
An Exploration of the Genetic Dynamics of Submerged Ceratophyllum demersum in a Freshwater Lake Under Conservation


Abstract

1. Introduction
2. Materials and Methods
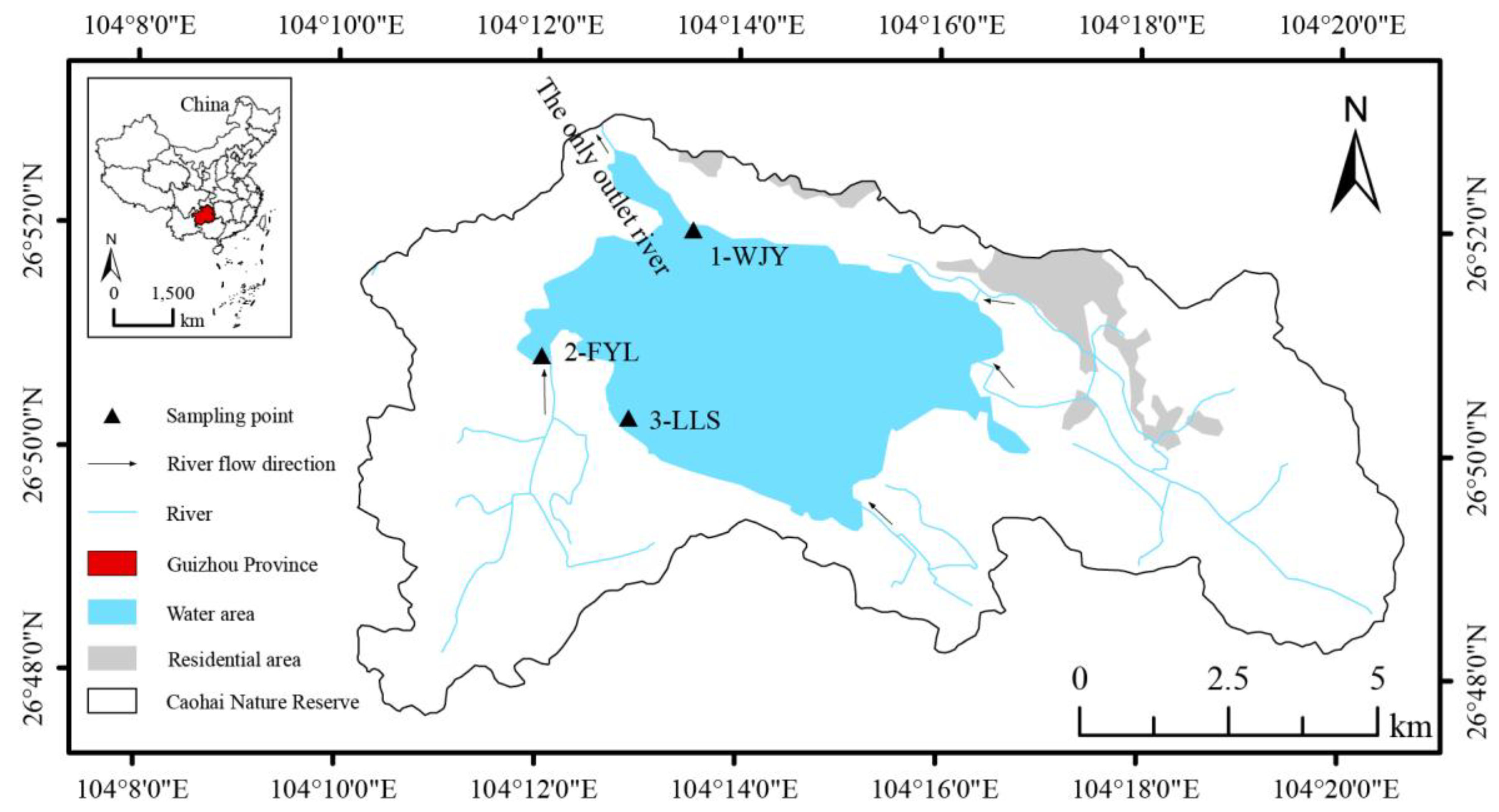
2.1. Sample Collection
2.2. DNA Extraction and PCR Amplification
2.3. Data Analysis
3. Results
3.1. Genetic Diversity
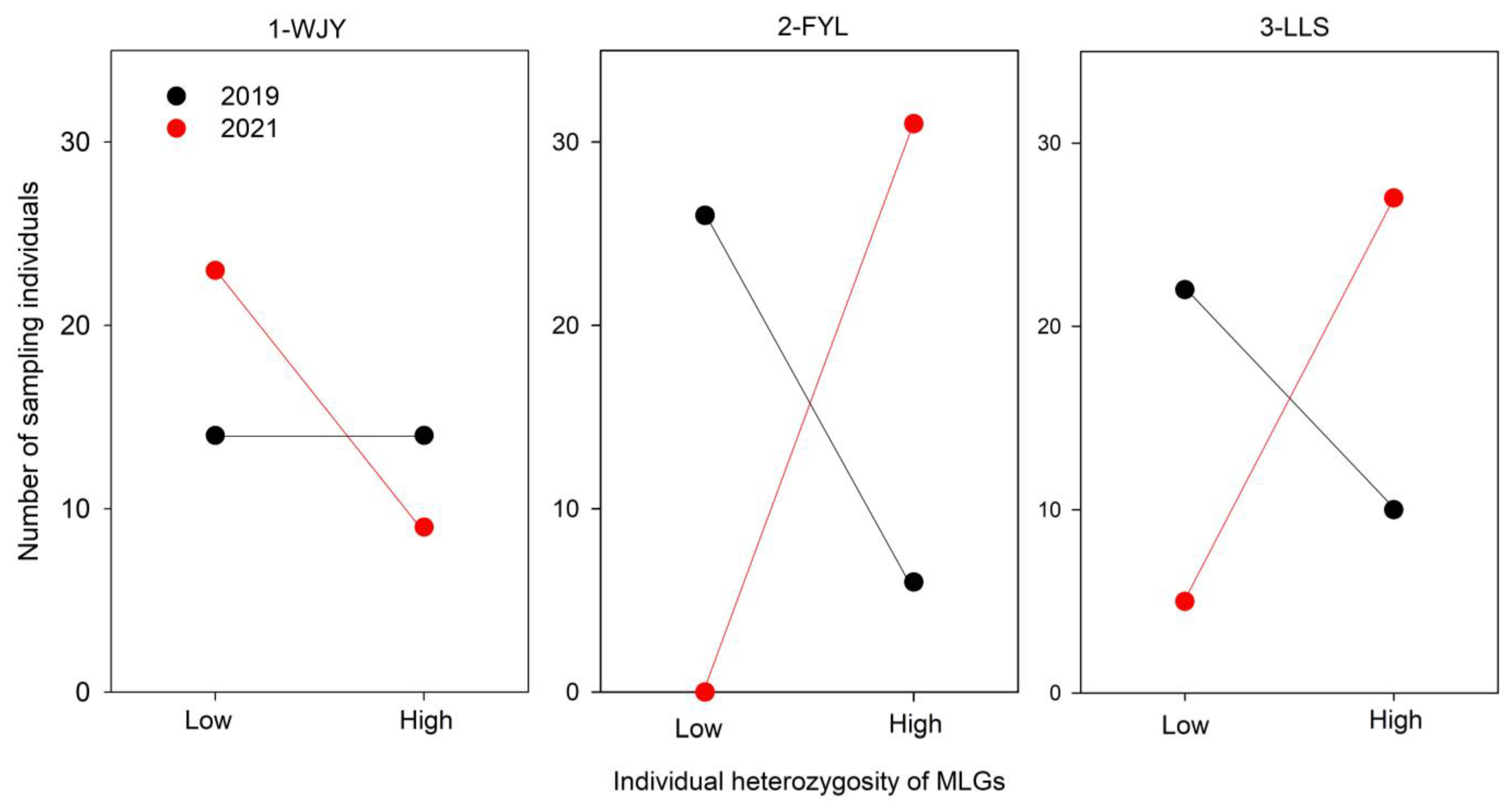
3.2. Clonal Diversity
3.3. Genetic Distance and Estimated Gene Flow Among Subpopulations
4. Discussion
4.1. Genetic and Clonal Diversity
4.2. Genetic Relationships
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Abbreviations
| SSR | Simple sequence repeats |
| MLG | Multi-locus genotype |
| PCoA | Principal coordinate analysis |
References
- Turner, M.G. Disturbance and landscape dynamics in a changing world. Ecology 2010, 91, 2833–2849. [Google Scholar] [CrossRef] [PubMed]
- Yuan, Z.J.; Wu, D.; Niu, L.L.; Ma, X.; Li, Y.; Hillman, A.L.; Abbott, M.B.; Zhou, A. Contrasting ecosystem responses to climatic events and human activity revealed by a sedimentary record from Lake Yilong, southwestern China. Sci. Total Environ. 2021, 783, 146922. [Google Scholar] [CrossRef]
- Cai, F.R. Study on Influencing Factors of Pro-Environmental Behavior of Bird-Watching Tourists in Caohai, Weining. Master’s Thesis, Yunnan University, Kunming, China, 2021. (In Chinese with English abstract). [Google Scholar] [CrossRef]
