Biodiversity Indicators Show Climate Change Will Alter Vegetation in Parks and Protected Areas



Abstract
:1. Introduction
2. Study Area
3. Data
3.1. The Dynamic Habitat Index
3.2. Protected Area and Ecological Classification Data
4. Methods
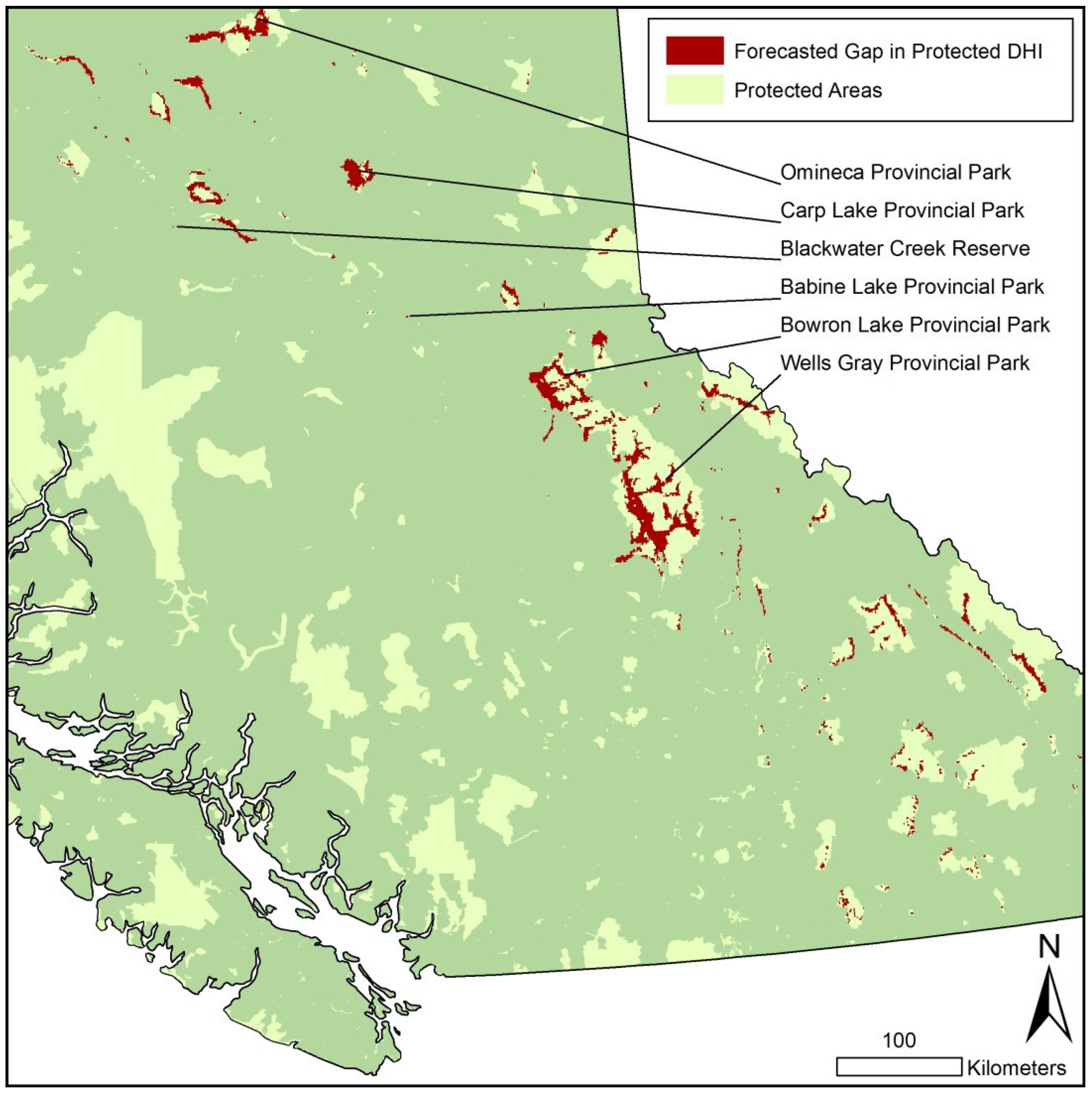
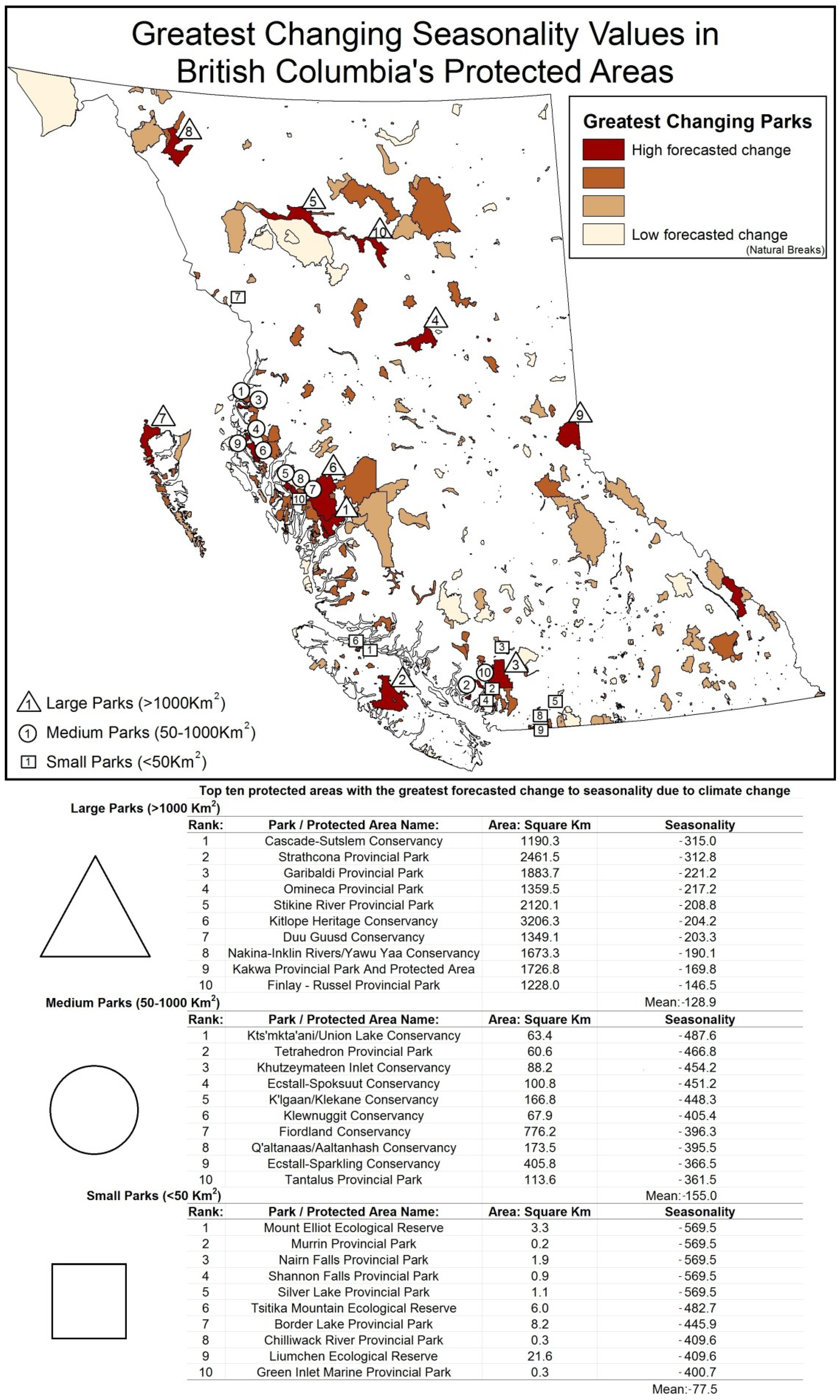
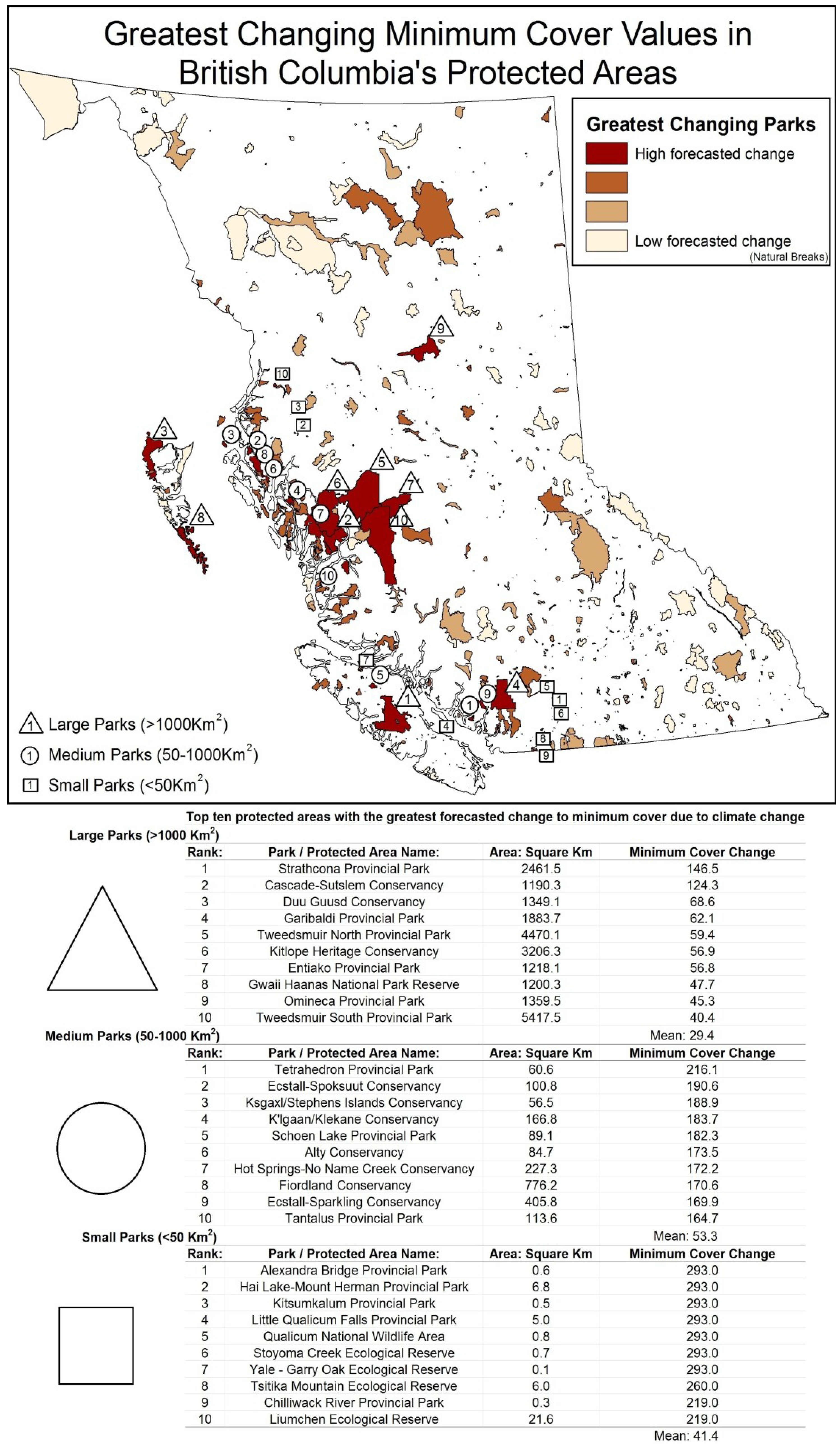
5. Results




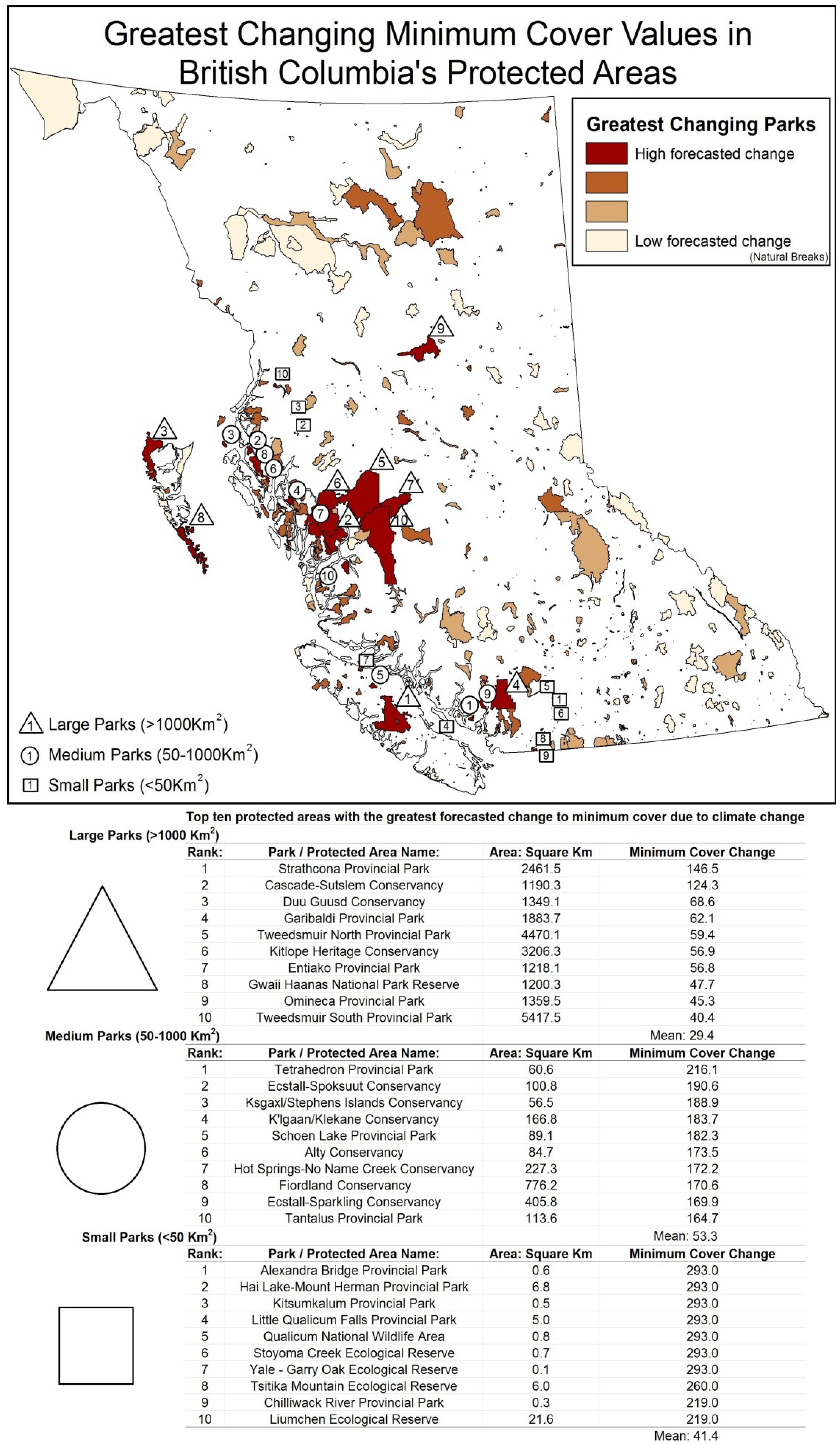

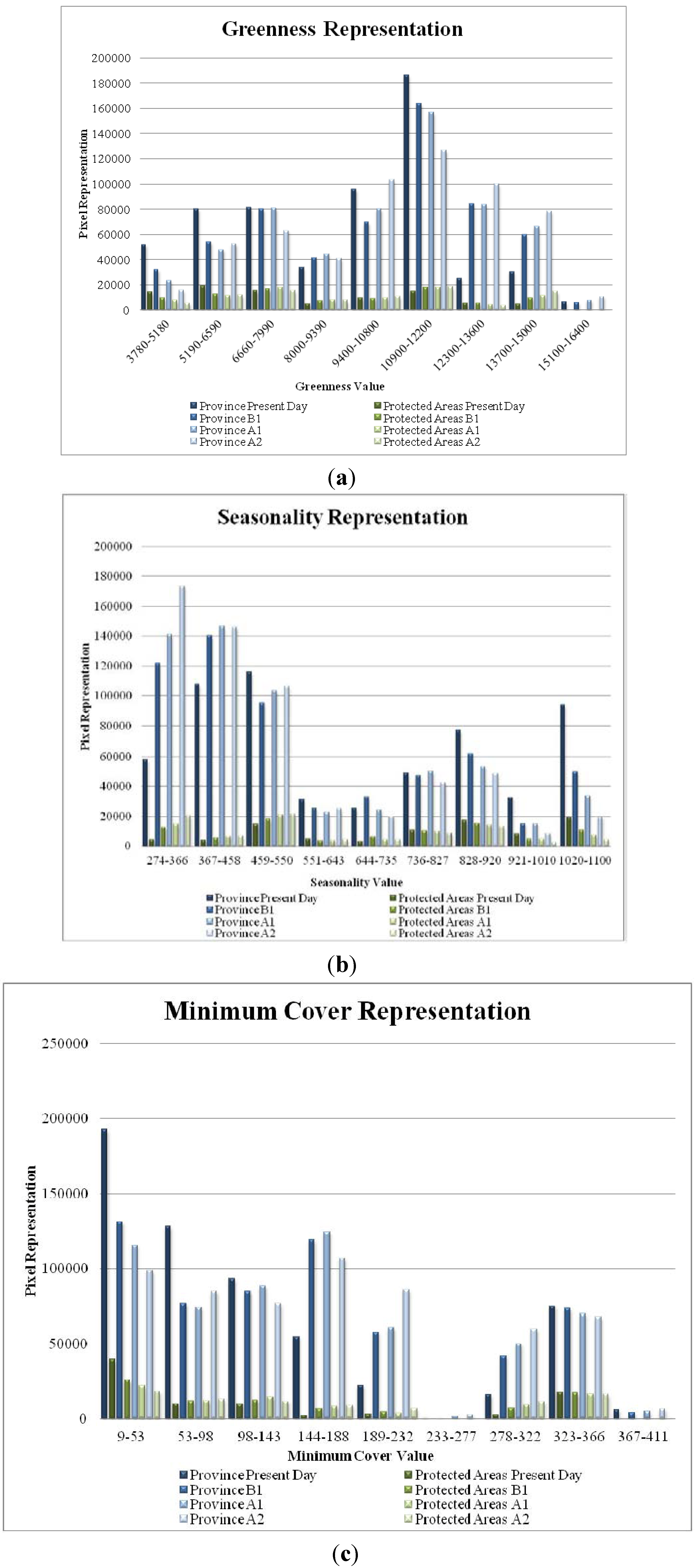{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Rank | BEC Zone | Mean Greenness Change | ||
| High Change Zone |  | 1 | Mountain Hemlock | 3437.4 |
| 2 | Coastal Mountain-heather Alpine | 2207.7 | ||
| 3 | Interior Mountain-heather Alpine | 1781.0 | ||
| 4 | Coastal Western Hemlock | 1712.7 | ||
