
Climate-Driven Range Shifts of the Endangered Cercidiphyllum japonicum in China: A MaxEnt Modeling Approach

 ,
,
Abstract
1. Introduction
- (1)
- to predict the potential suitable distribution areas of C. japonicum in China;
- (2)
- to identify the key environmental factors affecting its distribution;
- (3)
- to provide theoretical support and scientific basis for the conservation, population restoration, and management of C. japonicum.
2. Materials and Methods
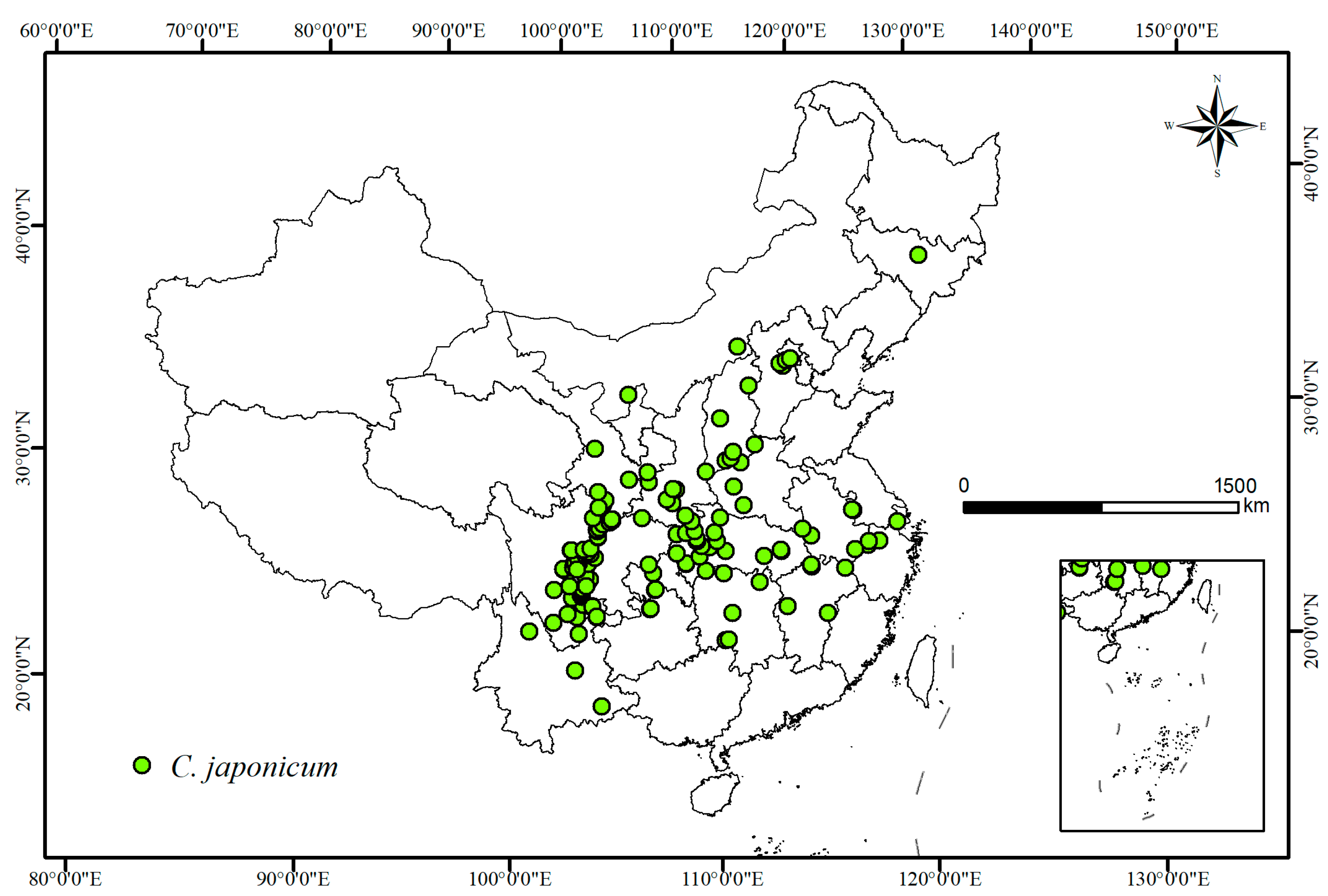
2.1. Species Data Sources and Processing
2.2. Environmental Factors
2.3. MaxEnt Modeling
2.4. Classification of Suitable Grades
3. Results
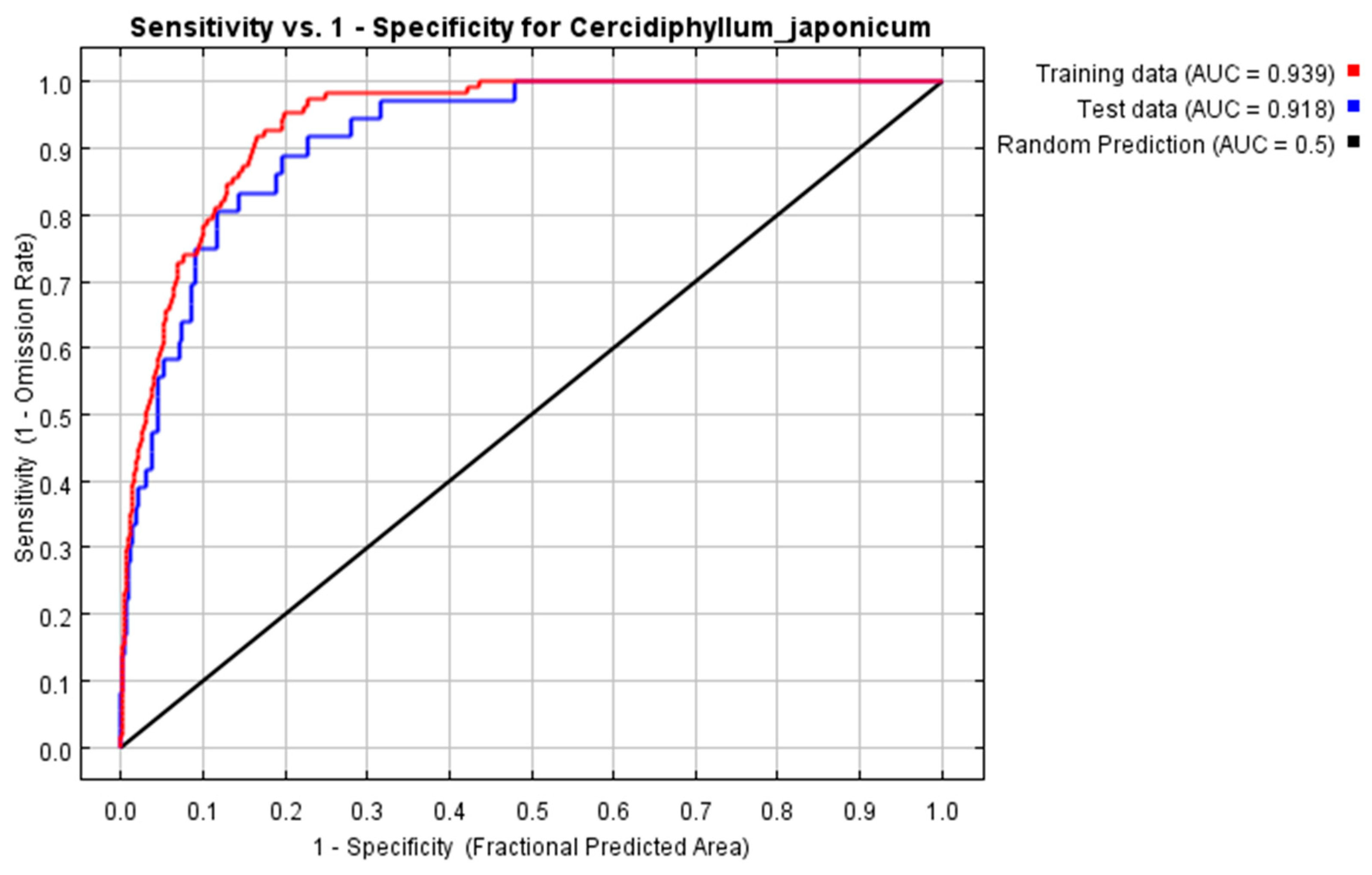
3.1. Model Optimization Results and Accuracy Evaluation
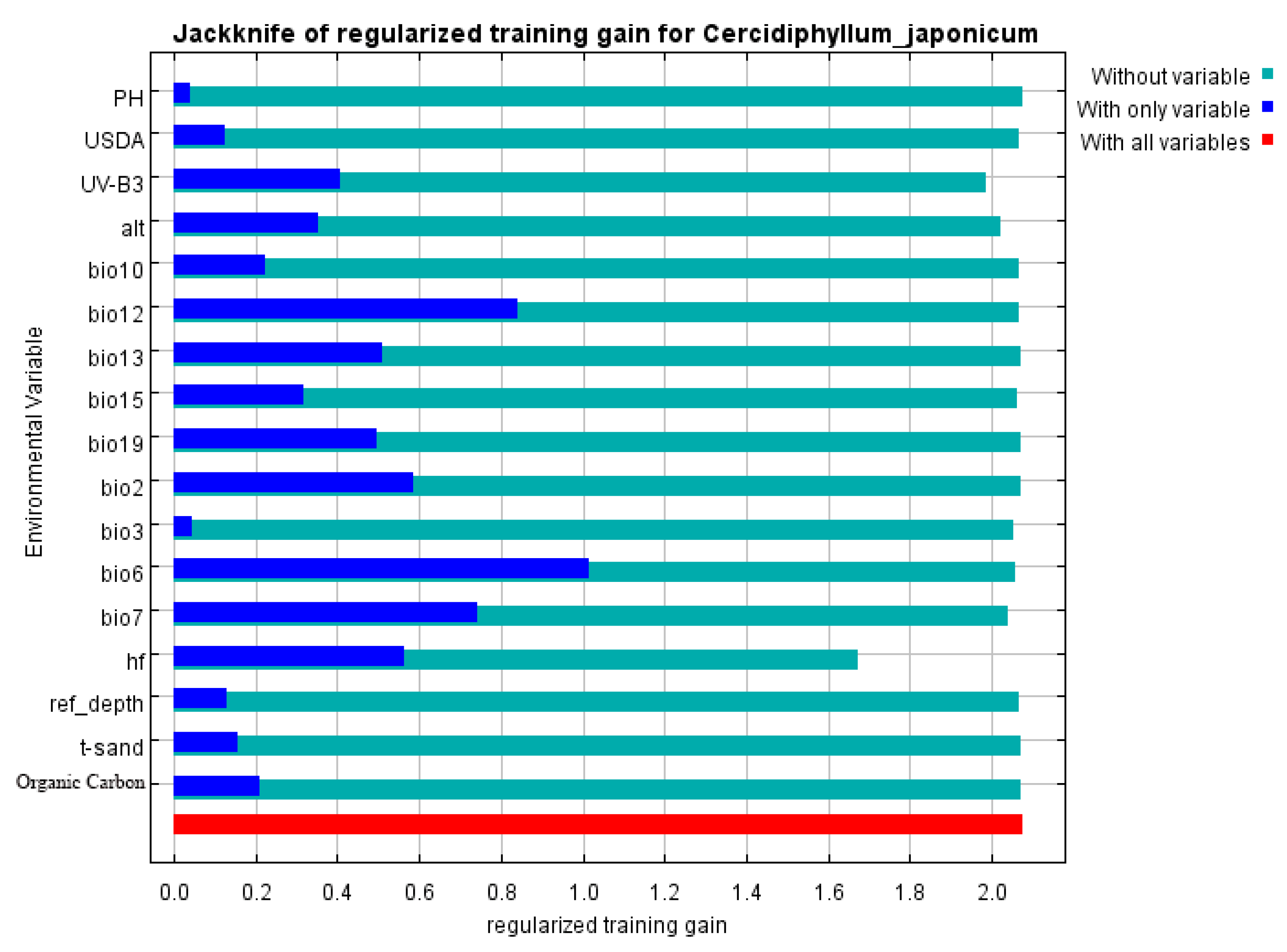
3.2. Contribution Rate of Environmental Variables
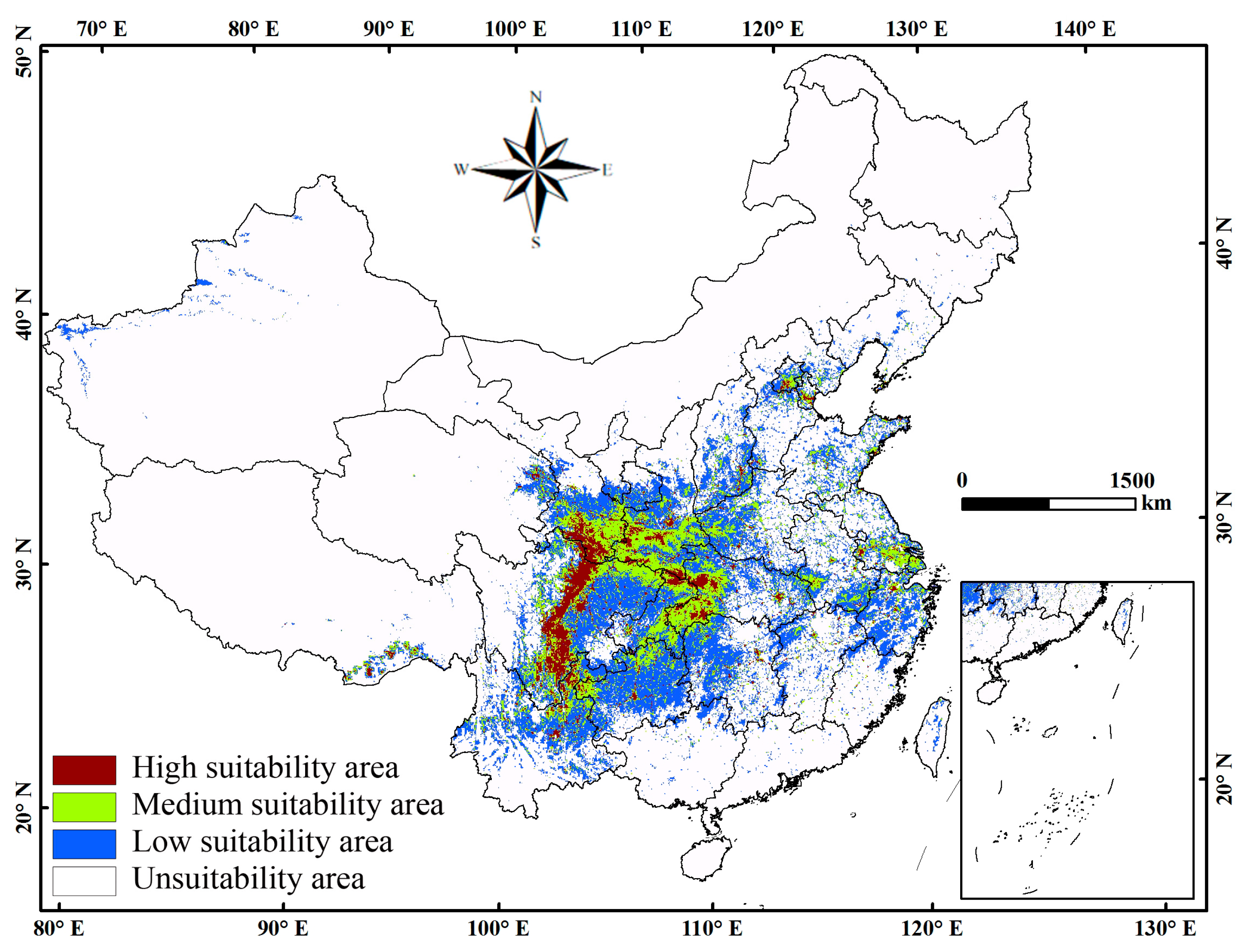
3.3. Current Distribution of C. japonicum Under Present Climate
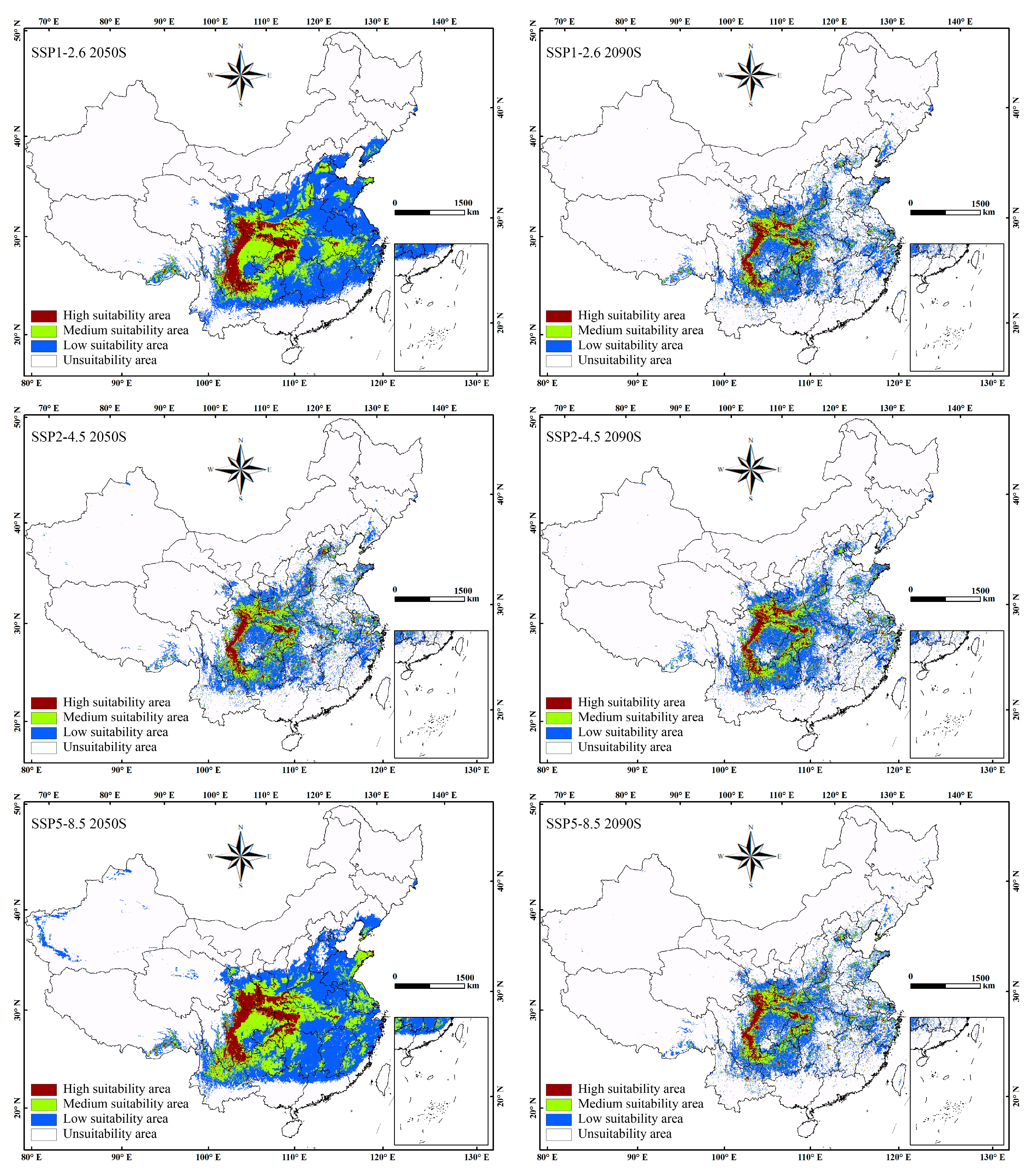
3.4. Potential Distribution of C. japonicum in Future Period
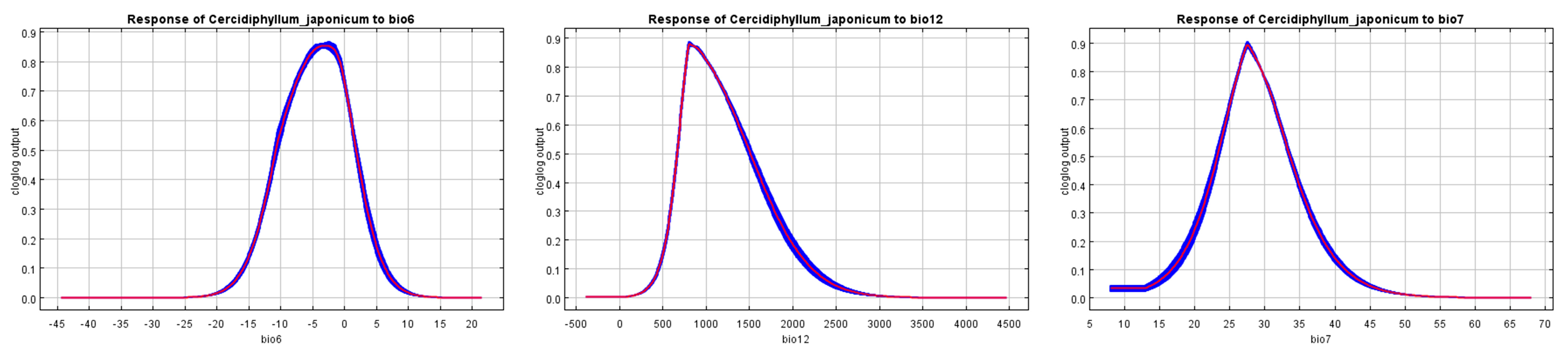
3.5. Environmental Variable Analysis
3.6. The Centroid Variation in the Potential Distribution of C. japonicum
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Conflicts of Interest
References
- Zhu, S.; Yin, P.; Yap, Z.; Qiu, Y. Chloroplast genomes of two extant species of Tertiary relict Cercidiphyllum (Cercidiphyllaceae): Comparative genomic and phylogenetic analyses. Mitochondrial Dna. Part B Resources 2019, 4, 1551–1552. [Google Scholar] [CrossRef]
- Zhang, J.; Zhang, B.; Qian, Z. Functional diversity of Cercidiphyllum japonicum, communities in the Shennongjia Reserve, central China. J. For. Res. 2015, 26, 171–177. [Google Scholar] [CrossRef]
- Sun, Z.; Kumar, R.M.S.; Li, J.; Yang, G.; Xie, Y. In Silico search and biological validation of MicroR171 family related to abiotic stress response in mulberry (Morus alba). Hortic. Plant J. 2022, 8, 184–194. [Google Scholar] [CrossRef]
- Song, N.; Zhang, J. Multivariate Analysis of the Endangered Medicinal Species Cercidiphyllum Japonicum Communities in the Shennongjia Re-serve, Central China. Cerne 2018, 24, 180–189. [Google Scholar] [CrossRef]
- Guan, B.; Chen, W.; Gong, X.; Wu, T.; Cai, Q.; Liu, Y.; Ge, G. Landscape connectivity of Cercidiphyllum japonicum, an endangered species and its implications for conservation. Ecol. Inform. 2016, 33, 51–56. [Google Scholar] [CrossRef]
- Song, X.; Yang, T.; Zhou, C.; Luo, S.; Dong, M.; Yan, X.; Lv, W.; Zheng, F.; Zhou, C.; Wei, Y. Differences in microbiome composition and transcriptome profiles between male and female Paederus fuscipes harbouring pederin-producing bacteria. Insect Mol. Biol. 2022, 31, 457–470. [Google Scholar] [CrossRef] [PubMed]
