
Genetic Impacts of Sustained Stock Enhancement on Wild Populations: A Case Study of Penaeus penicillatus in the Beibu Gulf, China

 , ,
, ,
Abstract
1. Introduction
2. Materials and Methods
2.1. Sample Collection
2.2. DNA Extraction and Microsatellite PCR
2.3. Data Analysis
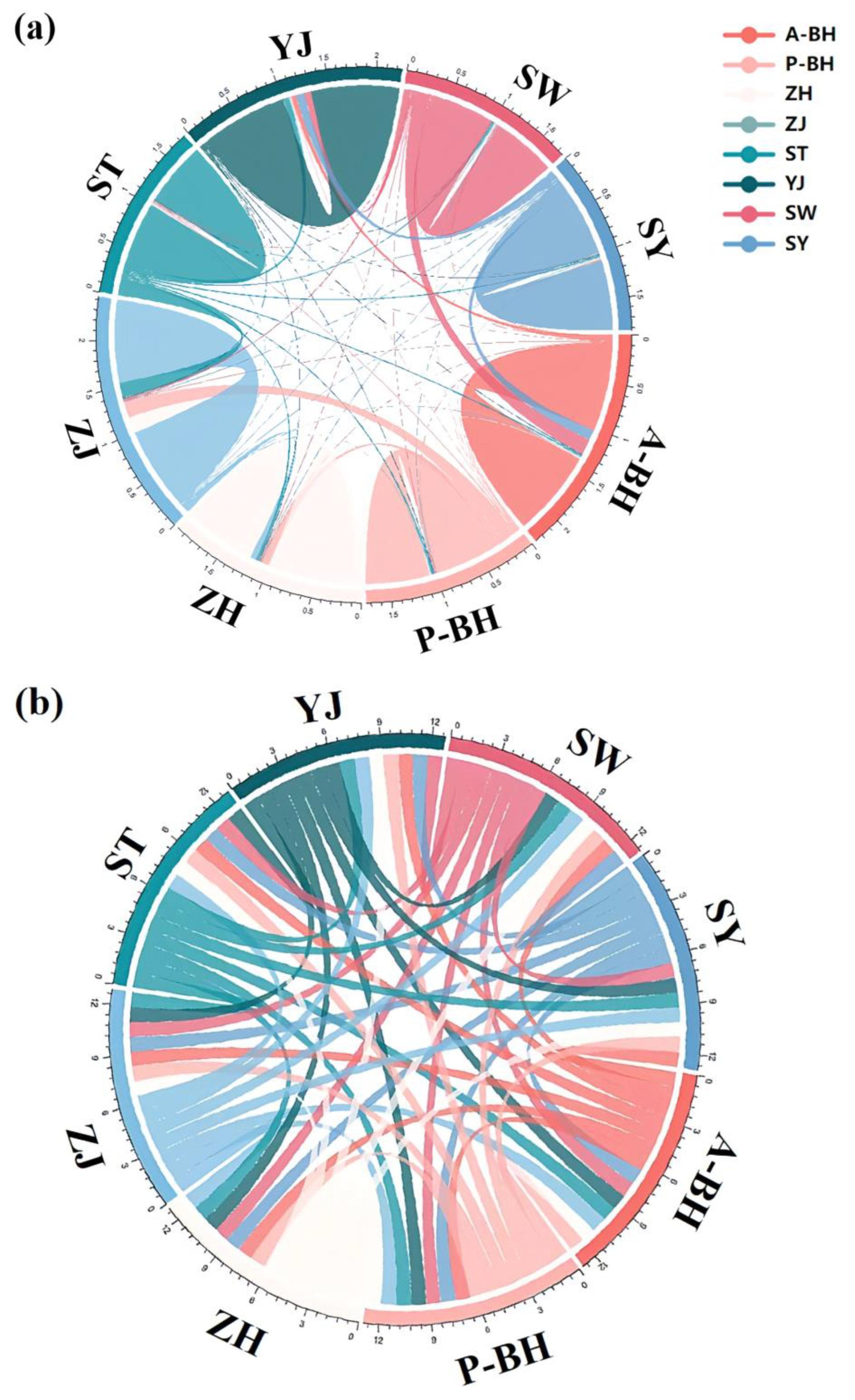
3. Results
3.1. Genetic Diversity
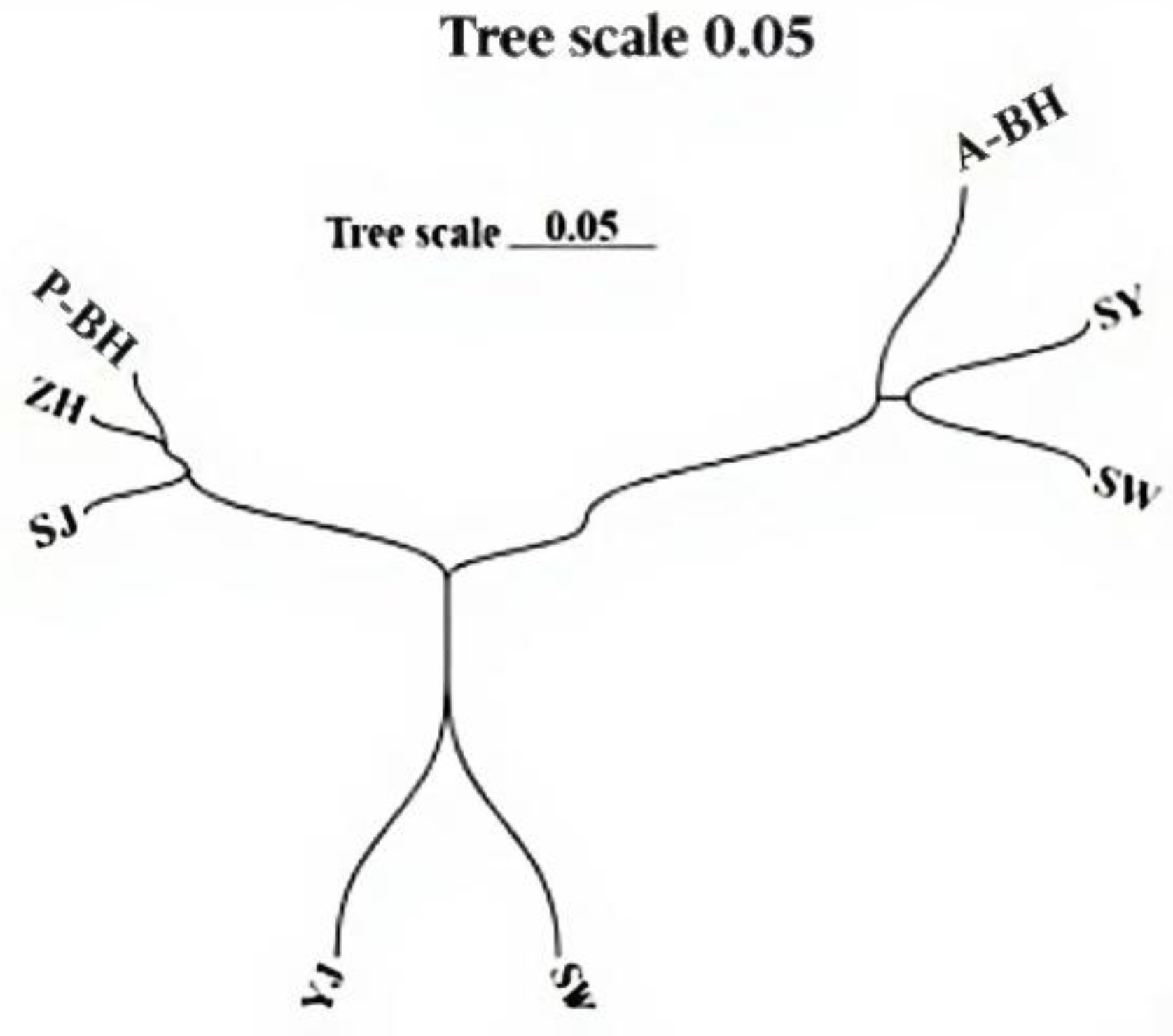
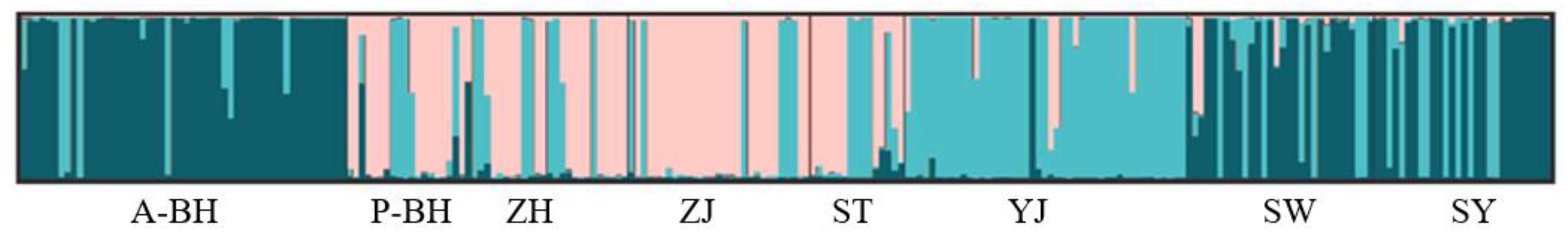
3.2. Genetic Structure
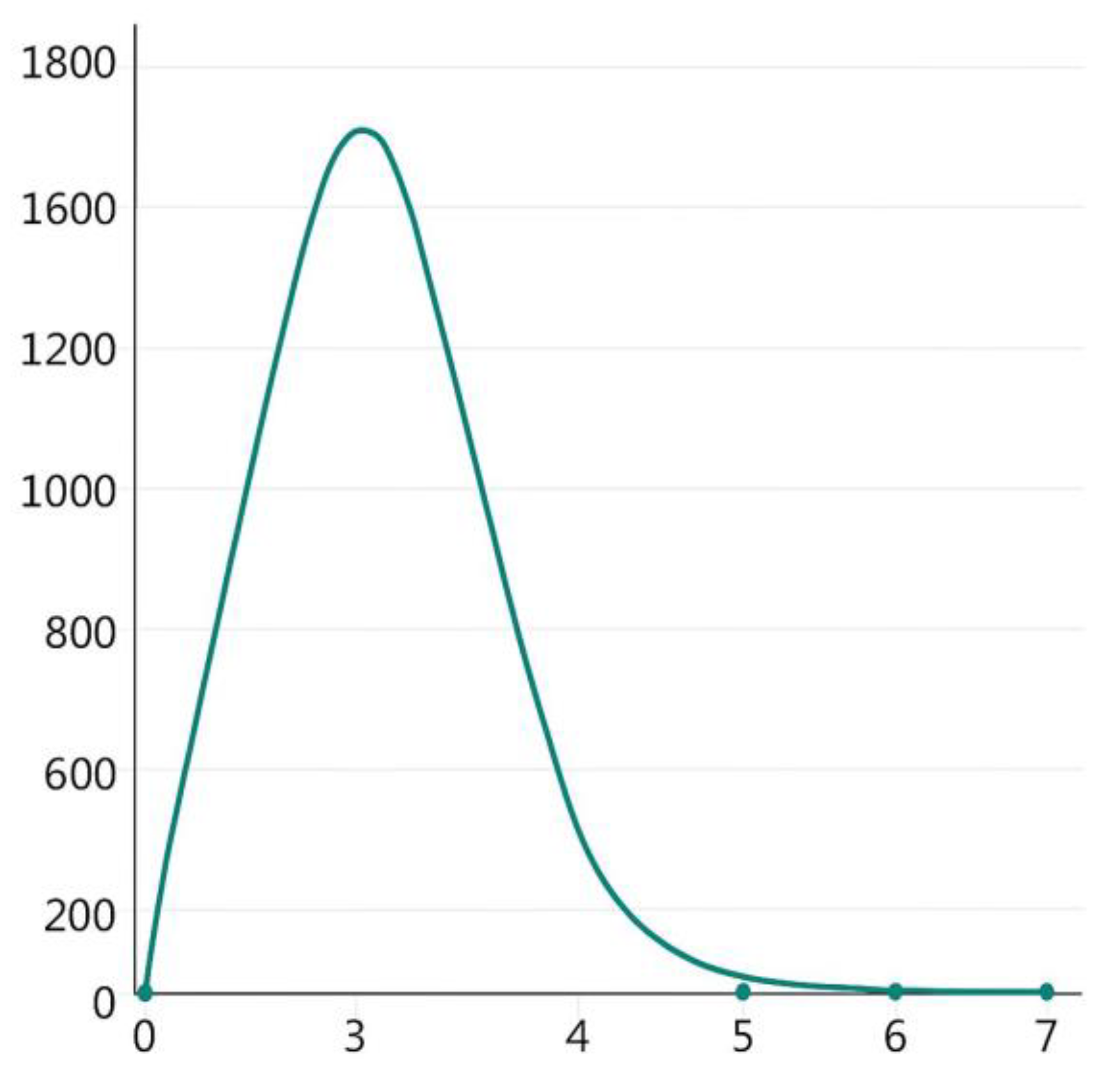
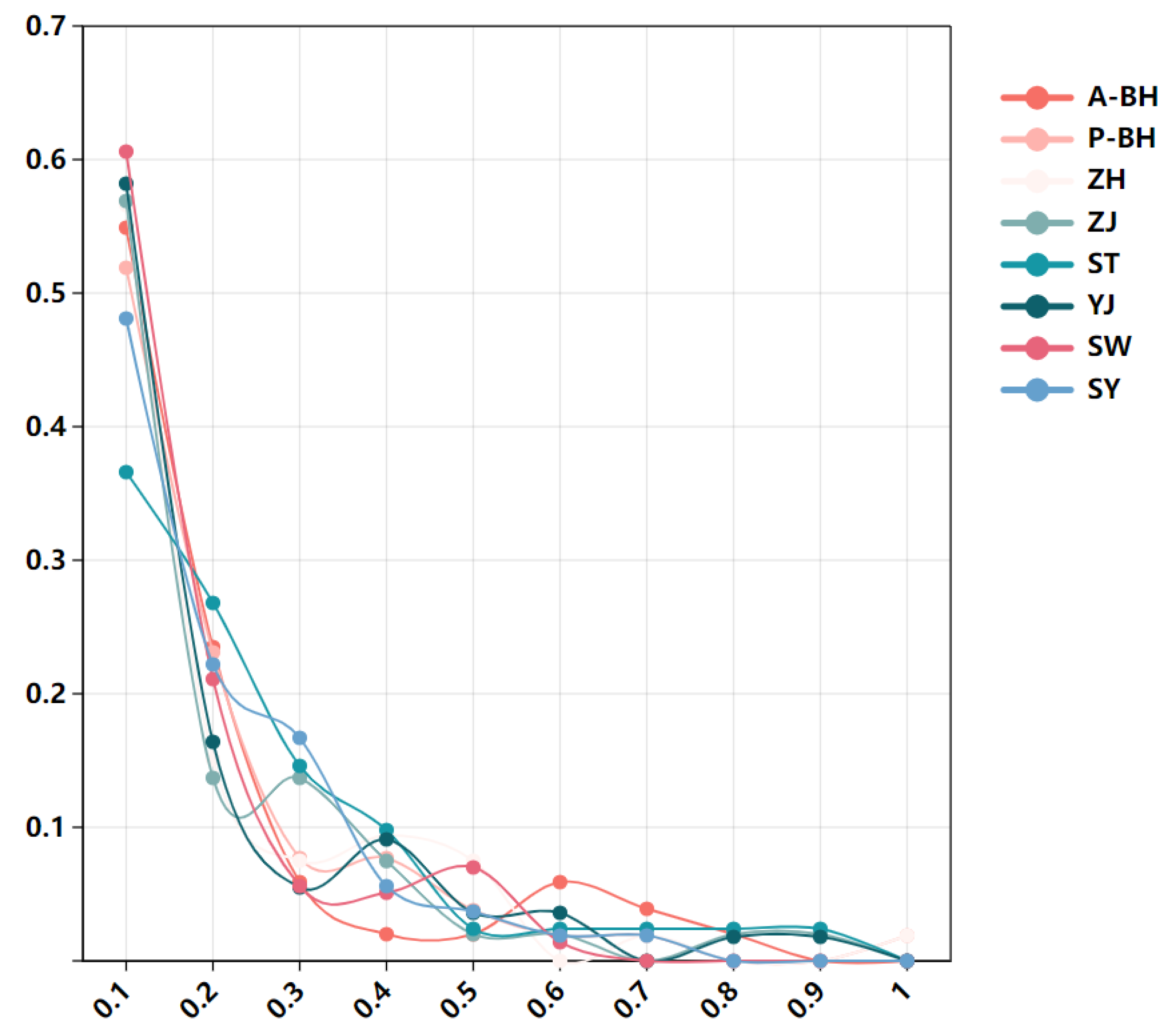
3.3. Demographic Pattern and Gene Flow
4. Discussion
4.1. Genetic Diversity
4.2. Genetic Structure
4.3. Gene Flow and Effective Population Size
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- FAO. The State of World Fisheries and Aquaculture 2022: Towards Blue Transformation; Food and Agriculture Organization of the United Nations: Rome, Italy, 2022. [Google Scholar]
- Han, H.; Chen, Z. The impact of socio-economic factors on the decline of fishery resources. Int. J. Mar. Sci. 2024, 14, 74. [Google Scholar] [CrossRef]
- Yin, J.; Xue, Y.; Xu, B.; Lin, J.; Zhang, M.; Liu, Y.; Wang, Q.; Chen, L.; Zhou, H.; Li, X.; et al. Potential impacts of ocean warming on energy flow and fisheries production in an overexploited ecosystem: Implication for effective fisheries management. Ecol. Indic. 2024, 158, 111433. [Google Scholar] [CrossRef]
- Basheer, V.S.; Ravi, C.; Sarkar, U.K. Conservation of genetic resources: Restoration of fish stocks. In Sustainable Management of Fish Genetic Resources; Springer Nature: Singapore, 2024; pp. 237–248. [Google Scholar]
- Simić, V.; Miljanović, B.; Petrović, A.; Radenković, M.; Stojković Piperac, M.; Veličković, T.; Jakovljević, M.; Simić, S. Inland fisheries in Serbia: Historical aspect, fish resources, management, and conservation. In Ecological Sustainability of Fish Resources of Inland Waters of the Western Balkans: Freshwater Fish Stocks, Sustainable Use and Conservation; Springer International Publishing: Cham, Switzerland, 2024; pp. 113–200. [Google Scholar]
- Bell, J.D.; Bartley, D.M.; Lorenzen, K. Restocking and stock enhancement of coastal fisheries: Potential, problems and progress. Fish. Res. 2006, 80, 1–8. [Google Scholar] [CrossRef]
- Masuda, T.; Shimono, Y.; Kishi, D.; Koizumi, I. Evaluation of genetic consequences of stocking on the southern-margin populations of white-spotted charr. Ecol. Evol. 2024, 14, e70140. [Google Scholar] [CrossRef]