| 5 | Engelmann Spruce - Subalpine Fir | 1466.8 | ||
| 6 | Sub-Boreal Spruce | 786.9 | ||
| 7 | Boreal White and Black Spruce | 759.6 | ||
| 8 | Interior Cedar - Hemlock | 676.3 | ||
| 9 | Boreal Altai Fescue Alpine | 613.9 | ||
| 10 | Bunchgrass | −498.5 | ||
| 11 | Interior Douglas-fir | −431.6 | ||
| 12 | Spruce - Willow - Birch | 407.3 | ||
| Low Change Zone | 13 | Montane Spruce | 227.8 | |
| 14 | Ponderosa Pine | −107.4 | ||
| 15 | Sub-Boreal Pine - Spruce | 53.3 | ||
| 16 | Coastal Douglas-fir | 0.0 | ||
| Rank | BEC Zone | Mean Seasonality Change | ||
| High Change Zone | | 1 | Mountain Hemlock | −257.4 |
| 2 | Coastal Western Hemlock | −192.4 | ||
| 3 | Spruce - Willow - Birch | −178.9 | ||
| 4 | Engelmann Spruce - Subalpine Fir | −165.4 | ||
| 5 | Coastal Mountain-heather Alpine | −134.1 | ||
| 6 | Interior Cedar - Hemlock | −106.5 | ||
| 7 | Sub-Boreal Spruce | −95.1 | ||
| 8 | Montane Spruce | −87.3 | ||
| 9 | Interior Mountain-heather Alpine | −63.1 | ||
| 10 | Boreal Altai Fescue Alpine | −61.5 | ||
| 11 | Boreal White and Black Spruce | −56.9 | ||
| 12 | Sub-Boreal Pine - Spruce | −48.4 | ||
| Low Change Zone | 13 | Interior Douglas-fir | −24.8 | |
| 14 | Ponderosa Pine | −0.3 | ||
| 15 | Bunchgrass | −0.2 | ||
| 16 | Coastal Douglas-fir | 0.0 | ||
| Rank | BEC Zone | Mean Minimum Cover Change | ||
| High Change Zone | | 1 | Mountain Hemlock | 90.8 |
| 2 | Coastal Western Hemlock | 87.8 | ||
| 3 | Sub-Boreal Pine - Spruce | 57.4 | ||
| 4 | Sub-Boreal Spruce | 53.9 | ||
| 5 | Coastal Douglas-fir | 48.5 | ||
| 6 | Montane Spruce | 41.3 | ||
| 7 | Interior Cedar - Hemlock | 41.1 | ||
| 8 | Coastal Mountain-heather Alpine | 41.0 | ||
| 9 | Boreal White and Black Spruce | 40.8 | ||
| 10 | Interior Douglas-fir | 39.4 | ||
| 11 | Engelmann Spruce - Subalpine Fir | 16.7 | ||
| 12 | Spruce - Willow - Birch | 14.7 | ||
| Low Change Zone | 13 | Bunchgrass | 12.9 | |
| 14 | Interior Mountain-heather Alpine | 7.4 | ||
| 15 | Ponderosa Pine | 4.3 | ||
| 16 | Boreal Altai Fescue Alpine | 4.1 |
6. Discussion
7. Conclusions
Acknowledgments
Conflict of Interest
References and Notes
- Barton, J.H. Biodiversity at Rio. BioScience 1992, 42, 773–776. [Google Scholar] [CrossRef]
- Aichi Biodiversity Target. Available online: http://www.cbd.int/sp/targets/ (accessed on 2 March 2013).