- Liu, X.; Qiu, Y.; Zheng, Z.L.; Hong, Q.; Zhang, Y.; Qian, Q.; Wan, B.; Chen, Q. Spatiotemporal changes in waterfowl habitat suitability in the Caohai Lake wetland and responses to human activities. Sustainability 2022, 14, 14409. [Google Scholar] [CrossRef]
- Wang, R.L.; Han, Y.; Fan, F.; Molinos, J.G.; Xu, J.; Wang, K.; Wang, D.; Mei, Z. Need to shift in river-lake connection scheme under the “ten-year fishing ban” in the Yangtze River, China. Ecol. Indic. 2022, 143, 109434. [Google Scholar] [CrossRef]
- Bornette, G.; Puijalon, S. Response of aquatic plants to abiotic factors: A review. Aquat. Sci. 2011, 73, 1–14. [Google Scholar] [CrossRef]
- Thomaz, S.M. Ecosystem services provided by freshwater macrophytes. Hydrobiologia 2023, 850, 2757–2777. [Google Scholar] [CrossRef]
- Hughes, A.R.; Stachowicz, J.J. Genetic diversity enhances the resistance of a seagrass ecosystem to disturbance. Proc. Natl. Acad. Sci. USA 2004, 101, 8998–9002. [Google Scholar] [CrossRef]
- Engloner, A.I.; Németh, K.; Kós, P.B.; Meglecz, E.; Bereczki, J. Genetic diversity of the submerged macrophyte Ceratophyllum demersum depends on habitat hydrology and habitat fragmentation. Front. Plant Sci. 2023, 14, 1277916. [Google Scholar] [CrossRef]
