Abiotic and Biotic Soil Characteristics in Old Growth Forests and Thinned or Unthinned Mature Stands in Three Regions of Oregon

Abstract
:Abbreviation
| df | degrees of freedom |
| CV | coefficient of variation |
| OG | old growth forest |
| FF | forest floor depth (mm) |
| TM | thinned mature forest |
| AMN | anaerobic mineralizable nitrogen (µg·gdw −1) |
| UM | unthinned mature forest |
| MAP | mean annual precipitation (mm) |
| BD | bulk density (g·mL−1) |
| MAT | mean annual temperature (°C) |
| DP | denitrification potential (nmol N·g dry mass−1·h−1) |
1. Introduction
- • Do unthinned mature stands differ from OG in any of the variables we measured?
- • Are thinned stands more or less similar to OG than unthinned stands?
- • Are differences among stand types, if they exist, consistent among regions?
2. Methods

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Region | MAP (mm) | MAT (°C) | MTWM (°C) | MTCM (°C) | Elevation (m) |
|---|---|---|---|---|---|
| Cascades | 1714 (1379–2261) | 9.6 (7.6–10.4) | 17.6 (15.8–18.3) | 3.0 (0.8–4.0) | 682–1111 |
| Coast Range | 2269 (1881–3367) | 10.7 (9.0–11.3) | 18.3 (16.0–19.3) | 4.4 (3.4–4.9) | 263–546 |
| Siskiyous | 869 (686–1159) | 10.1 (8.5–10.8) | 19.4 (17.6–20.2) | 2.4 (0.9–3.5) | 780–1032 |
3. Results
3.1. Univariate Analyses
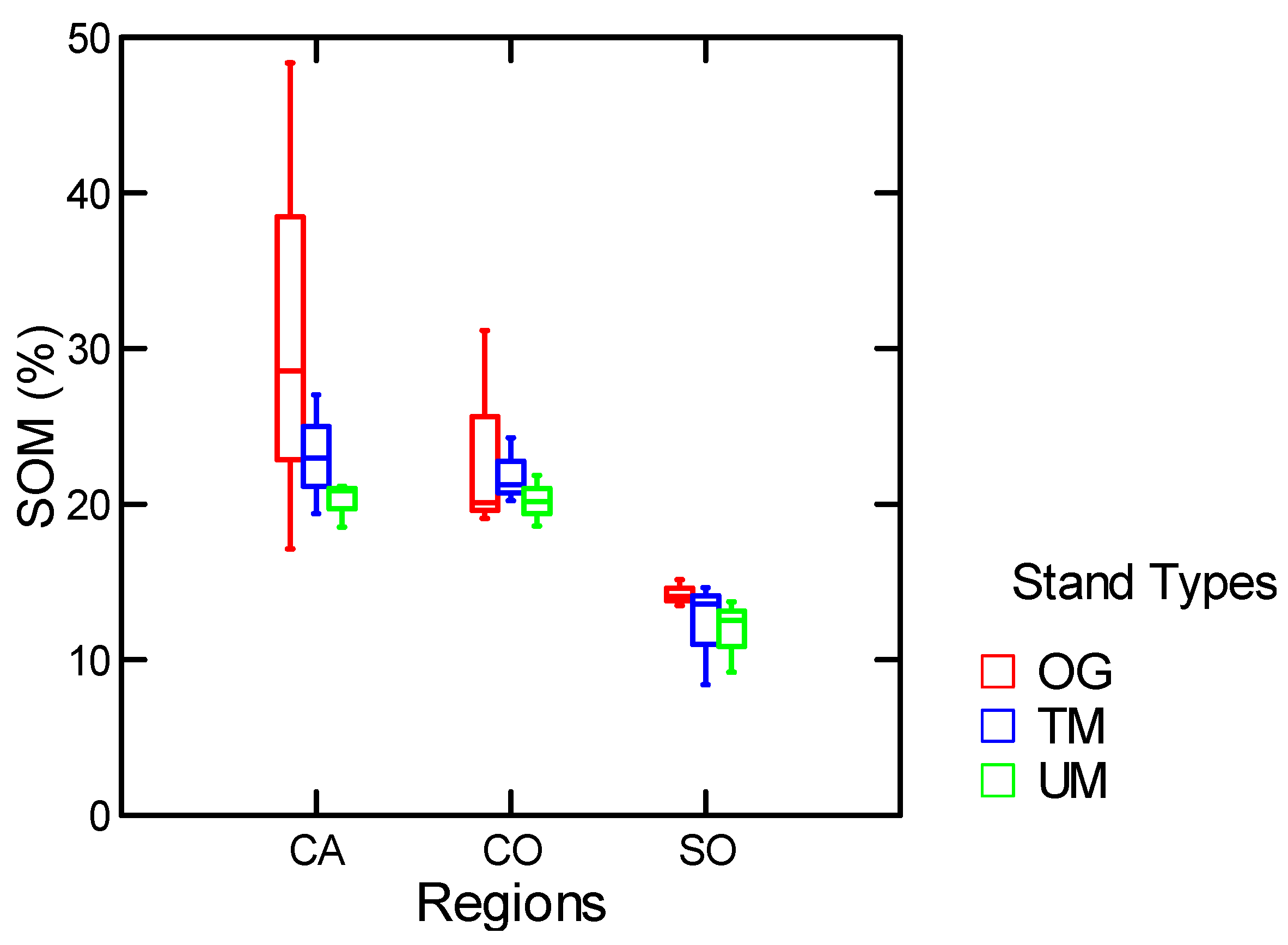
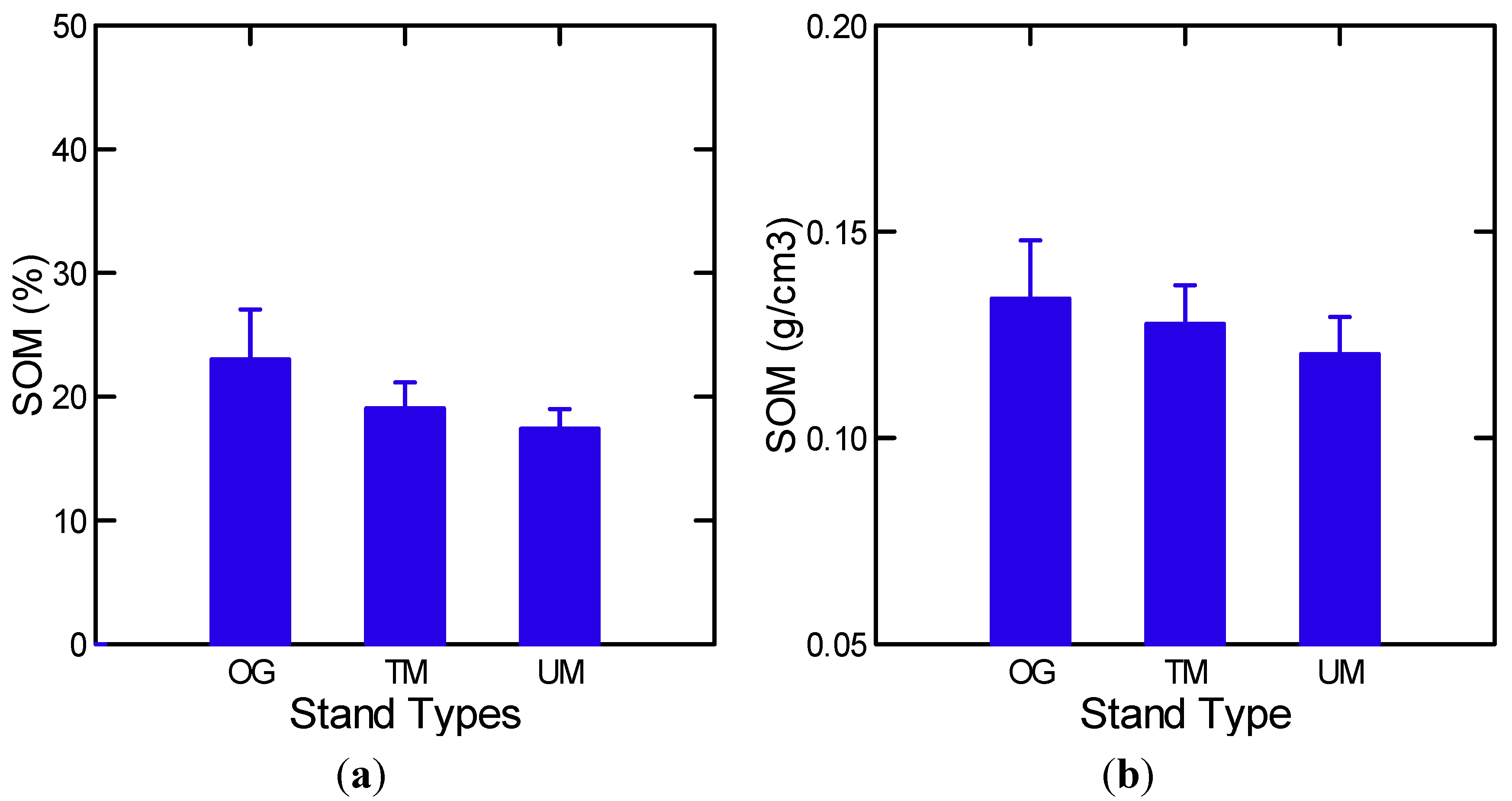
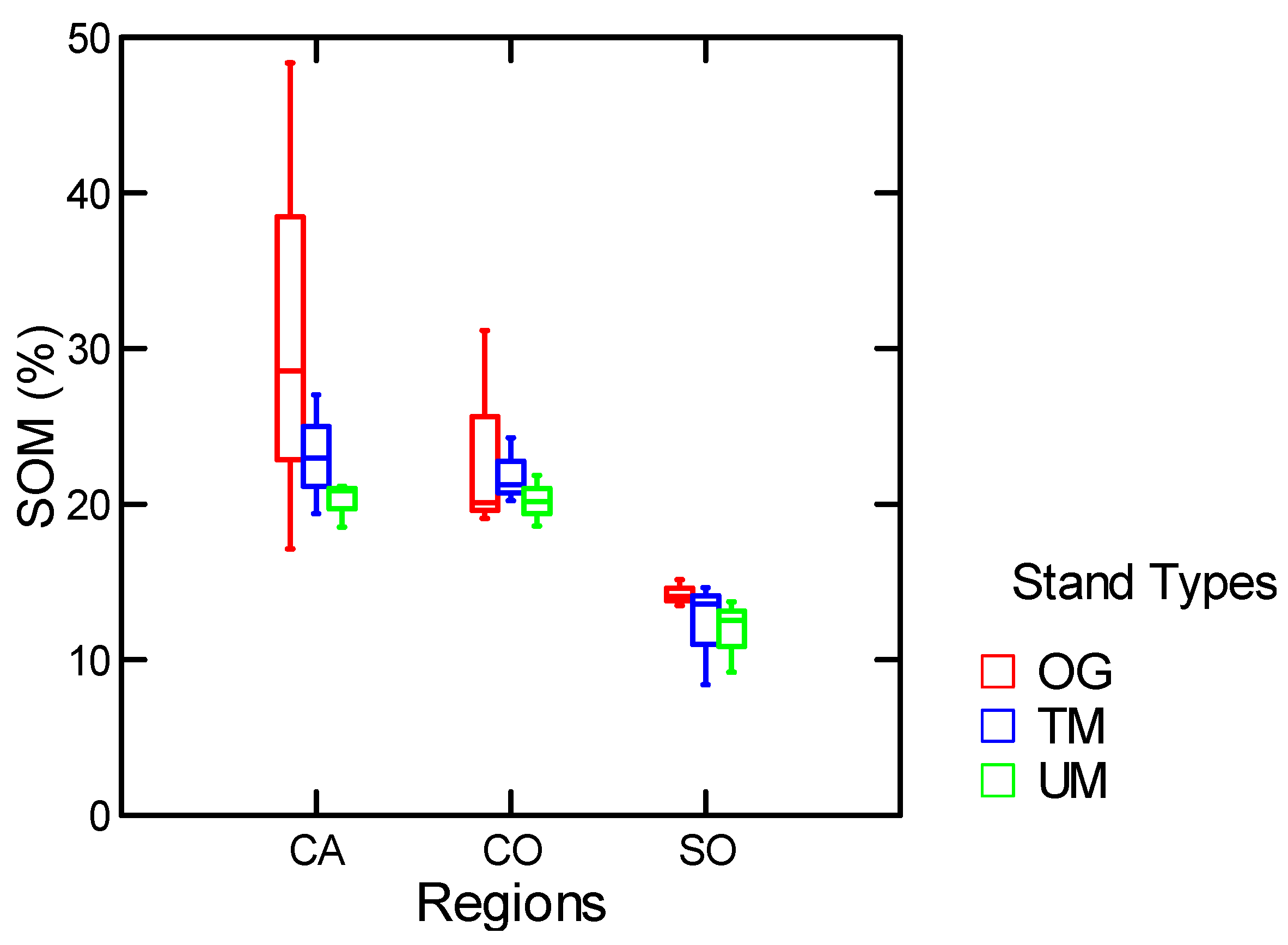
3.1.1. Mineral Soil Organic Matter and Forest Floor
| Response Variable | Factor | F | P |
|---|---|---|---|
| SOM % | Stand Types | 17.7 | 0.01 |
| SOM % | Stand Types w/o MR | 10.0 | 0.03 |
| SOM (g/cm3) | Stand Types | 1.2 | 0.38 |
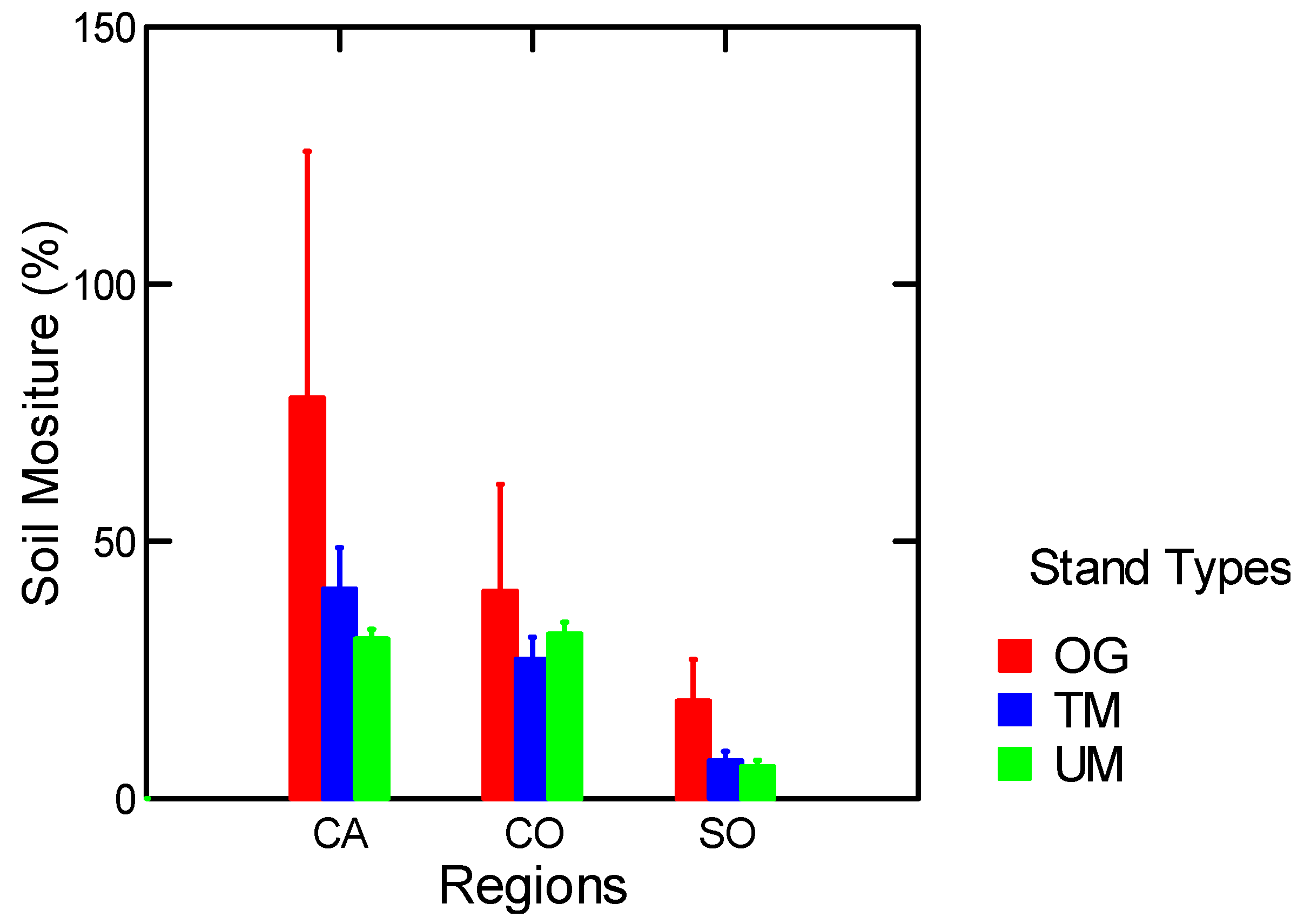
| Soil Moisture (%) | Stand Types | 15.7 | 0.01 |
