

Systematic Revision of the Genus Charmus Karsch, 1879 (Scorpiones: Buthidae), and Assessment of Its Phylogenetic Position Within Buthidae C. L. Koch, 1837 Using Ultraconserved Elements

 , ,
, ,
Abstract

1. Introduction
1.1. Systematics of the Genus Charmus
1.2. Phylogenetic Analysis of the Genus Charmus
2. Materials and Methods
2.1. Specimen Collection and Morphological Data
2.2. Repository
2.3. Taxon Sampling
2.4. Sanger Sequencing: DNA Extraction, Amplification and Sequencing
2.5. Sequence Alignment of the Sanger Sequenced Data
2.6. Genetic Divergence (p-Distance) of the Sanger Sequenced Data
2.7. Molecular Phylogenetic Analysis of the Sanger Sequenced Data
2.8. DNA Extractions and Shotgun Library Preparation
2.9. Hybridization Capture and Sequencing of UCEs
2.10. Data Processing and PHYLUCE Pipeline
2.11. Phylogenetic Analyses of the Generated UCE Matrices
3. Systematics
3.1. Charmus laneus Karsch, 1879

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| IES-631 (♂) | IES-633 (♂) | IES-628 (♀) | IES-629 (♀) | IES-632 | ||
|---|---|---|---|---|---|---|
| Carapace | L/W | 2.08/1.81 | 1.71/1.55 | 1.99/2.02 | 2.03/1.9 | 2.12/1.94 |
| Mesosoma | L | 4.48 | 3.62 | 4.6 | 5.35 | 2.47 |
| Tergite VII | L/W | 0.95/1.99 | 0.83/1.58 | 0.95/1.73 | 1.91/1.62 | 0.55/1.28 |
| Metasoma and telson | L | 8.55 | 7.83 | 10.47 | 8.84 | 6.09 |
| Segment I | L/W/D | 0.97/1.48/1.23 | 0.93/1.3/1.1 | 1.32/1.53/1.27 | 0.87/1.5/1.25 | 0.76/0.89/0.76 |
| Segment II | L/W/D | 1.21/1.4/1.2 | 0.99/1.22/1.15 | 1.61/1.57/1.42 | 1.05/1.52/1.26 | 0.86/0.87/0.80 |
| Segment III | L/W/D | 1.34/1.44/1.21 | 1.15/1.28/1.21 | 1.85/1.54/1.42 | 1.25/1.52/1.39 | 0.93/0.96/0.82 |
