
Mixed-Species Flock Diversity and Habitat Density Are Associated with Antipredator Behavior in Songbirds

 ,
,  , and
, and 
Abstract
1. Introduction
2. Materials and Methods
2.1. Study Site
2.2. Baseline and Predator Stimulus
2.3. Data Coding and Analysis
3. Results
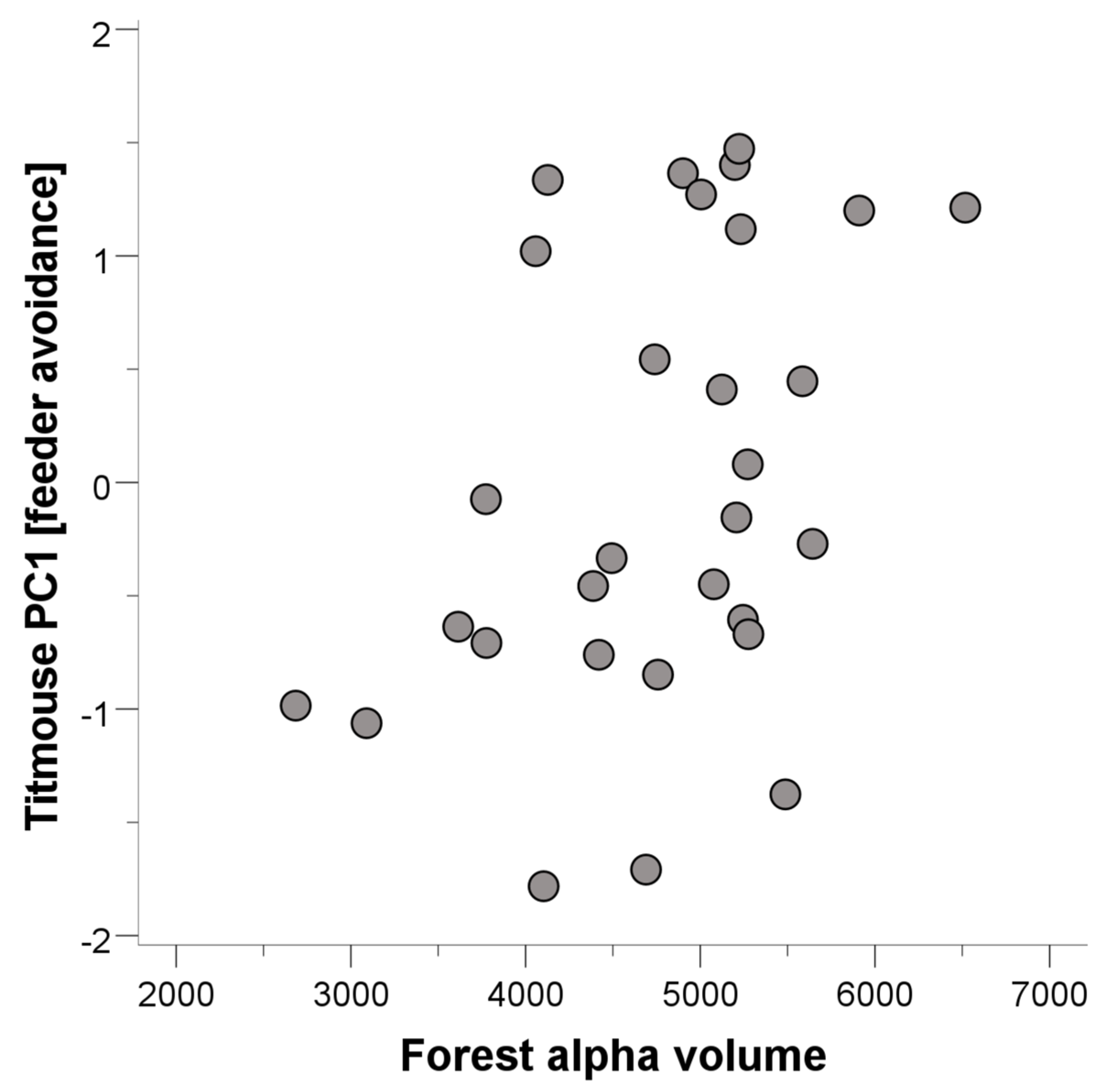
3.1. Tufted Titmice
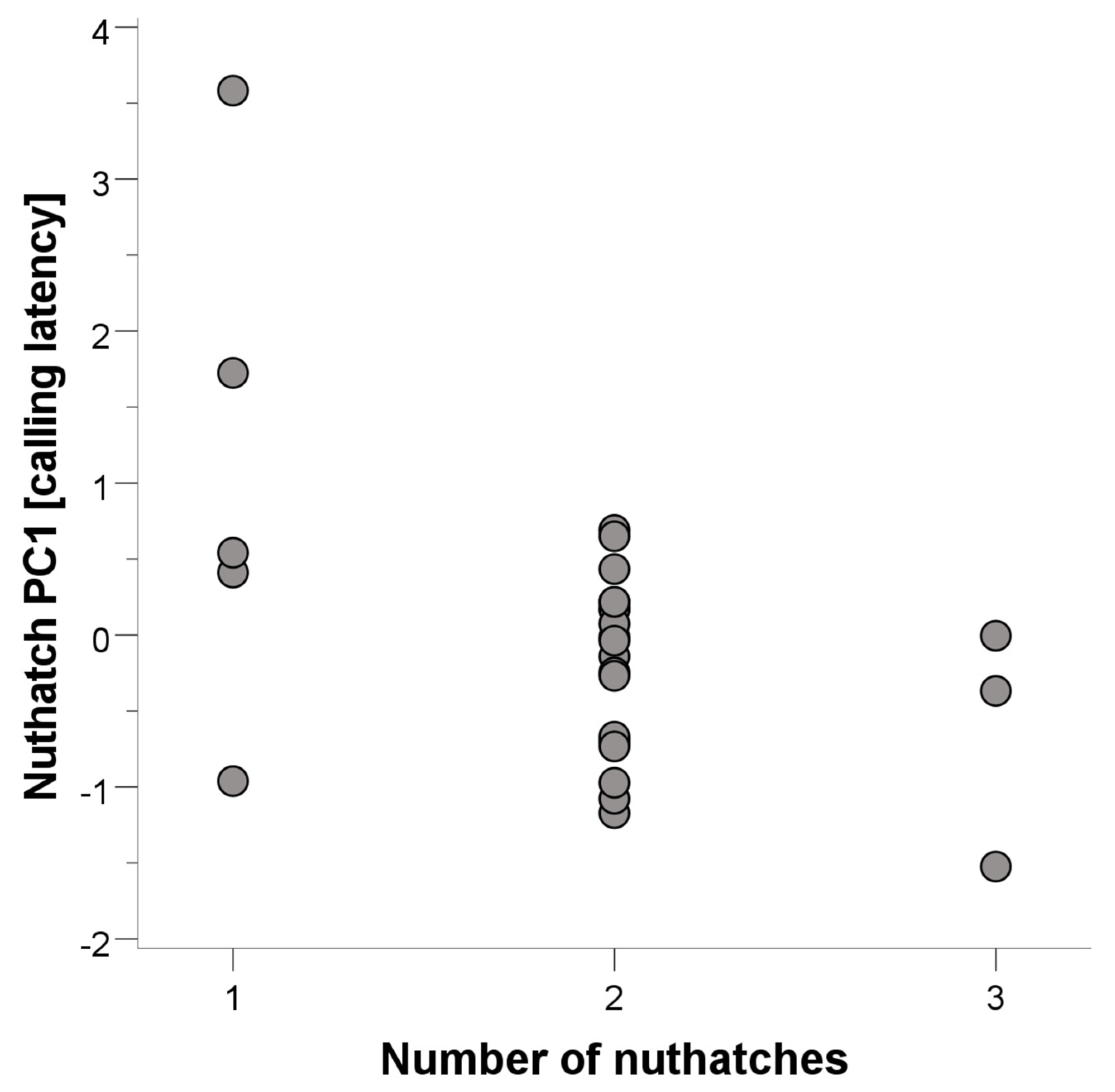
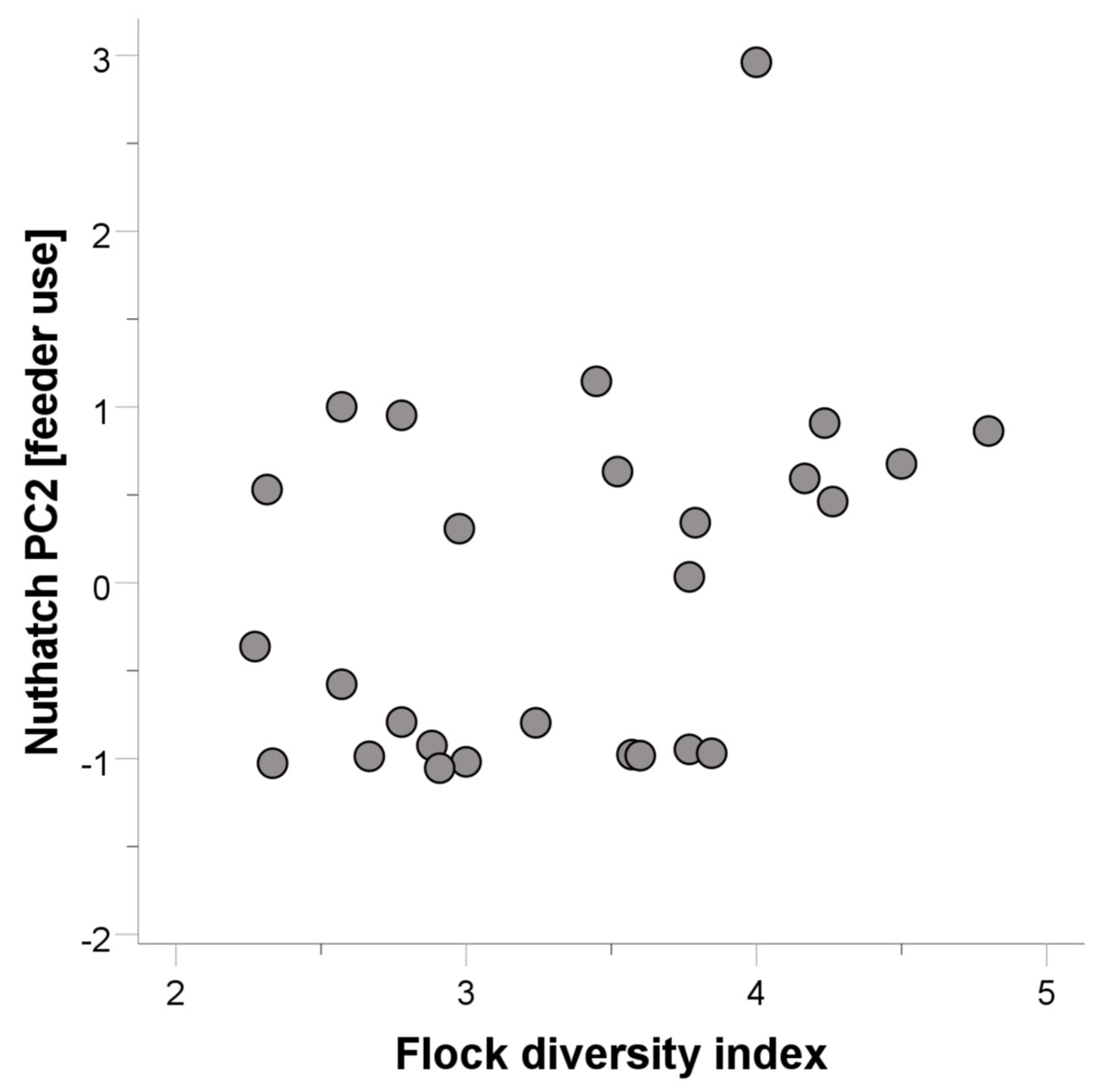
3.2. White-Breasted Nuthatches
3.3. Carolina Chickadees
4. Discussion
4.1. Social Variables
4.2. Physical Environmental Variables
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Adams, C.B.; Papeş, M.; Price, C.A.; Freeberg, T.M. Influence of social and physical environmental variation on antipredator behavior in mixed-species parid flocks. PLoS ONE 2023, 18, e0295910. [Google Scholar] [CrossRef] [PubMed]
- Carlson, N.V.; Freeberg, T.M.; Goodale, E.; Theo, A.H. Mixed-species groups and aggregations: Shaping ecological and behavioural patterns and processes. Philos. Trans. R. Soc. Lond. B 2023, 378, 20220093. [Google Scholar] [CrossRef]
- Coppinger, B.A.; Carlson, N.V.; Freeberg, T.M.; Sieving, K.E. Mixed-species groups and the question of dominance in the social ecosystem. Philos. Trans. R. Soc. Lond. B 2023, 378, 20220097. [Google Scholar] [CrossRef] [PubMed]
- Goodale, E.; Sridhar, H.; Sieving, K.E.; Bangal, P.; Colorado, G.J.; Farine, D.R.; Heymann, E.W.; Jones, H.H.; Krams, I.; Martínez, A.E.; et al. Mixed company: A framework for understanding the composition and organization of mixed-species animal groups. Biol. Rev. 2020, 95, 889–910. [Google Scholar] [CrossRef] [PubMed]
- Sridhar, H.; Shanker, K. Using intra-flock association patterns to understand why birds participate in mixed-species foraing flocks in terrestrial habitats. Behav. Ecol. Sociobiol. 2014, 68, 185–196. [Google Scholar] [CrossRef]
- Freeberg, T.M.; Adams, C.B.; Price, C.A.; Papeş, M. Mixed-species flock sizes and compositions influence flock members’ success in three field experiments with novel feeders. PLoS ONE 2024, 19, e0301270. [Google Scholar] [CrossRef]