| Soil Moisture (%) | Stand Types w/o MR | 7.2 | 0.05 |
| Soil Moisture (g/cm3) | Stand Types | 9.5 | 0.03 |
| Soil Moisture (g/cm3) | Stand Types w/o MR | 5.0 | 0.08 |
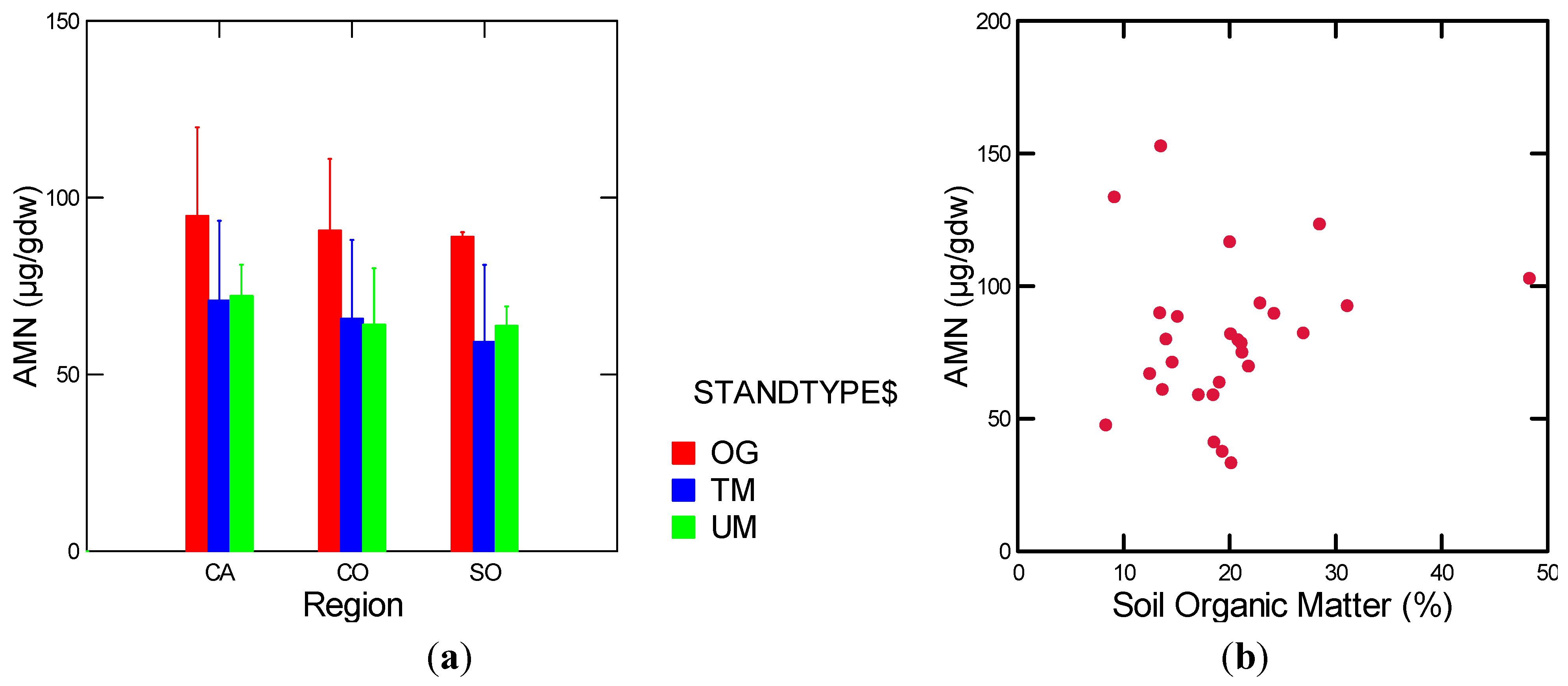
| AMN | Stand Types | 1.6 | 0.31 |
| AMN | Stand Types w/o TC | 16.5 | 0.01 |
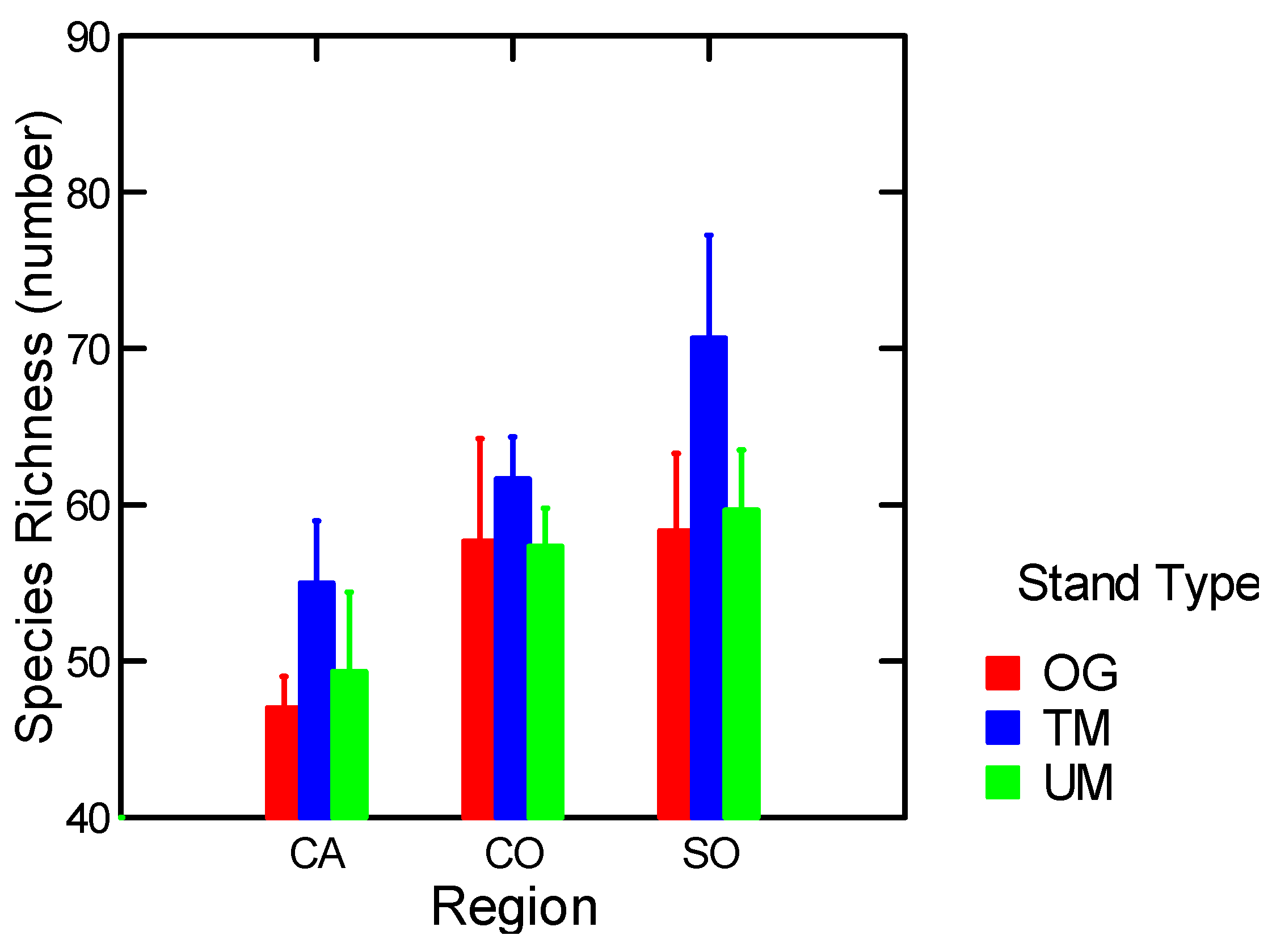
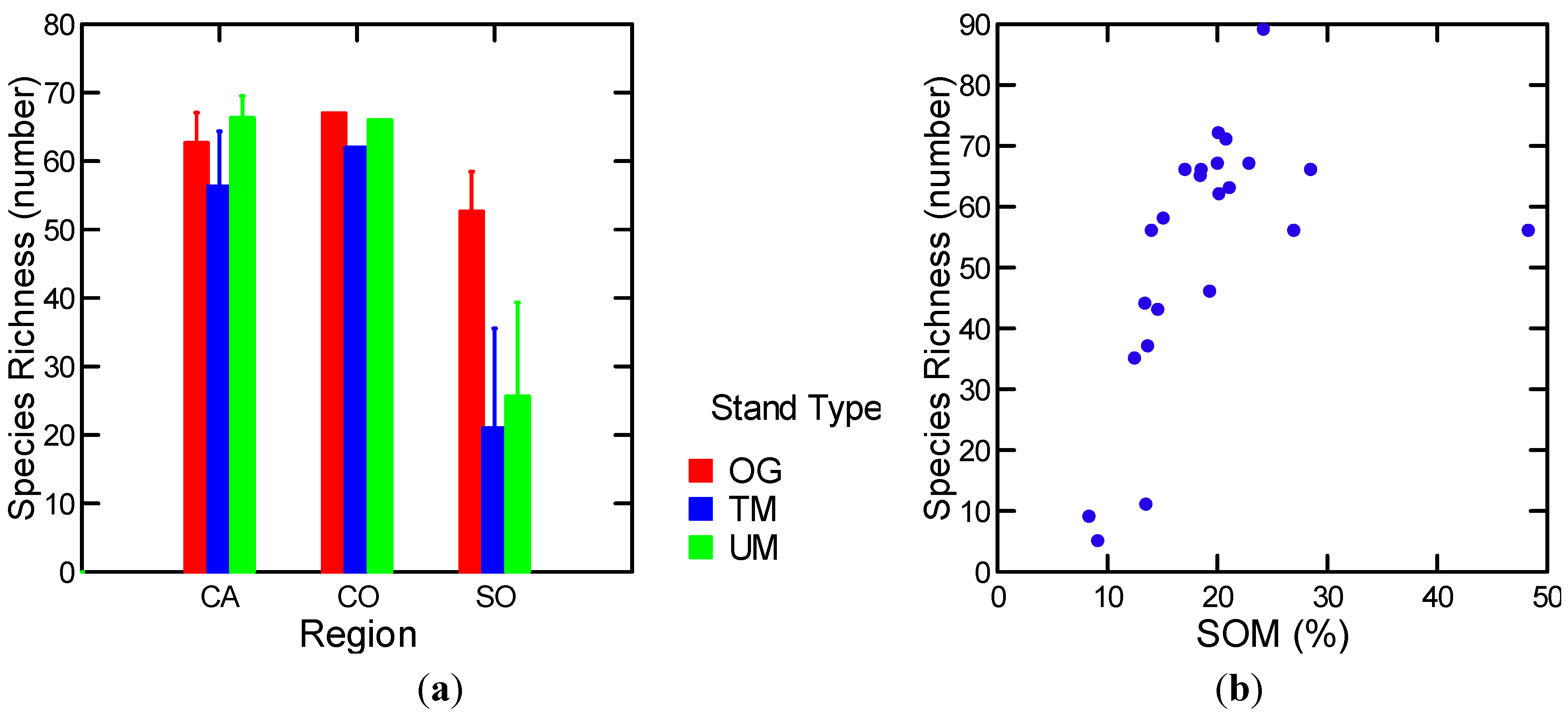
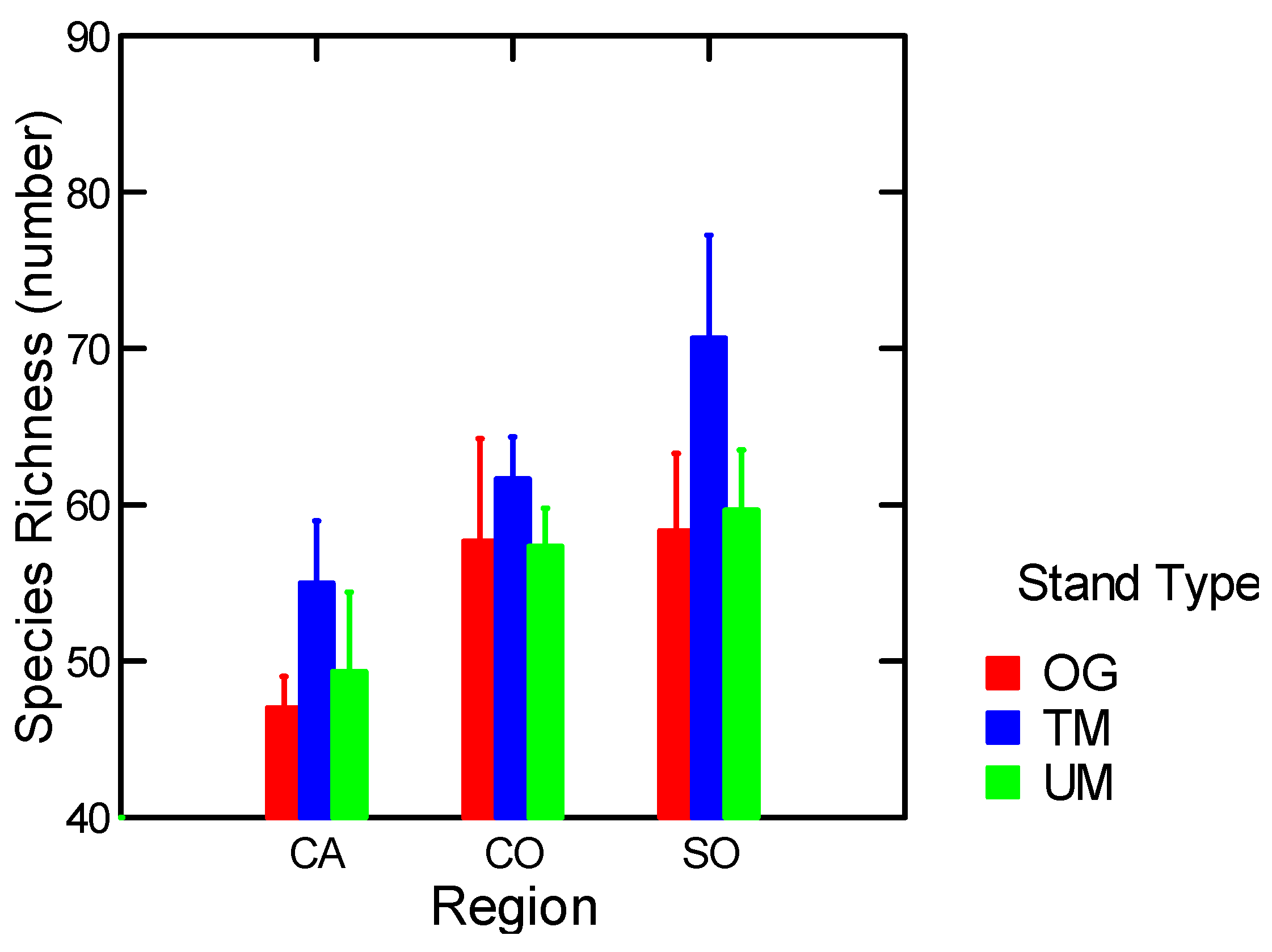
| Epigeic Macrofauna Richness | Stand Types | 9.8 | 0.03 |
| Dependent Variable | Data Base | Independent Variable(s) (sign of coefficient) | R2 | F | df | P |
|---|---|---|---|---|---|---|
| Log [SOM (%)] | All | Log [FF] (+) | 0.58 | 16.2 | 2, 24 | 0.000 |
| Log [elevation] (−) | ||||||
| SOM (%) | Coast Range | MAT (−) | 0.82 | 34.0 | 1, 7 | 0.001 |
| Log [SOM (%)] | All OG | Log [FF] (+) | 0.41 | 4.92 | 1, 7 | 0.062 |