| Segment IV | L/W/D | 1.51/1.48/1.18 | 1.35/1.29/1.24 | 2.12/1.54/1.42 | 1.59/1.52/1.36 | 1.06/0.95/0.85 |
| Segment V | L/W/D | 1.63/1.46/1.12 | 1.7/1.31/1.16 | 2.45/1.6/1.33 | 2.25/1.5/1.26 | 1.34/1.99/0.81 |
| Telson | L/W/D | 1.14/0.8/0.9 | 0.96/0.77/0.89 | 0.63/0.50/0.99 | 0.91/0.8/0.95 | 0.62/0.46/0.54 |
| Pedipalp | L | 6.63 | 5.29 | 6.78 | 6.54 | 4.03 |
| Femur | L/W | 1.72/0.51 | 1.35/0.48 | 1.65/0.54 | 1.72/0.55 | 0.98/0.39 |
| Patella | L/W | 2.00/0.78 | 1.62/0.69 | 2.15/0.79 | 1.97/0.75 | 1.26/0.46 |
| Chela | L | 2.91 | 2.32 | 2.98 | 2.85 | 1.79 |
| Manus | W/D | 0.6/0.6 | 0.48/0.54 | 0.63/0.66 | 0.66/0.64 | 0.37/0.39 |
| Movable finger | L | 1.65 | 1.75 | 1.91 | 2.04 | 1.08 |
| Pectine | L/W | 1.72/0.43 | 1.60/0.39 | 1.74/0.47 | 1.71/0.42 | 0.99/0.26 |
| Genital Operculum | L/W | 0.43/0.62 | 0.28/0.47 | 0.38/0.72 | 0.42/0.68 | 0.22/0.36 |
| Total | L | 15.11 | 13.16 | 17.06 | 16.22 | 10.68 |
| Pectinal teeth count | PTC | 17/16 | 15/16 | 16/17 | 16/17 | 17/17 |

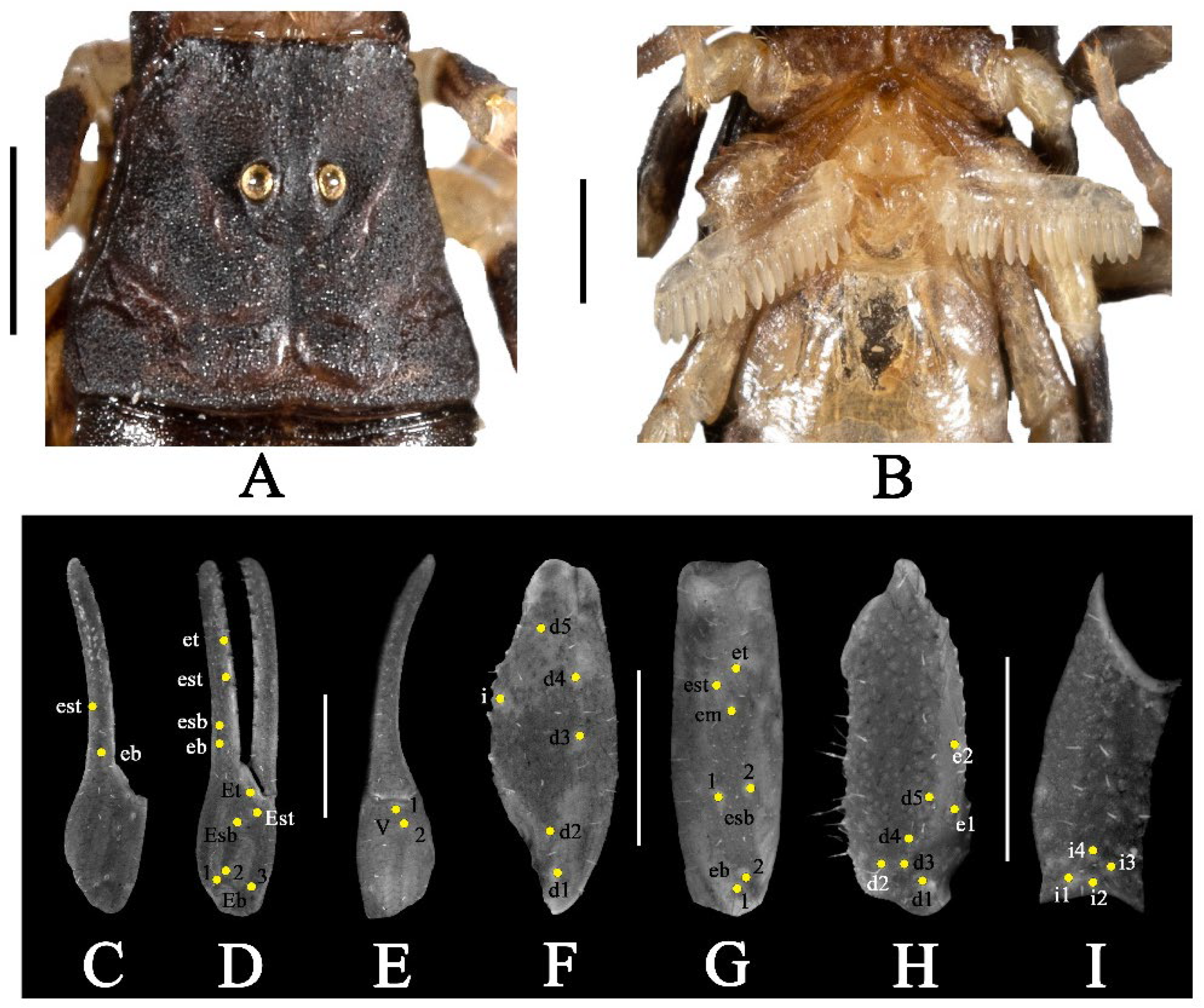

3.2. Charmus sinhagadensis Tikader and Bastawade, 1983
| BNHS SC 403 (♂) (Neotype) | IES-602(♂) | IES-587 (♂) | ||
|---|---|---|---|---|
| Carapace | L/W | 2.27/2.28 | 1.8/1.56 | 1.82/1.58 |
| Mesosoma | L | 4.78 | 3.32 | 3.23 |
| Tergite VII | L/W | 1.21/2.52 | 0.94/1.63 | 0.86/1.62 |
| Metasoma and telson | L | 10.62 | 8.53 | 8.48 |
| Segment I | L/W/D | 0.88/1.50/1.26 | 0.96/1.11/1.05 | 0.94/1.1/1.13 |
| Segment II | L/W/D | 1.29/1.31/1.27 | 1.12/1.11/1.05 | 1.15/1.1/1.14 |
| Segment III | L/W/D | 1.50/1.35/1.38 | 1.39/1.11/1.09 | 1.26/1.11/1.13 |