- Svancara, L.K.; Brannon, R.; Scott, J.M.; Groves, C.R.; Noss, R.F.; Pressey, R.L. Policy-Driven versus evidence-based conservation: A review of political targets and biological needs. BioScience 2005, 55, 989. [Google Scholar] [CrossRef]
- Andrew, M.E.; Wulder, M.A.; Coops, N.C. Patterns of protection and threats along productivity gradients in Canada. Biol. Conserv. 2011, 144, 2891–2901. [Google Scholar] [CrossRef]
- Pressey, R.L.; Cabeza, M.; Watts, M.E.; Cowling, R.M.; Wilson, K.A. Conservation planning in a changing world. Trends Ecol. Evol. 2007, 22, 583–592. [Google Scholar] [CrossRef]
- Kerr, J.T.; Deguise, I. Habitat loss and the limits to endangered species recovery. Ecol. Lett. 2004, 7, 1163–1169. [Google Scholar] [CrossRef]
- Hannah, L.; Midgley, G.F.; Sandy, A.; Araujo, M.B.; Hughes, G.; Enrique, M.-M.; Richard, P.; Paul, W. Protected area needs in a changing climate. Front. Ecol. Environ. 2007, 5, 131–138. [Google Scholar] [CrossRef]
- Lemieux, C.J.; Scott, D.J. Climate change, biodiversity conservation and protected area planning in Canada. Can. Geogr./Le Geogr. Can. 2005, 49, 384–397. [Google Scholar] [CrossRef]
- Visser, M.E. Keeping up with a warming world; assessing the rate of adaptation to climate change. Proc. R. Soc. 2008, 275, 649–659. [Google Scholar] [CrossRef]
- Harrison, P.; Berry, P.; Butt, N.; New, M. Modelling climate change impacts on species’ distributions at the European scale: Implications for conservation policy. Environ. Sci. Policy 2006, 9, 116–128. [Google Scholar] [CrossRef]
- Berry, P.M.; Dawson, T.P.; Harrison, P.A.; Pearson, R.G. Modelling potential impacts of climate change on the bioclimatic envelope of species in Britain and Ireland. Glob. Ecol. Biogeogr. 2002, 11, 453–462. [Google Scholar] [CrossRef]
- Midgley, G. Developing regional and species-level assessments of climate change impacts on biodiversity in the Cape Floristic Region. Biol. Conserv. 2003, 112, 87–97. [Google Scholar] [CrossRef]
- Halpin, P. Global climate change and natural-area protection: Management responses and research directions. Ecol. Appl. 1997, 7, 828–843. [Google Scholar] [CrossRef]
- United Nations Environment Programme (UNEP), Decisions Adopted by the Conference of the Parties to the Convention on Biological Diversity at its Seventh Meeting; UNEP: Kuala Lumpur, Malaysia, 2004.
- Environment Canada, Canadian Protected Areas Status Report; Environment Canada: Gatineau, QC, Canada, 2006.
- Canadian Biodiversity Strategy. Available online: http://www.cbin.ec.gc.ca/strategie-strategy (accessed on 22 October 2010).
- Iverson, L.R.; Prasad, A.M. Predicting abundance of 80 tree species following climate change in the eastern United States. Ecol. Monogr. 1998, 68, 465–485. [Google Scholar] [CrossRef]
- Fussel, H.M. Adaptation planning for climate change: Concepts, assessment approaches, and key lessons. Sustain. Sci. 2007, 2, 265–275. [Google Scholar] [CrossRef]
- Willis, S.G.; Hole, D.G.; Collingham, Y.C.; Hilton, G.; Rahbek, C.; Huntley, B. Assessing the impacts of future climate change on protected area networks: A method to simulate individual species’ responses. Environ. Manag. 2009, 43, 836–845. [Google Scholar]
- Hannah, L.; Midgley, G.F.; Lovejoy, T.; Bond, W.J.; Bush, M.; Lovett, J.C.; Scott, D.; Woodward, F.I. Conservation of biodiversity in a changing climate. Conserv. Biol. 2002, 16, 264–268. [Google Scholar] [CrossRef]
- Hawkins, B.A.; Field, R.; Cornell, H.V.; Currie, D.; Guégan, J.F.; Kaufman, D.M.; Kerr, J.T.; Mittelbach, G.G.; Oberdorff, T.; O’Brien, E.M.; et al. Energy, water, and broad-scale geographic patterns of species richness. Ecology 2003, 84, 3105–3117. [Google Scholar] [CrossRef]
- Araújo, M.B.; Rahbek, C. How does climate change affect biodiversity? Science 2006, 313, 1396–1397. [Google Scholar]
- Nagendra, H. Using remote sensing to assess biodiversity. Int. J. Remote Sens. 2001, 22, 2377–2400. [Google Scholar] [CrossRef]
- Turner, W.; Spector, S.; Gardiner, N. Remote sensing for biodiversity science and conservation. Trends Ecol. Evol. 2003, 18, 306–314. [Google Scholar]
- Waide, R.B.; Willig, M.R.; Steiner, C.F.; Mittelbach, G.; Gough, L.; Dodson, S.I.; Juday, G.P.; Parmenter, R. The relationship between productivity and species richness. Ann. Rev. Ecol. Syst. 1999, 30, 257–300. [Google Scholar] [CrossRef]