| Log [SOM (%)] | All TM | Log [FF ](+) | 0.79 | 27.0 | 1, 7 | 0.001 |
| Log [SOM (%)] | All UM | Log [FF] (+) | 0.74 | 8.64 | 2, 6 | 0.017 |
| Log [elevation] (−) | ||||||
| BD | All | SOM % (−) | 0.59 | 36.3 | 1, 25 | 0.000 |
| Log [FF] | All UM | Log [MAP] (+) | 0.62 | 11.2 | 1, 7 | 0.012 |
| Log [Soil Moisture(%)] | All | Log [SOM(%)] (+) | 0.89 | 194.0 | 1, 25 | 0.000 |
| Soil Moisture (g/cm3) | All | SOM (%) | 0.80 | 102.0 | 1, 25 | 0.000 |
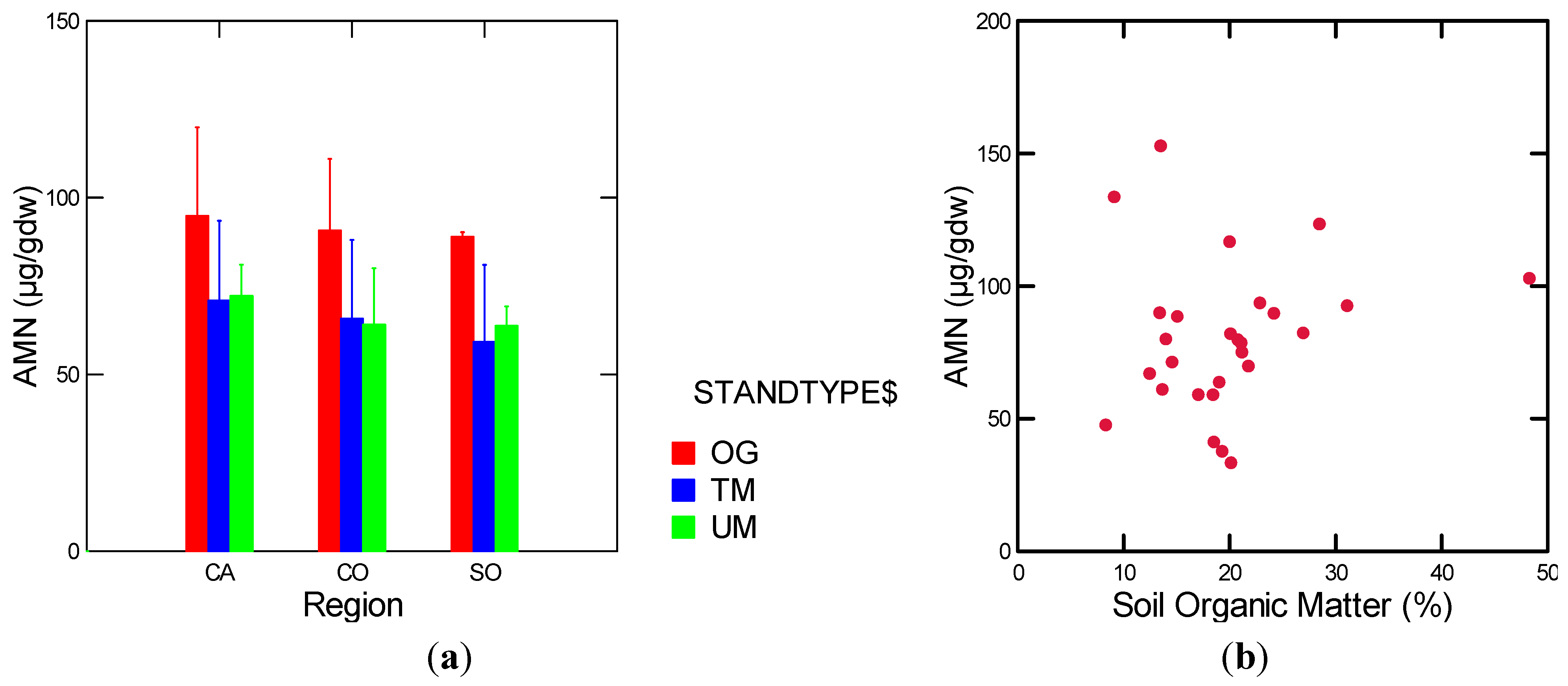
| AMN | w/o TCM | soil moisture (g/cm3)(+) MAP (−) | 0.41 | 7.69 | 2, 22 | 0.003 |
| Log [DP] | TM | Log [AMN] (+) | 0.61 | 4.7 | 2, 6 | 0.06 |
| Log [soil moisture (g/cm3)] (+) | ||||||
| Log [DP] | UM | Log [AMN] (+) | 0.49 | 6.7 | 1, 7 | 0.04 |
| Log [DP} | OG | Log [soil mesofauna (abundance)] (−) | 0.94 | 33.5 | 2, 4 | 0.003 |
| Log [soil moisture (g/cm3)] (+) |
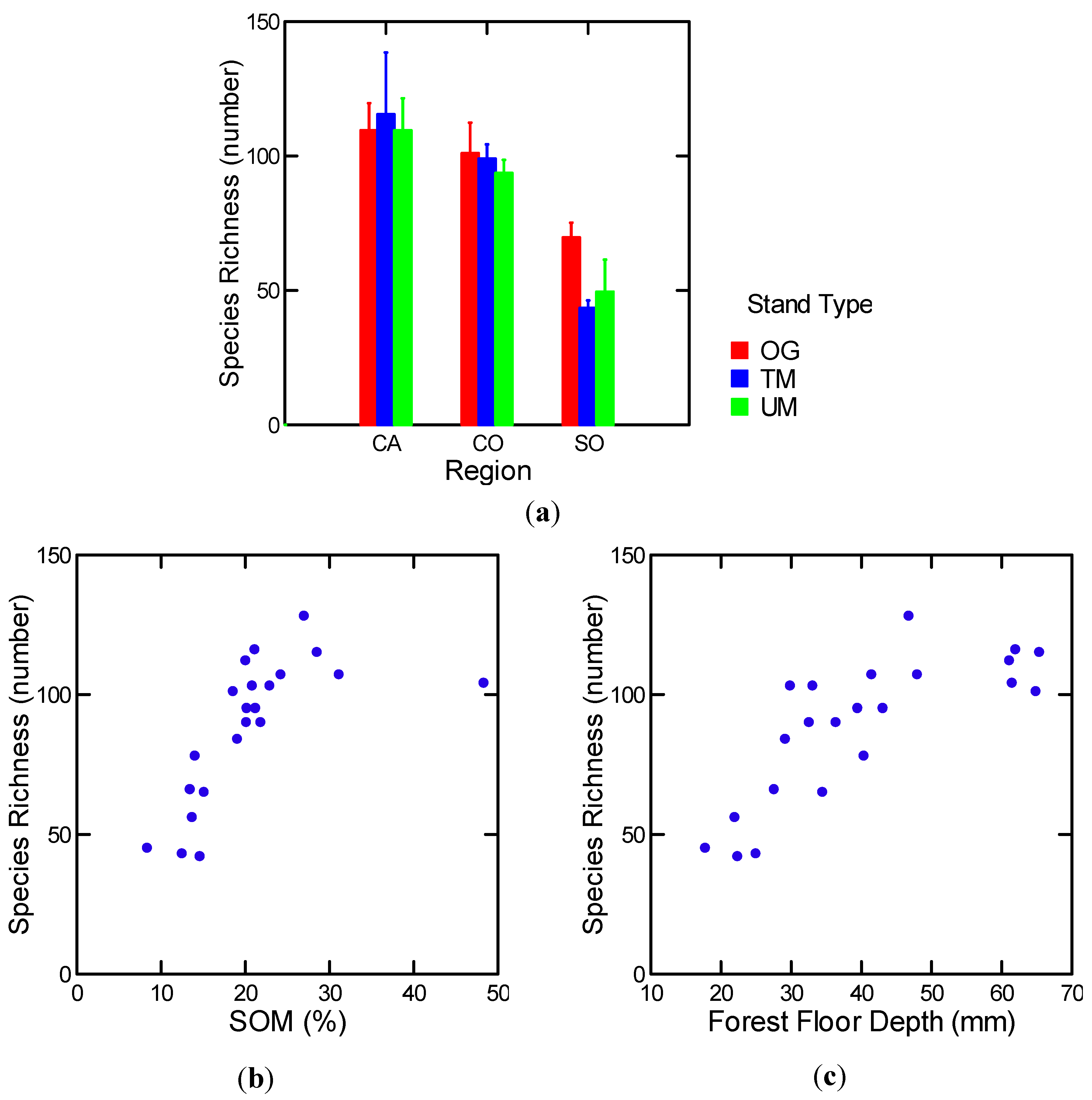