- Zhang, G.L.; Li, Z.B.; Wang, Z.L.; Lin, X.Y.; Wu, N. Study status and perspective of Fenneropenaeus penicillatus. Mod. Fish. Inform. 2010, 2010, 7–10. (In Chinese) [Google Scholar]
- Liang, H.F. Shrimp and Crab Biology; China Agriculture Press: Beijing, China, 2013. (In Chinese) [Google Scholar]
- Wang, J.Q.; Shih, Y.J.; Huang, L.M.; Li, J.; Li, W.W.; Shih, C.-H.; Chu, T.-J. Evaluating the effects related to restocking and stock replenishment of Penaeus penicillatus in the Xiamen Bay, China. J. Mar. Sci. Eng. 2021, 9, 1122. [Google Scholar] [CrossRef]
- Luo, Z.; Yang, C.; Wang, L.; Liu, Y.; Shan, B.; Liu, M.; Sun, D. Spatial and temporal distribution characteristics and diversity changes of fish resources in the coastal waters of the northern Gulf of Beibu. South J. Agric. 2023, 54, 1847–1857. (In Chinese) [Google Scholar]
- Information Office of the Ministry of Agriculture and Rural Affairs (MARA). China and Vietnam jointly hold joint stocking and conservation activities for fishery resources in the Gulf of Beibu Gulf. China Fish. 2017, 6, 4–5. (In Chinese) [Google Scholar]
- Wang, M.; Wang, W.; Xiao, G.; Liu, K.; Hu, Y.; Tian, T.; Kong, J.; Jin, X. Genetic diversity analysis of spawner and recaptured populations of Chinese shrimp (Fenneropenaeus chinensis) during stock enhancement in the Bohai Bay based on an SSR marker. Acta Oceanol. Sin. 2016, 35, 51–56. [Google Scholar] [CrossRef]
- Song, C.; Chen, P.; Lu, W.; Zhang, H.; Han, Q. Genetic diversity of a Phascolosoma esculenta population following stock enhancement in Yueqing Bay, East China Sea. Glob. Ecol. Conserv. 2024, 54, e03058. [Google Scholar] [CrossRef]
- Goldstein, D. Microsatellites: Evolution and Applications; Oxford University Press: Oxford, UK, 1999. [Google Scholar]
- Jarne, P.; Lagoda, P.J. Microsatellites, from molecules to populations and back. Trends Ecol. Evol. 1996, 11, 424–429. [Google Scholar] [CrossRef]
- Herkenhoff, M.E.; Bajay, M.M.; Costamilan, C.A.D.V.L. Genetic structure and biodiversity in wild Centropomus parallelus and in wild and recently domesticated Centropomus undecimallis populations. Life 2023, 13, 1595. [Google Scholar] [CrossRef]
- Tesfaye, G.; Curto, M.; Meulenbroek, P.; Englmaier, G.K.; Tibihika, P.D.; Alemayehu, E.; Getahun, A.; Meimberg, H. Genetic diversity of Nile tilapia (Oreochromis niloticus) populations in Ethiopia: Insights from nuclear DNA microsatellites and implications for conservation. Ecol. Evol. 2021, 21, 113. [Google Scholar] [CrossRef]
