
Relative Influence of Salinity in the Flow and Accumulation of Organic Carbon in Open-Water Karstic Mangroves

 ,
,
Abstract
1. Introduction
2. Materials and Methods
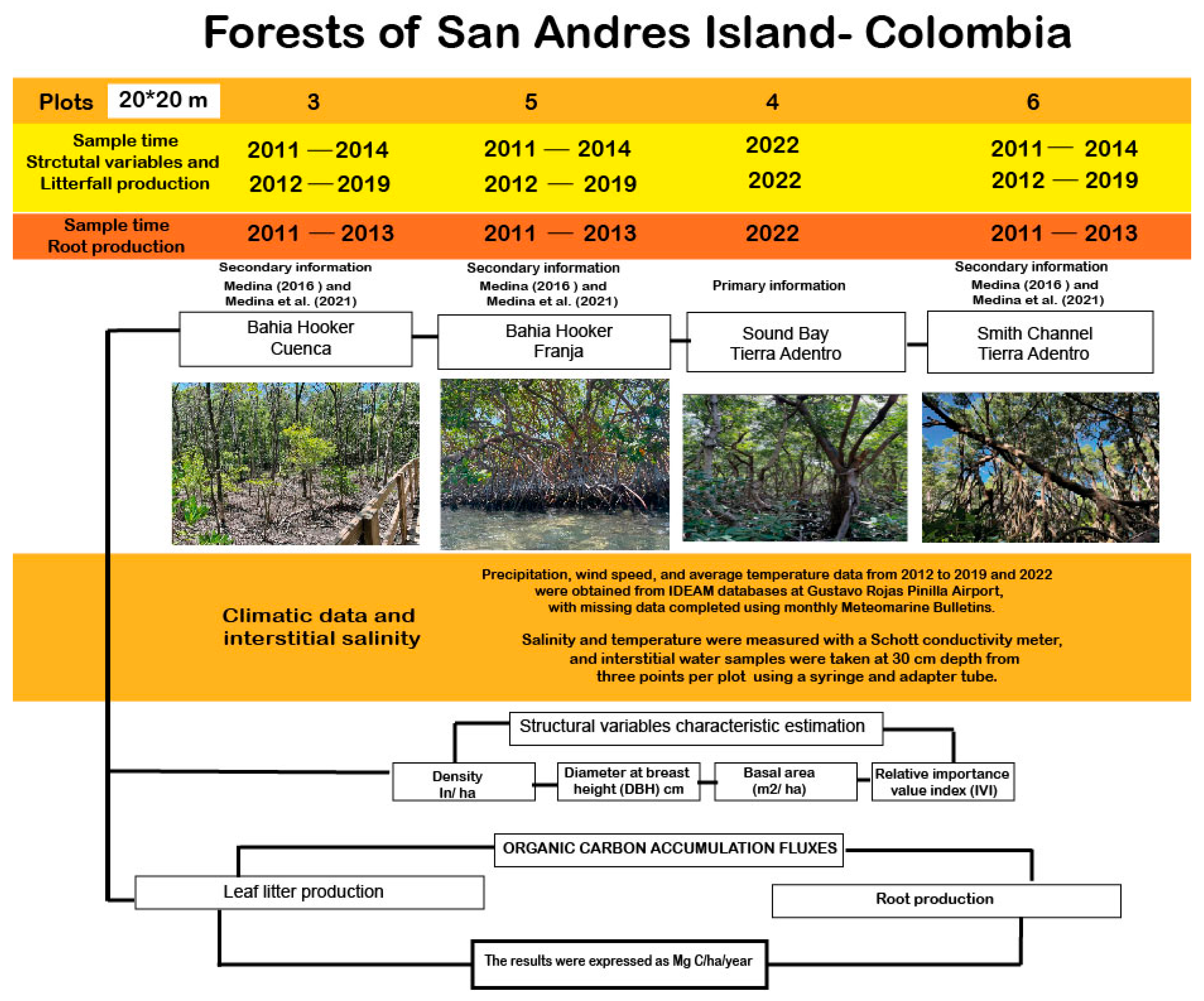
2.1. Study Area
2.2. Experiment Design
2.2.1. Forest Structure
2.2.2. Leaf Litter Production
2.2.3. Root Production
2.2.4. Climatic Data and Interstitial Salinity
2.3. Statistical Analysis
3. Results
3.1. Structure
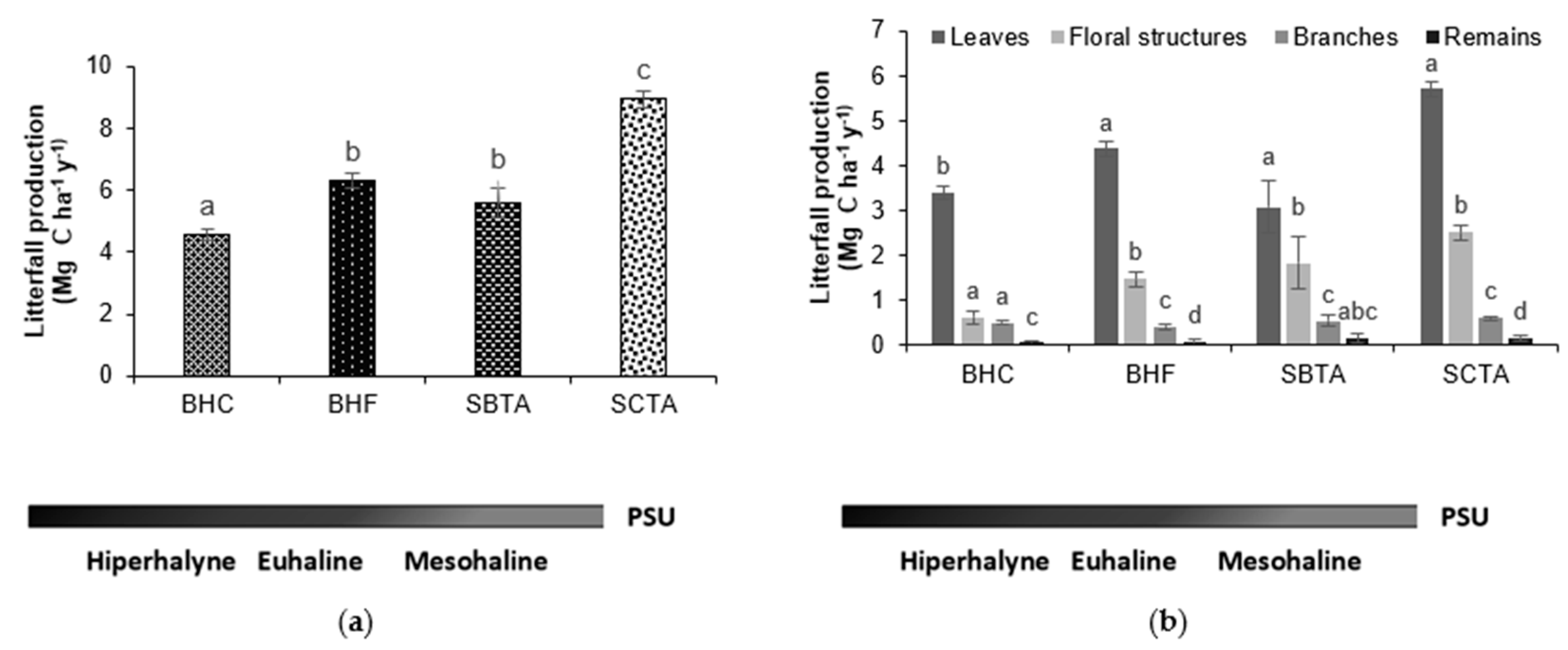
3.2. Carbon Content of Litterfall Production in Mangrove Forests with Different Salinity Gradients
3.3. Carbon Content in Multitemporal Litterfall Production
3.4. Carbon Content During Root Production in Forests with Different Salinity Gradients
3.5. Interstitial Salinity and Climatic Variables
3.6. Relationships of Climatic Variables and Interstitial Salinity to Litterfall Production
4. Discussion
4.1. Litter Production
4.1.1. Variation in Carbon Contents in Multitemporal and Multiannual Litterfall Production
4.1.2. Deltaic and Karstic Mangrove Analysis
4.2. Root Production
4.3. Potential Actions for the Conservation and Resilience of Mangrove Forests in the Face of Climate Change
5. Conclusions
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Bouillon, S. Storage beneath Mangroves. Nat. Geosci. 2011, 4, 282–283. [Google Scholar] [CrossRef]
- Duke, N.C.; Meynecke, J.-O.; Dittmann, S.; Ellison, A.M.; Anger, K.; Berger, U.; Cannicci, S.; Diele, K.; Ewel, K.C.; Field, C.D.; et al. A World Without Mangroves? Science 2007, 317, 41–42. [Google Scholar] [CrossRef] [PubMed]
- Palacios Peñaranda, M.L.; Cantera Kintz, J.R.; Peña Salamanca, E.J. Carbon Stocks in Mangrove Forests of the Colombian Pacific. Estuar. Coast. Shelf Sci. 2019, 227, 106299. [Google Scholar] [CrossRef]
- Kauffman, J.B.; Adame, M.F.; Arifanti, V.B.; Schile-Beers, L.M.; Bernardino, A.F.; Bhomia, R.K.; Donato, D.C.; Feller, I.C.; Ferreira, T.O.; Jesus Garcia, M.d.C.; et al. Total Ecosystem Carbon Stocks of Mangroves across Broad Global Environmental and Physical Gradients. Ecol. Monogr. 2020, 90, e01405. [Google Scholar] [CrossRef]
- Taillardat, P.; Friess, D.A.; Lupascu, M. Mangrove Blue Carbon Strategies for Climate Change Mitigation Are Most Effective at the National Scale. Biol. Lett. 2018, 14, 20180251. [Google Scholar] [CrossRef]
- Rodríguez-Rodríguez, J.A.; Mancera Pineda, J.E.; Melgarejo, L.M.; Medina Calderón, J.H. Functional Traits of Leaves and Forest Structure of Neotropical Mangroves under Different Salinity and Nitrogen Regimes. Flora Morphol. Distrib. Funct. Ecol. Plants 2018, 239, 52–61. [Google Scholar] [CrossRef]
- Herrera Silveira, J.A.; Camacho Rico, A.; Pech, E.; Pech, M.; Ramírez Ramírez, J.; Teutli Hernández, C. Dinámica Del Carbono (Almacenes y Flujos) En Manglares De. Terra Latinoamamericana 2016, 34, 61–72. [Google Scholar]
- Sitoe, A.; Mandlate, L.; Guedes, B. Biomass and Carbon Stocks of Sofala Bay Mangrove Forests. Forests 2014, 5, 1967–1981. [Google Scholar] [CrossRef]
- Agraz-Hernández, C.M.; Chan-Keb, C.A.; Chávez-Barrera, J.; Osti-Sáenz, J.; Expósito-Díaz, G.; Alonso-Campos, V.A.; Muñiz-Salazar, R.; Ruiz-Fernández, A.C.; Pérez-Bernal, L.H.; Sánchez-Cabeza, J.A.; et al. Carbon Stocks in a Mangrove Ecosystem in Northern Mexico: Environmental Changes for 35 Years. Rev. Mex. Biodivers. 2020, 91, GB2013. [Google Scholar] [CrossRef]