| Segment IV | L/W/D | 2.18/1.36/1.31 | 1.45/1.14/1.1 | 1.52/1.1/1.12 |
| Segment V | L/W/D | 2.56/1.36/1.26 | 1.87/1.15/1.08 | 1.78/1.1/1.07 |
| Telson | L/W/D | 1.24/0.84/1.05 | 0.93/0.81/0.73 | 1.17/0.76/0.93 |
| Pedipalp | L | 9.08 | 5.95 | 6.14 |
| Femur | L/W | 2.19/0.67 | 1.5/0.48 | 1.57/0.46 |
| Patella | L/W | 2.81/0.98 | 1.83/0.63 | 1.83/0.66 |
| Chela | L | 4.08 | 2.62 | 2.74 |
| Manus | W/D | 0.86/0.84 | 0.54/0.53 | 0.54/0.55 |
| Movable finger | L | 2.81 | 1.78 | 2.02 |
| Pectine | L/W | 2.04/0.52 | 1.54/0.37 | 1.6/0.36 |
| Genital Operculum | L/W | 0.43/0.96 | 0.35/0.56 | 0.37/0.55 |
| Total | L | 17.67 | 13.65 | 13.53 |
| Pectinal teeth count | PTC | 14/14 | 15/15 | 15/15 |

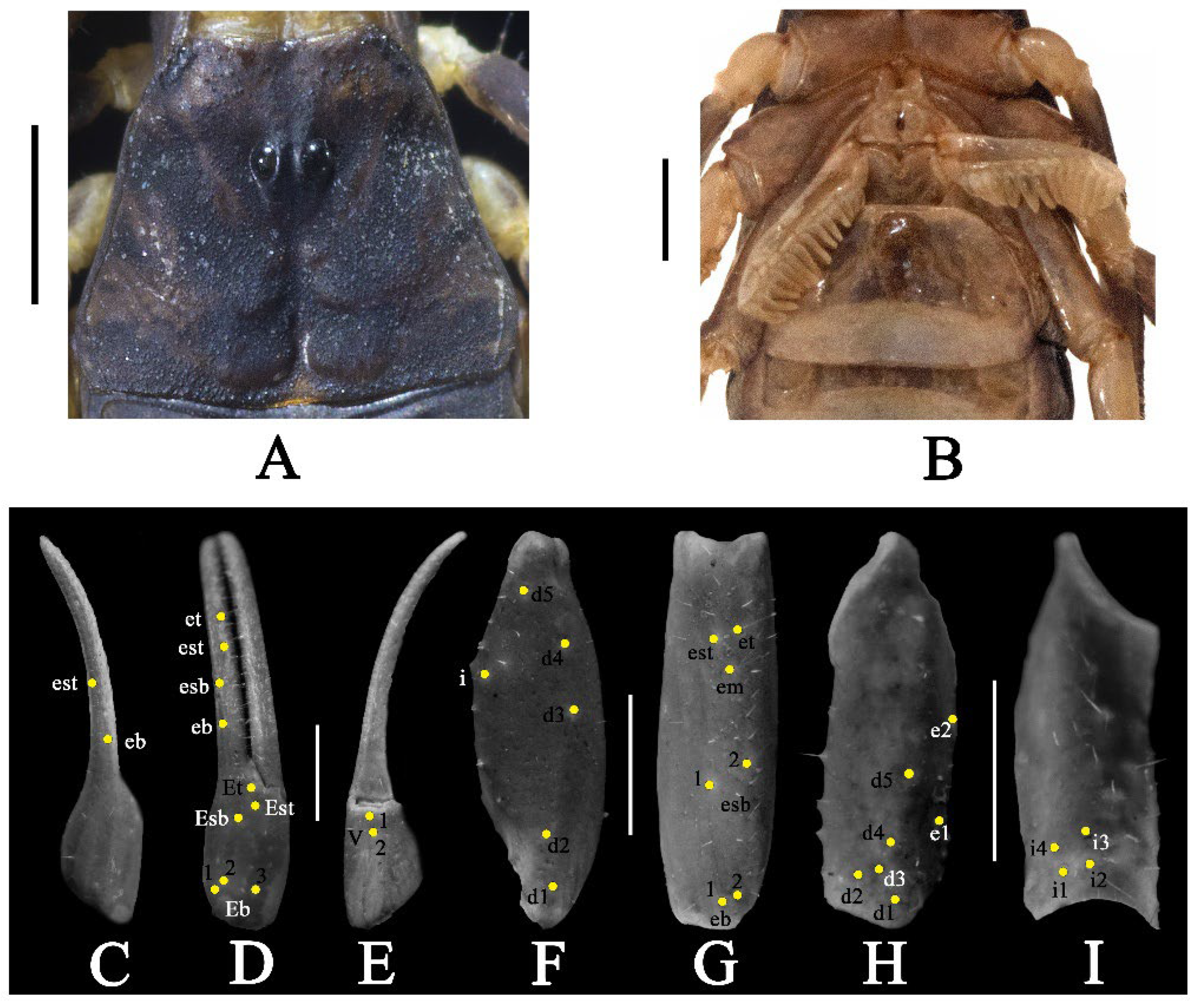

3.3. Charmus brignolii (Lourenço, 2000)
| IES-614 (♂) | IES-615 (♂) | IES-616 (♂) | IES-611 (♀) | IES-612 (♀) | ||
|---|---|---|---|---|---|---|
| Carapace | L/W | 1.96/1.73 | 2.09/1.83 | 2.08/1.87 | 1.98/1.89 | 2.08/1.80 |
| Mesosoma | L | 4.06 | 3.97 | 3.84 | 2.36 | 4.67 |
| Tergite VII | L/W | 0.92/2.04 | 0.86/2.04 | 0.83/2.26 | 0.55/1.24 | 1.16/2.33 |
| Metasoma and telson | L | 8.47 | 8.40 | 9.05 | 6.05 | 8.39 |
| Segment I | L/W/D | 0.79/1.32/0.96 | 0.82/1.41/1.1 | 0.85/1.53/1.04 | 1.09/1.4/1.19 | 0.81/1.39/1.06 |
| Segment II | L/W/D | 1.05/1.36/0.90 | 1.13/1.46/1.26 | 1.2/1.36/1.13 | 1.19/1.41/1.19 | 1.07/1.43/1.04 |
| Segment III | L/W/D | 1.19/1.42/1.05 | 1.33/1.54/1.32 | 1.43/1.48/1.2 | 1.83/1.41/1.18 | 1.26/1.45/1.27 |
| Segment IV | L/W/D | 1.52/1.42/1.20 | 1.5/1.51/1.24 | 1.57/1.62/1.24 | 1.94/1.41/1.17 | 1.44/1.57/1.33 |
| Segment V | L/W/D | 2.12/1.53/1.12 | 1.8/1.56/1.21 | 2.2/1.55/1.15 | - | 1.8/1.59/1.35 |