- Brunel, N.; Meza, F.; Ros, R.; Santibáñez, F. Effects of topsoil loss on wheat productivity in dryland zones of Chile. J. Soil. Sci. Plant Nut. 2011, 11, 129–137. [Google Scholar] [CrossRef]
- Guo, Y.; Li, X.; Zhao, Z.; Wei, H.; Gao, B.; Gu, W. Prediction of the potential geographic distribution of the ectomycorrhizal mushroom Tricholoma matsutake under multiple climate change scenarios. Sci. Rep.-Uk 2017, 7, 46221. [Google Scholar] [CrossRef]
- Merow, C.; Smith, M.J.; Silander, J.A., Jr. A practical guide to MaxEnt for modeling species’ distributions: What it does, and why inputs and settings matter. Ecography (Copenhagen) 2013, 36, 1058–1069. [Google Scholar] [CrossRef]
- Zhao, Z.; Xiao, N.; Shen, M.; Li, J. Comparison between optimized MaxEnt and random forest modeling in predicting potential distribution: A case study with Quasipaa boulengeri in China. Sci. Total Environ. 2022, 842, 156867. [Google Scholar] [CrossRef]
- Imani Wa Rusaati, B.; Won Kang, J. MaxEnt modeling for predicting the potential distribution of Lebrunia bushaie Staner (Clusiaceae) under different climate change scenarios in Democratic Republic of Congo. J. Asia-Pac. Biodivers. 2024, 17, 1–6. [Google Scholar] [CrossRef]
- Leng, W.; He, H.S.; Bu, R.; Dai, L.; Hu, Y.; Wang, X. Predicting the distributions of suitable habitat for three larch species under climate warming in Northeastern China. Forest Ecol. Manag. 2008, 254, 420–428. [Google Scholar] [CrossRef]
- Adams-Phillips, L.; Barry, C.; Giovannoni, J. Signal transduction systems regulating fruit ripening. Trends Plant Sci. 2004, 9, 331–338. [Google Scholar] [CrossRef]
- Wiśniewski, P.; Märker, M. Comparison of Topsoil Organic Carbon Stocks on Slopes under Soil-Protecting Forests in Relation to the Adjacent Agricultural Slopes. Forests 2021, 12, 390. [Google Scholar] [CrossRef]
- Iturbide, M.; Gutiérrez, J.M.; Alves, L.M.; Bedia, J.; Cerezo-Mota, R.; Cimadevilla, E.; Cofiño, A.S.; Di Luca, A.; Faria, S.H.; Gorodetskaya, I.V.; et al. An update of IPCC climate reference regions for subcontinental analysis of climate model data: Definition and aggregated datasets. Earth Syst. Sci. Data 2020, 12, 2959–2970. [Google Scholar] [CrossRef]
- Tu, W.; Xiong, Q.; Qiu, X.; Zhang, Y. Dynamics of invasive alien plant species in China under climate change scenarios. Ecol. Indic. 2021, 129, 107919. [Google Scholar] [CrossRef]
- Naasko, K.I.; Naylor, D.; Graham, E.B.; Couvillion, S.P.; Danczak, R.; Tolic, N.; Nicora, C.; Fransen, S.; Tao, H.; Hofmockel, K.S.; et al. Influence of soil depth, irrigation, and plant genotype on the soil microbiome, metaphenome, and carbon chemistry. Mbio 2023, 14, e175823. [Google Scholar] [CrossRef] [PubMed]
