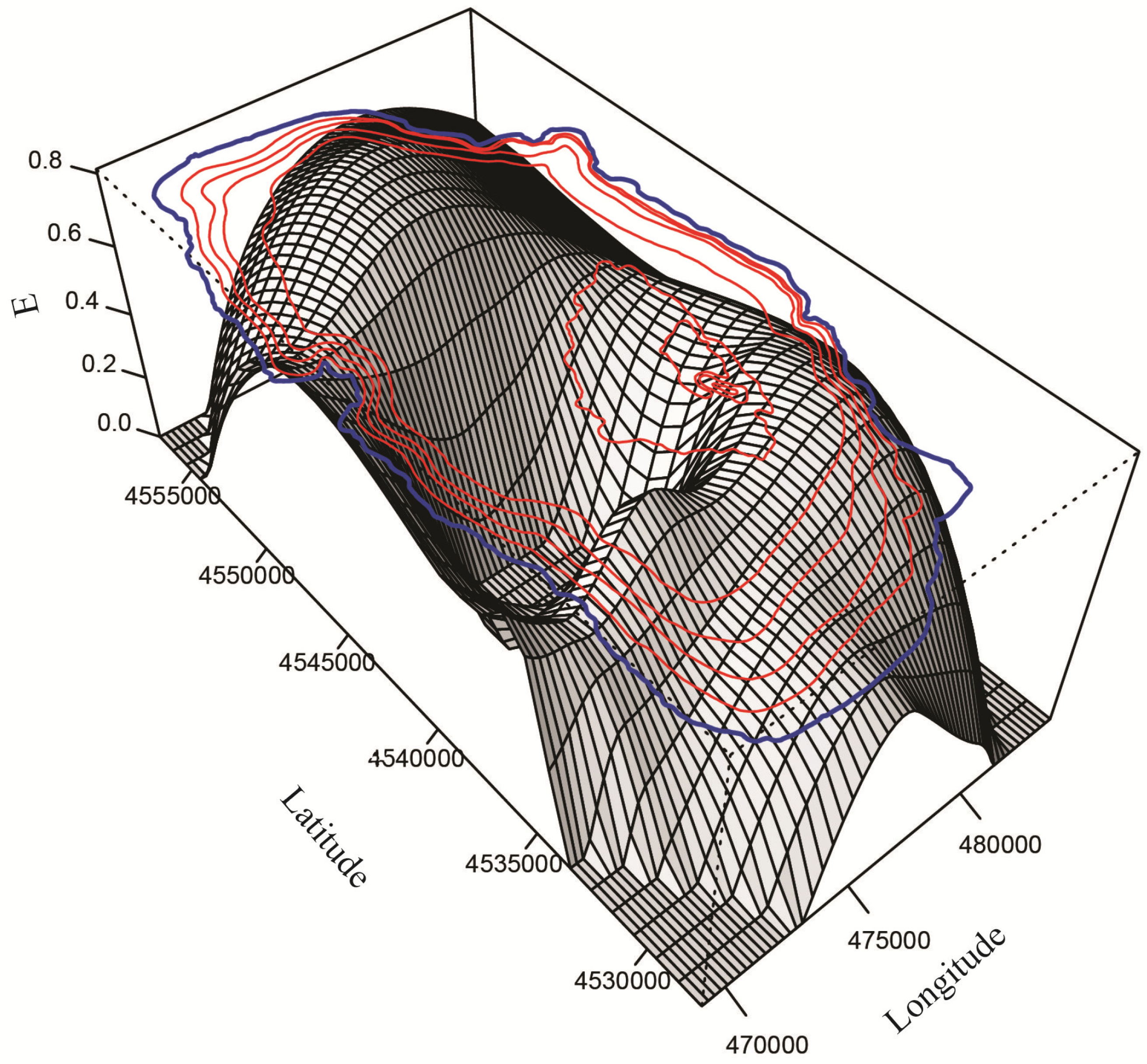
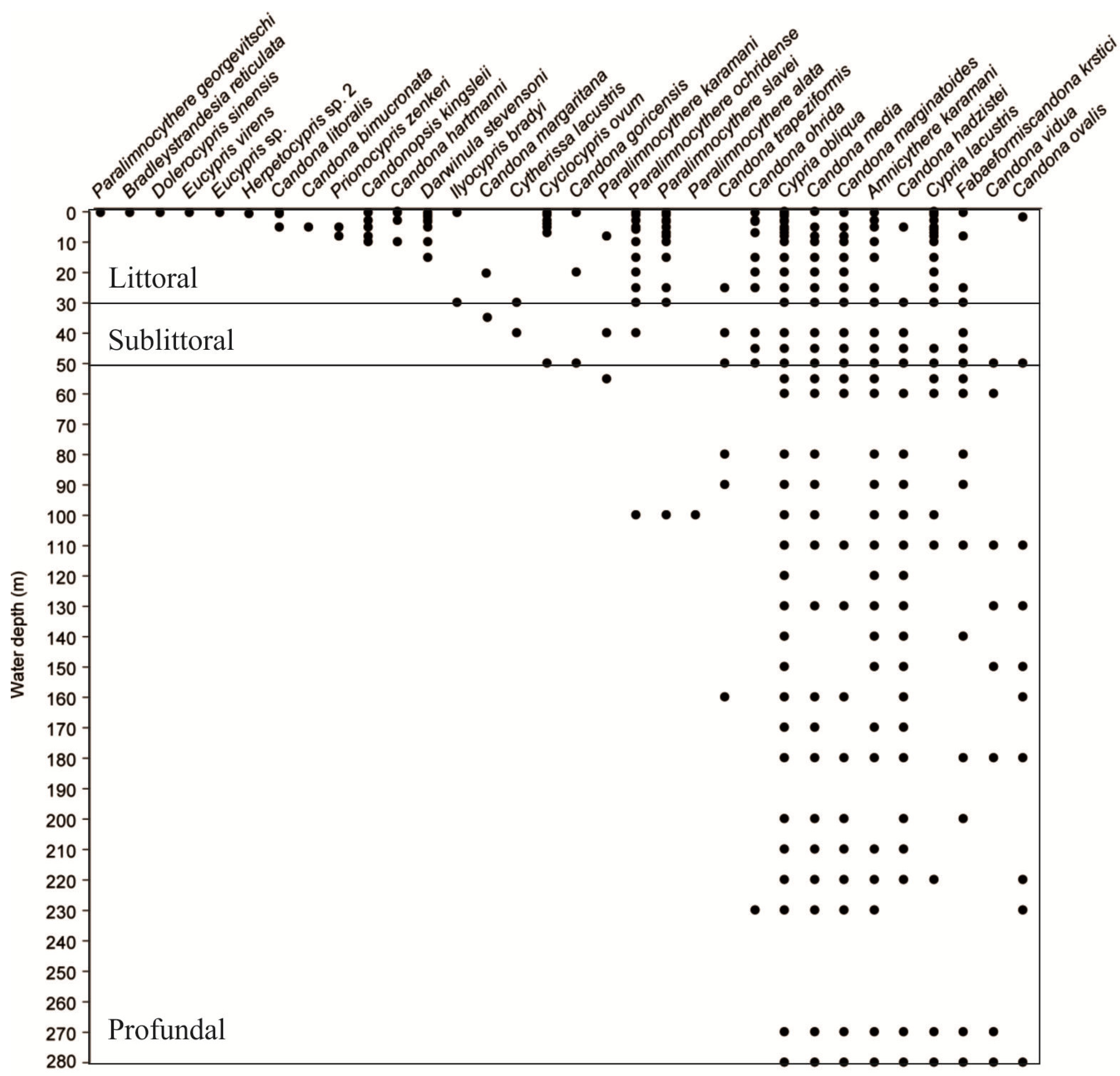
3.2.2. Vertical Distribution
3.2.2.1. Littoral Zone (≤30 m)
3.2.2.2. Sublittoral (30–50 m)
3.2.2.3. Profundal (50–280 m)
3.2.2.4. Littoral-Sublittoral Taxa (≤50 m)
3.2.2.5. Wide Depth-Range Taxa
3.3. Biogeography and Anthropogenic Impact on Ostracode Distribution
4. Conclusions
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Martens, K. Speciation in ancient lakes. Trends Ecol. Evol. 1997, 12, 177–182. [Google Scholar] [CrossRef] [PubMed]
- Schön, I.; Martens, K. Adaptive, pre-adaptive and non-adaptive components of radiations in ancient lakes: A review. Org. Divers. Evol. 2004, 4, 137–156. [Google Scholar] [CrossRef]
- Albrecht, C.; Trajanovski, S.; Kuhn, K.; Streit, B.; Wilke, T. Rapid evolution of an ancient lake species flock: Freshwater limpets (gastropoda: Ancylidae) in the balkan lake Ohrid. Org. Divers. Evol. 2006, 6, 294–307. [Google Scholar] [CrossRef]
- Von Rintelen, T.; Glaubrecht, M. Rapid evolution of sessility in an endemic species flock of the freshwater bivalve Corbicula from ancient lakes on sulawesi, Indonesia. Biol. Lett. 2006, 2, 73–77. [Google Scholar] [CrossRef] [PubMed]
- Lindhorst, K.; Vogel, H.; Krastel, S.; Wagner, B.; Hilgers, A.; Zander, A.; Schwenk, T.; Wessels, M.; Daut, G. Stratigraphic analysis of lake level fluctuations in lake Ohrid: An integration of high resolution hydro-acoustic data and sediment cores. Biogeosciences 2010, 7, 3651–3689. [Google Scholar] [CrossRef]
- Griffiths, H.I.; Reed, J.M.; Leng, M.J.; Ryan, S.; Petkovski, S. The recent palaeoecology and conservation status of balkan lake Dojran. Biol. Conserv. 2002, 104, 35–49. [Google Scholar] [CrossRef]