- Bouillon, S.; Borges, A.V.; Castañeda-Moya, E.; Diele, K.; Dittmar, T.; Duke, N.C.; Kristensen, E.; Lee, S.Y.; Marchand, C.; Middelburg, J.J.; et al. Mangrove Production and Carbon Sinks: A Revision of Global Budget Estimates. Global Biogeochem. Cycles 2008, 22, e912910. [Google Scholar] [CrossRef]
- Rovai, A.S.; Twilley, R.R.; Worthington, T.A.; Riul, P. Brazilian Mangroves: Blue Carbon Hotspots of National and Global Relevance to Natural Climate Solutions. Front. For. Global Change 2022, 4, 787533. [Google Scholar] [CrossRef]
- Twilley, R.R.; Castañeda-Moya, E.; Rovai, A. Productivity and Carbon Dynamics in Mangrove Wetlands. In Mangrove Ecosystems: A Global Biogeographic Perspective; Rivera-Monroy, V., Lee, S., Kristensen, E., Twilley, R., Eds.; Springer: Cham, Switherland, 2017. [Google Scholar]
- Medina-Calderón, J.H.; Mancera-Pineda, J.E.; Castañeda-Moya, E.; Rivera-Monroy, V.H. Hydroperiod and Salinity Interactions Control Mangrove Root Dynamics in a Karstic Oceanic Island in the Caribbean Sea (San Andres, Colombia). Front. Mar. Sci. 2021, 7, 598132. [Google Scholar] [CrossRef]
- Twilley, R.R.; Rovai, A.S.; Riul, P. Coastal Morphology Explains Global Blue Carbon Distributions. Front. Ecol. Environ. 2018, 16, 503–508. [Google Scholar] [CrossRef]
- Mckee, K.L.; Cahoon, D.R.; Feller, I.C. Caribbean Mangroves Adjust to Rising Sea Level through Biotic Controls on Change in Soil Elevation. Global Ecol. Biogeogr. 2007, 16, 545–556. [Google Scholar] [CrossRef]
- Day, J.W.; Coronado-Molina, C.; Vera-Herrera, F.R.; Twilley, R.; Rivera-Monroy, V.H.; Alvarez-Guillen, H.; Day, R.; Conner, W. Aquatic Botany a 7 Year Record of Above-Ground Net Primary Production in a Southeastern Mexican Mangrove Forest. Aquat. Bot. 1996, 55, 39–60. [Google Scholar] [CrossRef]
- Torres-Fernández del Campo, J.; Olvera-Vargas, M.; Figueroa-Rangel, B.L.; Cuevas-Guzmán, R.; Iñiguez-Dávalos, L.I. Patterns of Spatial Diversity and Structure of Mangrove Vegetation in Pacific West-Central Mexico. Wetlands 2018, 38, 919–931. [Google Scholar] [CrossRef]
- Restrepo, J.D.; Kjerfve, B. The Pacific and Caribbean Rivers of Colombia: Water Discharge, Sediment Transport and Dissolved Loads. In Environmental Geochemistry in Tropical and Subtropical Environments; Springer: Berlin/Heidelberg, Germany, 2004; pp. 169–187. [Google Scholar]
- Riascos, J.M.; Blanco-Libreros, J.F. Pervasively High Mangrove Productivity in a Major Tropical Delta throughout an ENSO Cycle (Southern Caribbean, Colombia). Estuar. Coast. Shelf Sci. 2019, 227, 106301. [Google Scholar] [CrossRef]
- Pérez-Ceballos, R.; Rivera-Rosales, K.; Zaldívar-Jiménez, A.; Canales-delgadillo, J.; Brito-Pérez, R.; Del Ángel, L.A.; Merino-Ibarra, M. Efecto de La Restauración Hidrológica Sobre La Productividad de Raíces Subterráneas En Los Manglares de Laguna de Términos, México. Bot. Sci. 2018, 96, 569–581. [Google Scholar] [CrossRef]
- Arnaud, M.; Morris, P.J.; Baird, A.J.; Dang, H.; Nguyen, T.T. Fine Root Production in a Chronosequence of Mature Reforested Mangroves. New Phytol. 2021, 232, 1591–1602. [Google Scholar] [CrossRef]