- Wei, X.; Xu, D.; Liu, Q.; Wu, Y.; Zhuo, Z. Predicting the potential distribution range of Batocera horsfieldi under CMIP6 climate change using the MaxEnt model. J. Econ. Entomol. 2024, 117, 187–198. [Google Scholar] [CrossRef]
- Liu, D.; Zeng, Y.; Qiu, C.; Lin, Q. Molecular Cloning and Adversity Stress Expression Analysis of SPDS Genes in Mulberry (Morus notabilis). Russ. J. Plant Physiol. 2021, 68, 1186–1193. [Google Scholar] [CrossRef]
- Li, H.; Wang, Q.; Li, M.; Zang, X.; Wang, Y. Identification of urban waterlogging indicators and risk assessment based on MaxEnt Model: A case study of Tianjin Downtown. Ecol. Indic. 2024, 158, 111354. [Google Scholar] [CrossRef]
- Lihui, M.; Xiaoli, L.; Jie, C.; Youke, W.; Jingui, Y. Effects of Slope Aspect and Rainfall on Belowground Deep Fine Root Traits and Aboveground Tree Height. Front. Plant Sci. 2021, 12, 684468. [Google Scholar] [CrossRef] [PubMed]
- Gao, H.; Qian, Q.; Liu, L.; Xu, D. Predicting the Distribution of Sclerodermus sichuanensis (Hymenoptera: Bethylidae) under Climate Change in China. Insects 2023, 14, 475. [Google Scholar] [CrossRef] [PubMed]
- Shishir, S.; Mollah, T.H.; Tsuyuzaki, S.; Wada, N. Predicting the probable impact of climate change on the distribution of threatened Shorea robusta forest in Purbachal, Bangladesh. Glob. Ecol. Conserv. 2020, 24, e1250. [Google Scholar] [CrossRef]
- Vanhaelewyn, L.; Prinsen, E.; Van Der Straeten, D.; Vandenbussche, F. Hormone-controlled UV-B responses in plants. J. Exp. Bot. 2016, 67, 4469–4482. [Google Scholar] [CrossRef]
- Jain, M.; Bansal, J.; Rajkumar, M.S.; Sharma, N.; Khurana, J.P.; Khurana, P. Draft genome sequence of Indian mulberry (Morus indica) provides a resource for functional and translational genomics. Genomics 2022, 114, 110346. [Google Scholar] [CrossRef]
- He, D.; Jiang, M.; Wei, X. A dendroclimatic investigation of radial growth—climate relationships for the riparian species Cercidiphyllum japonicum in the Shennongjia area, central China. Trees (Berlin, West) 2012, 26, 503–512. [Google Scholar] [CrossRef]
- Tatsumi, C.; Azuma, W.A.; Ogawa, Y.; Komada, N. Nitrogen Availability and Microbial Communities of Canopy Soils in a Large Cercidiphyllum japonicum Tree of a Cool-Temperate Old Growth Forest. Microb. Ecol. 2021, 82, 919–931. [Google Scholar] [CrossRef]
- Zhu, S.; Chen, J.; Zhao, J.; Comes, H.P.; Li, P.; Fu, C.; Xie, X.; Lu, R.; Xu, W.; Feng, Y.; et al. Genomic insights on the contribution of balancing selection and local adaptation to the long-term survival of a widespread living fossil tree, Cercidiphyllum japonicum. New Phytol. 2020, 228, 1674–1689. [Google Scholar] [CrossRef]
