
Soil Nitrogen Transformation Pathways Shift Following Deep Tillage in Coastal Wetlands Invaded by Spartina alterniflora

{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
1. Introduction
2. Materials and Methods
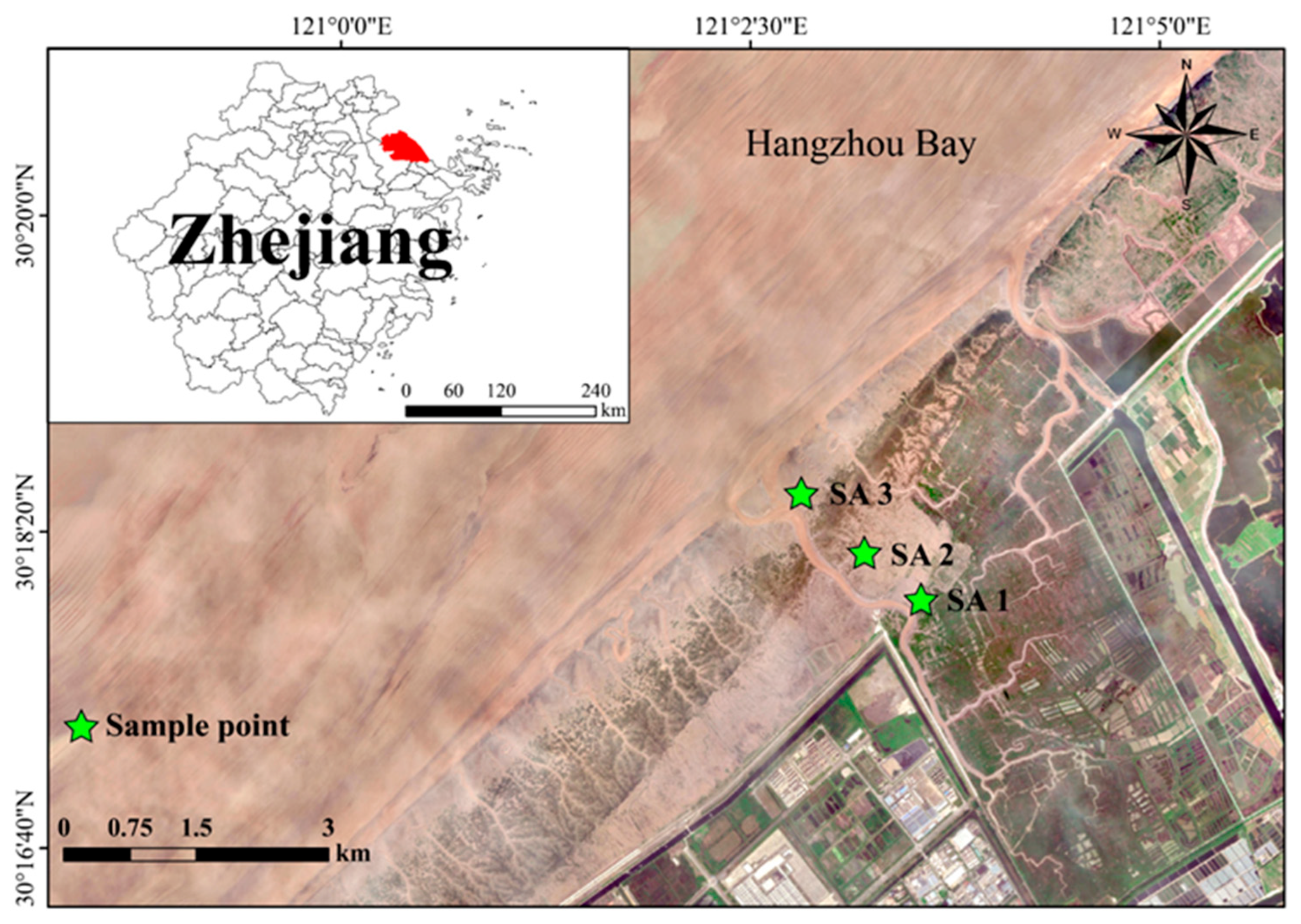
2.1. Study Area
2.2. Experimental Design and Soil Sampling
2.3. Soil Physicochemical Analysis
2.4. Determination of Soil Nitrogen Cycling Rates
2.5. Statistical Analyses
3. Result
3.1. Effects of Deep Tillage on Soil Physicochemical Properties
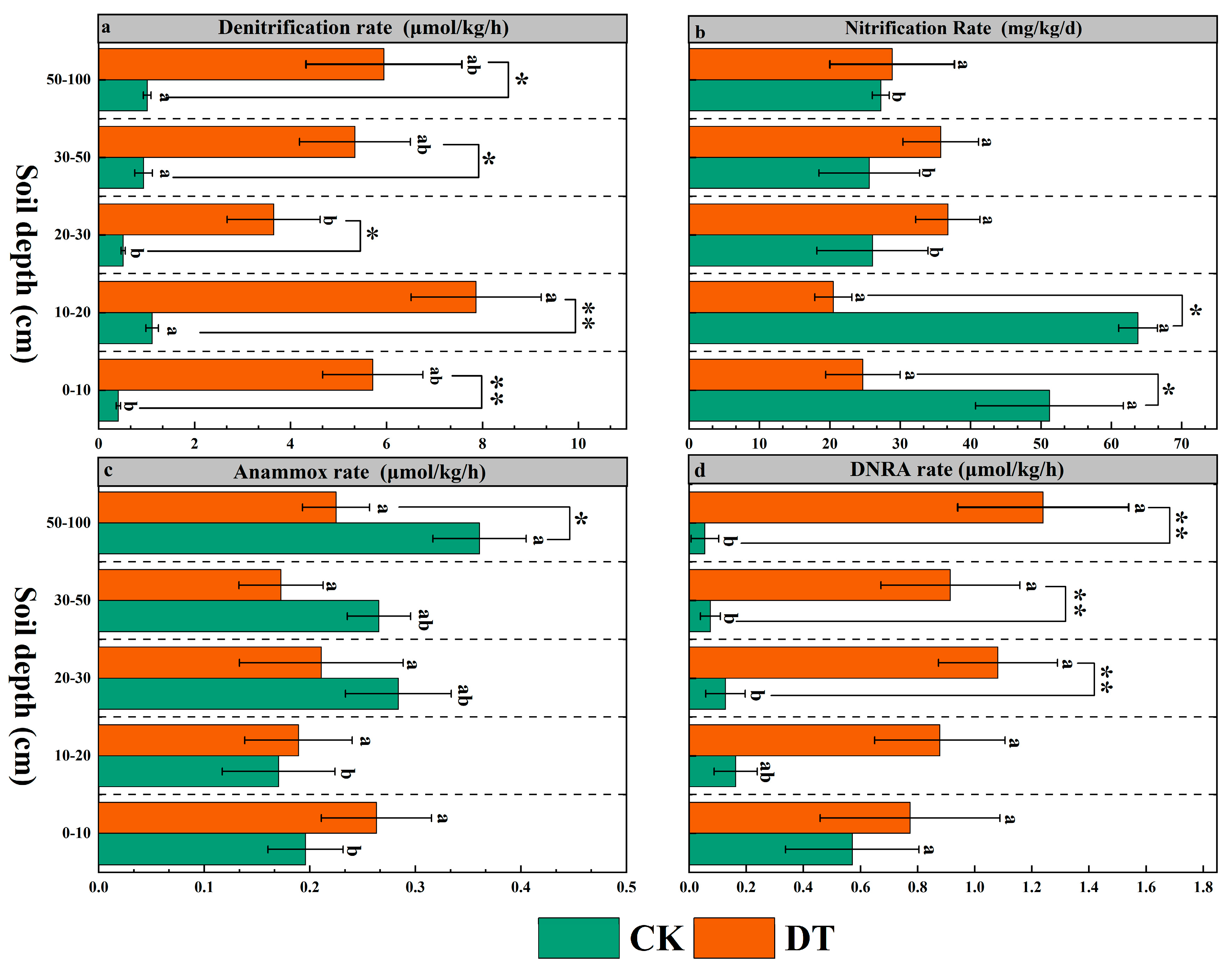
3.2. Changes in Nitrification, Denitrification, Anammox, and DNRA Rates
3.3. Correlations Between Nitrogen Transformation Processes and Soil Physicochemical Properties
4. Discussion
4.1. Effects of Deep Tillage on Soil Properties and Nitrogen Transformation
4.2. Regulation of Nitrogen Transformation by Soil Properties
4.3. Implications for Coastal Wetland Restoration
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Murray, N.J.; Worthington, T.A.; Bunting, P.; Duce, S.; Hagger, V.; Lovelock, C.E.; Lucas, R.; Saunders, M.I.; Sheaves, M.; Spalding, M. High-resolution mapping of losses and gains of Earth’s tidal wetlands. Science 2022, 376, 744–749. [Google Scholar] [CrossRef]
- Yuan, J.; Ding, W.; Liu, D.; Kang, H.; Freeman, C.; Xiang, J.; Lin, Y. Exotic Spartina alterniflora invasion alters ecosystem–atmosphere exchange of CH4 and N2O and carbon sequestration in a coastal salt marsh in China. Glob. Change Biol. 2015, 21, 1567–1580. [Google Scholar] [CrossRef]
- Hinshaw, S.E.; Tatariw, C.; Flournoy, N.; Kleinhuizen, A.; Taylor, C.; Sobecky, P.A.; Mortazavi, B. Vegetation loss decreases salt marsh denitrification capacity: Implications for marsh erosion. Environ. Sci. Technol. 2017, 51, 8245–8253. [Google Scholar] [CrossRef]
- Yu, X.; Yang, J.; Liu, L.; Tian, Y.; Yu, Z. Effects of Spartina alterniflora invasion on biogenic elements in a subtropical coastal mangrove wetland. Environ. Sci. Pollut. Res. 2015, 22, 3107–3115. [Google Scholar] [CrossRef] [PubMed]
- Meng, W.; Feagin, R.A.; Innocenti, R.A.; Hu, B.; He, M.; Li, H. Invasion and ecological effects of exotic smooth cordgrass Spartina alterniflora in China. Ecol. Eng. 2020, 143, 105670. [Google Scholar] [CrossRef]
- Fu, S.; Zheng, S.; Gao, W.; Wang, A.; Ma, X.; Sun, L.; Sun, T.; Shao, D. Effects of the water-sediment regulation scheme on the expansion of Spartina alterniflora at the Yellow River Estuary, China. Front. Environ. Sci. 2021, 9, 642442. [Google Scholar] [CrossRef]