- Frogley, M.R.; Preece, R.C. A faunistic review of the modern and fossil molluscan fauna from lake pamvotis, ioannina, an ancient lake in NW Greece: Implications for endemism in the Balkans. In Balkan Biodiversity—Pattern and Process in the European Hotspot; Griffiths, H.I., Kryštufek, B., Reed, J.M., Eds.; Kluwer Academic: Dordrecht, The Netherlands, 2004; pp. 243–260. [Google Scholar]
- Reed, J.M.; Kryštufek, B.; Eastwood, W.J. The physical geography of the Balkans and nomenclature of place names. In Balkan Biodiversity—Pattern and Process in the European Hotspot; Griffiths, H.I., Kryštufek, B., Reed, J.M., Eds.; Kluwer Academic: Dordrecht, The Netherlands, 2004; pp. 9–22. [Google Scholar]
- Albrecht, C.; Hauffe, T.; Schreiber, K.; Trajanovski, S.; Wilke, T. Mollusc biodiversity and endemism in the potential ancient lake Trichonis, Greece. Malacologia 2009, 51, 357–375. [Google Scholar] [CrossRef]
- Albrecht, C.; Wilke, T. Ancient lake Ohrid: Biodiversity and evolution. Hydrobiologia 2008, 615, 103–140. [Google Scholar] [CrossRef]
- Trajanovski, S.; Albrecht, C.; Schreiber, K.; Schultheiß, R.; Stadler, T.; Benke, M.; Wilke, T. Testing the spatial and temporal framework of speciation in an ancient lake species flock: The leech genus Dina (hirudinea: Erpobdellidae) in lake Ohrid. Biogeosciences 2010, 7, 3387–3402. [Google Scholar] [CrossRef]
- Wagner, B.; Reicherter, K.; Daut, G.; Wessels, M.; Matzinger, A.; Schwalb, A.; Spirkovski, Z.; Sanxhaku, M. The potential of lake Ohrid for long-term palaeoenvironmental reconstructions. Palaeogeogr. Palaeoclimatol. Palaeoecol. 2008, 259, 341–356. [Google Scholar] [CrossRef]
- Belmecheri, S.; Namiotko, T.; Robert, C.; von Grafenstein, U.; Danielopol, D.L. Climate controlled ostracod preservation in lake Ohrid (Albania, Macedonia). Palaeogeogr. Palaeoclimatol. Palaeoecol. 2009, 277, 236–245. [Google Scholar] [CrossRef]