- Gerber, R.; Piscart, C.; Roussel, J.; Georges, R.; Houet, T.; Royer, J.; Bergerot, B. Landscape models can predict the distribution of aquatic insects across agricultural areas. Landsc. Ecol. 2023, 38, 2917–2929. [Google Scholar] [CrossRef]
- Sharma, R.; Khan, S.; Kaul, V. Predicting the potential habitat suitability and distribution of “Weed-Onion” (Asphodelus tenuifolius Cavan.) in India under predicted climate change scenarios. J. Agric. Food Res. 2023, 14, 100697. [Google Scholar] [CrossRef]
- Deng, X.; Xu, D.; Liao, W.; Wang, R.; Zhuo, Z. Predicting the distributions of Scleroderma guani (Hymenoptera: Bethylidae) under climate change in China. Ecol. Evol. 2022, 12, e9410. [Google Scholar] [CrossRef] [PubMed]
- Liu, Z.; Zhang, Q.; Wu, X.; Yu, W.; Guo, S. Insecticidal Mechanism of Wintergreen Oil Against the Health Pest Paederus fuscipes (Coleoptera: Staphylinidae). J. Med. Entomol. 2018, 55, 155–162. [Google Scholar] [CrossRef] [PubMed]
- Liu, S.; Chen, T.; Ye, D.; Chen, Q.; Ni, J.; Rao, M. Prediction of distributional patterns of four major Camellia oilseed species in China under climate and land use changes. Ecol. Indic. 2023, 155, 110996. [Google Scholar] [CrossRef]
- Ab Lah, N.Z.; Yusop, Z.; Hashim, M.; Mohd Salim, J.; Numata, S. Predicting the Habitat Suitability of Melaleuca cajuputi Based on the MaxEnt Species Distribution Model. Forests 2021, 12, 1449. [Google Scholar] [CrossRef]
- Zhuo, Z.; Xu, D.; Pu, B.; Wang, R.; Ye, M. Predicting distribution of Zanthoxylum bungeanum Maxim. in China. Bmc Ecol. 2020, 20, 46. [Google Scholar] [CrossRef]
- Soilhi, Z.; Sayari, N.; Benalouache, N.; Mekki, M. Predicting current and future distributions of Mentha pulegium L. in Tunisia under climate change conditions, using the MaxEnt model. Ecol. Inform. 2022, 68, 101533. [Google Scholar] [CrossRef]
- Mapunda, K.K.; Andrew, S.M. Predicting the distribution of critically endangered tree species Karomia gigas under climate change in Tanzania. Ecol. Eng. 2023, 195, 107065. [Google Scholar] [CrossRef]
- Kirillov, V.; Massalova, V.; Pathak, A.; Ivashchenko, A.; Kabanova, S.; Rakhimzhanov, A.; Stikhareva, T. Insights into ex situ conservation and MaxEnt-based habitat suitability modeling of Prunus ulmifolia Franch. (Rosaceae Juss.): A Central Asian relict species. Trees For. People 2025, 20, 100901. [Google Scholar] [CrossRef]
- Curty, C.; Engel, N. Detection, isolation and structure elucidation of a chlorophyll a catabolite from autumnal senescent leaves of Cercidiphyllum japonicum. Phytochemistry 1996, 42, 1531–1536. [Google Scholar] [CrossRef]
- Xia, Z.; Dai, X.; Fan, W.; Liu, C.; Zhang, M.; Bian, P.; Zhou, Y.; Li, L.; Zhu, B.; Liu, S.; et al. Chromosome-level Genomes Reveal the Genetic Basis of Descending Dysploidy and Sex Determination in Morus Plants. Genom. Proteom. Bioinf 2022, 20, 1119–1137. [Google Scholar] [CrossRef]