- Yang, W.; An, S.; Zhao, H.; Xu, L.; Qiao, Y.; Cheng, X. Impacts of Spartina alterniflora invasion on soil organic carbon and nitrogen pools sizes, stability, and turnover in a coastal salt marsh of eastern China. Ecol. Eng. 2016, 86, 174–182. [Google Scholar] [CrossRef]
- Gao, D.; Li, X.; Lin, X.; Wu, D.; Jin, B.; Huang, Y.; Liu, M.; Chen, X. Soil dissimilatory nitrate reduction processes in the Spartina alterniflora invasion chronosequences of a coastal wetland of southeastern China: Dynamics and environmental implications. Plant Soil 2017, 421, 383–399. [Google Scholar] [CrossRef]
- Liao, C.; Luo, Y.; Jiang, L.; Zhou, X.; Wu, X.; Fang, C.; Chen, J.; Li, B. Invasion of Spartina alterniflora enhanced ecosystem carbon and nitrogen stocks in the Yangtze Estuary, China. Ecosystems 2007, 10, 1351–1361. [Google Scholar] [CrossRef]
- Zeng, D.; Miao, J.; Wu, G.; Zhan, X. Nitrogen removal, microbial community and electron transport in an integrated nitrification and denitrification system for ammonium-rich wastewater treatment. Int. Biodeterior. Biodegrad. 2018, 133, 202–209. [Google Scholar] [CrossRef]
- Cao, S.; Du, R.; Zhou, Y. Coupling anammox with heterotrophic denitrification for enhanced nitrogen removal: A review. Crit. Rev. Environ. Sci. Technol. 2021, 51, 2260–2293. [Google Scholar] [CrossRef]
- Rohe, L.; Apelt, B.; Vogel, H.-J.; Well, R.; Wu, G.-M.; Schlüter, S. Denitrification in soil as a function of oxygen availability at the microscale. Biogeosciences 2021, 18, 1185–1201. [Google Scholar] [CrossRef]
- Simek, M.; Cooper, J.E.; Picek, T.; Santrucckova, H. Denitrification in arable soils in relation to their physico-chemical properties and fertilization practice. Soil Biol. Biochem. 2000, 32, 101–110. [Google Scholar] [CrossRef]
- Anderson, C.R.; Peterson, M.E.; Frampton, R.A.; Bulman, S.R.; Keenan, S.; Curtin, D. Rapid increases in soil pH solubilise organic matter, dramatically increase denitrification potential and strongly stimulate microorganisms from the Firmicutes phylum. PeerJ 2018, 6, e6090. [Google Scholar] [CrossRef] [PubMed]
- Wendeborn, S. The chemistry, biology, and modulation of ammonium nitrification in soil. Angew. Chem. Int. Ed. 2020, 59, 2182–2202. [Google Scholar] [CrossRef]
- Kumar, M.; Lin, J.G. Co-existence of anammox and denitrification for simultaneous nitrogen and carbon removal—Strategies and issues. J. Hazard. Mater. 2010, 178, 1–9. [Google Scholar] [CrossRef] [PubMed]