- Vásquez, C.; Quinones, R.A.; Brante, A.; Hernandez-Miranda, E. Genetic diversity and resilience in benthic marine populations. Rev. Chil. Hist. Nat. 2023, 96, 4. [Google Scholar] [CrossRef]
- Wang, S.M.; Dou, H.S. Memoirs of Lakes in China; Science Press: Beijing, China, 1998. (In Chinese) [Google Scholar]
- Peng, F.C.; He, T.R.; Li, Z.J.; Chen, M.; Qian, X.; Zeng, L.; Xu, Y. Enrichment characteristics and risk assessment of Hg in bird feathers from Caohai wetland in Guizhou Province, China. Acta Geochim. 2018, 37, 526–536. [Google Scholar] [CrossRef]
- Tu, S.L. Effects of Reed (Phragmites australis) Expansion on Wintering Behavior of Black-Necked Crane (Grus nigricollis) in Caohai Lake Reserve, Guizhou Province. Master’s Thesis, Guizhou Normal University, Guiyang, China, 2023. (In Chinese with English abstract). [Google Scholar]
- Xu, L.; Guo, Y.M.; Liu, D.; Li, R.; Du, M. Temporal and influencing factors of Caohai water quality in recent ten years. Environ. Prot. Sci. 2024, 50, 127–133, (In Chinese with English abstract). [Google Scholar] [CrossRef]
- Flora of China Editorial Committee. Flora of China. 2024. Available online: http://www.iplant.cn (accessed on 29 March 2024). (In Chinese).