- Muhammad-Nor, S.M.; Huxham, M.; Salmon, Y.; Duddy, S.J.; Mazars-Simon, A.; Mencuccini, M.; Meir, P.; Jackson, G. Exceptionally High Mangrove Root Production Rates in the Kelantan Delta, Malaysia; An Experimental and Comparative Study. For. Ecol. Manag. 2019, 444, 214–224. [Google Scholar] [CrossRef]
- Castañeda-Moya, E.; Twilley, R.R.; Rivera-Monroy, V.H. Allocation of Biomass and Net Primary Productivity of Mangrove Forests along Environmental Gradients in the Florida Coastal Everglades, USA. For. Ecol. Manag. 2013, 307, 226–241. [Google Scholar] [CrossRef]
- Castañeda-Moya, E.; Twilley, R.R.; Rivera-Monroy, V.H.; Marx, B.D.; Coronado-Molina, C.; Ewe, S.M.L. Patterns of Root Dynamics in Mangrove Forests Along Environmental Gradients in the Florida Coastal Everglades, USA. Ecosystems 2011, 14, 1178–1195. [Google Scholar] [CrossRef]
- Medina Calderon, J.H. Estructura, Biomasa y Producción Primaria Neta. Ph.D. Thesis, Universidad Nacional de Colombia, Santa Marta, Colombia, 2016. [Google Scholar]
- Gavio, B.; Palmer-Cantillo, S.; Mancera, J.E. Historical Analysis (2000–2005) of the Coastal Water Quality in San Andrés Island, SeaFlower Biosphere Reserve, Caribbean Colombia. Mar. Pollut. Bull. 2010, 60, 1018–1030. [Google Scholar] [CrossRef]
- Vargas, G. Geología y Aspectos Geográficos de La Isla de San Andrés, Colombia. Geol. Colomb. 2004, 29, 71–87. [Google Scholar]
- CORALINA. INVEMAR Atlas de La Reserva de Biósfera Seaflower. In Archipiélago de San, Providencia y Santa Catalina. Instituto de Investigaciones y Costeras “José Benito Vives De Andréis”—INVEMAR—Y Para El Desarrollo Sostenible Del Archipiélago de San, Providencia y Santa Catalina—CORALINA; Gómez López, D.I., Segura Quintero, C., Sierra Correa, P.C., Garay Tinoco, J., Eds.; Serie de Publicaciones Especiales de INVEMAR # 28; CORALINA: Santa Marta, Colombia, 2012. [Google Scholar]
- Mckee, K.L.; Mendelssohn, I.A.; Hester, M.W. Reexamination of Pore Water Sulfide Concentrations and Redox Potentials Near the Aerial Roots of Rhizophora mangle and Avicennia germinans. Am. J. Bot. 1988, 75, 1352–1359. [Google Scholar] [CrossRef]
- Carriker, M.R. Ecology of Estuarine Benthic Invertebrates: A Perspective. Estuaries 1967, 83, 442–487. [Google Scholar]
- Howard, J.; Hoyt, S.; Isensee, K.; Telszewski, M.; Pidgeon, E. Coastal Blue Carbon Methods for Assessing Carbon Stocks and Emissions Factors in Mangroves, Tidal Salt Marshes, and Seagrass Meadows. 2014. Available online: https://unesdoc.unesco.org/ark:/48223/pf0000372868 (accessed on 16 December 2024).
- Feller, I.C.; Lovelock, C.E.; Berger, U.; McKee, K.L.; Joye, S.B.; Ball, M.C. Biocomplexity in Mangrove Ecosystems. Ann. Rev. Mar. Sci. 2010, 2, 395–417. [Google Scholar] [CrossRef]
- R Core Team R: A Language and Environment for Statistical Computing; R Foundation for Statistical Computing: Vienna, Austria, 2021.
- Ball, M.C. Comparative Ecophysiology of Mangrove Forest and Tropical Lowland Moist Rainforest. In Tropical Forest Plant Ecophysiology; Springer: Boston, MA, USA, 1996; pp. 461–496. [Google Scholar]
- Adame, M.F.; Fry, B. Source and Stability of Soil Carbon in Mangrove and Freshwater Wetlands of the Mexican Pacific Coast. Wetl. Ecol. Manag. 2016, 24, 129–137. [Google Scholar] [CrossRef]
- Coronado-Molina, C.; Alvarez-Guillen, H.; Day, J.W.; Reyes, E.; Perez, B.C.; Vera-Herrera, F.; Twilley, R. Litterfall Dynamics in Carbonate and Deltaic Mangrove Ecosystems in the Gulf of Mexico. Wetl. Ecol. Manag. 2012, 20, 123–136. [Google Scholar] [CrossRef]