- Rutting, T.; Boeckx, P.; Muller, C.; Klemedtsson, L. Assessment of the importance of dissimilatory nitrate reduction to ammonium for the terrestrial nitrogen cycle. Biogeosciences 2011, 8, 1779–1791. [Google Scholar] [CrossRef]
- Wang, S.; Pi, Y.; Song, Y.; Jiang, Y.; Zhou, L.; Liu, W.; Zhu, G. Hotspot of dissimilatory nitrate reduction to ammonium (DNRA) process in freshwater sediments of riparian zones. Water Res. 2020, 173, 115539. [Google Scholar] [CrossRef]
- Herbert, E.R.; Boon, P.; Burgin, A.J.; Neubauer, S.C.; Franklin, R.B.; Ardón, M.; Hopfensperger, K.N.; Lamers, L.P.; Gell, P. A global perspective on wetland salinization: Ecological consequences of a growing threat to freshwater wetlands. Ecosphere 2015, 6, 1–43. [Google Scholar] [CrossRef]
- Ghaly, A.; Ramakrishnan, V. Nitrogen sources and cycling in the ecosystem and its role in air, water and soil pollution: A critical review. J. Pollut. Eff. Control. 2015, 3, 1–26. [Google Scholar]
- Cao, M.; Cui, L.; Sun, H.; Zhang, X.; Zheng, X.; Jiang, J. Effects of Spartina alterniflora invasion on soil microbial community structure and ecological functions. Microorganisms 2021, 9, 138. [Google Scholar] [CrossRef] [PubMed]
- Sheng, Y.; Luan, Z.; Yan, D.; Li, J.; Xie, S.; Liu, Y.; Chen, L.; Li, M.; Wu, C. Effects of Spartina alterniflora invasion on soil carbon, nitrogen and phosphorus in Yancheng coastal wetlands. Lands 2022, 11, 2218. [Google Scholar] [CrossRef]
- Zhang, C.B.; Liu, W.L.; Luo, B.; Guan, M.; Wang, J.; Ge, Y.; Chang, J. Spartina alterniflora invasion impacts denitrifying community diversity and functioning in marsh soils. Geoderma 2020, 375, 114456. [Google Scholar] [CrossRef]
- Wigand, C.; Brennan, P.; Stolt, M.; Holt, M.; Ryba, S. Soil respiration rates in coastal marshes subject to increasing watershed nitrogen loads in southern New England, USA. Wetlands 2009, 29, 952–963. [Google Scholar] [CrossRef]
- Zhang, Y.; Wang, Y.; Li, R.; Zhu, H.; Zhang, F.; Huang, B.; Peng, X. Effects of Spartina alterniflora invasion on active soil organic carbon in the coastal marshes. Chin. J. Soil. Sci. 2012, 43, 102–106. [Google Scholar]
- Lopez Rosas, H.; Moreno-Casasola, P.; Espejel Gonzalez, V.E. Shade treatment affects structure and recovery of invasive C 4 African grass E chinochloa pyramidalis. Ecol. Evol. 2015, 5, 1327–1342. [Google Scholar] [CrossRef]
- Shin, W.; Kim, J.; Song, Y.; Kang, H.; Byun, C. Soil tillage effect on the control of invasive Spartina anglica in a coastal wetland. Wetl. Ecol. Manag. 2024, 32, 397–408. [Google Scholar] [CrossRef]