- Zhao, Y.; Zhu, X.; Li, Z.; Xu, W.; Dong, J.; Wei, H.; Li, Y.; Li, X. Genetic diversity and structure of Chinese grass shrimp, Palaemonetes sinensis, inferred from transcriptome-derived microsatellite markers. BMC Genet. 2019, 20, 75. [Google Scholar] [CrossRef]
- Liu, S. Development of Microsatellite Markers and Their Application in Stock Enhancement of Penaeus penicillatus. Master’s Thesis, Tianjin Agricultural University, Tianjin, China, 2019. (In Chinese). [Google Scholar]
- Sheng, Q.; Ru, H.; Li, Y.; Ni, Z.H. Current Status and Analysis of the Distribution Pattern of National Aquatic Germplasm Resource Conservation Areas in China. J. Fish. China 2019, 43, 62–80. (In Chinese) [Google Scholar]
- Zhu, Y. Study on the Deep Western Boundary Current in the South China Sea. Ph.D. Thesis, Ocean University of China, Qingdao, China, 2015. (In Chinese). [Google Scholar]
- Rousset, F. Genepop’007: A complete re-implementation of the genepop software for Windows and Linux. Mol. Ecol. Resour. 2008, 8, 103–106. [Google Scholar] [CrossRef] [PubMed]
- Van Oosterhout, C.; Hutchinson, W.F.; Wills, D.M. MICRO-CHECKER: Software for identifying and correcting genotyping errors in microsatellite data. Mol. Ecol. Notes 2004, 4, 535–538. [Google Scholar] [CrossRef]
- Goudet, J. FSTAT (version 1.2): A computer program to calculate F-statistics. J. Hered. 1995, 86, 485–486. [Google Scholar] [CrossRef]
- Rice, W.R. Analyzing tables of statistical tests. Evolution 1989, 43, 223–225. [Google Scholar] [CrossRef]
- Excoffier, L.; Lischer, H.E.L. Arlequin suite ver 3.5: A new series of programs to perform population genetics analyses under Linux and Windows. Mol. Ecol. Resour. 2010, 10, 564–567. [Google Scholar] [CrossRef]
- Populations, version 1.2.30; Free Software Under GNU GPL; Olivier Langella: Gif-sur-Yvette, France, 1999. Available online: https://bioinformatics.org/ (accessed on 8 August 2024).
- Saitou, N.; Nei, M. The neighbor-joining method: A new method for reconstructing phylogenetic trees. Mol. Biol. Evol. 1987, 4, 406–425. [Google Scholar] [CrossRef]
- Evanno, G.; Regnaut, S.; Goudet, J. Detecting the number of clusters of individuals using the software STRUCTURE: A simulation study. Mol. Ecol. 2005, 14, 2611–2620. [Google Scholar] [CrossRef]
- Earl, D.A.; Holdt, B.M. STRUCTURE HARVESTER: A website and program for visualizing STRUCTURE output and implementing the Evanno method. Conserv. Genet. Resour. 2012, 4, 359–361. [Google Scholar] [CrossRef]