| Telson | L/W/D | 0.91/0.71/0.83 | 1.13/0.82/0.76 | 1.04/0.78/0.90 | - | 1.14/0.81/0.84 |
| Pedipalp | L | 6.47 | 6.37 | 6.68 | 7.42 | 8.36 |
| Femur | L/W | 1.66/0.48 | 1.68/0.46 | 1.66/0.47 | 1.51/0.79 | 1.71/0.52 |
| Patella | L/W | 1.94/0.71 | 1.95/0.76 | 2.07/0.74 | 3.08/1.11 | 2/0.76 |
| Chela | L | 2.87 | 2.74 | 2.95 | 2.83 | 4.65 |
| Manus | W/D | 0.57/0.54 | 0.56/0.53 | 0.65/0.59 | 0.93/0.62 | 0.58/0.55 |
| Movable finger | L | 2 | 1.64 | 2.02 | 1.91 | 2.01 |
| Pectine | L/W | 1.78/0.45 | 1.77/0.4 | 1.82/0.41 | 1.63/0.4 | 1.97/0.39 |
| Genital Operculum | L/W | 0.43/0.54 | 0.45/0.65 | 0.38/0.59 | 0.36/0.64 | 0.48/0.66 |
| Total | L | 14.49 | 14.46 | 14.97 | 10.39 | 15.14 |
| Pectinal teeth count | PTC | 16/16 | 16/16 | 17/17 | 16/17 | 16/16 |
| IES-608 (unidentified sex) | IES-609 (♀) | IES-610 (♀) | IES-613 (♀) | IES-617 (♀) | ||
| Carapace | L/W | 2.04/1.83 | 2.06/1.75 | 2.08/1.81 | 1.87/1.70 | 1.99/1.83 |
| Mesosoma | L | 3.94 | 4.15 | 3.81 | 3.92 | 3.64 |
| Tergite VII | L/W | 0.85/1.65 | 0.95/2.19 | 0.83/1.81 | 0.89/1.89 | 0.76/1.96 |
| Metasoma and telson | L | 8.86 | 8.63 | 7.93 | 8.76 | 8.54 |
| Segment I | L/W/D | 0.83/1.37/1.12 | 0.8/1.36/1.19 | 0.81/1.39/1.16 | 0.9/1.36/1.09 | 0.82/1.36/1.15 |
| Segment II | L/W/D | 1.15/1.4/1.18 | 1.00/1.4/1.27 | 1.17/1.37/1.22 | 1.07/1.44/1.1 | 1.03/1.44/1.19 |
| Segment III | L/W/D | 1.27/1.46/1.24 | 1.18/1.46/1.39 | 1.26/1.48/1.32 | 1.19/1.51/1.22 | 1.20/1.42/1.22 |
| Segment IV | L/W/D | 1.52/1.49/1.23 | 1.54/1.44/1.39 | 1.34/1.5/1.3 | 1.48/1.56/1.14 | 1.52/1.48/1.24 |
| Segment V | L/W/D | 2.22/1.49/1.17 | 2.25/1.54/1.35 | 1.73/1.55/1.28 | 2.22/1.57/1.18 | 2.18/1.43/1.25 |