- Freeberg, T.M.; Eppert, S.K.; Sieving, K.E.; Lucas, J.R. Diversity in mixed species groups improves success in a novel feeder test in a wild songbird community. Sci. Rep. 2017, 7, 43014. [Google Scholar] [CrossRef]
- Page, S.E. The Diversity Bonus: How Great Teams Pay Off in the Knowledge Economy; Princeton University Press: Princeton, NJ, USA, 2017. [Google Scholar]
- Barber, J.R.; Crooks, K.R.; Fristrup, K.M. The costs of chronic noise exposure for terrestrial organisms. Trends Ecol. Evol. 2010, 25, 180–189. [Google Scholar] [CrossRef]
- Chan, A.A.Y.-H.; Giraldo-Perez, P.; Smith, S.; Blumstein, D.T. Anthropogenic noise affects risk assessment and attention: The distracted prey hypothesis. Biol. Lett. 2010, 6, 458–461. [Google Scholar] [CrossRef]
- Jung, H.; Sherrod, A.; LeBreux, S.; Price, J.M.; Freeberg, T.M. Traffic noise and responses to a simulated approaching avian predator in mixed-species flocks of chickadees, titmice, and nuthatches. Ethology 2020, 126, 620–629. [Google Scholar] [CrossRef]
- Senzaki, M.; Yamaura, Y.; Francis, C.D.; Nakamura, F. Traffic noise reduces foraging efficiency in wild owls. Sci. Rep. 2016, 6, 30602. [Google Scholar] [CrossRef]
- Richardson, K.E.; Roche, D.P.; Mugel, S.G.; Lancaster, N.D.; Sieving, K.E.; Freeberg, T.M.; Lucas, J.R. Social dynamics of core members in mixed-species bird flocks change across a gradient of foraging habitat quality. PLoS ONE 2022, 17, e0262385. [Google Scholar]
- He, P.; Maldonado-Chaparro, A.A.; Farine, D.R. The role of habitat configuration in shaping social structure: A gap in studies of animal social complexity. Behav. Ecol. Sociobiol. 2019, 73, 9. [Google Scholar] [CrossRef]
- Herbert-Read, J.E.; Kremer, L.; Bruintjes, R.; Radford, A.N.; Ioannou, C.C. Anthropogenic noise pollution from pile-driving disrupts the structure and dynamics of Fish Shoals. Proc. R. Soc. B Biol. Sci. 2017, 284, 20171627. [Google Scholar] [CrossRef] [PubMed]
- Ekman, J. Ecology of non-breeding social systems of Parus. Wilson Bull. 1989, 101, 263–288. [Google Scholar]
- Smith, S.M. The Black-Capped Chickadee: Behavioral Ecology and Natural History; Cornell University Press: Ithaca, NY, USA, 1991. [Google Scholar]