3.1.2. Among and within Stand Distribution of Soil Organic Matter
3.1.3. Soil Moisture
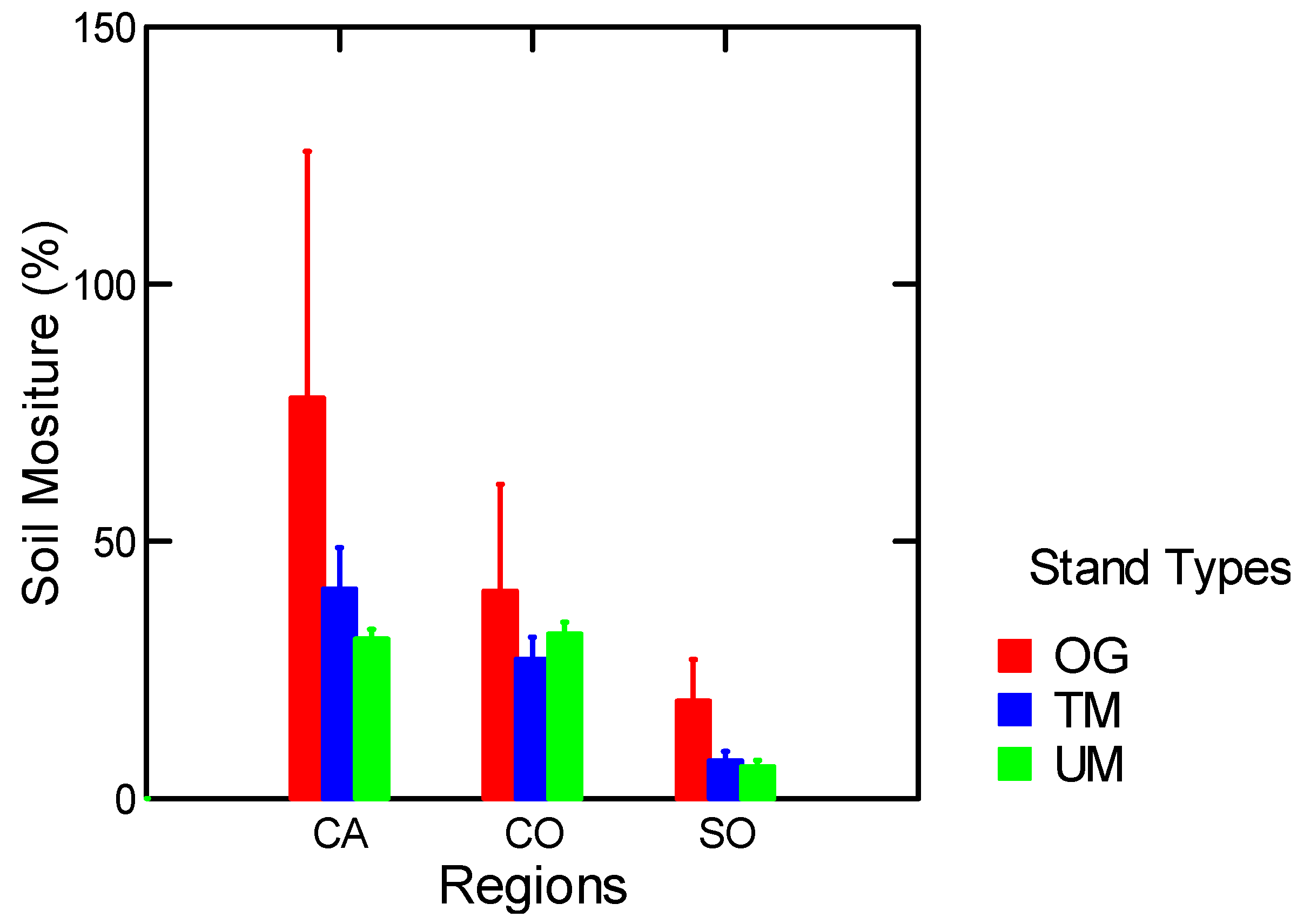
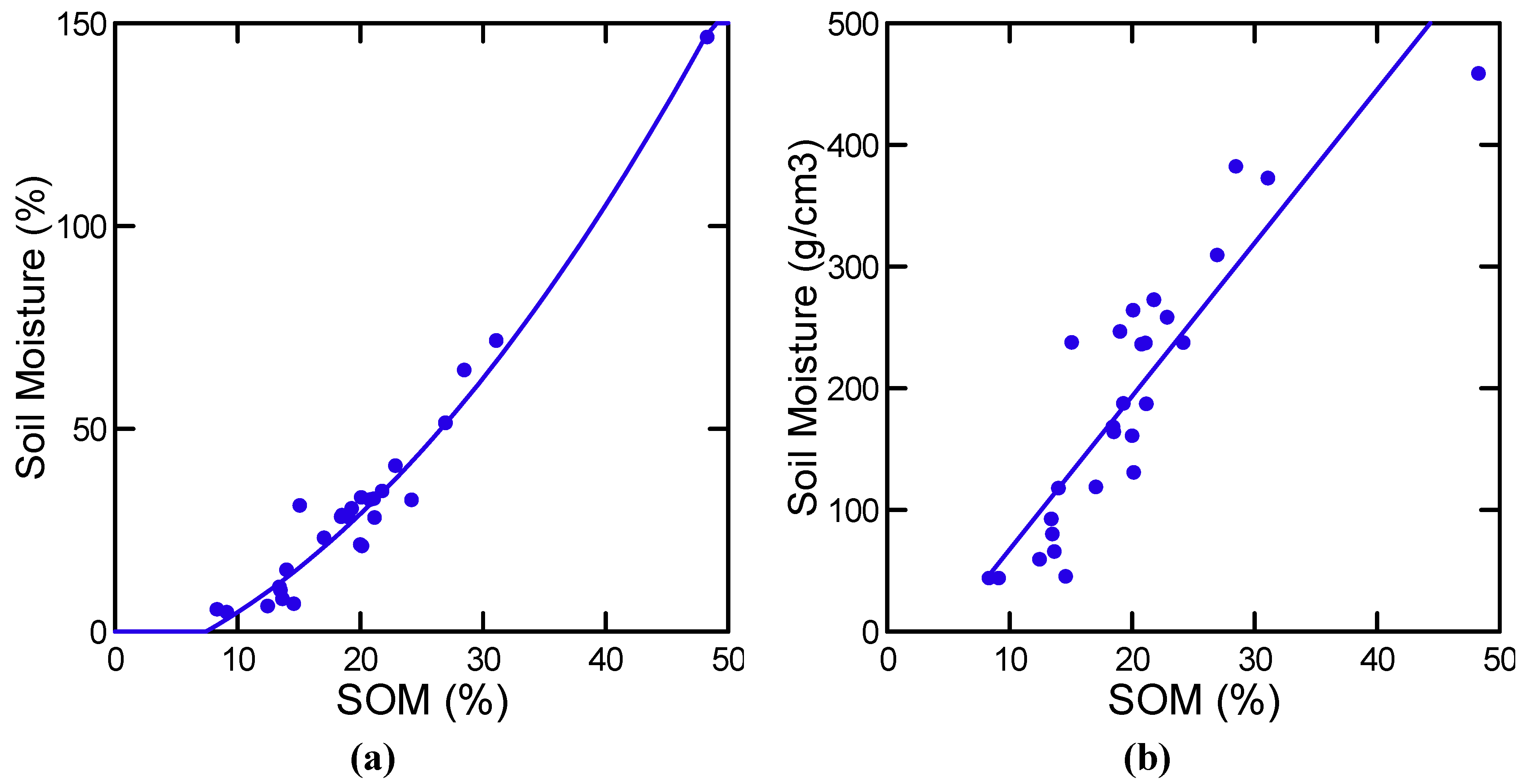
3.1.4. Nitrogen
3.1.5. Arthropods
3.1.5.1. Epigeic Macrofauna
| Dependent Variable | Data Base | Independent Variable (s) (sign of coefficient) | R2 | F | df | P |
|---|---|---|---|---|---|---|
| Epigeic macrofauna: Richness | All | FF (−) | 0.34 | 13.04 | 1, 25 | 0.001 |
| Log [Epigeic macrofauna: Abundance] | All | Log [FF] (−) | 0.20 | 6.27 | 1, 25 | 0.019 |
| Log [FF mesofauna richness] | 25 stands | Log [FF] (+) | 0.75 | 28.6 | 2, 19 | 0.000 |
| Log [SOM%] (+) | ||||||
| FF mesofauna abundance | 25stands | Soil moisture (%) (+) | 0.73 | 25.6 | 2, 19 | 0.000 |
| MAP (+) | ||||||
| Log [soil mesofauna richness] | 25 stands | Log [soil moisture (g/cm3)] (+) | 0.50 | 19.1 | 1, 21 | 0.000 |