- Stankovič, S. The Balkan Lake Ohrid and Its Living World; W. Junk: Den Haag, The Netherlands, 1960; Volume IX, p. 357. [Google Scholar]
- Salemaa, H. Lake Ohrid. Arch. Hydrobiol. 1994, 44, 55–64. [Google Scholar]
- Klie, W. Zur kenntnis der ostracoden-gattung Limnocythere. Z Wiss Zool Abt A 1934, 3, 534–544. (In German) [Google Scholar]
- Klie, W. Studien über ostracoden aus dem Ohridsee: II. Limnocytherinae und Cytherinae. Arch. Hydrobiol. 1939, 35, 631–646. (In German) [Google Scholar]
- Klie, W. Studien über ostracoden aus dem Ohridsee: I. Candocyprinae. Arch. Hydrobiol. 1939, 35, 28–45. (In German) [Google Scholar]
- Klie, W. Studien über ostracoden aus dem Ohridsee: III. Erster nachtrag. Arch. Hydrobiol. 1942, 38, 254–259. (In German) [Google Scholar]
- Holmes, P.F. Ostracoda of lake Ohrid. Arch. Hydrobiol. 1937, 31, 484–500. [Google Scholar]
- Mikulić, F. Nove Candona vrste iz Ohridskog jezera. Bull Mus natn Hist nat 1961, 17, 87–107. (In German) [Google Scholar]
- Petkovski, T.K. Zur kenntnis der crustaceen des Prespasees. Fragm Balc. Mus. Maced. Sci. Nat. 1960, 3, 117–131. (In German) [Google Scholar]
- Petkovski, T.K. Zwei neue ostracoden aus dem Ohrid- und Prepasee. Izd Inst Piscic RP Maced 1960, 3, 57–66. (In German) [Google Scholar]
- Petkovski, T.K. Süsswasserostracoden aus Jugoslawien VII. Fragm Balc Mus Maced Sci. Nat. 1960, 3, 99–108. (In German) [Google Scholar]
- Petkovski, T.K. Zwei neue Limnocythere-arten aus Mazedonien (crustacea-ostracoda). Musei Maced. Sci. Nat. 1969, 12, 1–18. (In German) [Google Scholar]
- Petkovski, T.K. Einige neue und bemerkenswerte Candoninae aus dem Ohridsee und einigen anderen fundorten in Europa. Musei Maced. Sci. Nat. 1969, 11, 81–111. (In German) [Google Scholar]