- Goodale, E.; Beauchamp, G.; Ruxton, G.D. Mixed-Species Groups of Animals: Behavior, Community Structure, and Conservation; Academic Press: London, UK, 2017. [Google Scholar]
- Moynihan, M. The organization and probable evolution of some mixed species flocks of neotropical birds. Smithson. Misc. Collect. 1962, 143, 7. [Google Scholar]
- Sridhar, H.; Beauchamp, G.; Shanker, K. Why do birds participate in mixed-species foraging flocks? A large-scale synthesis. Anim. Behav. 2009, 78, 337–347. [Google Scholar] [CrossRef]
- Krams, I.; Krama, T.; Freeberg, T.M.; Kullberg, C.; Lucas, J.R. Linking social complexity and vocal complexity: A parid perspective. Philos. Trans. R. Soc. London. Ser. B Biol. Sci. 2012, 367, 1879–1891. [Google Scholar] [CrossRef]
- Lucas, J.R.; Freeberg, T.M. “Information” and the chick-a-dee call: Communicating with a complex vocal system. In Ecology and Behavior of Chickadees and Titmice: An Integrated Approach; Otter, K., Ed.; Oxford University Press: Oxford, UK, 2007; pp. 199–213. [Google Scholar]
- Courter, J.R.; Ritchison, G. Alarm calls of tufted titmice convey information about predator size and threat. Behav. Ecol. 2010, 21, 936–942. [Google Scholar] [CrossRef]
- Soard, C.M.; Ritchison, G. ‘Chick-a-dee’ calls of Carolina chickadees convey information about degree of threat posed by avian predators. Anim. Behav. 2009, 78, 1447–1453. [Google Scholar] [CrossRef]
- Templeton, C.N.; Greene, E. Nuthatches eavesdrop on variations in heterospecific chickadee mobbing alarm calls. Proc. Natl. Acad. Sci. USA 2007, 104, 5479–5482. [Google Scholar] [CrossRef] [PubMed]
- Templeton, C.N.; Greene, E.; Davis, K. Allometry of alarm calls: Black-capped chickadees encode information about predator size. Science 2005, 308, 1934–1937. [Google Scholar] [CrossRef]
- Coppinger, B.A.; Davis, J.; Freeberg, T.M. Flockmate familiarity affects note composition of chick-a-dee calls. Acta Ethologica 2019, 22, 73–77. [Google Scholar] [CrossRef]