- Latta, G.; Hailemariam, T.; Barrett, T. Mapping and imputing potential productivity of Pacific Northwest forests using climate variables. Can. J. For. Res. 2009, 39, 1197–1207. [Google Scholar] [CrossRef]
- Running, S.W.; Hunt, E.R. Generalization of a Forest Ecosystem Process Model for Other Biomes, BIOME-BGC, and an Application for Global-Scale Models; San Diego Academic press: San Diego, MA, USA, 1993; pp. 141–158. [Google Scholar]
- Shilling, F. Do habitat conservation plans protect endangered species? Science 1997, 276, 1662–1663. [Google Scholar] [CrossRef]
- Algar, A.C.; Kharouba, H.M.; Young, E.R.; Kerr, J.T. Predicting the future of species diversity: Macroecological theory, climate change, and direct tests of alternative forecasting methods. Ecography 2009, 32, 22–33. [Google Scholar] [CrossRef]
- Fitterer, J.L.; Nelson, T.A.; Coops, N.C.; Wulder, M.A. Modelling the ecosystem indicators of British Columbia using Earth observation data and terrain indices. Ecol. Indic. 2012, 20, 151–162. [Google Scholar]
- Fontana, F.M.; Coops, N.C.; Khlopenkov, K.V.; Trishchenko, A.P.; Riffler, M.; Wulder, M.A. Generation of a novel 1km NDVI data set over Canada, the northern United States, and Greenland based on historical AVHRR data. Remote Sens. Environ. 2012, 121, 171–185. [Google Scholar]
- Slayback, D.A.; Pinzon, J.E.; Los, S.O.; Tucker, C.J. Northern hemisphere photosynthetic trends 1982–99. Glob. Change Biol. 2003, 9, 1–15. [Google Scholar] [CrossRef]
- Xiao, J.; Moody, A. Geographical distribution of global greening trends and their climatic correlates: 1982–1998. Int. J. Remote Sens. 2005, 26, 2371–2390. [Google Scholar] [CrossRef]
- Waser, L.T.; Stofer, S.; Schwarz, M.; Küchler, M.; Ivits, E.; Scheidegger, C. Prediction of biodiversity—regression of lichen species richness on remote sensing data. Community Ecol. 2004, 5, 121–133. [Google Scholar] [CrossRef]
- Puumalainen, J.; Kennedy, P.; Folving, S. Monitoring forest biodiversity: A European perspective with reference to temperate and boreal forest zone. J. Environ. Manag. 2003, 67, 5–14. [Google Scholar] [CrossRef]
- Fitterer, J.L.; Nelson, T.A.; Coops, N.C.; Wulder, M.A.; Mahony, N.A. Exploring the ecological processes driving geographical patterns of breeding bird richness in British Columbia, Canada. Ecol. Appl. 2012, 20, 151–162. [Google Scholar]
- Running, S.W.; Nemani, R.R. Relating seasonal patterns of the AVHRR vegetation index to simulated photosynthesis and transpiration of forests in different climates. Remote Sens. Environ. 1988, 24, 347–367. [Google Scholar] [CrossRef]
- Zhang, X.; Tarpley, D.; Sullivan, J.T. Diverse responses of vegetation phenology to a warming climate. Geophys. Res. Lett. 2007, 34, 1–5. [Google Scholar]
- Herrmann, S.; Anyamba, A.; Tucker, C. Recent trends in vegetation dynamics in the African Sahel and their relationship to climate. Glob. Environ. Change Part A 2005, 15, 394–404. [Google Scholar] [CrossRef]
- Kawabata, A.; Ichii, K.; Yamaguchi, Y. Global monitoring of interannual changes in vegetation activities using NDVI and its relationships to temperature and precipitation. Int. J. Remote Sens. 2001, 22, 1377–1382. [Google Scholar]
- Nemani, R.R.; Keeling, C.D.; Hashimoto, H.; Jolly, W.M.; Piper, S.C.; Tucker, C.J.; Myneni, R.B.; Running, S.W. Climate-driven increases in global terrestrial net primary production from 1982 to 1999. Science 2003, 300, 1560. [Google Scholar] [CrossRef]
- Churkina, G.; Running, S. Investigating the balance between timber harvest and productivity of global coniferous forests under global change. Clim. Change 2000, 47, 167–191. [Google Scholar] [CrossRef]
- Hall, R.J.; Raulier, F.; Price, D.T.; Arsenault, E.; Bernier, P.Y.; Case, B.S.; Guo, X. Integrating remote sensing and climate data with process-based models to map forest productivity within west-central Alberta’s boreal forest: Ecoleap-West 1. For. Chronicle 2006, 82, 159–176. [Google Scholar]
- Holmes, K. Forecasting impacts of climate change on indicators of British Columbia’s Biodiversity. MSc thesis, University of Victoria, Victoria, BC, Canada, 2012. [Google Scholar]
- Austin, M.A.; Buffett, D.A.; Nicolson, D.J.; Scudder, G.G.E.; Stevens, V. Taking Nature’s Pulse: The Status of Biodiversity in British Columbia; publisher: Victoria, BC, Canada, 2008; p. 268. Available online: http://www.biodiversitybc.org/ (accessed on 5 March 2013).