- Jombart, T. adegenet: A R package for the multivariate analysis of genetic markers. Bioinformatics 2008, 24, 1403–1405. [Google Scholar] [CrossRef]
- R Core Team. R: A Language and Environment for Statistical Computing; R Foundation for Statistical Computing: Vienna, Austria, 2013. [Google Scholar]
- Piry, S.; Luikart, G.; Cornuet, J.M. BOTTLENECK: A computer program for detecting recent reductions in the effective population size using allele frequency data. J. Hered. 1999, 90, 502–503. [Google Scholar] [CrossRef]
- Do, C.; Waples, R.S.; Peel, D.; Macbeth, G.M.; Tillett, B.J.; Ovenden, J.R. NeEstimator V2: Re-implementation of software for the estimation of contemporary effective population size (Ne) from genetic data. Mol. Ecol. Resour. 2014, 14, 209–214. [Google Scholar] [CrossRef] [PubMed]
- Wilson, G.A.; Rannala, B. Bayesian inference of recent migration rates using multilocus genotypes. Genetics 2003, 163, 1177–1191. [Google Scholar] [CrossRef] [PubMed]
- Beerli, P.; Felsenstein, J. Maximum likelihood estimation of migration matrix and effective population sizes in n subpopulations by way of a coalescent approach. Proc. Natl. Acad. Sci. USA 2001, 98, 4563–4568. [Google Scholar] [CrossRef]
- Beerli, P. Comparison of Bayesian and maximum-likelihood inference of population genetic parameters. Bioinformatics 2006, 22, 2011. [Google Scholar] [CrossRef]
- Beerli, P.; Mashayekhi, S.; Sadeghi, M.; Khodaei, M.; Shaw, K. Population Genetic Inference with MIGRATE. Curr. Protoc. Bioinform. 2019, 68, e87. [Google Scholar] [CrossRef]
- IBM Corp. IBM SPSS Statistics for Windows; Version 25.0; IBM Corp.: Armonk, NY, USA, 2017. [Google Scholar]
- Yuan, Y. Development of Microsatellites and Analysis of Genetic Diversity in Male and Female Populations of Penaeus penicillatus. Master’s Thesis, Jimei University, Xiamen, China, 2016. (In Chinese). [Google Scholar]
- Ada, A.M.; Vandergast, A.G.; Fisher, R.N.; Suárez, A.V.; Bohonak, A.J. Conservation genetics of the endangered California freshwater shrimp (Syncaris pacifica): Watershed and stream networks define gene pool boundaries. Conserv. Genet. 2024, 25, 1021–1033. [Google Scholar] [CrossRef]
- Knibb, W.; Whatmore, P.; Lamont, R.; Nguyen, N.H.; Nguyen, T.T.T.; Moore, S. Can genetic diversity be maintained in long-term mass-selected populations without pedigree information?—A case study using banana shrimp Fenneropenaeus merguiensis. Aquaculture 2014, 428, 71–78. [Google Scholar] [CrossRef]