- Coppinger, B.A.; Sanchez de-Launay, A.; Freeberg, T.M. Carolina chickadee (Poecile carolinensis) calling behavior in response to threats and in flight: Flockmate familiarity matters. J. Comp. Psychol. 2018, 132, 16–23. [Google Scholar] [CrossRef] [PubMed]
- Freeberg, T.M. Social complexity can drive vocal complexity: Group size influences vocal information in Carolina chickadees. Psychol. Sci. 2006, 17, 557–561. [Google Scholar] [CrossRef] [PubMed]
- Matthysen, E. Behavioral and ecological correlates of territory quality in the Eurasian Nuthatch (Sitta europaea). Auk 1990, 107, 86–95. [Google Scholar]
- Mostrom, A.M.; Curry, R.L.; Lohr, B.S. Carolina chickadee: Poecile carolinensis. In Birds of the World; Poole, A.F., Gill, F.B., Eds.; Version 1.0.; Cornell Lab of Ornithology: Ithaca, NY, USA, 2002. [Google Scholar] [CrossRef]
- Bartmess-LeVasseur, J.; Branch, C.L.; Browning, S.A.; Owens, J.L.; Freeberg, T.M. Predator stimuli and calling behavior of Carolina chickadees (Poecile carolinensis), tufted titmice (Baeolophus bicolor), and white-breasted nuthatches (Sitta carolinensis). Behav. Ecol. Sociobiol. 2010, 64, 1187–1198. [Google Scholar] [CrossRef]
- Smith, D.W.; Linnartz, N.W. The southern hardwood region. In Regional Silviculture of the United States, 2nd ed.; Barrett, J.W., Ed.; John Wiley & Sons: New York, NY, USA, 1980; pp. 145–230. [Google Scholar]
- Owens, J.L.; Stec, C.L.; O’Hatnick, A. The effects of extended exposure to traffic noise on parid social and risk-taking behavior. Behav. Process. 2012, 91, 61–69. [Google Scholar] [CrossRef]
- de Gasperis, S.R.; De Zan, L.R.; Battisti, C.; Reichegger, I.; Carpaneto, G.M. Distribution and abundance of hole-nesting birds in Mediterranean forests: Impact of past management patterns on habitat preference. Ornis Fennica 2016, 93, 100–110. [Google Scholar] [CrossRef]
- Sieving, K.E.; Contreras, T.A.; Maute, K.L. Heterospecific facilitation of forest-boundary crossing by mobbing understory birds in North-Central Florida. Auk 2004, 121, 738–751. [Google Scholar] [CrossRef]
- Edelsbrunner, H.; Kirkpatrick, D.; Seidel, R. On the shape of a set of points in the plane. IEEE Trans. Inf. Theory 1983, 29, 551–559. [Google Scholar] [CrossRef]