| Log [soil mesofauna richness] | w/o MR OG | Log [SOM (%)] (+) | 0.65 | 36.1 | 1, 20 | 0.000. |
3.1.5.2. Forest Floor Mesofauna
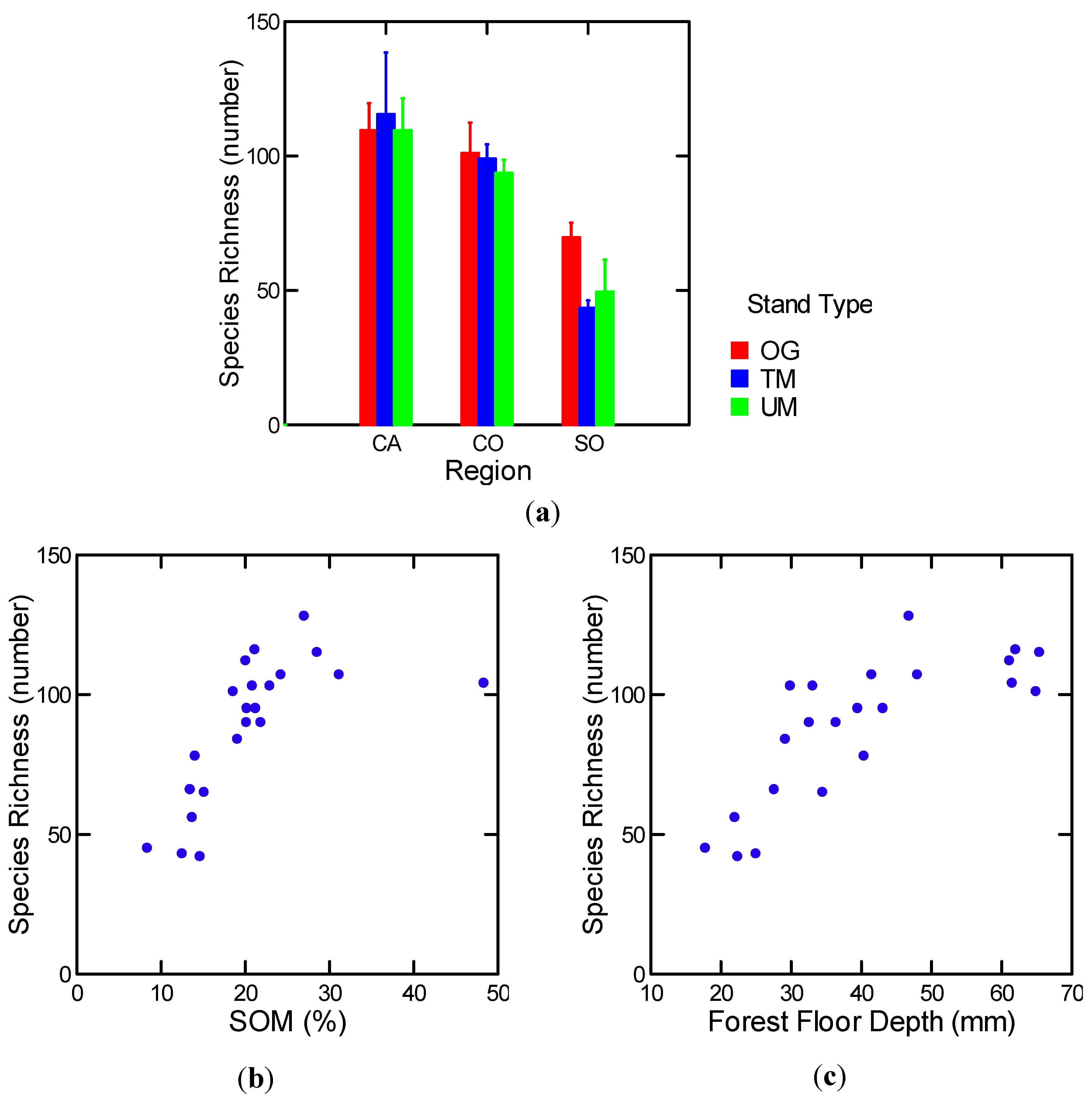
3.1.5.3. Soil Mesofauna
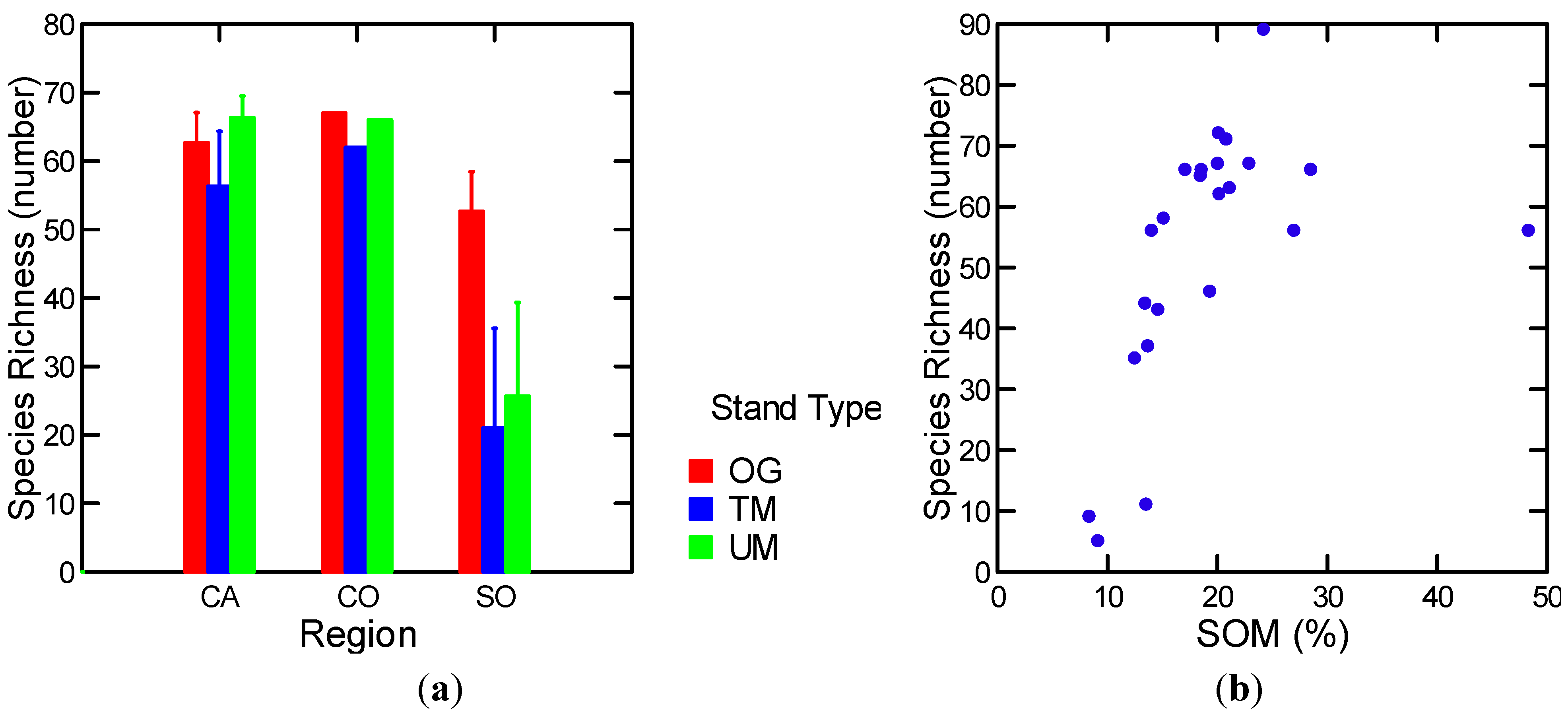
3.2. Multivariate Analysis of Variance
3.2.1. With Soil Mesofauna
| Models | H-L Trace | F | df | OG vs. TM | OG vs. UM | UM vs. TM | |
|---|---|---|---|---|---|---|---|
| With Soil Mesofauna (7 sites) | Model 1 | 0.019 | 3.79 | 6, 14 | 0.004 | 0.004 | 0.460 |
| Without Soil Mesofauna, with FF mesofauna (7 sites) | Model 2 | 0.010 | 4.42 | 6, 14 | 0.0012 | 0.010 | 0.626 |
| Model 3 | 0.047 | 2.90 | 8, 12 | 0.006 | 0.029 | 0.710 | |
| Without soil or FF mesofauna (9 sites) | Model 4 | 0.012 | 4.1 | 4, 28 | 0.009 | 0.048 | 0.076 |
3.2.2. Without Soil Mesofauna, with Forest Floor Mesofauna
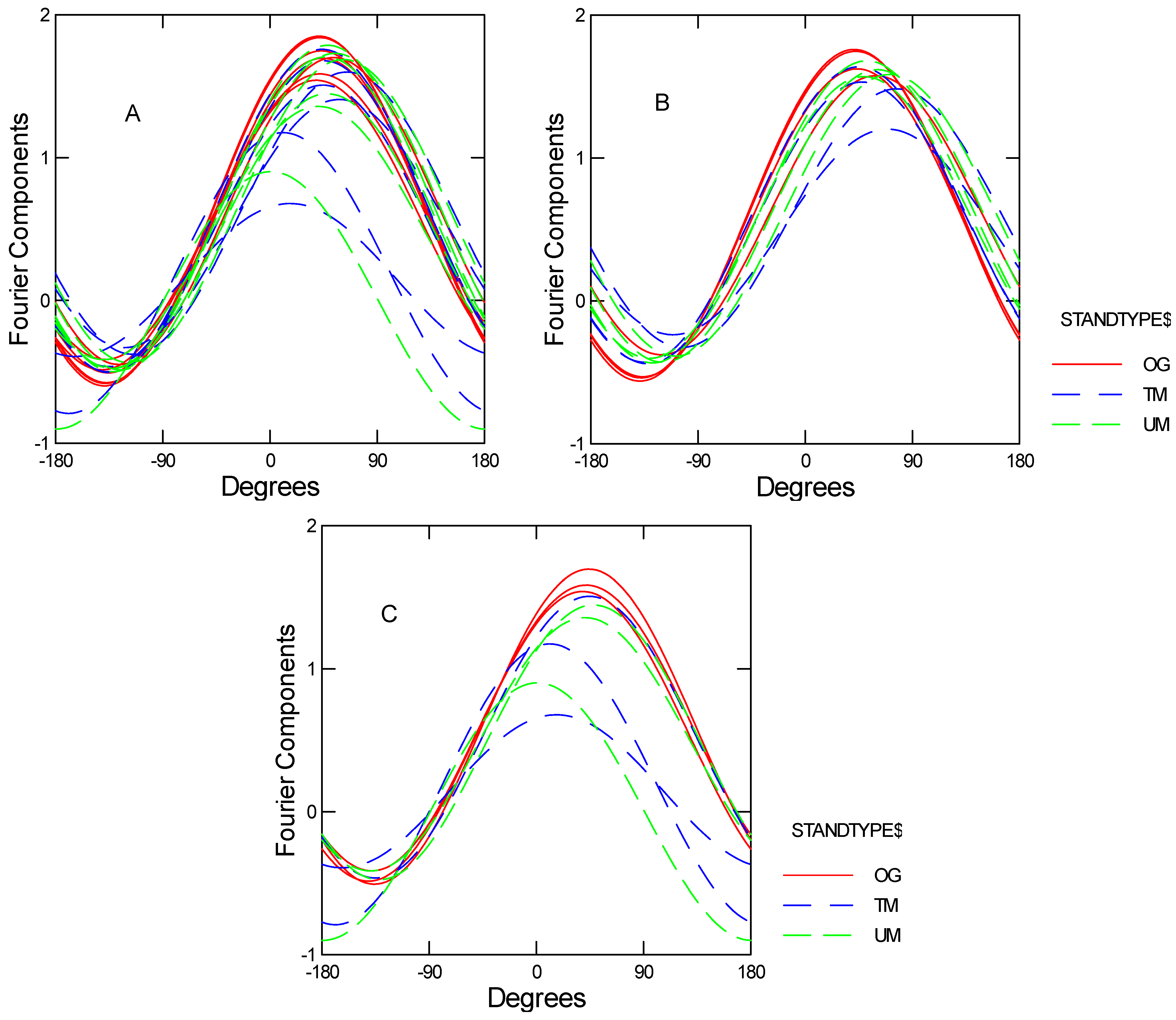