- Chen, Z.Z.; Lin, Z.J.; Qiu, Y.S. Evaluation of sustainability of fisheries resources for South China Sea based on the AHP. J. Nat. Resour. 2010, 25, 249–257. [Google Scholar]
- Su, L.; Chen, Z.Z.; Zhang, K.; Xu, Y.W.; Qiu, Y.S. Establishment of quality status evaluation system of fishery resources in Beibu Gulf based on bottom trawl survey data. J. Guangdong Ocean Univ. 2021, 41, 10–16. (In Chinese) [Google Scholar]
- Liu, L.; Lin, L.; Li, C.H.; Liu, Y.; Xu, S.N.; Zhou, Y.B. Effect assessment of marine fishery stock enhancement: A review of the literature. Guangdong Agr. Sci. 2014, 41, 133–137+143. (In Chinese) [Google Scholar]
- Blankenship, H.L.; Leber, K.M. A responsible approach to marine stock enhancement. Am. Fish. Soc. Symp. 1995, 15, 167–175. [Google Scholar]
- Bell, J.D.; Leber, K.M.; Blankenship, H.L.; Loneragan, N.R.; Masuda, R. A new era for restocking, stock enhancement and sea ranching of coastal fisheries resources. Rev. Fish. Sci. 2008, 16, 1–9. [Google Scholar] [CrossRef]
- Berg, E.E.; Hamrick, J.L. Quantification of genetic diversity at allozyme loci. Can. J. For. Res. 1997, 27, 415–424. [Google Scholar] [CrossRef]
- Zhang, X.; Sun, D.; Liu, Y.; Wang, Y.; Zhou, Q. Genetic Diversity and Population Structure of Portunus trituberculatus in Released and Wild Populations Based on Microsatellite DNA Markers from the Yangtze Estuary. Sustainability 2023, 15, 11142. [Google Scholar]
- Jarosz, D.F.; Taipale, M.; Lindquist, S. Protein homeostasis and the phenotypic manifestation of genetic diversity: Principles and mechanisms. Annu. Rev. Genet. 2010, 44, 189–216. [Google Scholar] [CrossRef]
- Liu, S.; Sun, D.; Liu, Y.; Yang, C.; Shan, B.; Zhou, W. Evaluation of stock enhancement effectiveness for Penaeus penicillatus in the Beibu Gulf using microsatellite markers. J. Ocean Univ. China (Nat. Sci. Ed.) 2019, 49 (Suppl. S2), 37–45. (In Chinese) [Google Scholar]
- Zhou, Y.; Wang, X.; Li, J.; Zhang, H.; Liu, P. Development of SSR Markers from Genomic Data for Litopenaeus vannamei and Analysis of Genetic Diversity in Different Cultured Populations. J. Fish. China 2023, 47, 108602. [Google Scholar]
- Zhang, M.; Zhao, Y.; Shan, B.; Chen, D.; Huang, W.; Liu, M.; Lin, X. Microsatellite-Marker-Based Evaluation of Stock Enhancement for Kuruma Prawn Penaeus japonicus in Beibu Gulf, South China Sea. Fishes 2023, 8, 568. [Google Scholar] [CrossRef]