- Ribeiro, R.d.A.; Rovai, A.S.; Twilley, R.R.; Castañeda-Moya, E. Spatial Variability of Mangrove Primary Productivity in the Neotropics. Ecosphere 2019, 10, e02841. [Google Scholar] [CrossRef]
- Lopes, D.M.S.; Tognella, M.M.P.; Falqueto, A.R.; Soares, M.L.G. Salinity Variation Effects on Photosynthetic Responses of the Mangrove Species Rhizophora mangle L. Growing in Natural Habitats. Photosynthetica 2019, 57, 1142–1155. [Google Scholar] [CrossRef]
- Agraz Hernández, C.M.; García Zaragoza, C.; Iriarte-Vivar, S.; Flores-Verdugo, F.J.; Moreno Casasola, P. Forest Structure, Productivity and Species Phenology of Mangroves in the La Mancha Lagoon in the Atlantic Coast of Mexico. Wetl. Ecol. Manag. 2011, 19, 273–293. [Google Scholar] [CrossRef]
- Torres, J.R.; Sánchez-Mejía, Z.M.; Arreola-Lizárraga, J.A.; Galindo-Félix, J.I.; Mascareño-Grijalva, J.J.; Rodríguez-Pérez, G. Environmental Factors Controlling Structure, Litter Productivity, and Phenology of Mangroves in Arid Region of the Gulf of California. Acta Oecol. 2022, 117, 103861. [Google Scholar] [CrossRef]
- Bompy, F.; Lequeue, G.; Imbert, D.; Dulormne, M. Increasing Fluctuations of Soil Salinity Affect Seedling Growth Performances and Physiology in Three Neotropical Mangrove Species. Plant Soil. 2014, 380, 399–413. [Google Scholar] [CrossRef]
- Jithesh, M.N.; Prashanth, S.R.; Sivaprakash, K.R.; Parida, A. Monitoring Expression Profiles of Antioxidant Genes to Salinity, Iron, Oxidative, Light and Hyperosmotic Stresses in the Highly Salt Tolerant Grey Mangrove, Avicennia marina (Forsk.) Vierh. by MRNA Analysis. Plant Cell Rep. 2006, 25, 865–876. [Google Scholar] [CrossRef]
- Takemura, T.; Hanagata, N.; Sugihara, K.; Baba, S.; Karube, I.; Dubinsky, Z. Physiological and Biochemical Responses to Salt Stress in the Mangrove, Bruguiera gymnorrhiza. Aquat. Bot. 2000, 68, 15–28. [Google Scholar] [CrossRef]
- Mitsch, W.J.; Gosselink, J.G. The Value of Wetlands: Importance of Scale and Landscape Setting. Ecol. Econ. 2000, 35, 25–33. [Google Scholar] [CrossRef]
- Naidoo, G.; Tuffers, A.V.; von Willert, D.J. Changes in Gas Exchange and Chlorophyll Fluorescence Characteristics of Two Mangroves and a Mangrove Associate in Response to Salinity in the Natural Environment. Trees 2002, 16, 140–146. [Google Scholar] [CrossRef]
- Goussi, R.; Manaa, A.; Derbali, W.; Cantamessa, S.; Abdelly, C.; Barbato, R. Comparative Analysis of Salt Stress, Duration and Intensity, on the Chloroplast Ultrastructure and Photosynthetic Apparatus in Thellungiella salsuginea. J. Photochem. Photobiol. B 2018, 183, 275–287. [Google Scholar] [CrossRef]
- Torres, J.R.; Barba, E.; Choix, F.J. Production and Biomass of Mangrove Roots in Relation to Hydroperiod and Physico-Chemical Properties of Sediment and Water in the Mecoacan Lagoon, Gulf of Mexico. Wetl. Ecol. Manag. 2019, 27, 427–442. [Google Scholar] [CrossRef]
- Parida, A.K.; Das, A.B.; Sanada, Y.; Mohanty, P. Effects of Salinity on Biochemical Components of the Mangrove, Aegiceras corniculatum. Aquat. Bot. 2004, 80, 77–87. [Google Scholar] [CrossRef]
- Zlatev, Z.S.; Yordanov, I.T. Effects of Soil Drought on Photosynthesis and Chlorophyll Fluorescence in Bean Plants. Bulg. J. Plant Physiol. 2004, 30, 3–18. [Google Scholar]