- Islam, K.N.; Rana, L.R.S.; Islam, K.; Hossain, M.S.; Hossain, M.M.; Hossain, M.A. Climate change and the distribution of two Ficus spp. in Bangladesh—Predicting the spatial shifts. Trees For. People 2021, 4, 100086. [Google Scholar] [CrossRef]
- Chakraborty, A.; Joshi, P.K.; Sachdeva, K. Predicting distribution of major forest tree species to potential impacts of climate change in the central Himalayan region. Ecol. Eng. 2016, 97, 593–609. [Google Scholar] [CrossRef]
- Cajigas Gandia, A.; Alonso Bosch, R.; Mancina, C.A.; Herrel, A. Climatic variation along the distributional range in Cuban Anolis lizards: Species and ecomorphs under future scenarios of climate change. Glob. Ecol. Conserv. 2023, 42, e2401. [Google Scholar] [CrossRef]
- Lin, L.; Xiao-Long, J.; Kai-Qi, G.; Amy, B.; Min, D. Climate change impacts the distribution of Quercus section Cyclobalanopsis (Fagaceae), a keystone lineage in East Asian evergreen broadleaved forests. Plant Diversity 2023, 45, 552–568. [Google Scholar] [CrossRef] [PubMed]
- Swart, C.; Donaldson, J.; Barker, N. Predicting the distribution of Encephalartos latifrons, a critically endangered cycad in South Africa. Biodivers. Conserv. 2018, 27, 1961–1980. [Google Scholar] [CrossRef]
- Zhao, H.; Xian, X.; Guo, J.; Yang, N.; Zhang, Y.; Chen, B.; Huang, H.; Liu, W. Monitoring the little fire ant, Wasmannia auropunctata (Roger 1863), in the early stage of its invasion in China: Predicting its geographical distribution pattern under climate change. J. Integr. Agric. 2023, 22, 2783–2795. [Google Scholar] [CrossRef]
- Ries, G.; Heller, W.; Puchta, H.; Sandermann, H.; Seidlitz, H.K.; Hohn, B. Elevated UV-B radiation reduces genome stability in plants. Nature 2000, 406, 98–101. [Google Scholar] [CrossRef] [PubMed]
- Oishy, M.N.; Shemonty, N.A.; Fatema, S.I.; Mahbub, S.; Mim, E.L.; Raisa, M.B.H.; Anik, A.H. Unravelling the effects of climate change on the soil-plant-atmosphere interactions: A critical review. Soil. Environ. Health 2025, 3, 100130. [Google Scholar] [CrossRef]
- Zheng, S.; Zhu, Y.; Liu, C.; Fan, W.; Xiang, Z.; Zhao, A. Genome-wide identification and characterization of genes involved in melatonin biosynthesis in Morus notabilis (wild mulberry). Phytochemistry 2021, 189, 112819. [Google Scholar] [CrossRef]
- Gao, H. Method of improving the conversion of Cadmium-containing plant biomass energy under the background of soil pollution. Energy Rep. 2022, 8, 10803–10811. [Google Scholar] [CrossRef]
- Hu, X.; Zhang, C.; Shu, Z.; Zhang, Y.; Li, B.; Chu, C. Habitat analysis and conservation priorities for the endangered spotted linsang in China. Glob. Ecol. Conserv. 2024, 56, e3320. [Google Scholar] [CrossRef]