- Premachandra, H.K.A.; Becker, A.; Taylor, M.D.; Knibb, W. Eastern King Prawn (Penaeus plebejus) Stock Enhancement—Genetic Evidence That Hatchery-Bred Prawns Have Survived in the Wild After Release. Front. Genet. 2022, 13, 975174. [Google Scholar] [CrossRef]
- Zhang, Y.W.; Pan, X.F.; Wang, X.A.; Fan, W.; Yang, J.X. Restocking of Anabarilius grahami in Lake Fuxian, Southwest China: Morphological and Genetic Effects. Zool. Res. 2020, 41, 741–747. [Google Scholar] [CrossRef]
- An, H.-S.; Hong, S.-W.; Lee, J.-U.; Park, J.-Y.; Kim, K.-K. Genetic diversity of wild and farmed black sea bream (Acanthopagrus schlegelii) populations in Jeju. Anim. Cells Syst. 2010, 14, 37–44. [Google Scholar] [CrossRef]
- Bohonak, A.J. Dispersal, Gene Flow, and Population Structure. Q. Rev. Biol. 1999, 74, 21–45. [Google Scholar] [CrossRef]
- Wei, X.; Fu, Z.; Li, J.; Ye, Y. Genetic Diversity and Structure Revealed by Genomic Microsatellite Markers of Mytilus unguiculatus in the Coast of China Sea. Animals 2023, 13, 1609. [Google Scholar] [CrossRef]
- Li, X.; Gao, S.; Zhao, M.; Zhang, Y.; Liu, Y. SSR Marker-Based Genetic Resource Assessment of the Rainbow Clam Moerella iridescens along the Coasts of China: Implications for Strategy of Conservation Management. Front. Mar. Sci. 2022, 9, 843312. [Google Scholar] [CrossRef]
- Shan, B.; Liu, Y.; Song, N.; Ji, D.; Yang, C.; Zhao, Y.; Gao, T.; Sun, D. Genetic Diversity and Population Structure of Black Sea Bream (Acanthopagrus schlegelii) Based on Mitochondrial Control Region Sequences: The Genetic Effect of Stock Enhancement. Reg. Stud. Mar. Sci. 2020, 35, 101188. [Google Scholar] [CrossRef]
- Hsu, T.-H.; Huang, C.-W.; Lin, C.-H.; Lee, H.-T.; Pan, C.-Y. Tracing the Origin of Fish without Hatchery Information: Genetic Management of Stock Enhancement for Mangrove Red Snapper (Lutjanus argentimaculatus) in Taiwan. Fish. Aquat. Sci. 2020, 23, 13. [Google Scholar] [CrossRef]
- Gonzalez, E.B.; Taniguchi, N.; Umino, T. Can Ordinary Single-Day Egg Collection Increase the Effective Population Size in Broodstock Management Programs? Breeder-Offspring Assignment in Black Sea Bream (Acanthopagrus schlegelii) through Two-Hourly Intervals. Aquaculture 2010, 308, 12–19. [Google Scholar] [CrossRef]
- Wang, W.; Cui, Y.; Jiang, L.; Wang, Y.; Wang, H.; Li, H. Estimation of Effective Population Size of Cynoglossus semilaevis. J. Fish. China 2019, 43, 997–1004. (In Chinese) [Google Scholar]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Time | After Releasing | Before Releasing | ||||||