- Lou, J. The Quantitative Study of Aquatic Vegetation Community and Human Disturbance in Caohai, Guizhou. Master’s Thesis, Guizhou University, Guiyang, China, 2006. (In Chinese with English abstract). [Google Scholar]
- Dai, L.L.; Chen, X.; Li, L.; Liu, C.; Yuan, G. Aquatic plant diversity and community succession in Caohai wetland, Guizhou Province. Acta Hydrobiol. Sin. 2020, 44, 869–876, (In Chinese with English abstract). [Google Scholar]
- Triest, L.; Tran Thi, V.; Le Thi, D.; Sierens, T.; Geert, A.V. Genetic differentiation of submerged plant populations and taxa between habitats. Hydrobiologia 2010, 656, 15–27. [Google Scholar] [CrossRef]
- Cao, W.; Wang, Q.; Cao, Q.J. Fine-scale spatial genetic structure and gene dispersal in lake populations of submerged species. J. Oceanol. Limnol. 2025, 43, 831–847. [Google Scholar] [CrossRef]
- Les, D.H. Breeding systems, population structure, and evolution in hydrophilous angiosperms. Ann. Mo. Bot. Gard. 1988, 75, 819–835. [Google Scholar] [CrossRef]
- Foroughi, M.; Najafi, P.; Toghiani, A.; Honarjoo, N. Analysis of pollution removal from wastewater by Ceratophyllum demersum. Afr. J. Biotechnol. 2010, 9, 2125–2128. [Google Scholar]
- Dai, Y.; Jia, C.; Liang, W.; Hu, S.; Wu, Z. Effects of the submerged macrophyte Ceratophyllum demersum L. on restoration of a eutrophic waterbody and its optimal coverage. Ecol. Eng. 2012, 40, 113–116. [Google Scholar] [CrossRef]
- Les, D.H. Genetic diversity in the monoecious hydrophile Ceratophyllum (Ceratophyllaceae). Am. J. Bot. 1991, 78, 1070–1082. [Google Scholar] [CrossRef]
- Reynolds, C.; Cumming, G.S. Seed dispersal by waterbirds in southern Africa: Comparing the roles of ectozoochory and endozoochory. Freshw. Biol. 2016, 61, 349–361. [Google Scholar] [CrossRef]
- Capers, R.S. Macrophyte colonization in a freshwater tidal wetland (Lyme, CT, USA). Aquat. Bot. 2003, 77, 325–338. [Google Scholar] [CrossRef]
- Engloner, A.I.; Németh, K.; Bereczki, J. The genetic diversity of the macrophyte Ceratophyllum demersum in Backwaters reflects differences in the hydrological connectivity and water flow rate of habitats. Plants 2024, 13, 2220. [Google Scholar] [CrossRef] [PubMed]
- Chao, F.; Jiang, X.; Wang, X.; Lu, B.; Liu, J.; Xia, P. Water level fluctuation rather than eutrophication induced the extinction of submerged plants in Guizhou’s Caohai Lake: Implications for lake management. Water 2024, 16, 772. [Google Scholar] [CrossRef]
- Zhu, Y. Dynamics of Waterbirds Community and Their Exposure Risk of Heavy Metal Exposure in Caohai Wetland, Guizhou Province, China. Master’s Thesis, Guizhou University, Guiyang, China, 2020. (In Chinese with English abstract). [Google Scholar] [CrossRef]