- Quadros, A.F.; Nordhaus, I.; Reuter, H.; Zimmer, M. Modelling of Mangrove Annual Leaf Litterfall with Emphasis on the Role of Vegetation Structure. Estuar. Coast. Shelf Sci. 2019, 218, 292–299. [Google Scholar] [CrossRef]
- Moreno, E.; Guerrero, A.; Gutiérrez, M.D.C.; Ortiz, C.A.; Palma, D.J. Los Manglares de Tabasco, Una Reserva Natural de Carbono. Madera Bosques 2002, 8, 115–128. [Google Scholar] [CrossRef]
- Yáñez-Arancibia, A.; Twilley, R.R.; Lara-Domínguez, A.L. Los Ecosistemas de Manglar Frente al Cambio Climático Global. Madera Bosques 1998, 4, 3–19. [Google Scholar] [CrossRef]
- McKee, K.L. Seedling Recruitment Patterns in a Belizean Mangrove Forest: Effects of Establishment Ability and Physico-Chemical Factors. Oecologia 1995, 101, 448–460. [Google Scholar] [CrossRef]
- Sierra-Rozo, O.; Mancera-Pineda, J.E.; Santos-Martínez, A. Velocidad de Descomposición de la Hojarasca en Diferentes Sustratos de Manglar, Durante la Época de Lluvias en San Andrés Isla, Caribe Colombiano. Bol. Investig. Mar. Cost. 2009, 1, 59–84. [Google Scholar] [CrossRef]
- Núñez-Ravelo, F.; Ugas-Pérez, M.; Calderón-Castellanos, R.; Rivas-Meriño, F. Cuantificación Del Carbono Orgánico y Materia Orgánica En Suelos No Rizosféricos o Cubiertos Por Avicennia germinans (L.) y Conocarpus erectus (L.) Emplazados En Boca de Uchire, Laguna de Unare, Estado de Anzoátegui, Venezuela. Rev. Geogr. Am. Cent. 2021, 1, 371–398. [Google Scholar] [CrossRef]
- García-Hansen, I.; Gaviria-Chiquazuque, J.F.; Prada-Triana, M.C.; Alvarez-León, R. Producción de Hojarasca de Los Manglares de La Isla de San Andrés, Caribe Colombiano. Rev. Biol. Trop. 2002, 50, 273–291. [Google Scholar]
- Pezeshki, S.R.; DeLaune, R.D.; Meeder, J.F. Carbon Assimilation and Biomass Partitioning in Avicennia germinans and Rhizophora mangle Seedlings in Response to Soil Redox Conditions. Environ. Exp. Bot. 1997, 37, 161–171. [Google Scholar] [CrossRef]
- Krauss, K.W.; Twilley, R.R.; Doyle, T.W.; Gardiner, E.S. Leaf Gas Exchange Characteristics of Three Neotropical Mangrove Species in Response to Varying Hydroperiod. Tree Physiol. 2006, 26, 959–968. [Google Scholar] [CrossRef] [PubMed]
- Sherman, R.E.; Fahey, T.J.; Martinez, P. Spatial Patterns of Biomass and Aboveground Net Primary Productivity in a Mangrove Ecosystem in the Dominican Republic. Ecosystems 2003, 6, 384–398. [Google Scholar] [CrossRef]
- Chen, G.C.; Tam, N.F.Y.; Ye, Y. Summer Fluxes of Atmospheric Greenhouse Gases N2O, CH4 and CO2 from Mangrove Soil in South China. Sci. Total Environ. 2010, 408, 2761–2767. [Google Scholar] [CrossRef]
- Agraz Hernández, C.M.; Chan Keb, C.A.; Iriarte-Vivar, S.; Posada Venegas, G.; Serratos, B.V.; Osti Sáenz, J. Phenological Variation of Rhizophora Mangle and Ground Water Chemistry Associated to Changes of the Precipitation Variación Fenológica de Rhizophora mangle y Química Del Agua Intersticial Asociada a Cambios de La Precipitación. Hidrobiológica 2015, 25, 49–61. [Google Scholar]
- Lovelock, C.E.; Feller, I.C. Photosynthetic Performance and Resource Utilization of Two Mangrove Species Coexisting in a Hypersaline Scrub Forest. Oecologia 2003, 134, 455–462. [Google Scholar] [CrossRef]