- Wang, K.; Wang, S.; Zhang, X.; Wang, W.; Li, F.; Dong, L.; Kong, F.; Xi, M. Potential ecological impacts of physical control on Spartina alterniflora in coastal wetland: Migration and transformation of nutrients and the response of bacterial community structure. J. Clean. Prod. 2023, 398, 136556. [Google Scholar] [CrossRef]
- Song, X.; Wang, Y.; Mei, B.; Zhang, X.; Zheng, P.; Song, W.; Tian, B.; Han, G.; Xie, B. Effects of Spartina alterniflora control on soil carbon and nitrogen in coastal wetlands. Watershed Ecol. Environ. 2024, 6, 54–62. [Google Scholar] [CrossRef]
- Wei, W.; Mei, X.; Dai, Z.; Tang, Z. Recent morphodynamic evolution of the largest uninhibited island in the Yangtze (Changjiang) estuary during 1998–2014: Influence of the anthropogenic interference. Cont. Shelf Res. 2016, 124, 83–94. [Google Scholar] [CrossRef]
- Song, Z.; Shi, W.; Zhang, J.; Hu, H.; Zhang, F.; Xu, X. Transport mechanism of suspended sediments and migration trends of sediments in the central Hangzhou Bay. Water 2020, 12, 2189. [Google Scholar] [CrossRef]
- Takamoto, A.; Takahashi, T.; Togami, K. Estimation models from soil pH with a solid-to-liquid ratio of 1: 2.5 to pH measured by other methods using soils in Japan. Soil Sci. Plant Nutr. 2023, 69, 190–198. [Google Scholar] [CrossRef]
- Smith, J.L.; Doran, J.W. Measurement and use of pH and electrical conductivity for soil quality analysis. Methods Assess. Soil Qual. 1997, 49, 169–185. [Google Scholar]
- Yao, X.; Yu, K.; Wang, G.; Deng, Y.; Lai, Z.; Chen, Y.; Jiang, Y.; Liu, J. Effects of soil erosion and reforestation on soil respiration, organic carbon and nitrogen stocks in an eroded area of Southern China. Sci. Total Environ. 2019, 683, 98–108. [Google Scholar] [CrossRef] [PubMed]
- Xie, C.; Xu, J.; Tang, J.; Baig, S.A.; Xu, X. Comparison of phosphorus determination methods by ion chromatography and molybdenum blue methods. Commun. Soil Sci. Plant Anal. 2013, 44, 2535–2545. [Google Scholar] [CrossRef]
- Konen, M.E.; Jacobs, P.M.; Burras, C.L.; Talaga, B.J.; Mason, J.A. Equations for predicting soil organic carbon using loss-on-ignition for north central US soils. Soil Sci. Soc. Am. J. 2002, 66, 1878–1881. [Google Scholar] [CrossRef]
- Hart, S.C.; Stark, J.M.; Davidson, E.A.; Firestone, M.K. Nitrogen mineralization, immobilization, and nitrification. Methods Soil Anal. Part 2 Microbiol. Biochem. Prop. 1994, 5, 985–1018. [Google Scholar]