- Saghai-Maroof, M.A.; Soliman, K.M.; Jorgensen, R.A.; Allard, R.W. Ribosomal DNA spacer-length polymorphisms in barley: Mendelian inheritance, chromosomal location, and population dynamics. Proc. Natl. Acad. Sci. USA 1984, 81, 8014–8018. [Google Scholar] [CrossRef]
- Peakall, R.; Smouse, P.E. GenAlEx 6.5: Genetic analysis in excel. Population genetic software for teaching and research-an update. Bioinformatics 2012, 28, 2537–2539. [Google Scholar] [CrossRef] [PubMed]
- Hardy, O.J.; Vekemans, X. SPAGeDi: A versatile computer program to analyse spatial genetic structure at the individual or population levels. Mol. Ecol. Notes 2002, 2, 618–620. [Google Scholar] [CrossRef]
- Meirmans, P.G. GENODIVE version 3.0: Easy-to-use software for the analysis of genetic data of diploids and polyploids. Mol. Ecol. Resour. 2020, 20, 1126–1131. [Google Scholar] [CrossRef]
- Dorken, M.E.; Eckert, C.G. Severely reduced sexual reproduction in northern populations of a clonal plant, Decodon verticillatus (Lythraceae). J. Ecol. 2001, 89, 339–350. [Google Scholar] [CrossRef]
- Hämmerli, A.; Reusch, T.B.H. Inbreeding depression influences genet size distribution in a marine angiosperm. Mol. Ecol. 2003, 12, 619–629. [Google Scholar] [CrossRef]
- Li, Y.X.; Zhao, X.Y.; Xia, M.L.; Wei, X.; Hou, H. Temperature is a cryptic factor shaping the geographical pattern of genetic variation in Ceratophyllum demersum across a subtropical freshwater lake. Plant Divers. 2024, 46, 630–639. [Google Scholar] [CrossRef]
- Cao, Q.J.; Mei, F.; Wang, L. Population genetic structure in six sympatric and widespread aquatic plants inhabiting diverse lake environments in China. Ecol. Evol. 2017, 7, 5713–5723. [Google Scholar] [CrossRef]
- Hidding, B.; Meirmans, P.G.; Klaassen, M.; Ouborg, T.d.B.N.J.; Wagemaker, C.A.M.; Nolet, B.A. The effect of herbivores on genotypic diversity in a clonal aquatic plant. Oikos 2014, 123, 1112–1120. [Google Scholar] [CrossRef]
- Wu, Z.B.; Zuo, J.C.; Ma, J.M.; Wu, J.; Cheng, S.; Liang, W. Establishing submersed macrophytes via sinking and colonization of shoot fragments clipped off manually. Wuhan Univ. J. Nat. Sci. 2007, 12, 553–557. [Google Scholar] [CrossRef]
- Bilton, D.T.; Paula, J.; Bishop, J.D.D. Dispersal, genetic differentiation and speciation in estuarine organisms. Estuar. Coast. Shelf Sci. 2002, 55, 937–952. [Google Scholar] [CrossRef]
- Cao, Q.J.; Liu, B.; Hu, F. Effects of hydrological connection and human disturbance on genetic variation of submerged Vallisneria natans populations in four lakes in China. J. Oceanol. Limnol. 2021, 39, 1403–1416. [Google Scholar] [CrossRef]
- Young, A.; Boyle, T.; Brown, T. The population genetic consequences of habitat fragmentation for plants. Trends Ecol. Evol. 1996, 11, 413–418. [Google Scholar] [CrossRef]