- Mikulić, F.; Pljakić, M.A. Die merkmale der kvalitativen distribution der endemischen Candonaarten im Ochridsee. Ekologija 1970, 5, 101–115. (In German) [Google Scholar]
- Wagner, B.; Lotter, A.; Nowaczyk, N.; Reed, J.; Schwalb, A.; Sulpizio, R.; Valsecchi, V.; Wessels, M.; Zanchetta, G. A 40,000-year record of environmental change from ancient lake Ohrid (Albania and Macedonia). J. Paleolimnol. 2009, 41, 407–430. [Google Scholar] [CrossRef]
- Lorenschat, J.; Scharf, B.; Petkovski, T.; Viehberg, F.; Schwalb, A. Scientific collaboration on past speciation conditions in Ohrid (SCOPSCO): Recent and fossil ostracods from lake Ohrid as indicators of past environments: A coupled ecological and molecular genetic approach with deep-time perspective. Joannea Geologie und Palaontologie 2011, 11, 113–115. [Google Scholar]
- Lorenschat, J.; Schwalb, A. Autecology of the extant ostracod fauna of Lake Ohrid and adjacent waters—A key to paleoenvironmental reconstruction. Belg. J. Zool. 2013, 143, 42–68. [Google Scholar]
- Lindhorst, K.; Krastel, S.; Reicherter, K.; Stipp, M.; Wagner, B.; Schwenk, T. Sedimentary and Tectonic Evolution of Lake Ohrid (Macedonia/Albania). Basin Res. 2014. [Google Scholar] [CrossRef]
- Hauffe, T.; Albrecht, C.; Schreiber, K.; Birkhofer, K.; Trajanovski, S.; Wilke, T. Spatially explicit analysis of gastropod biodiversity in ancient Lake Ohrid. Biogeosciences 2011, 8, 175–188. [Google Scholar] [CrossRef]
- Spirkovski, Z.; Avramovski, O.; Kodzoman, A. Watershed management in the lake Ohrid region of Albania and Macedonia. Lakes Reserv. Res. Manag. 2001, 6, 237–242. [Google Scholar] [CrossRef]