- Kyle, S.C. Do Carolina chickadees (Poecile carolinensis) and tufted titmice (Baeolophus bicolor) use predator eyes in risk assessment? Anim. Cogn. 2021, 24, 533–540. [Google Scholar] [CrossRef]
- Kyle, S.C.; Freeberg, T.M. Do Carolina chickadees (Poecile carolinensis) and tufted titmice (Baeolophus bicolor) attend to the head or body orientation of a perched avian predator? J. Comp. Psychol. 2016, 130, 145–152. [Google Scholar] [CrossRef] [PubMed]
- Bateson, M.; Martin, P. Measuring Behaviour: An Introductory Guide, 4th ed.; Cambridge University Press: Cambridge, UK, 2021. [Google Scholar]
- Grubb, T.C., Jr.; Pravosudov, V.V. White-breasted Nuthatch (Sitta carolinensis). In Birds of the World; Poole, A.F., Ed.; Version 1.0.; Cornell Lab of Ornithology: Ithaca, NY, USA, 2020. [Google Scholar] [CrossRef]
- Burghardt, G.M.; Bartmess-LeVasseur, J.M.; Browning, S.A.; Morrison, K.E.; Stec, C.L.; Zachau, C.E.; Freeberg, T.M. Perspectives—Minimizing observer bias in behavioral studies: A review and recommendations. Ethology 2012, 118, 511–517. [Google Scholar] [CrossRef]
- Simpson, E.H. Measurement of diversity. Nature 1949, 163, 688. [Google Scholar] [CrossRef]
- Ritchison, G. Vocalizations of the white-breasted nuthatch. Wilson Bull. 1983, 95, 440–451. [Google Scholar]
- Greenberg, R.S. Birds of many feathers: The formation and structure of mixed species flocks of forest birds. In On the Move: How and Why Animals Travel in Groups; Boinski, S., Garber, P.A., Eds.; University of Chicago Press: Chicago, IL, USA, 2001; pp. 521–558. [Google Scholar]
- Nolen, M.T.; Lucas, J.R. Asymmetries in mobbing behaviour and correlated intensity during predator mobbing by nuthatches, chickadees and titmice. Anim. Behav. 2009, 77, 1137–1146. [Google Scholar] [CrossRef]
- James, W.R.; McClintock, J.B. Anti-predator responses of amphipods are more effective in the presence of conspecific chemical cues. Hydrobiologia 2017, 797, 277–288. [Google Scholar] [CrossRef]
- Yorzinski, J.L.; Patricelli, G.L. Birds adjust acoustic directionality to beam their antipredator calls to predators and conspecifics. Proc. R. Society. B Biol. Sci. 2010, 277, 923–932. [Google Scholar] [CrossRef]