- Poveda, G.; Mesa, Ó.J. Las Fases Extremas Del Fenómeno ENSO (El Niño y La Niña) y Su Influencia Sobre La Hidrología de Colombia. Ing. Hidrául. Méx. 1996, 9, 21–37. [Google Scholar]
- Sánchez, A.R.; Pineda, J.E.M.; Casas, X.M.; Calderón, J.H.M. Influence of Edaphic Salinity on Leaf Morphoanatomical Functional Traits on Juvenile and Adult Trees of Red Mangrove (Rhizophora mangle): Implications with Relation to Climate Change. Forests 2021, 12, 1586. [Google Scholar] [CrossRef]
- Adame, M.F.; Kauffman, J.B.; Medina, I.; Gamboa, J.N.; Torres, O.; Caamal, J.P.; Reza, M.; Herrera-Silveira, J.A. Carbon Stocks of Tropical Coastal Wetlands within the Karstic Landscape of the Mexican Caribbean. PLoS ONE 2013, 8, e56569. [Google Scholar] [CrossRef]
- Torres, J.R.; Barba-Macías, E.; Sánchez, A.J. Tres Años de Producción de Hojarasca Del Manglar y Su Relación Con Las Condiciones Ambientales En La Laguna Mecoacán, Golfo de México. Ecosistemas 2023, 32, 2368. [Google Scholar] [CrossRef]
- Saravanakumar, A.; Mayalagu, R. Seasonal Variations in Physico-Chemical Characteristics of Water, Sediment and Soil Texture in Arid Zone Mangroves of Kachchh-Gujarat. J. Environ. Biol. 2008, 29, 725–732. [Google Scholar]
- Rovai, A.S.; Riul, P.; Twilley, R.R.; Castañeda-Moya, E.; Rivera-Monroy, V.H.; Williams, A.A.; Simard, M.; Cifuentes-Jara, M.; Lewis, R.R.; Crooks, S.; et al. Scaling Mangrove Aboveground Biomass from Site-level to Continental-scale. Global Ecol. Biogeogr. 2016, 25, 286–298. [Google Scholar] [CrossRef]
- Huxham, M.; Whitlock, D.; Githaiga, M.; Dencer-Brown, A. Carbon in the Coastal Seascape: How Interactions Between Mangrove Forests, Seagrass Meadows and Tidal Marshes Influence Carbon Storage. Curr. For. Rep. 2018, 4, 101–110. [Google Scholar] [CrossRef]
- Twilley, R.R. Properties of Mangrove Ecosystems Related to the Energy Signature of Coastal Environments. In Maximum Power: The Ideas and Applications of H. T. odum; Twilley Robert, R., Ed.; University Press of Colorado: Denver, CO, USA, 1995; pp. 43–62. [Google Scholar]
- Saintilan, N.; Khan, N.S.; Ashe, E.; Kelleway, J.J.; Rogers, K.; Woodroffe, C.D.; Horton, B.P. Thresholds of Mangrove Survival under Rapid Sea Level Rise. Science 2020, 368, 1118–1121. [Google Scholar] [CrossRef] [PubMed]
- Rovai, A.S.; Twilley, R.R.; Castañeda-Moya, E.; Riul, P.; Cifuentes-Jara, M.; Manrow-Villalobos, M.; Horta, P.A.; Simonassi, J.C.; Fonseca, A.L.; Pagliosa, P.R. Global Controls on Carbon Storage in Mangrove Soils. Nat. Clim. Change 2018, 8, 534–538. [Google Scholar] [CrossRef]
- Rajaniemi, T.K.; Allison, V.J. Abiotic Conditions and Plant Cover Differentially Affect Microbial Biomass and Community Composition on Dune Gradients. Soil. Biol. Biochem. 2009, 41, 102–109. [Google Scholar] [CrossRef]
- Adame, M.F.; Teutli, C.; Santini, N.S.; Caamal, J.P.; Zaldívar-Jiménez, A.; Herńndez, R.; Herrera-Silveira, J.A. Root Biomass and Production of Mangroves Surrounding a Karstic Oligotrophic Coastal Lagoon. Wetlands 2014, 34, 479–488. [Google Scholar] [CrossRef]
- Meera, S.P.; Bhattacharyya, M.; Kumar, A. Dynamics of Mangrove Functional Traits under Osmotic and Oxidative Stresses. Plant Growth Regul. 2023, 101, 285–306. [Google Scholar] [CrossRef]
- Nguyen, H.T.; Stanton, D.E.; Schmitz, N.; Farquhar, G.D.; Ball, M.C. Growth Responses of the Mangrove Avicennia marina to Salinity: Development and Function of Shoot Hydraulic Systems Require Saline Conditions. Ann. Bot. 2015, 115, 397–407. [Google Scholar] [CrossRef]