- Xue, D.; Yu, H.; Fang, Y.; Shan, J.; Xi, D.; Wang, Y.; Kuzyakov, Y.; Wang, Z.-L. 15N-tracer approach to assess nitrogen cycling processes: Nitrate reduction, anammox and denitrification in different pH cropland soils. Catena 2020, 193, 104611. [Google Scholar] [CrossRef]
- Hou, L.; Zheng, Y.; Liu, M.; Li, X.; Lin, X.; Yin, G.; Gao, J.; Deng, F.; Chen, F.; Jiang, X. Anaerobic ammonium oxidation and its contribution to nitrogen removal in China’s coastal wetlands. Sci. Rep. 2015, 5, 15621. [Google Scholar] [CrossRef]
- Thamdrup, B.; Dalsgaard, T. Production of N2 through anaerobic ammonium oxidation coupled to nitrate reduction in marine sediments. Appl. Environ. Microbio. 2002, 68, 1312–1318. [Google Scholar] [CrossRef]
- Yin, G.; Hou, L.; Liu, M.; Liu, Z.; Gardner, W.S. A novel membrane inlet mass spectrometer method to measure 15NH4+ for isotope-enrichment experiments in aquatic ecosystems. Environ. Sci. Technol. 2014, 48, 9555–9562. [Google Scholar] [CrossRef] [PubMed]
- Li, S.; Twilley, R.R. Nitrogen dynamics of inundated sediments in an emerging coastal deltaic floodplain in mississippi river delta using isotope pairing technique to test response to nitrate enrichment and sediment organic matter. Estuar. Coasts. 2021, 44, 1899–1915. [Google Scholar] [CrossRef]
- Sanchez, G. PLS path modeling with R. Berkeley Trowchez Ed. 2013, 383, 551. [Google Scholar]
- Chin, W.W.; Marcolin, B.L.; Newsted, P.R. A partial least squares latent variable modeling approach for measuring interaction effects: Results from a Monte Carlo simulation study and an electronic-mail emotion/adoption study. Inf. Syst. Res. 2003, 14, 189–217. [Google Scholar] [CrossRef]
- Afthanorhan, A.; Awang, Z.; Aimran, N. Five common mistakes for using partial least squares path modeling (PLS-PM) in management research. Contemp. Manag. Res. 2020, 16, 255–278. [Google Scholar] [CrossRef]
- Kang, E.; Li, Y.; Zhang, X.; Yan, Z.; Wu, H.; Li, M.; Yan, L.; Zhang, K.; Wang, J.; Kang, X. Soil pH and nutrients shape the vertical distribution of microbial communities in an alpine wetland. Sci. Total Environ. 2021, 774, 145780. [Google Scholar] [CrossRef]
- Xu, X.; Zhang, Q.; Tan, Z.; Li, Y.; Wang, X. Effects of water-table depth and soil moisture on plant biomass, diversity, and distribution at a seasonally flooded wetland of Poyang Lake, China. Chin. Geogr. Sci. 2015, 25, 739–756. [Google Scholar] [CrossRef]
- Kuzyakov, Y.; Blagodatskaya, E. Microbial hotspots and hot moments in soil: Concept and review. Soil Biol. Biochem. 2015, 83, 184–199. [Google Scholar] [CrossRef]