- Kerr, J.; Cihlar, J. Patterns and causes of species endangerment in Canada. Ecol. Appl. 2004, 14, 743–753. [Google Scholar] [CrossRef]
- Mote, P.W.; Parson, E.A.; Hamlet, A.F.; Keeton, W.S.; Lettenmaier, D.; Mantua, N.; Miles, E.L.; Peterson, D.W.; Peterson, D.L.; Slaughter, R.; et al. Preparing for climatic change: The water, salmon, and forests of the Pacific Northwest. Clim. Change 2003, 61, 45–88. [Google Scholar] [CrossRef]
- Coops, N.C.; Wulder, M.A.; Duro, D.; Han, T.; Berry, S. The development of a Canadian dynamic habitat index using multi-temporal satellite estimates of canopy light absorbance. Ecol. Indic. 2008, 8, 754–766. [Google Scholar] [CrossRef]
- Summary of the Park and Protected Areas System. BC Parks, Ministry of Environment. Available online: http://www.env.gov.bc.ca/bcparks/aboutBCParks/prk_desig.html/ (accessed on 5 March 2013).
- Coops, N.C.; Wulder, M.A.; Iwanicka, D. Demonstration of a satellite-based index to monitor habitat at continental-scales. Ecol. Indic. 2009, 9, 948–958. [Google Scholar] [CrossRef]
- Andrew, M.E.; Wulder, M.A.; Coops, N.C.; Baillargeon, G. Beta-diversity gradients of butterflies along productivity axes. Glob. Ecol. Biogeogr. 2012, 21, 352–364. [Google Scholar] [CrossRef]
- Pettorelli, N.; Vik, J.O.; Mysterud, A.; Gaillard, J.-M.; Tucker, C.J.; Stenseth, N.C. Using the satellite-derived NDVI to assess ecological responses to environmental change. Trends Ecol. Evol. 2005, 20, 503–510. [Google Scholar] [CrossRef]
- Berry, S.; Mackey, B.; Brown, T. Potential applications of remotely sensed vegetation greenness to habitat analysis and the conservation of dispersive fauna. Pac. Conserv. Biol. 2007, 13, 120–127. [Google Scholar]
- Liu, J.; Cihlar, J.; Chen, W. Net primary productivity distribution in the BOREAS region from a process model using satellite and surface data. J. Geophys. Res. 1999, 104, 27735–27754. [Google Scholar] [CrossRef]
- Huston, M. A general hypothesis of species diversity. Am. Soc. Nat. 1979, 113, 81–101. [Google Scholar]
- Tilman, D.; Wedin, D.; Knops, J. Productivity and sustainability influenced by biodiversity in grassland ecosystems. Nature 1996, 379, 718–720. [Google Scholar] [CrossRef]
- Los, S.O.; Collatz, G.J.; Bounoua, L.; Sellers, P.J.; Tucker, C.J. Global interannual variations in sea surface temperature and land surface vegetation, air temperature, and precipitation. J. Clim. 2001, 14, 1535–1549. [Google Scholar] [CrossRef]
- Shaver, G.R.; Chapin, F., III; Gartner, B.L. Factors limiting seasonal growth and peak biomass accumulation in Eriophorum. vaginatum in Alaskan tussock tundra. J. Ecol. 1986, 74, 257–278. [Google Scholar] [CrossRef]
- Breiman, L.; Friedman, J.H.; Olshen, R.A.; Stone, C.G. Classification and Regression Trees; Wadsworth International Group: Belmont, CA, USA, 1984. [Google Scholar]
- Wang, T.; Hamann, A.; Splittlehouse, D. ClimateWNA: A program to generate high-resolution climate data for climate change studies and applications in western North America. Available online: http://www.genetics.forestry.ubc.ca/cfcg/ClimateWNA/help.htm/ (accessed on 21 March 2011).
- Murdock, T.; Bürger, G. Research Plan for Regional Climate Impacts. Pacific Climate Impacts Consortium 2010. Available online: http://www.pacificclimate.org/ (accessed on 21 March 2011).
- Monserud, R.A.; Yang, Y.; Huang, S.; Tchebakova, N. Potential change in lodgepole pine site index and distribution under climate change in Alberta. Can. J. For. Res. 2008, 38, 343–352. [Google Scholar] [CrossRef]
- Mote, P.; Salathé, E.; Peacock, C. Scenarios of future climate for the Pacific Northwest. Climate Impacts Group. University of Washington, 2005. Available online: http://www.cses.washington.edu/db/pdf/kc05scenarios462.pdf (accessed on 5 March 2013).
- GeoBC British Columbia Data Distribution Service. Available online: https://www.apps.gov.bc.ca/pub/dwds/home.so/ (accessed on 29 July 2012).
- B.C. Ministry of Forests Biogeoclimatic Zones of British Columbia. Available online: http://www.for.gov.bc.ca/hfd/library/documents/treebook/biogeo/biogeo.htm/ (accessed 21 March 2011).
- Ministry of Environment. Preparing for Climate Change, British Columbia’s Adaptation Strategy; Government of British Columbia: Victoria, BC, Canada, 2010. Available online: http://www.livesmartbc.ca/attachments/Adaptation_Strategy.pdf (accessed on 5 March 2013).