- Mangini, G.G.; Gandoy, F.A.; Areta, J.I.; Blendinger, P.G. Benefits of foraging in mixed-species flocks depend on species role and foraging strategy. Ibis 2023, 165, 629–646. [Google Scholar] [CrossRef]
- Carere, C.; Maestripieri, D. Animal Personalities: Behavior, Physiology, and Evolution; University of Chicago Press: Chicago, IL, USA, 2013. [Google Scholar]
- Sih, A.; Bell, A.; Johnson, J.C. Behavioral syndromes: An ecological and evolutionary overview. Trends Ecol. Evol. 2004, 19, 372–378. [Google Scholar] [CrossRef] [PubMed]
- Sih, A.; Watters, J.V. The mix matters: Behavioural types and group dynamics in water striders. Behaviour 2005, 142, 1417–1431. [Google Scholar]
- Morse, D.H. Ecological aspects of some mixed-species foraging flocks of birds. Ecol. Monogr. 1970, 40, 119–168. [Google Scholar] [CrossRef]
- Sullivan, K.A. Information exploitation by downy woodpeckers in mixed-species flocks. Behaviour 1984, 91, 294–311. [Google Scholar] [CrossRef]
- Meaux, E.; He, C.; Zeng, X.; He, R.; Jiang, A.; Goodale, E. Audience effects in a group-living bird: How contact call rate is affected by vegetation and group size and composition. Ecol. Evol. 2023, 13, e9909. [Google Scholar] [CrossRef]
- Grime, J.P. Control of species density in herbaceous vegetation. J. Environ. Manag. 1973, 1, 151–167. [Google Scholar]
- Moi, D.A.; García-Rios, R.; Hong, Z.; Daquila, B.V.; Mormul, R.P. Intermediate disturbance hypothesis in ecology: A literature review. Ann. Zool. Fenn. 2020, 57, 67–78. [Google Scholar] [CrossRef]
- Grade, A.M.; Sieving, K.E. When the birds go unheard: Highway noise disrupts information transfer between bird species. Biol. Lett. 2016, 12, 20160113. [Google Scholar] [CrossRef]


{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Species | PC Score | Predictor Variable | ß | t | p |
|---|---|---|---|---|---|
| Carolina chickadees | 1 | Alpha volume | 0.232 | 1.164 | 0.255 |
| Traffic noise (dB) | 0.170 | 0.825 | 0.417 | ||
| N. chickadees | −0.213 | −0.814 | 0.423 | ||
| Flock size | 0.102 | 0.360 | 0.722 | ||