- Rütting, T.; Schleusner, P.; Hink, L.; Prosser, J.I. The contribution of ammonia-oxidizing archaea and bacteria to gross nitrification under different substrate availability. Soil Biol. Biochem. 2021, 160, 108353. [Google Scholar] [CrossRef]
- Afzal, M.R.; Naz, M.; Ashraf, W.; Du, D. The legacy of plant invasion: Impacts on soil nitrification and management implications. Plants 2023, 12, 2980. [Google Scholar] [CrossRef]
- Bateman, E.; Baggs, E. Contributions of nitrification and denitrification to N2O emissions from soils at different water-filled pore space. Biol. Fertil. Soils 2005, 41, 379–388. [Google Scholar] [CrossRef]
- Hu, B.L.; Shen, L.D.; Zheng, P.; Hu, A.H.; Chen, T.T.; Cai, C.; Liu, S.; Lou, L.P. Distribution and diversity of anaerobic ammonium-oxidizing bacteria in the sediments of the Qiantang River. Environ. Microbiol. Rep. 2012, 4, 540–547. [Google Scholar] [CrossRef] [PubMed]
- Philippot, L.; Cuhel, J.; Saby, N.P.; Chèneby, D.; Chronakova, A.; Bru, D.; Arrouays, D.; Martin-Laurent, F.; Simek, M. Mapping field-scale spatial patterns of size and activity of the denitrifier community. Environ. Microbiol. 2009, 11, 1518–1526. [Google Scholar] [CrossRef] [PubMed]
- Burgin, A.J.; Hamilton, S.K. Have we overemphasized the role of denitrification in aquatic ecosystems? A review of nitrate removal pathways. Front. Ecol. Environ. 2007, 5, 89–96. [Google Scholar] [CrossRef]
- Kartal, B.; Kuenen, J.v.; Van Loosdrecht, M. Sewage treatment with anammox. Science 2010, 328, 702–703. [Google Scholar] [CrossRef]
- Davidson, E.A.; Hart, S.C.; Firestone, M.K. Internal cycling of nitrate in soils of a mature coniferous forest. Ecology 1992, 73, 1148–1156. [Google Scholar] [CrossRef]
- Aber, J.; McDowell, W.; Nadelhoffer, K.; Magill, A.; Berntson, G.; Kamakea, M.; McNulty, S.; Currie, W.; Rustad, L.; Fernandez, I. Nitrogen saturation in temperate forest ecosystems: Hypotheses revisited. BioScience 1998, 48, 921–934. [Google Scholar] [CrossRef]
- Zheng, L.; Wang, X.; Ding, A.; Yuan, D.; Tan, Q.; Xing, Y.; Xie, E. Ecological insights into community interactions, assembly processes and function in the denitrifying phosphorus removal activated sludge driven by phosphorus sources. Front. Microbiol. 2021, 12, 779369. [Google Scholar] [CrossRef]
- Asghar, H.N.; Setia, R.; Marschner, P. Community composition and activity of microbes from saline soils and non-saline soils respond similarly to changes in salinity. Soil Biol. Biochem. 2012, 47, 175–178. [Google Scholar] [CrossRef]