- Soares, L.; Cazetta, E.; Santos, L.R.; Franca, D.d.S.; Gaiotto, F.A. Anthropogenic disturbances eroding the genetic diversity of a threatened palm tree: A multiscale approach. Front. Genet. 2019, 10, 1090. [Google Scholar] [CrossRef]




{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Characteristic | 2019 | 2021 | ||||
|---|---|---|---|---|---|---|
| 1-WJY | 2-FYL | 3-LLS | 1-WJY | 2-FYL | 3-LLS | |
| Water depth (m) | 1.2–2.0 | <0.5 | <0.8 | 1.3–2.1 | <0.6 | <0.9 |
| TP in water (mg L−1) | 0.092 | 0.101 | 0.097 | 0.095 | 0.099 | 0.096 |
| TN in water (mg L−1) | 1.067 | 1.367 | 1.384 | 1.301 | 1.660 | 1.598 |
| Genetic | Clonal | ||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Allelic | Ramet Level | Genet Level | |||||||||||
| Subpopulation-Year | N | P (%) | Na | Nt | Np | Ar | Ho | He | Ho | He | G | Ge | R |
| 1-WJY-2019 a | 28 | 88.9 | 3.000 B | 27 | 1 | 2.98 B | 0.645 A | 0.527 A | 0.640 A | 0.597 B | 7 | 3.7 | 0.222 |
| 1-WJY-2021 | 32 | 100 | 3.667 C | 33 | 7 | 3.45 C | 0.618 A | 0.400 A | 0.648 A | 0.500 A | 12 | 4.8 | 0.355 |
| 2-FYL-2019 | 32 | 77.8 | 1.778 A | 16 | 9 | 1.75 A | 0.587 A | 0.319 A | 0.587 A | 0.354 A | 7 | 3.5 | 0.194 |
| 2-FYL-2021 | 31 | 88.9 | 1.889 A | 17 | 10 | 1.89 A | 0.735 A | 0.397 A | 0.778 A | 0.476 A | 4 | 2.8 | 0.100 |
| 3-LLS-2019 | 32 | 100 | 3.889 C * | 35 | 12 | 3.87 C * | 0.614 A | 0.472 A | 0.676 A | 0.640 B | 9 | 2.4 | 0.258 |
| 3-LLS-2021 | 32 | 88.9 | 2.556 B | 23 | 0 | 2.56 B | 0.736 A | 0.481 A | 0.667 A | 0.570 A | 3 | 1.6 | 0.065 |
| Average—2019 | 31 | 88.9 | 2.889 (1.060) b | 26 (9.5) | 7 (5.7) | 2.87 (1.06) | 0.615 (0.029) | 0.440 (0.108) | 0.634 (0.045) | 0.531 (0.154) | 7.7 (1.2) | 3.2 (0.7) | 0.225 (0.032) |
| Average—2021 | 32 | 92.6 | 2.704 (0.898) | 24 (8.1) | 6 (5.1) | 2.63 (0.78) | 0.696 (0.068) | 0.426 (0.048) | 0.697 (0.070) | 0.515 (0.049) | 6.3 (4.9) | 3.1 (1.6) | 0.174 (0.158) |
| Total population—2019 | 92 | 100 | 4.333 | 39 | 6 | 4.28 | 0.614 | 0.452 | 0.635 | 0.575 | 19 | 9.1 | 0.198 |
| Total population—2021 | 95 | 100 | 4.222 | 38 | 4 | 4.17 | 0.697 | 0.591 | 0.679 | 0.581 | 17 | 7.5 | 0.170 |
| Subpopulation | ||||||||||
|---|---|---|---|---|---|---|---|---|---|---|
| 1-WJY | 2-FYL | 3-LLS | ||||||||
| 2019 | 2021 | 2019 | 2021 | 2019 | 2021 b | |||||
| Locus | Allele 1 | Allele 1 | Allele 2 | Allele 1 | Allele 2 | Allele 1 | Allele 2 | Allele 1 | Allele 2 | |
| CdL024 | 121 a (0.017) | 125 (0.016) | 127 (0.500) | 119 (0.047) | 127 (0.016) | |||||
| CdL034 | 256 (0.500) | 266 (0.500) | 268 (1.000) | 260 (0.047) | 268 (0.016) | |||||
| CdL041 | 155 (0.113) | 163 (0.016) | ||||||||
| CdL042 | 100 (0.031) | 110 (0.047) | 108 (1.000) | 100 (0.500) | 112 (0.500) | 114 (0.016) | ||||