- Vovides, A.G.; Berger, U.; Grueters, U.; Guevara, R.; Pommerening, A.; Lara-Domínguez, A.L.; López-Portillo, J. Change in Drivers of Mangrove Crown Displacement along a Salinity Stress Gradient. Funct. Ecol. 2018, 32, 2753–2765. [Google Scholar] [CrossRef]
- Pi, N.; Tam, N.F.Y.; Wu, Y.; Wong, M.H. Root Anatomy and Spatial Pattern of Radial Oxygen Loss of Eight True Mangrove Species. Aquat. Bot. 2009, 90, 222–230. [Google Scholar] [CrossRef]
- Ola, A.; Gauthier, A.R.G.; Xiong, Y.; Lovelock, C.E. The Roots of Blue Carbon: Responses of Mangrove Stilt Roots to Variation in Soil Bulk Density. Biol. Lett. 2019, 15, 20180866. [Google Scholar] [CrossRef] [PubMed]
- López Rodríguez, A.; Cecilia, Á.; Correa, S.; Cristina, P.; Ortiz, H.; Guzmán, M.; Zapata, L. Ordenamiento Ambiental de Los Manglares Del Archipiélago San Andrés, Providencia y Santa Catalina Item Type Book/Monograph/Conference Proceedings. 2009. Available online: https://centrodocumentacion.invemar.org.co/cgi-bin/koha/opac-detail.pl?biblionumber=16502 (accessed on 16 December 2024).
- Ahmed, S.; Sarker, S.K.; Friess, D.A.; Kamruzzaman, M.; Jacobs, M.; Islam, M.A.; Alam, M.A.; Suvo, M.J.; Sani, M.N.H.; Dey, T.; et al. Salinity Reduces Site Quality and Mangrove Forest Functions. From Monitoring to Understanding. Sci. Total Environ. 2022, 853, 158662. [Google Scholar] [CrossRef] [PubMed]
- Arnaud, M.; Krause, S.; Norby, R.J.; Dang, T.H.; Acil, N.; Kettridge, N.; Gauci, V.; Ullah, S. Global Mangrove Root Production, Its Controls and Roles in the Blue Carbon Budget of Mangroves. Global Change Biol. 2023, 29, 3256–3270. [Google Scholar] [CrossRef]
- Krauss, K.W.; McKee, K.L.; Lovelock, C.E.; Cahoon, D.R.; Saintilan, N.; Reef, R.; Chen, L. How Mangrove Forests Adjust to Rising Sea Level. New Phytol. 2014, 202, 19–34. [Google Scholar] [CrossRef]
- Salmo, S.G.; Lovelock, C.; Duke, N.C. Vegetation and Soil Characteristics as Indicators of Restoration Trajectories in Restored Mangroves. Hydrobiologia 2013, 720, 1–18. [Google Scholar] [CrossRef]
- McKee, K.L.; Krauss, K.W.; Cahoon, D.R. Does Geomorphology Determine Vulnerability of Mangrove Coasts to Sea-Level Rise? In Dynamic Sedimentary Environments of Mangrove Coasts; Elsevier: Amsterdam, The Netherlands, 2021; pp. 255–272. [Google Scholar]
- Lang’at, J.K.S.; Kirui, B.K.Y.; Skov, M.W.; Kairo, J.G.; Mencuccini, M.; Huxham, M. Species Mixing Boosts Root Yield in Mangrove Trees. Oecologia 2013, 172, 271–278. [Google Scholar] [CrossRef]
- Hayes, M.A.; Jesse, A.; Tabet, B.; Reef, R.; Keuskamp, J.A.; Lovelock, C.E. The Contrasting Effects of Nutrient Enrichment on Growth, Biomass Allocation and Decomposition of Plant Tissue in Coastal Wetlands. Plant Soil 2017, 416, 193–204. [Google Scholar] [CrossRef]
- Cormier, N.; Twilley, R.R.; Ewel, K.C.; Krauss, K.W. Fine Root Productivity Varies along Nitrogen and Phosphorus Gradients in High-Rainfall Mangrove Forests of Micronesia. Hydrobiologia 2015, 750, 69–87. [Google Scholar] [CrossRef]
- Breithaupt, J.L.; Steinmuller, H.E. Refining the Global Estimate of Mangrove Carbon Burial Rates Using Sedimentary and Geomorphic Settings. Geophys. Res. Lett. 2022, 49, e2022GL100177. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Forest | Salinity (PSU) | Species | Density () | DBH (≥2.5 cm) | Basal Area () | IVI |