- Wang, B.; Lin, X. Exotic Spartina alterniflora invasion enhances sediment N-loss while reducing N retention in mangrove wetland. Geoderma 2023, 431, 116362. [Google Scholar] [CrossRef]
- Gao, G.-F.; Li, P.-F.; Zhong, J.-X.; Shen, Z.-J.; Chen, J.; Li, Y.-T.; Isabwe, A.; Zhu, X.-Y.; Ding, Q.-S.; Zhang, S. Spartina alterniflora invasion alters soil bacterial communities and enhances soil N2O emissions by stimulating soil denitrification in mangrove wetland. Sci. Total Environ. 2019, 653, 231–240. [Google Scholar] [CrossRef] [PubMed]
- Butterbach-Bahl, K.; Baggs, E.M.; Dannenmann, M.; Kiese, R.; Zechmeister-Boltenstern, S. Nitrous oxide emissions from soils: How well do we understand the processes and their controls? Philos. Trans. R. Soc. B 2013, 368, 20130122. [Google Scholar] [CrossRef] [PubMed]
- Kraft, B.; Strous, M.; Tegetmeyer, H.E. Microbial nitrate respiration–genes, enzymes and environmental distribution. J. Biotech. 2011, 155, 104–117. [Google Scholar] [CrossRef] [PubMed]
- Van den Berg, E.M.; Rombouts, J.L.; Kuenen, J.G.; Kleerebezem, R.; van Loosdrecht, M.C. Role of nitrite in the competition between denitrification and DNRA in a chemostat enrichment culture. Amb. Express 2017, 7, 1–7. [Google Scholar] [CrossRef]
- Philippot, L.; Raaijmakers, J.M.; Lemanceau, P.; Van Der Putten, W.H. Going back to the roots: The microbial ecology of the rhizosphere. Nat. Rev. Microbiol. 2013, 11, 789–799. [Google Scholar] [CrossRef]


Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Gao, J.; Jiang, P.; Li, J.; Wu, M.; Shao, X.; Li, N. Soil Nitrogen Transformation Pathways Shift Following Deep Tillage in Coastal Wetlands Invaded by Spartina alterniflora. Diversity 2025, 17, 473. https://doi.org/10.3390/d17070473
Gao J, Jiang P, Li J, Wu M, Shao X, Li N. Soil Nitrogen Transformation Pathways Shift Following Deep Tillage in Coastal Wetlands Invaded by Spartina alterniflora. Diversity. 2025; 17(7):473. https://doi.org/10.3390/d17070473
Chicago/Turabian StyleGao, Jingwen, Pengcheng Jiang, Junzhen Li, Ming Wu, Xuexin Shao, and Niu Li. 2025. "Soil Nitrogen Transformation Pathways Shift Following Deep Tillage in Coastal Wetlands Invaded by Spartina alterniflora" Diversity 17, no. 7: 473. https://doi.org/10.3390/d17070473
APA StyleGao, J., Jiang, P., Li, J., Wu, M., Shao, X., & Li, N. (2025). Soil Nitrogen Transformation Pathways Shift Following Deep Tillage in Coastal Wetlands Invaded by Spartina alterniflora. Diversity, 17(7), 473. https://doi.org/10.3390/d17070473