| Telson | L/W/D | 1.1/0.75/0.84 | 1.09/0.75/0.89 | 0.99/0.76/0.79 | 0.99/0.79/0.96 | 1/0.71/0.94 |
| Pedipalp | L | 6.09 | 6.42 | 6.17 | 6.08 | 6.16 |
| Femur | L/W | 1.61/0.51 | 1.65/0.51 | 1.65/0.49 | 1.6/0.48 | 1.54/0.53 |
| Patella | L/W | 1.77/0.71 | 1.94/0.75 | 1.85/0.68 | 1.89/0.70 | 1.93/0.75 |
| Chela | L | 2.71 | 2.83 | 2.67 | 2.59 | 2.69 |
| Manus | W/D | 0.52/0.61 | 0.56/0.5 | 0.57/0.61 | 0.56/0.6 | 0.61/0.58 |
| Movable finger | L | 2.09 | 1.84 | 1.88 | 1.72 | 1.96 |
| Pectine | L/W | 1.67/0.39 | 1.57/0.37 | 1.61/0.4 | 1.55/0.3 | 1.55/0.34 |
| Genital Operculum | L/W | 0.4/0.59 | 0.44/0.62 | 0.44/0.6 | 0.33/0.68 | 0.36/0.53 |
| Total | L | 14.84 | 14.84 | 13.82 | 14.55 | 14.17 |
| Pectinal teeth count | PTC | 17/17 | 16/17 | 16/16 | 16/17 | 16/16 |
| IES-475 (♂) | IES-479(♂) | IES-483(♂) | IES-474(♀) | |||
|---|---|---|---|---|---|---|
| Carapace | L/W | 2.1/1.85 | 2.06/1.85 | 1.64/1.42 | 1.69/1.49 | |
| Mesosoma | L/W | 4.55 | 4.10 | 2.98 | 2.48 | |
| Tergite VII | L/W | 1.13/2.43 | 0.93/1.95 | 0.80/1.47 | 0.7/1.78 | |
| Metasoma and telson | L | 8.88 | 8.41 | 7.59 | 7.32 | |
| Segment I | L/W/D | 0.84/1.47/1.26 | 0.94/1.41/1.11 | 0.85/1.21/0.98 | 0.73/1.17/1.03 | |
| Segment II | L/W/D | 1.15/1.53/1.37 | 1.12/1.46/1.19 | 1.00/1.12/1.09 | 1.01/1.26/1.12 | |
| Segment III | L/W/D | 1.24/1.58/1.50 | 1.2/1.5/1.29 | 1.07/1.2/1.12 | 1.16/1.2/1.21 | |
| Segment IV | L/W/D | 1.65/1.6/1.50 | 1.141/1.52/1.36 | 1.18/1.26/1.14 | 1.25/1.22/1.15 | |
| Segment V | L/W/D | 2.06/1.58/1.51 | 1.95/1.57/1.42 | 1.86/1.27/1.16 | 1.74/1.22/1.13 | |