3.2.3. Without Soil or Forest Floor Mesofauna: All Nine Sites
4. Discussion
4.1. Differences in Stand Types
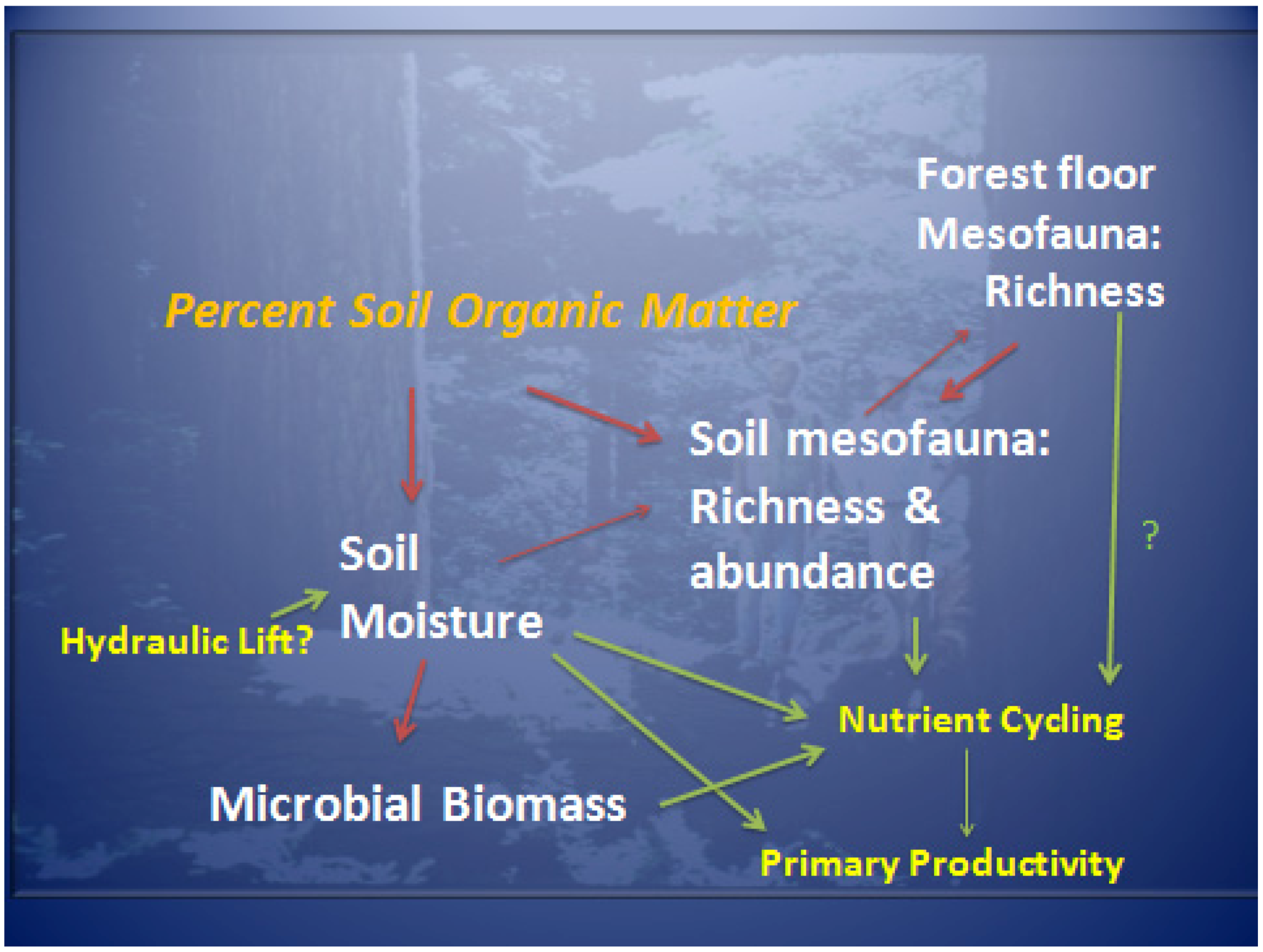
4.2. Soil Water and Extended Sampling
4.3. Outliers, Stand History, and Biological Legacies
4.4. Landscape Legacies
4.5. Resilience: Are Mature Stands on a Trajectory Toward Old Growth Conditions?
Acknowledgements
References
- Strittholt, J.R.; Dellasalla, D.A.; Jiang, H. Status of mature and old-growth forests in the Pacific Northwest. Cons. Biol. 2006, 20, 363–374. [Google Scholar] [CrossRef]
- FEMAT (Forest Ecosystem Assessment Team), Forest Ecosystem Management: An Ecological, Economic, and Social Assessment; Report of the FEMAT; U.S. Government Printing Office: Washington, DC, USA, 1993.
- Sabine, C.L.; Heimann, M.; Artaxo, P.; Bakker, D.C.E.; Chen, C.T.A.; Field, C.B.; Gruber, N.; Le Quere, C.; Prinn, R.G.; Richey, J.E.; Lankao, P.R.; Sathaye, J.A.; Valentini, R. Current status and past trends on the global carbon cycle. In The Global Carbon Cycle SCOPE 62; Field, C.B., Raupach, M.R., Eds.; Island Press: Washington, DC, USA, 2004; pp. 17–44. [Google Scholar]
- Jandl, R.; Linder, M.; Vesterdal, L.; Bauwens, B.; Baritz, R.; Hagedorn, F.; Johnson, D.W.; Minkkinen, K.; Byrne, K.A. How strongly can management influence soil carbon sequestration? Geoderma 2007, 137, 253–268. [Google Scholar]
- Bradford, J.D.; Kastendick, D.N. Age-related patterns of forest complexity and carbon storage in pine and aspen-birch ecosystems in northern Minnesota, USA. Can. J. For. Res. 2010, 40, 401–409. [Google Scholar] [CrossRef]
- Johnson, D.W.; Murphy, J.D.; Walker, R.F.; Glass, D.; Miller, W.W. Wildfire effects on forest carbon and nutrient budgets. Ecol. Eng. 2007, 31, 183–192. [Google Scholar] [CrossRef]
- Bormann, B.T.; Homann, P.S.; Darbyshire, R.L.; Morrissette, B.A. Intense wildfire sharply reduces mineral soil C and N: The first evidence. Can. J. For. Res. 2008, 38, 2771–2783. [Google Scholar] [CrossRef]
- Johnson, D.W.; Curtis, P.S. Effects of forest management on soil C and N storage: Meta analysis. For. Ecol. Manag. 2001, 140, 227–238. [Google Scholar] [CrossRef]
- Sanchez, F.G.; Tiarks, A.E.; Kranabetter, J.M.; Page-Dumroese, D.S.; Powers, R.F.; Sanborn, P.T.; Chapman, W.K. Effects of organic matter removal and soil compaction on fifth-year mineral soil carbon and nitrogen contents for sites across the United States and Canada. Can. J. For. Res. 2006, 36, 565–576. [Google Scholar] [CrossRef]
- Nave, L.E.; Vance, E.D.; Swanston, C.W.; Curtis, P.S. Harvest impacts on soil carbon storage in temperate forests. For. Ecol. Manag. 2010, 259, 857–866. [Google Scholar] [CrossRef]
- Nave, L.E.; Vance, E.D.; Swanston, C.W.; Curtis, P.S. Fire effects on temperate forest soil C and N storage. Ecol. Appl. 2011, 21, 1189–1201. [Google Scholar] [CrossRef]
- Natural Resources Conservation Service. Distribution Maps of Dominant Soil Orders. Available online: http://soils.usda.gov/technical/classification/orders/ (accessed on 6 September 2012).
- Pregitzer, K.S.; Euskirchen, E.S. Carbon cycling and storage in world forests: Biome patterns related to forest age. Glob. Chang. Biol. 2004, 10, 2052–2077. [Google Scholar] [CrossRef]
- Griffiths, R.P.; Swanson, A.K. Forest soil characteristics in a chronosequence of harvested Douglas-fir forests. Can. J. For. Res. 2001, 31, 1871–1879. [Google Scholar] [CrossRef]
- Buddle, C.M.; Langor, D.W.; Pohl, G.R.; Spence, J.R. Arthropod responses to harvesting and wildfire: Implications for emulation of natural disturbance in forest management. Biol. Conserv. 2006, 128, 36–357. [Google Scholar]
- Pohl, G.R.; Langor, D.W.; Spence, J.R. Rove beetles and ground beetles (Coleoptera: Staphylinidae, Carabidae) as indicators of harvest and regeneration practices in western Canadian foothills forests. Biol. Conserv. 2007, 137, 294–307. [Google Scholar] [CrossRef]
- Baker, S.C.; Grove, S.J.; Forster, L.; Bonham, K.J.; Bashford, D. Short-term tresponses of ground-active beetles to alternative silvicultural systems in the Warra Silvicultural Systems Trial, Tasmania, Australia. For. Ecol. Manag. 2009, 258, 444–459. [Google Scholar] [CrossRef]
- Légaré, J.-P.; Hébert, C.; Ruel, J.-C. Alternative silvicultural practices in irregular Boreal forests: Response of beetle assemblages. Silva Fenn. 2011, 45, 937–956. [Google Scholar]
- Hayes, J.P.; Chan, S.S.; Emmingham, W.H.; Tappeiner, J.C.; Kellog, L.D.; Bailey, J.D. Wildlife response to thinning young forests in the Pacific Northwest. J. For. 1997, 95, 28–33. [Google Scholar]
- Lindh, B.C.; Muir, P.S. Understory vegetation in young Douglas-fir forests: Does thinning help restore old-growth composition? For. Ecol. Manag. 2004, 192, 285–296. [Google Scholar] [CrossRef]
- Bailey, J.D.; Mayrsohn, C.; Doescher, P.S.; St. Pierre, E.; Tappeiner, J.C. Understory vegetation in old and young Douglas-fir forests of western Oregon. For. Ecol. Manag. 1998, 112, 289–302. [Google Scholar] [CrossRef]
- Bailey, J.D.; Tappeiner, J.C. Effects of thinning on structural development in 40- to 100-year-old Douglas-fir stands in western Oregon. For. Ecol. Manag. 1998, 108, 99–113. [Google Scholar] [CrossRef]
- Hagar, J.C.; McComb, W.C.; Emmingham, W.H. Bird communities in commercially thinned and unthinned Douglas-fir stands of western Oregon. Wildl. Soc. Bull. 1996, 24, 353–366. [Google Scholar]
- Hayes, J.P.; Weikel, J.M.; Huso, M.M.P. Response of birds to thinning in young Douglas-fir forests. Ecol. Appl. 2003, 13, 1222–1232. [Google Scholar] [CrossRef]
- Manning, T.; Hagar, J.C.; McComb, B.C. Thinning of young Douglas-fir forests decreases density of northern flying squirrels in the Oregon Cascades. For. Ecol. Manag. 2012, 264, 115–124. [Google Scholar] [CrossRef]
- Holloway, G.L.; Smith, W.P. A meta-analysis of forest age and structure effects on northern flying squirrel densities. J. Wildl. Manag. 2011, 75, 668–674. [Google Scholar] [CrossRef]
- Carey, A.B.; Johnson, M.L. Small mammals in managed, naturally young, and old-growth forests. Ecol. Appl. 1995, 5, 336–352. [Google Scholar] [CrossRef]
- Lorenz, K.; Lal, R. Carbon Sequestration in Forest Ecosystems; Springer: New York, NY, USA, 2010. [Google Scholar]
- Main, M. Personal Communication. Consulting Forester: Ashland, OR, USA, March 2012. [Google Scholar]
- Wang, T.; Hamann, A.; Spittlehouse, D. ClimateWNA: A Program to Generate High-Resolution Climate Data for Climate Change Studies and Applications in Western North America; ClimateWNA v. 4.60. University of British Columbia: Vancouver, Canada, 2010. Available online: http://www.genetics.forestry.ubc.ca/cfcg/ClimateWNA/ClimateWNA.html (accessed on 14 September, 2012).
- Wang, T.; Hamann, A.; Spittlehouse, D.; Aitken, S.N. Development of scale-free climate data for western Canada for use in resource management. Int. J. Climatol. 2006, 26, 383–397. [Google Scholar] [CrossRef]
- Natural Resources Conservation Service. Soil Surveys for Oregon. Available online: http://www.or.nrcs.usda.gov/pnw_soil/or_data.html/ (accessed on 6 September 2012).
- Natural Resources Conservation Service. Oregon Soil Surveys Data. Available online: http://www.or.nrcs.usda.gov/pnw_soil/or_data.html/ (accessed on 6 September 2012).
- Keeny, D.; Bremner, J. A chemical index of soil nitrogen availability. Nature 1966, 211, 892–893. [Google Scholar] [CrossRef]
- Binkley, D.; Hart, S.C. The components of nitrogen availability assessments in forest soils. Adv. Soil Sci. 1989, 10, 57–112. [Google Scholar] [CrossRef]
- Groffman, P.M.; Tiedje, J.M. Denitrification in north temperate forest soils: Relationships between denitrification and environmental factors at the landscape scale. Soil Biol. Biochem. 1989, 21, 621–626. [Google Scholar] [CrossRef]
- Griffiths, R.P.; Homann, P.S.; Riley, R. Denitrification enzyme activity of Douglas-fir and red alder forest soils of the Pacific Northwest. Soil Biol. Biochem. 1998, 30, 1147–1157. [Google Scholar] [CrossRef]
- Spence, J.R.; Niemala, J.K. Sampling carabid assemblages with pitfall traps: The madness and the method. Can. Entomol. 1994, 126, 881–894. [Google Scholar] [CrossRef]
- Winter, J.P.; Voroney, R.P. Microarthropods in soil and litter. In Soil Sampling and Methods of Analysis; Carter, M.R., Ed.; Canadian Society of Soil Science, CRC Press: Boca Raton, FL, USA, 1993; pp. 333–340. [Google Scholar]
- Systat Software, Version 13; Cranes Software International Ltd.: Bangalore, India, 2008.