{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Bio-Climatic Variables | Abbreviation | Percent Contribution/% | Permutation Importance/% |
|---|---|---|---|
| Annual Precipitation | Bio12 | 38.4 | 33.8 |
| Min Temperature of Coldest Month | Bio6 | 19.5 | 0.1 |
| Temperature Annual Range (bio5–bio6) | Bio7 | 12.9 | 10.5 |
| Mean Temperature of Warmest Quarter | Bio10 | 10.1 | 10.4 |
| Mean Temperature of Driest Quarter | Bio9 | 8.7 | 0 |
| Isothermality (bio2/bio7) (×100) | Bio3 | 4.3 | 4.7 |
| Precipitation of Coldest Quarter | Bio19 | 1.7 | 7.6 |
| Precipitation Seasonality (Coefficient of Variation) | Bio15 | 1.3 | 2.1 |
| Temperature Seasonality (SD ×100) | Bio4 | 0.8 | 3.8 |
| Precipitation of Wettest Month | Bio13 | 0.8 | 12.8 |
| Mean Temperature of Coldest Quarter | Bio11 | 0.7 | 0.1 |
| Temperature Mean Diurnal Range (Mean of Monthly [max temp–min temp]) | Bio2 | 0.3 | 5.6 |
| Precipitation of Warmest Quarter | Bio18 | 0.3 | 5.7 |
| Precipitation of Driest Quarter | Bio17 | 0.1 | 1.2 |
| Max Temperature of Warmest Month | Bio5 | 0.1 | 0 |
| Precipitation of Driest Month | Bio14 | 0.1 | 1.2 |
| Mean Temperature of Wettest Quarter | Bio8 | 0.1 | 0.3 |
| Annual Mean Temperature | Bio1 | 0 | 0.1 |
| Precipitation of Wettest Quarter | Bio16 | 0 | 0 |
| Variable Classification | Environmental Variables | Unit | Abbreviation |
|---|---|---|---|
| Bio-climatic variables | Mean Temperature of Warmest Quarter | mm | Bio10 |
| Annual Precipitation | mm | Bio12 | |
| Precipitation of Wettest Month | mm | Bio13 | |
| Precipitation Seasonality (Coefficient of Variation) | mm | Bio15 | |
| Precipitation of Coldest Quarter | mm | Bio19 | |
| Mean Diurnal Range (Mean of Monthly [max temp–min temp]) | °C | Bio2 | |
| Isothermality (bio2/bio7) (×100) | % | Bio3 | |
| Min Temperature of Coldest Month | °C | Bio6 | |
| Temperature Annual Range (bio5–bio6) | °C | Bio7 | |
| Soil variables | Soil Reference Depth | m | Ref-depth |
| Soil Acidity and Alkalinity | N/A | pH | |
| Upper Soil Sediment Content | %wt. | T-sand | |
| Organic Carbon Content | %wt. | TOC | |
| Soil Evaluation Indicators | N/A | USDA | |
| Terrain variables | Altitude | m | Alt |
| physical/solar radiation variable | Ultraviolet-B Radiation | nm | UV-B |
| Human variables | Human Footprint | N/A | Hf |
| Variable Classification | Abbreviation | Percent Contribution/% | Permutation Importance/% |
|---|---|---|---|
| Annual Precipitation | Bio12 | 34 | 9.3 |
| Min Temperature of Coldest Month | Bio6 | 19.5 | 14.9 |
| Human Footprint | hf | 15 | 15 |
| Altitude | alt | 12.6 | 23.7 |