- Wagner, B.; Wilke, T.; Krastel, S.; Zanchetta, G.; Sulpizio, R.; Reicherter, K.; Leng, M.; Grazhdani, A.; Trajanovski, S.; Levkov, Z.; Reed, J.; Wonik, T. Deep drilling of ancient Lake Ohrid capture over 1 million years of evolution and global climate cycling. EOS Trans. 2014, 95, 25–26. [Google Scholar] [CrossRef]
- Leng, M.J.; Baneschi, I.; Zanchetta, G.; Jex, C.N.; Wagner, B.; Vogel, H. Late quaternary palaeoenvironmental reconstruction from lakes Ohrid and Prespa (Macedonia/Albania border) using stable isotopes. Biogeosciences 2010, 7, 3109–3122. [Google Scholar] [CrossRef]
- Popovska, C.; Bonacci, O. Basic data on the hydrology of lakes Ohrid and Prespa. Hydrol. Process 2007, 21, 658–664. [Google Scholar] [CrossRef]
- Matzinger, A.; Spirkovski, Z.; Patceva, S.; Wüest, A. Sensitivity of ancient lake Ohrid to local anthropogenic impacts and global warming. J. Great Lakes Res. 2006, 32, 158–179. [Google Scholar] [CrossRef]
- Matter, M.; Anselmetti, F.S.; Jordanoska, B.; Wagner, B.; Wessels, M.; Wüest, A. Carbonate sedimentation and effects of eutrophication observed at the kališta subaquatic springs in lake Ohrid (Macedonia). Biogeosciences 2010, 7, 4715–4747. [Google Scholar] [CrossRef]
- Vogel, H.; Wessels, M.; Albrecht, C.; Stich, H.B.; Wagner, B. Spatial variability of recent sedimentation in lake Ohrid (Albania/Macedonia)—A complex interplay of natural and anthropogenic factors and their possible impact on biodiversity patterns. Biogeosciences 2010, 7, 3333–3342. [Google Scholar] [CrossRef]
- Talevska, M.; Petrovic, D.; Milosevic, D.; Talevski, T.; Maric, D.; Talevska, A. Biodiversity of macrophyte vegetation from lake Prespa, lake Ohrid and lake Skadar. Biotech. Biotech. Equip. 2007, 23, 931–935. [Google Scholar] [CrossRef]
- Data Publisher for Earth and Environmental Science. Available online: http://doi.pangaea.de/doi:10.1594/PANGAEA.819871?format=html (accessed on 3 July 2014).