- Lowry, R. Concepts and Applications of Inferential Statistics. 2003. Available online: http://vassarstats.net/textbook/ (accessed on 14 September 2012).
- Latta, G.; Temesgen, H.; Adams, D.; Barret, T. Analysis o potential impacts of climate change on forests of the United States Pacific Northwest. For. Ecol. Manag. 2010, 259, 720–729. [Google Scholar] [CrossRef]
- Andrews, D.F. Plots of high dimensional data. Biometrics 1972, 28, 125–136. [Google Scholar] [CrossRef]
- Waring, R.H.; Franklin, J.F. The evergreen forests of the Pacific Northwest. Science 1979, 204, 1380–1386. [Google Scholar]
- Edmonds, R.L.; Binkley, D.; Feller, M.C.; Sollins, P.; Abee, A.; Myrold, D.D. Nutrient cycling: effects on productivity of Pacific Northwest forests. In Maintaining The Long-Term Productivity of Pacific Northwest Forest Ecosystems; Perry, D.A., Meurisse, R., Thomas, B., Miller, R., Boyle, J., Means, J., Perry, C.R., Powers, R.F., Eds.; Timber Press: Portland, OR, USA, 1989; pp. 17–35. [Google Scholar]
- Shumway, J.; Atkinson, W.A. Predicting nitrogen fertilizer response in unthinned stands of Douglas-fir. Comm. Soil Sci. Plan. 1978, 9, 529–539. [Google Scholar] [CrossRef]
- Powers, R.F. Mineralizable nitrogen as an index of nitrogen availability to forest trees. Soil Sci. Soc. Am. J. 1980, 44, 1314–1320. [Google Scholar] [CrossRef]
- Nielsen, U.N.; Ayers, E.; Wall, D.H.; Bardgett, R.D. Soil biodiversity and carbon cycling: A review and synthesis of studies examining diversity-function relationships. Eur. J. Soil Sci. 2011, 62, 105–116. [Google Scholar]
- Luyssaert, S.; Schulze, E.-D.; Börner, A.; Knohl, A.; Hessenmöller, D.; Law, B.E.; Ciais, P.; Grace, J. Old-growth forests as global carbon sinks. Nature 2008, 455, 213–215. [Google Scholar]
- Fogel, R.; Cromack, K., Jr. Effect of habitat and substrate quality on Douglas-fir litter decomposition in western Oregon. Can. J. Bot. 1977, 55, 1632–1640. [Google Scholar] [CrossRef]
- Silver, W.L.; Miya, R.K. Global patterns in root decomposition: Comparisons of climate and litter quality effects. Oecologia 2001, 129, 407–419. [Google Scholar]
- Palmroth, S.; Oren, R.; McCarthy, H.R.; Johnsen, K.H.; Finzi, A.C.; Butnor, J.R.; Ryan, M.G.; Schlesinger, W.H. Aboveground sink strength in forests controls the allocation of carbon belowground and its CO2-induced enhancement. Proc. Natl. Acad. Sci. USA 2006, 103, 19362–19367. [Google Scholar]
- Bauhus, J.; Paré, D.; Côté, L. Effects of tree species, stand age, and soil type on soil microbial biomass and its activity in southern boreal forest. Soil Biol. Biochem. 1998, 30, 1077–1089. [Google Scholar] [CrossRef]
- Griffiths, R.P.; Bradshaw, G.A.; Marks, B.; Lienkaemper, G.W. Spatial distribution of ectomycorrhizal mats in coniferous forests of the Pacific Northwest, USA. Plant Soil 1996, 180, 147–158. [Google Scholar] [CrossRef]
- Vogt, K.; Vogt, D.L.; Brown, S.; Tilley, J.P.; Edmonds, R.; Silver, W.P.; Siccama, T.G. Dynamics of forest floor and soil organic matter accumulation in boreal, temperate, and tropical forests. In Soil Management and Greenhouse Effect; Lal, R., Kimble, J., Levine, E., Stewart, B.A., Eds.; CRC Press: New York, NY, USA, 1995; pp. 159–178. [Google Scholar]
- Aber, J.D.; Melillo, J.M.; McClaugherty, C.A. Predicting long-term patterns of mass-loss, nitrogen dynamics, and soil organic matter formation from initial fine litter chemistry in temperate forest ecosystems. Can. J. Bot. 1990, 68, 2201–2208. [Google Scholar] [CrossRef]
- Hattenschiwiler, S.; Tiunov, A.V.; Scheu, S. Biodiversity and litter decomposition in terrestrial ecosystems. Annu. Rev. Ecol. Evol. Syst. 2005, 36, 191–218. [Google Scholar] [CrossRef]
- Climate Impacts Group. About Pacific Northwest Climate; Climate Impacts Group: Seattle, WA, USA, 2011. Available online: http://cses.washington.edu/cig/pnwc/pnwc.shtml (accessed on 6 September 2012).
- Warren, J.M.; Meinzer, F.C.; Brooks, J.R.; Domec, J.C. Vertical stratification of soil water storage and release dynamics in Pacific Northwest coniferous forests. Agr. For. Meteorol. 2005, 130, 39–58. [Google Scholar] [CrossRef]
- Brooks, J.R.; Meinzer, F.C.; Coulombe, R.; Gregg, J. Hydraulic redistribution of soil water during summer drought in two contrasting Pacific Northwest coniferous forests. Tree Physiol. 2002, 22, 1107–1117. [Google Scholar] [CrossRef]
- Prieto, I.; Armas, C.; Pugnaire, F.I. Water release through plant roots: New insights into its consequences at the plant and ecosystem level. New Phytol. 2012, 193, 830–841. [Google Scholar] [CrossRef]
- Neumann, R.B.; Cardon, Z.G. The magnitude of hydraulic redistribution by plant roots: A review and synthesis of empirical and modeling studies. New Phytol. 2012, 194, 337–352. [Google Scholar] [CrossRef]
- Sollins, P.; Cline, S.P.; Verhoeven, T.; Sachs, D.; Spycher, G. Patterns of log decay in old-growth Douglas-fir forests. Can. J. For. Res. 1987, 17, 1585–1595. [Google Scholar] [CrossRef]
- Amaranthus, M.P.; Parrish, D.; Perry, D.A. Decaying logs as moisture reservoirs following drought and wildfire. In Proceedings of Watershed 89: Stewardship of Soil, Water, and Air Resources, Juneau, AK, USA, 21–23 March 1989; Alexander, E.B., Ed.; U.S. Department of Agriculture Forest Service, Alaska Region: Juneau, AK, USA, 1989. RlO-MB-77. [Google Scholar]
- Harmon, M.E.; Franklin, J.F.; Swanson, F.J.; Sollins, P.; Gregory, S.V.; Lattin, J.D.; Anderson, N.H.; Cline, S.P.; Aumen, N.G.; Sedell, J.R.; Lienkaemper, G.W.; Cromack, K., Jr.; Cummins, K.W. Ecology of coarse woody debris in temperate ecosystems. Adv. Ecol. Res. 1986, 15, 133–302. [Google Scholar] [CrossRef]
- Olson, J.S. Energy storage and the balance of producers and decomposers in ecological systems. Ecology 1966, 44, 322–331. [Google Scholar] [CrossRef]
- Perry, D.A.; Hessburg, P.F.; Skinner, C.N.; Spies, T.A.; Stephens, S.L.; Taylor, A.H.; Franklin, J.F.; McComb, B.; Riegel, G. The ecology of mixed severity fire regimes in Washington, Oregon, and Northern California. For. Ecol. Manag. 2011, 262, 703–717. [Google Scholar] [CrossRef]
- Agee, J.K. Fire Ecology of Pacific Northwest Forests; Island Press: Washington, DC, USA, 1993. [Google Scholar]
- Moritz, M.A.; Pariesen, M.-A.; Batllori, E.; Krawchuk, M.A.; Van Doren, J.; Ganz, D.J.; Hayhoe, K. Climate change and disruptions to global fire activity. Ecosphere 2012, 3, 1–22. [Google Scholar]
- Spies, T.A.; Franklin, J.F. The structure of natural young, mature, and old-growth forests in Oregon and Washington. In Wildlife and Vegetation of Unmanaged Douglas-fir Forests; General Technical Report PNW-GTR-285; USDA Forest Service, Pacific Northwest Research Station: Portland, OR, USA, 1991; pp. 91–110. [Google Scholar]
- Lindenmeyer, D.B.; Franklin, J.F.; Lohmus, A.; Baker, S.C.; Bauhus, J.; Beese, W.; Brodie, A.; Kiehl, B.; Kouki, J.; Marinez Pastur, G.; et al. A major shift to the retention approach for forestry can help resolve some global forest sustainability issues. Conserv. Lett. 2012. [Google Scholar] [CrossRef]
- Hart, S.C. Email message. Personal Communication. University of California: Merced, CA, USA, August 2012. [Google Scholar]
- Madson, S.L. Correlation between Structural Heterogeneity and Arthropod Biodiversity: Implications for Management of Pacific Northwest Forests. M.Sc. Thesis, Oregon State University, Corvallis, OR, USA, 1997; pp. 1–118. [Google Scholar]
© 2012 by the authors; licensee MDPI, Basel, Switzerland. This article is an open-access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/3.0/).
Share and Cite
Perry, D.A.; Griffiths, R.P.; Moldenke, A.R.; Madson, S.L. Abiotic and Biotic Soil Characteristics in Old Growth Forests and Thinned or Unthinned Mature Stands in Three Regions of Oregon. Diversity 2012, 4, 334-362. https://doi.org/10.3390/d4030334
Perry DA, Griffiths RP, Moldenke AR, Madson SL. Abiotic and Biotic Soil Characteristics in Old Growth Forests and Thinned or Unthinned Mature Stands in Three Regions of Oregon. Diversity. 2012; 4(3):334-362. https://doi.org/10.3390/d4030334
Chicago/Turabian StylePerry, David A., Robert P. Griffiths, Andrew R. Moldenke, and Stephanie L. Madson. 2012. "Abiotic and Biotic Soil Characteristics in Old Growth Forests and Thinned or Unthinned Mature Stands in Three Regions of Oregon" Diversity 4, no. 3: 334-362. https://doi.org/10.3390/d4030334
APA StylePerry, D. A., Griffiths, R. P., Moldenke, A. R., & Madson, S. L. (2012). Abiotic and Biotic Soil Characteristics in Old Growth Forests and Thinned or Unthinned Mature Stands in Three Regions of Oregon. Diversity, 4(3), 334-362. https://doi.org/10.3390/d4030334