| Flock diversity index | −0.185 | −0.739 | 0.467 | ||
| Tufted titmice | 1 | Alpha volume | 0.432 | 2.368 | 0.025 |
| Traffic noise (dB) | 0.304 | 1.557 | 0.133 | ||
| N. titmice | −0.227 | −0.916 | 0.369 | ||
| Flock size | 0.132 | 0.458 | 0.651 | ||
| Flock diversity index | −0.180 | −0.759 | 0.455 | ||
| 2 | Alpha volume | 0.266 | 1.341 | 0.192 | |
| Traffic noise (dB) | 0.143 | 0.667 | 0.511 | ||
| N. titmice | 0.216 | 0.794 | 0.435 | ||
| Flock size | −0.013 | −0.042 | 0.967 | ||
| Flock diversity index | −0.109 | −0.419 | 0.679 | ||
| White-breasted nuthatches | 1 | Alpha volume | −0.176 | −0.909 | 0.374 |
| Traffic noise (dB) | 0.150 | 0.789 | 0.439 | ||
| N. nuthatches | −0.408 | −2.105 | 0.047 | ||
| Flock size | −0.030 | −0.143 | 0.888 | ||
| Flock diversity index | −0.305 | −1.517 | 0.144 | ||
| 2 | Alpha volume | −0.097 | −0.480 | 0.636 | |
| Traffic noise (dB) | −0.218 | −1.097 | 0.285 | ||
| N. nuthatches | −0.276 | −1.372 | 0.185 | ||
| Flock size | 0.151 | 0.696 | 0.494 | ||
| Flock diversity index | 0.425 | 2.036 | 0.055 | ||
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Frazier, E.K.; Selman, Z.A.; Price, C.A.; Papeş, M.; Freeberg, T.M. Mixed-Species Flock Diversity and Habitat Density Are Associated with Antipredator Behavior in Songbirds. Diversity 2025, 17, 363. https://doi.org/10.3390/d17050363
Frazier EK, Selman ZA, Price CA, Papeş M, Freeberg TM. Mixed-Species Flock Diversity and Habitat Density Are Associated with Antipredator Behavior in Songbirds. Diversity. 2025; 17(5):363. https://doi.org/10.3390/d17050363
Chicago/Turabian StyleFrazier, Eric K., Zaharia A. Selman, Charles A. Price, Monica Papeş, and Todd M. Freeberg. 2025. "Mixed-Species Flock Diversity and Habitat Density Are Associated with Antipredator Behavior in Songbirds" Diversity 17, no. 5: 363. https://doi.org/10.3390/d17050363
APA StyleFrazier, E. K., Selman, Z. A., Price, C. A., Papeş, M., & Freeberg, T. M. (2025). Mixed-Species Flock Diversity and Habitat Density Are Associated with Antipredator Behavior in Songbirds. Diversity, 17(5), 363. https://doi.org/10.3390/d17050363