- Meisch, C. Freshwater Ostracoda of Western and Central Europe; Spektrum Akademischer Verlag: Berlin, Germany, 2000; p. 522. [Google Scholar]
- Namiotko, T.; Danielopol, D.L.; Belmecheri, S.; Gross, M.; von Grafenstein, U. On the Leptocytheridae Ostracods of the Long-Lived Lake Ohrid: A Reappraisal of their Taxonomic Assignment and Biogeographic Origin. Int. Rev. Hydrobiol. 2012, 97, 356–374. [Google Scholar] [CrossRef]
- Petkovski, T.K.; Scharf, B.; Keyser, D. New and little known species of the genus Candona (Crustacea, Ostracoda) from Macedonia and other Balkan areas. Limnologica 2002, 32, 114–130. [Google Scholar] [CrossRef]
- Krebs, C.J. Ecological Methodology; Harper and Row Publishers: New York, NY, USA, 1989. [Google Scholar]
- Magurran, A. Ecological Diversity and Its Measurement; Princeton Universuty Press: Princeton, NJ, USA, 1988. [Google Scholar]
- Cleveland, W.S.; Devlin, S.J. Locally weighted regression: An approach to regression analysis by local fitting. J. Am. Stat. Assoc. 1988, 83, 596–610. [Google Scholar] [CrossRef]
- Wood, S. Generalized Additive Models: An Introduction with R, 1st ed.; Chapman and Hall: London, UK, 2006. [Google Scholar]
- R Core Team. R: A Language and Environment for Statistical Computing, R Foundation for Statistical Computing: Vienna, Austria, 2009.
- Karanovic, I. Recent Freshwater Ostracods of the World. Crustacea, Ostracoda, Podocopida; Springer: Berlin, Germany, 2012; p. 608. [Google Scholar]
- Frogley, M.R.; Griffiths, H.I.; Martens, K.; Holmes, J.A.; Chivas, A.R. Modern and fossil ostracods from ancient lakes. In The Ostracoda: Applications in Quaternary Research; Holmes, J.A., Chivas, A.R., Eds.; American Geophysical Union: Washington, DC, USA, 2002; pp. 167–184. [Google Scholar]
- Gliozzi, E.; Grossi, F. Late messinian lago-mare ostracod palaeoecology: A correspondence analysis approach. Palaeogeogr. Palaeoclimatol. Palaeoecol. 2008, 264, 288–295. [Google Scholar] [CrossRef]
- Martens, K. Ostracod speciation in ancient lakes: A review. In Speciation in Ancient Lakes; Martens, K., Goddeeris, B., Coulter, G., Eds.; Archiv für Hydrobiologie: Stuttgart, Germany, 1994; Volume 44, pp. 203–222. [Google Scholar]
- Martens, K.; Schön, I. Crustacean biodiversity in ancient lakes: A review. Crustaceana 1999, 72, 899–910. [Google Scholar] [CrossRef]
- Griffiths, H.I.; Frogley, M.R. Fossil ostracods, faunistics and the evolution of regional biodiversity. In Balkan Biodiversity—Pattern and Process in the European Hotspot; Griffiths, H.I., Krystufek, B., Reed, J.M., Eds.; Kluwer Academic Publishers: Dordrecht, The Netherlands, 2004; pp. 261–272. [Google Scholar]
- Griffiths, H.I.; Brancelj, A. The (palaeo-)biogeography of Candona bimucronata: An unusual candonid ostracod from the Balkans. Crustaceana 1996, 69, 882–889. [Google Scholar] [CrossRef]
- Klie, W. Krebstiere Oder Crustacea III: Ostracoda, Muschelkrebse. Die Tierwelt Deutschlands und der Angrenzenden Meeresteile; Verlag Von Gustav Fischer: Jena, Germany, 1938; p. 230. (In German) [Google Scholar]