- Lemieux, C.J.; Beechey, T.J.; Gray, P.A. Prospects for Canada’s protected areas in an era of climate change. Land Use Policy 2011, 28, 928–941. [Google Scholar] [CrossRef]
- Johnson, C.J.; Parker, K.L.; Heard, D.C.; Seip, D.R. Movements, foraging habits, and habitat use strategies of northern woodland caribou during winter: Implications for forest practices in British Columbia. JEM 2004, 5, 22–35. [Google Scholar]
- Food and Agriculture Organization. Assessing forest degradation: Towards the Development of Globally Applicable Guidelines; Food Resources Assessment Working Paper 177; Food and Agriculture Organization: Rome, Italy, 2011; p. 99. Available online: http://www.fao.org/docrep/015/i2479e/i2479e00.pdf (accessed on 5 March 2013).
- Flather, C.; Wilson, K. Identifying gaps in conservation networks: Of indicators and uncertainty in geographic-based analyses. Ecol. Appl. 1997, 7, 531–542. [Google Scholar] [CrossRef]
- Ministry of Lands Parks and Housing. Wells Gray Provincial Park Master Plan. Province of British Columbia: Victoria, BC, Canada, 1986; p. 55. Available online: http://www.env.gov.bc.ca/bcparks/planning/mgmtplns/wellsgray/wells_gray_mp.pdf (accessed on 5 March 2013).
- Ministry of Water, Land and Air Protection. Management Direction for Chopaka. East. Site, South. Okanagan Grasslands Protected Area; Province of British Columbia: Victoria, BC, Canada, 2003. Available online: http://www.env.gov.bc.ca/bcparks/planning/mgmtplns/s_okanpa/chopeast.pdf (accessed on 5 March 2013).
- Klanderud, K. Climate change effects on species interactions in an alpine plant community. J. Ecol. 2005, 93, 127–137. [Google Scholar] [CrossRef]
- Brink, V. A directional change in the subalpine forest-heath ecotone in Garibaldi Park, British Columbia. Ecology 1959, 40, 10–16. [Google Scholar] [CrossRef]
- Wang, T.; Campbell, E.M.; O’Neill, G.A.; Aitken, S.N. Projecting future distributions of ecosystem climate niches: Uncertainties and management applications. For. Ecol. Manag. 2012, 279, 128–140. [Google Scholar] [CrossRef]
- Williams Creek Ecological Reserve—Detailed Description. BC Parks, Ministry of Environment. Available online: http://www.env.gov.bc.ca/bcparks/eco_reserve/williamscrk_er.html/ (accessed on 5 March 2013).
- Kerr, J.T. Global Change Impacts on Biodiversity: The View from Canada. In Coast to Coast Seminar Series; Sage: Ottawa, ON, Canada, 2012. [Google Scholar]
- Garry Oak Ecosystems Recovery Team. Species at Risk. Available online: http://www.goert.ca/about/species_at_risk.php/ (accessed on 5 March 2013).
- Hamann, A.; Smets, P.; Yanchuk, A.; Aitken, S. An ecogeographic framework for in situ conservation of forest trees in British Columbia. Can. J. For. Res. 2005, 35, 2553–2561. [Google Scholar] [CrossRef]
- Species at Risk and Local Government. A Primer for British Columbia. Available online: http://www.speciesatrisk.bc.ca/ (accessed on 8 May 2012).
- Redfeldt, G.E.; Crookston, N.L.; Sáenz-Romero, C.; Campbell, E.M. North American vegetation model for land-use planning in a changing climate: a solution to large scale classification problems. Ecol. Appl. 2012, 22, 119–141. [Google Scholar] [CrossRef]
- Marmion, M.; Parviainen, M.; Luoto, M.; Heikkinen, R.K.; Thuiller, W. Evaluation of consensus methods in predictive species distribution modelling. Divers. Distrib. 2009, 15, 59–69. [Google Scholar] [CrossRef]
- Pearson, R.G.; Dawson, T.P. Predicting the impacts of climate change on the distribution of species: Are bioclimate envelope models useful? Glob. Ecol. Biogeogr. 2003, 12, 361–371. [Google Scholar] [CrossRef]
- Austin, M. Spatial prediction of species distribution: An interface between ecological theory and statistical modelling. Ecol. Model. 2002, 157, 101–118. [Google Scholar] [CrossRef]
- Heikkinen, R.K.; Luoto, M.; Araujo, M.B.; Virkkala, R.; Thuiller, W.; Sykes, M.T. Methods and uncertainties in bioclimatic envelope modelling under climate change. Progress Phys. Geogr. 2006, 30, 751–777. [Google Scholar] [CrossRef]
© 2013 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/3.0/).
Share and Cite
Holmes, K.R.; Nelson, T.A.; Coops, N.C.; Wulder, M.A. Biodiversity Indicators Show Climate Change Will Alter Vegetation in Parks and Protected Areas. Diversity 2013, 5, 352-373. https://doi.org/10.3390/d5020352
Holmes KR, Nelson TA, Coops NC, Wulder MA. Biodiversity Indicators Show Climate Change Will Alter Vegetation in Parks and Protected Areas. Diversity. 2013; 5(2):352-373. https://doi.org/10.3390/d5020352
Chicago/Turabian StyleHolmes, Keith R., Trisalyn A. Nelson, Nicholas C. Coops, and Michael A. Wulder. 2013. "Biodiversity Indicators Show Climate Change Will Alter Vegetation in Parks and Protected Areas" Diversity 5, no. 2: 352-373. https://doi.org/10.3390/d5020352
APA StyleHolmes, K. R., Nelson, T. A., Coops, N. C., & Wulder, M. A. (2013). Biodiversity Indicators Show Climate Change Will Alter Vegetation in Parks and Protected Areas. Diversity, 5(2), 352-373. https://doi.org/10.3390/d5020352