| Ultraviolet-B Radiation | UV-B | 7.5 | 16.3 |
| Temperature Annual Range (Bio5–Bio6) | Bio7 | 5.7 | 9.6 |
| Isothermality (Bio2/Bio7) (×100) | Bio3 | 2.5 | 0.8 |
| Soil Reference Depth | Ref-depth | 1.2 | 0.5 |
| Organic Carbon Content | TOC | 0.4 | 0.5 |
| Mean Temperature of Warmest Quarter | Bio10 | 0.4 | 1.2 |
| Precipitation Seasonality (Coefficient of Variation) | Bio15 | 0.3 | 1.1 |
| Precipitation of Coldest Quarter | Bio19 | 0.2 | 1 |
| Soil Evaluation Indicators | USDA | 0.2 | 0.9 |
| Upper Soil Sediment Content | T-sand | 0.2 | 1.2 |
| Soil Acidity and Alkalinity | PH | 0.1 | 0.1 |
| Mean Diurnal Range (Mean of Monthly [max temp–min temp]) | Bio2 | 0.1 | 3.2 |
| Precipitation of Wettest Month | Bio13 | 0.1 | 0.6 |
| Province | High Suitable Area (km2) | Medium Suitable Area (km2) | Low Suitable Area (km2) | No Suitable Area (km2) | Percentage of High Suitable Areas in Province (%) | Percentage of Total Suitable Areas in China (%) |
|---|---|---|---|---|---|---|
| Sichuan | 3650 | 4296 | 7471 | 10,811 | 13.916 | 58.781 |
| Gansu | 1419 | 2324 | 3354 | 16,824 | 5.932 | 29.668 |
| Shanxi | 1080 | 3430 | 2899 | 4331 | 9.199 | 63.109 |
| Hubei | 977 | 2746 | 3440 | 2952 | 9.659 | 70.816 |
| Yunnan | 634 | 1673 | 5782 | 11,657 | 3.211 | 40.965 |
| Chongqing | 458 | 1506 | 1911 | 581 | 10.278 | 86.961 |
| Guizhou | 306 | 1535 | 5551 | 1802 | 3.328 | 80.400 |
| Hunan | 213 | 754 | 4633 | 5568 | 1.907 | 50.143 |
| Henan | 181 | 1116 | 3504 | 4489 | 1.948 | 51.679 |
| Jiangsu | 180 | 912 | 1276 | 3186 | 3.241 | 42.636 |
| Xizang | 168 | 354 | 726 | 64,601 | 0.255 | 1.895 |
| Zhejiang | 139 | 551 | 2668 | 1993 | 2.598 | 62.755 |
| Shaanxi | 130 | 576 | 2739 | 5746 | 1.414 | 37.482 |
| Qinghai | 128 | 260 | 1070 | 39,634 | 0.311 | 3.548 |
| Shandong | 115 | 603 | 2123 | 5993 | 1.302 | 32.160 |
| Anhui | 107 | 818 | 2231 | 4541 | 1.390 | 41.003 |
| Jiangxi | 89 | 289 | 2017 | 6401 | 1.012 | 27.228 |
| Tianjin | 85 | 148 | 188 | 281 | 12.108 | 59.972 |
| Beijing | 81 | 200 | 411 | 300 | 8.165 | 69.758 |
| Hebei | 49 | 370 | 1904 | 8981 | 0.433 | 20.550 |
| Fujian | 25 | 60 | 636 | 5508 | 0.401 | 11.575 |
| Liaoning | 20 | 101 | 541 | 8320 | 0.223 | 7.370 |
| Guangxi | 15 | 48 | 778 | 11,207 | 0.125 | 6.980 |
| Shanghai | 12 | 172 | 110 | 41 | 3.582 | 87.761 |
| Ningxia | 8 | 66 | 457 | 2503 | 0.264 | 17.502 |
| Jilin | 2 | 3 | 87 | 12,171 | 0.016 | 0.750 |
| Taiwan | 2 | 9 | 173 | 1631 | 0.110 | 10.138 |
| Guangdong | 1 | 19 | 145 | 8637 | 0.011 | 1.875 |
| Heilongjiang | 1 | 0 | 10 | 31,278 | 0.003 | 0.035 |
| Hainan | 0 | 0 | 0 | 1561 | 0.000 | 0.000 |
| Inner Mongolia | 0 | 0 | 68 | 74,302 | 0.000 | 0.091 |
| Xinjiang | 0 | 6 | 948 | 100,204 | 0.000 | 0.943 |