|---|---|---|---|---|---|---|---|---|
| 2024 | 2017 | 2017 | 2017 | 2017 | 2017 | 2017 | 2017 | |
| Site | Beihai (A-BH) | Beihai (P-BH) | Yangjiang (YJ) | Zhuhai (ZH) | Shantou (ST) | Shanwei (SW) | Sanya (SY) | Zhanjiang (ZJ) |
| Number (n) | 52 | 20 | 45 | 25 | 15 | 30 | 28 | 29 |
| Locus | A-BH n = 52 | P-BH n = 20 | ZH n = 25 | ZJ n = 29 | ST n = 15 | YJ n = 45 | SW n = 30 | SY n = 28 | |
|---|---|---|---|---|---|---|---|---|---|
| CMDX27 | Na | 6 | 7 | 8 | 7 | 5 | 6 | 9 | 7 |
| Ar | 4.99 | 6.68 | 6.64 | 5.81 | 4.56 | 7.35 | 6.43 | 7.02 | |
| Ho | 0.60 | 0.50 | 0.56 | 0.48 | 0.47 | 0.69 | 0.53 | 0.68 | |
| He | 0.60 | 0.79 | 0.79 | 0.77 | 0.77 | 0.60 | 0.77 | 0.76 | |
| Fis | −0.005 | 0.355 | 0.275 | 0.366 | 0.375 | −0.163 | 0.292 | 0.097 | |
| CMDX33 | Na | 5 | 6 | 7 | 7 | 4 | 7 | 4 | 4 |
| Ar | 4.14 | 5.50 | 5.64 | 5.95 | 5.65 | 3.50 | 3.87 | 5.13 | |
| Ho | 0.44 | 0.50 | 0.44 | 0.48 | 0.07 | 0.40 | 0.30 | 0.39 | |
| He | 0.50 | 0.73 | 0.67 | 0.75 | 0.44 | 0.66 | 0.62 | 0.60 | |
| Fis | 0.113 | 0.302 | 0.333 | 0.35 | 0.846 | 0.386 | 0.505 | 0.334 | |
| CMDX74 | Na | 6 | 5 | 4 | 6 | 5 | 5 | 11 | 6 |
| Ar | 5.61 | 4.25 | 3.45 | 5.22 | 3.81 | 7.58 | 5.77 | 7.21 | |
| Ho | 0.62 | 0.10 | 0.04 | 0.07 | 0.07 | 0.02 | 0.80 | 0.54 | |
| He | 0.70 | 0.51 | 0.52 | 0.42 | 0.57 | 0.33 | 0.66 | 0.78 | |
| Fis | 0.116 | 0.800 | 0.922 | 0.832 | 0.879 | 0.932 | −0.235 | 0.299 | |
| CMDX71 | Na | 8 | 9 | 8 | 10 | 8 | 12 | 10 | 8 |
| Ar | 5.73 | 7.99 | 6.89 | 8.19 | 8.02 | 8.29 | 6.76 | 8.68 | |
| Ho | 0.56 | 0.70 | 0.48 | 0.83 | 0.87 | 0.78 | 0.70 | 0.68 | |
| He | 0.49 | 0.80 | 0.80 | 0.79 | 0.81 | 0.82 | 0.74 | 0.75 | |
| Fis | −0.152 | 0.103 | 0.392 | −0.071 | −0.108 | 0.040 | 0.036 | 0.086 | |
| CMDX59 | Na | 8 | 9 | 11 | 11 | 8 | 10 | 11 | 9 |
| Ar | 5.78 | 8.81 | 9.45 | 8.96 | 6.91 | 8.25 | 8.36 | 8.59 | |
| Ho | 0.66 | 0.80 | 0.80 | 0.83 | 0.80 | 0.84 | 0.70 | 0.96 | |
| He | 0.63 | 0.88 | 0.87 | 0.85 | 0.84 | 0.79 | 0.80 | 0.85 | |
| Fis | −0.056 | 0.070 | 0.059 | 0.005 | 0.019 | −0.088 | 0.116 | −0.150 | |
| CMDX25 | Na | 5 | 4 | 6 | 3 | 3 | 4 | 8 | 7 |
| Ar | 4.12 | 3.69 | 4.74 | 2.98 | 3.62 | 7.07 | 5.98 | 5.97 | |
| Ho | 0.67 | 0.50 | 0.72 | 0.41 | 0.67 | 0.58 | 0.57 | 0.89 | |
| He | 0.60 | 0.57 | 0.62 | 0.56 | 0.55 | 0.59 | 0.78 | 0.79 | |