- Karanovic, I.; Petkovski, T.K. Two interesting ostracod species from Montenegro (SE Europe). Ann. Limnol. 1999, 35, 123–132. [Google Scholar] [CrossRef]
- Pérez, L.; Lorenschat, J.; Bugja, R.; Brenner, M.; Scharf, B.; Schwalb, A. Distribution, diversity and ecology of modern freshwater ostracodes (Crustacea), and hydrochemical characteristics of Lago Petén Itzá, Guatemala. J. Limnol. 2010, 69, 146–159. [Google Scholar] [CrossRef]
- Pérez, L.; Lorenschat, J.; Massaferro, J.; Pailles, C.; Sylvestre, F.; Hollwedel, W.; Brandorff, G.-O.; Brenner, M.; Islebe, G.; Lozano, M.S. Bioindicators of climate and trophic state in lowland and highland aquatic ecosystems of the Northern Neotropicos. Revista de Biología Tropical 2013, 61, 603–644. [Google Scholar] [PubMed]
- Pérez, L.; Frenzel, P.; Brenner, M.; Escobar, J.; Hoelzmann, P.; Scharf, B.; Schwalb, A. Late Quaternary (24–10 ka BP) environmental history of the Neotropical lowlands inferred from ostracodes in sediments of Lago Petén Itzá, Guatemala. J. Paleolimnol. 2011, 46, 59–74. [Google Scholar] [CrossRef]
- Külköylüoğlu, O.; Sari, N. Ecological characteristics of the freshwater ostracoda in Bolu region (Turkey). Hydrobiologia 2012, 688, 37–46. [Google Scholar] [CrossRef]
- Kostoski, G.; Albrecht, C.; Trajanovski, S.; Wilke, T. A freshwater biodiversity hotspot under pressure—Assessing threats and identifying conservation needs for ancient lake Ohrid. Biogeosciences 2010, 7, 3999–4015. [Google Scholar] [CrossRef]
- Pieri, V.; Vandekerkhove, J.; Goi, D. Ostracoda (Crustacea) as indicators for surface water quality: A case study from the ledra river basin (NE Italy). Hydrobiologia 2012, 688, 25–35. [Google Scholar] [CrossRef]
© 2014 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/3.0/).
Share and Cite
Lorenschat, J.; Pérez, L.; Correa-Metrio, A.; Brenner, M.; Von Bramann, U.; Schwalb, A. Diversity and Spatial Distribution of Extant Freshwater Ostracodes (Crustacea) in Ancient Lake Ohrid (Macedonia/Albania). Diversity 2014, 6, 524-550. https://doi.org/10.3390/d6030524
Lorenschat J, Pérez L, Correa-Metrio A, Brenner M, Von Bramann U, Schwalb A. Diversity and Spatial Distribution of Extant Freshwater Ostracodes (Crustacea) in Ancient Lake Ohrid (Macedonia/Albania). Diversity. 2014; 6(3):524-550. https://doi.org/10.3390/d6030524
Chicago/Turabian StyleLorenschat, Julia, Liseth Pérez, Alexander Correa-Metrio, Mark Brenner, Ullrich Von Bramann, and Antje Schwalb. 2014. "Diversity and Spatial Distribution of Extant Freshwater Ostracodes (Crustacea) in Ancient Lake Ohrid (Macedonia/Albania)" Diversity 6, no. 3: 524-550. https://doi.org/10.3390/d6030524
APA StyleLorenschat, J., Pérez, L., Correa-Metrio, A., Brenner, M., Von Bramann, U., & Schwalb, A. (2014). Diversity and Spatial Distribution of Extant Freshwater Ostracodes (Crustacea) in Ancient Lake Ohrid (Macedonia/Albania). Diversity, 6(3), 524-550. https://doi.org/10.3390/d6030524