| Hong Kong | 0 | 0 | 0 | 52 | 0.000 | 0.000 |
| China | 10,275 | 24,945 | 59,851 | 458,035 | 0.107 | 0.989 |
| Predicted Area (km2) | Comparison with Current Distribution (%) | ||||||
|---|---|---|---|---|---|---|---|
| Decade | Scenarios | High Suitable | Medium Suitable | Low Suitable | High Suitable | Medium Suitable | Low Suitable |
| current | 10,275 | 24,945 | 59,851 | ||||
| 2050s | SSP1-2.6 | 16,821 | 43,086 | 94,738 | 63.708 | 72.724 | 58.290 |
| SSP2-4.5 | 10,160 | 23,674 | 62,092 | −1.119 | −5.095 | 3.744 | |
| SSP5-8.5 | 16,405 | 47,259 | 95,573 | 59.659 | 89.452 | 59.684 | |
| 2090s | SSP1-2.6 | 10,452 | 20,266 | 58,844 | 1.722 | −18.757 | −1.682 |
| SSP2-4.5 | 10,663 | 21,919 | 61,827 | 3.776 | −12.131 | 3.301 | |
| SSP5-8.5 | 10,029 | 21,086 | 56,258 | −2.394 | −15.470 | −6.003 | |
| Environmental Variables | Suitable Range | Optimum Value |
|---|---|---|
| Min Temperature of Coldest Month (bio6)/°C | −12.225–3.132 | −3.175 |
| Annual Precipitation (bio12)/mm | 611.111–1732.320 | 813.131 |
| Temperature Annual Range (bio5–bio6) (bio7)/°C | 21.343–35.940 | 27.456 |
| Scene | Period of Time | Angle/° | Direction | Displacement/km |
|---|---|---|---|---|
| SSP1-2.6 | Contemporary to 2050s | 115.06 | Northwest | 127.19 |
| 2050s to 2090s | 288.92 | Southeast | 118.46 | |
| Contemporary to 2090s | 168.45 | Northwest | 15.78 | |
| SSP2-4.5 | Contemporary to 2050s | 17.47 | Northeast | 138.74 |
| 2050s to 2090s | 107.23 | Northwest | 317.18 | |
| Contemporary to 2090s | 130.89 | Northwest | 345.65 | |
| SSP5-8.5 | Contemporary to 2050s | 127.76 | Northwest | 159.50 |
| 2050s to 2090s | 135.19 | Northwest | 154.47 | |
| Contemporary to 2090s | 20.60 | Northeast | 21.84 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Jiang, Y.; Zhang, H.; Cui, J.; Zheng, L.; Ning, B.; Xu, D. Climate-Driven Range Shifts of the Endangered Cercidiphyllum japonicum in China: A MaxEnt Modeling Approach. Diversity 2025, 17, 467. https://doi.org/10.3390/d17070467
Jiang Y, Zhang H, Cui J, Zheng L, Ning B, Xu D. Climate-Driven Range Shifts of the Endangered Cercidiphyllum japonicum in China: A MaxEnt Modeling Approach. Diversity. 2025; 17(7):467. https://doi.org/10.3390/d17070467
Chicago/Turabian StyleJiang, Yuanyuan, Honghua Zhang, Jun Cui, Lei Zheng, Bingqian Ning, and Danping Xu. 2025. "Climate-Driven Range Shifts of the Endangered Cercidiphyllum japonicum in China: A MaxEnt Modeling Approach" Diversity 17, no. 7: 467. https://doi.org/10.3390/d17070467
APA StyleJiang, Y., Zhang, H., Cui, J., Zheng, L., Ning, B., & Xu, D. (2025). Climate-Driven Range Shifts of the Endangered Cercidiphyllum japonicum in China: A MaxEnt Modeling Approach. Diversity, 17(7), 467. https://doi.org/10.3390/d17070467