| Fis | −0.138 | 0.109 | −0.186 | 0.248 | −0.255 | 0.004 | 0.258 | −0.144 | |
| CMDX102 | Na | 4 | 4 | 3 | 3 | 3 | 4 | 6 | 6 |
| Ar | 3.51 | 3.44 | 2.79 | 2.85 | 3.12 | 5.62 | 4.77 | 4.74 | |
| Ho | 0.64 | 0.20 | 0.04 | 0.10 | 0.27 | 0.04 | 0.53 | 0.46 | |
| He | 0.54 | 0.19 | 0.19 | 0.22 | 0.35 | 0.42 | 0.71 | 0.50 | |
| Fis | −0.191 | 0.195 | 0.784 | 0.529 | 0.211 | 0.892 | 0.234 | 0.064 | |
| CMDX126 | Na | 9 | 8 | 6 | 4 | 5 | 7 | 12 | 7 |
| Ar | 6.64 | 7.18 | 5.12 | 3.99 | 4.98 | 8.63 | 5.98 | 6.97 | |
| Ho | 0.69 | 0.40 | 0.20 | 0.38 | 0.33 | 0.71 | 0.53 | 0.68 | |
| He | 0.79 | 0.77 | 0.66 | 0.71 | 0.77 | 0.73 | 0.80 | 0.79 | |
| Fis | 0.126 | 0.468 | 0.693 | 0.457 | 0.552 | 0.018 | 0.324 | 0.123 | |
| Mean | Na | 6.4 | 6.5 | 6.3 | 6.4 | 5.1 | 6.9 | 8.9 | 6.8 |
| Ar | 5.02 | 5.94 | 5.59 | 5.49 | 5.08 | 7.03 | 5.99 | 6.79 | |
| Ho | 0.60 | 0.45 | 0.41 | 0.45 | 0.44 | 0.51 | 0.58 | 0.66 | |
| He | 0.60 | 0.66 | 0.64 | 0.63 | 0.64 | 0.63 | 0.64 | 0.62 |
| Linkage Disequilibrium Method (LD) (CL = 95%) | |
|---|---|
| A-BH | 10.1~39.6 |
| P-BH | 8.5~96.4 |
| ZH | 5.0~250.7 |
| ZJ | 10.3~32.6 |
| ST | 2.1~15.4 |
| YJ | 11.6~39.3 |
| SW | 11.8~49.0 |
| SY | 4.3~17.7 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Wu, Y.; Sun, D.; Wang, L.; Liu, Y.; Yang, C.; Liu, M.; Xie, Q.; Chen, C.; Zou, J.; Zhang, D.; et al. Genetic Impacts of Sustained Stock Enhancement on Wild Populations: A Case Study of Penaeus penicillatus in the Beibu Gulf, China. Diversity 2025, 17, 511. https://doi.org/10.3390/d17080511
Wu Y, Sun D, Wang L, Liu Y, Yang C, Liu M, Xie Q, Chen C, Zou J, Zhang D, et al. Genetic Impacts of Sustained Stock Enhancement on Wild Populations: A Case Study of Penaeus penicillatus in the Beibu Gulf, China. Diversity. 2025; 17(8):511. https://doi.org/10.3390/d17080511
Chicago/Turabian StyleWu, Yaxuan, Dianrong Sun, Liangming Wang, Yan Liu, Changping Yang, Manting Liu, Qijian Xie, Cheng Chen, Jianwei Zou, Dajuan Zhang, and et al. 2025. "Genetic Impacts of Sustained Stock Enhancement on Wild Populations: A Case Study of Penaeus penicillatus in the Beibu Gulf, China" Diversity 17, no. 8: 511. https://doi.org/10.3390/d17080511
APA StyleWu, Y., Sun, D., Wang, L., Liu, Y., Yang, C., Liu, M., Xie, Q., Chen, C., Zou, J., Zhang, D., & Shan, B. (2025). Genetic Impacts of Sustained Stock Enhancement on Wild Populations: A Case Study of Penaeus penicillatus in the Beibu Gulf, China. Diversity, 17(8), 511. https://doi.org/10.3390/d17080511