| CdL060 | 140 (0.047) | 185 (0.813) | 203 (0.500) | |||||||
| CdL070 | 184 (0.047) | 196 (0.500) | 184 (0.500) | 190 (0.016) | 192 (0.047) | |||||
| CdL077 | 269 (0.500) | 253 (0.500) | ||||||||
| CdL087 | 207 (0.018) | 217 (0.016) | 205 (0.047) | 207 (0.031) | ||||||
| CdL093 | 212 (0.033) | 234 (0.500) | 236 (0.500) | 212 (0.806) | 246 (0.194) | 164 (0.031) | 250 (0.031) | |||
| Subpopulation-Year | 1-WJY-2019 | 2-FYL-2019 | 3-LLS-2019 | 1-WJY-2021 | 2-FYL-2021 | 3-LLS-2021 |
|---|---|---|---|---|---|---|
| 1-WJY-2019 | 0 | |||||
| 2-FYL-2019 | 0.042 | 0 | ||||
| 3-LLS-2019 | 0.060 | 0.012 | 0 | |||
| 1-WJY-2021 | 0.034 | 0 | ||||
| 2-FYL-2021 | 0.847 | 0.773 | 0 | |||
| 3-LLS-2021 | 0.178 | 0.277 | 0.692 | 0 |
| 2019 | 2021 | ||||||||
|---|---|---|---|---|---|---|---|---|---|
| Locus | Fis | Fit | Fst | Nm | Locus | Fis | Fit | Fst | Nm |
| CdL024 | 0.337 | 0.391 | 0.082 | 2.799 | CdL024 | −0.450 | 0.023 | 0.326 | 0.516 |
| CdL034 | −0.689 | −0.631 | 0.035 | 6.962 | CdL034 | −0.941 | 0.005 | 0.487 | 0.263 |
| CdL041 | 0.933 | 0.948 | 0.224 | 0.867 | CdL041 | 0.134 | 0.184 | 0.058 | 4.090 |
| CdL042 | 0.355 | 0.434 | 0.122 | 1.794 | CdL042 | −0.449 | 0.122 | 0.394 | 0.384 |
| CdL060 | 0.094 | 0.159 | 0.071 | 3.249 | CdL060 | −0.653 | −0.152 | 0.303 | 0.575 |
| CdL070 | −0.721 | −0.678 | 0.025 | 9.799 | CdL070 | −0.806 | −0.479 | 0.181 | 1.132 |
| CdL077 | −0.611 | −0.569 | 0.026 | 9.190 | CdL077 | −0.794 | −0.491 | 0.169 | 1.227 |
| CdL087 | −0.711 | −0.702 | 0.006 | 43.733 | CdL087 | −0.726 | −0.512 | 0.124 | 1.768 |
| CdL093 | −0.720 | −0.676 | 0.026 | 9.510 | CdL093 | −0.630 | −0.151 | 0.294 | 0.600 |
| Mean (SE) | −0.193 (0.210) | −0.147 (0.211) | 0.069 (0.023) | 9.767 (4.403) | Mean (SE) | −0.591 (0.105) | −0.161 (0.091) | 0.260 (0.046) | 1.173 (0.398) |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Wang, Q.; Cao, Q.-J. An Exploration of the Genetic Dynamics of Submerged Ceratophyllum demersum in a Freshwater Lake Under Conservation. Diversity 2025, 17, 413. https://doi.org/10.3390/d17060413
Wang Q, Cao Q-J. An Exploration of the Genetic Dynamics of Submerged Ceratophyllum demersum in a Freshwater Lake Under Conservation. Diversity. 2025; 17(6):413. https://doi.org/10.3390/d17060413
Chicago/Turabian StyleWang, Qian, and Qian-Jin Cao. 2025. "An Exploration of the Genetic Dynamics of Submerged Ceratophyllum demersum in a Freshwater Lake Under Conservation" Diversity 17, no. 6: 413. https://doi.org/10.3390/d17060413
APA StyleWang, Q., & Cao, Q.-J. (2025). An Exploration of the Genetic Dynamics of Submerged Ceratophyllum demersum in a Freshwater Lake Under Conservation. Diversity, 17(6), 413. https://doi.org/10.3390/d17060413