|---|---|---|---|---|---|---|
| BHC | 62.4 ± 10.4 | Ag | 2242 ± 32.4 | 9.1 ± 0.3 | 19.8 ± 0.5 | 221 |
| Lr | 125 ± 55.9 | 10.8 ± 1.1 | 1.3 ± 0.08 | 37 | ||
| Rm | 183 ± 67.7 | 7.1 ± 0.5 | 0.8 ± 0.04 | 42 | ||
| Total | 2550 ± 71.7 a | 9.1 ± 0.3 a | 21.9 ± 0.5 b | 300 | ||
| BHF | 37.5 ± 5.8 | Ag | 215 ± 0.5 | 8.8 ± 0.8 | 1.7 ± 0.2 | 26 |
| Lr | 920 ± 1.7 | 8.1 ± 0.3 | 6.3 ± 0.3 | 104 | ||
| Rm | 2520 ± 2.3 | 6.8 ± 0.1 | 10.9 ± 0.2 | 170 | ||
| Total | 3655 ± 43.7 ab | 7.3 ± 0.1a | 18.9 ± 0.4 b | 300 | ||
| SBTA | 11.5 ± 7.5 | Ag | 356 ± 0.5 | 14.8 ± 1.1 | 9.8 ± 1.2 | 133 |
| Lr | 188 ± 0.8 | 27.6 ± 2 | 13.2 ± 2 | 99 | ||
| Rm | 194 ± 1.1 | 13.04 ± 1.5 | 4.2 ± 0.8 | 68 | ||
| Total | 738 ± 8.8 b | 18.5 ± 0.73 b | 27.3 ± 0.4 b | 300 | ||
| SCTA | 9.6 ± 6.3 | Lr | 75 ± 13 | 34.1 ± 1.4 | 9.6 ± 2.6 | 61 |
| Rm | 313 ± 24 | 28.5 ± 0.7 | 28.2 ± 0.1 | 182 | ||
| Other spp. | 16 ± 9 | 7.2 ± 0.4 | 58 | |||
| Total | 404 ± 26 ab | 23.3 ± 8.2 b | 37.8 ± 2.5 b | 300 | ||
| Df | 3 | 3 | 3 | |||
| P | 0.011 | 0.006 | 0.057 | |||
| X2 | 11.192 | 12.4 | 7.526 | |||
| Size of Roots | ||||
|---|---|---|---|---|
| Forest | <2 mm | 2–5 mm | >5 mm | Total |
| BHC | 0.08 ± 0.06 | 0.09 ± 0.07 | 0.07 ± 0.10 | 0.24 ± 0.20 ab |
| BHF | 0.48 ± 0.30 | 0.28 ± 0.10 | 0.54 ± 0.34 | 1.30 ± 0.5 a |
| SBTA | 0.41 ± 0.08 | 0.24 ± 0.07 | 0.54 ± 0.36 | 1.19 ± 0.46 ab |
| SCTA | 0.16 ± 0.03 | 0.10 ± 0.02 | 0.16 ± 0.06 | 0.41 ± 0.08 b |
| P | 0.0008 | 0.00025 | 0.024 | 0.0052 |
| Df | 3 | 3 | 3 | 3 |
| X2 | 16.77 | 19.21 | 9.45 | 12.76 |
| Forest | PSU Annual | PSU by Period | |
|---|---|---|---|
| Wet | Dry | ||
| BHC | 62.3 ± 10.5 a | 60.7 ± 0.9 a | 62.5 ± 1.0 a |
| BHF | 37.4 ± 5.7 b | 36.6 ± 0.4 b | 39.0 ± 0.3 b |
| SBTA | 11.5 ± 7.4 c | 9.4 ± 0.6 c | 17.0 ± 0.8 d |
| SCTA | 9.6 ± 6.2 d | 9.5 ± 0.4 c | 9.8 ± 0.5 c |
| Production Forest | Ws Prom m/s | P Mm | T °C | |||
|---|---|---|---|---|---|---|
| R | p | R | p | R | p | |
| BHC | −0.73 | 0.039 * | 0.16 | 0.696 | 0.10 | 0.819 |
| BHF | 0.46 | 0.251 | 0.12 | 0.782 | −0.36 | 0.379 |
| SCTA | 0.38 | 0.349 | 0.43 | 0.292 | −0.23 | 0.580 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Quintero Alvarado, A.P.; Medina Calderón, J.H.; Mancera-Pineda, J.E. Relative Influence of Salinity in the Flow and Accumulation of Organic Carbon in Open-Water Karstic Mangroves. Diversity 2025, 17, 360. https://doi.org/10.3390/d17050360
Quintero Alvarado AP, Medina Calderón JH, Mancera-Pineda JE. Relative Influence of Salinity in the Flow and Accumulation of Organic Carbon in Open-Water Karstic Mangroves. Diversity. 2025; 17(5):360. https://doi.org/10.3390/d17050360
Chicago/Turabian StyleQuintero Alvarado, Angélica Paola, Jairo Humberto Medina Calderón, and José Ernesto Mancera-Pineda. 2025. "Relative Influence of Salinity in the Flow and Accumulation of Organic Carbon in Open-Water Karstic Mangroves" Diversity 17, no. 5: 360. https://doi.org/10.3390/d17050360
APA StyleQuintero Alvarado, A. P., Medina Calderón, J. H., & Mancera-Pineda, J. E. (2025). Relative Influence of Salinity in the Flow and Accumulation of Organic Carbon in Open-Water Karstic Mangroves. Diversity, 17(5), 360. https://doi.org/10.3390/d17050360