| Telson | L/W/D | 1.16/0.89/0.94 | 1.07/0.82/0.95 | 0.97/0.68/0.90 | 0.78/0.66/0.79 | |
| Pedipalp | L | 6.72 | 6.48 | 5.29 | 4.19 | |
| Femur | L/W | 1.61/0.59 | 1.60/0.48 | 1.34/0.47 | 0.91/0.43 | |
| Patella | L/W | 2.09/0.78 | 1.98/0.77 | 1.64/0.58 | 1.02/0.59 | |
| Chela | L | 3.02 | 2.90 | 2.31 | 2.26 | |
| Manus | W/D | 0.61/0.64 | 0.67/0.75 | 0.47/0.52 | 0.43/0.47 | |
| Movable finger | L | 2.08 | 1.91 | 1.53 | 1.56 | |
| Pectine | L/W | 1.57/0.36 | 1.46/0.35 | 1.11/0.26 | 1.31/0.31 | |
| Genital Operculum | L/W | 0.28/0.47 | 0.44/0.58 | 0.29/0.38 | 0.32/0.42 | |
| Total | L | 15.53 | 14.57 | 12.21 | 11.49 | |
| Pectinal teeth count | PTC | 16/16 | 15/16 | 15/17 | 15/15 | |
| IES-476(♀) | IES-477(♀) | IES-481 (♀) | IES-482 (♀) | IES-586 (unidentidied sex) | ||
| Carapace | L/W | 1.67/1.41 | 2.06/1.8 | 2.14/1.9 | 1.65/1.4 | 1.88/1.70 |
| Mesosoma | L/W | 3.03 | 6.21 | 4.26 | 2.62 | 3.23 |
| Tergite VII | L/W | 0.8/1.45 | 1.18/2.37 | 0.93/2.24 | 0.72/1.57 | 0.91/1.8 |
| Metasoma and telson | L | 7.40 | 8.70 | 9.39 | 7.11 | 8.42 |
| Segment I | L/W/D | 0.67/1.05/0.98 | 0.69/1.47/1.27 | 0.91/1.43/1.26 | 0.73/1.19/0.93 | 0.83/1.15/1.06 |
| Segment II | L/W/D | 0.9/1.12/1.01 | 1.35/1.5/1.32 | 1.12/1.32/1.22 | 0.96/1.14/0.93 | 1.12/1.11/1.08 |
| Segment III | L/W/D | 0.96/1.16/1.04 | 1.44/1.58/1.39 | 1.31/1.51/1.32 | 1.00/1.81/1.14 | 1.19/1.16/1.07 |
| Segment IV | L/W/D | 1.36/1.22/1.05 | 1.49/1.53/1.42 | 1.61/1.47/1.45 | 1.06/1.18/1.15 | 1.48/1.13/1.05 |
| Segment V | L/W/D | 2.01/1.23/1.13 | 1.96/1.62/1.27 | 2.26/1.56/1.44 | 1.71/1.16/0.99 | 1.91/1.17/1.04 |
| Telson | L/W/D | 0.89/0.64/0.74 | 1.17/0.82/0.96 | 1.09/0.85/0.96 | 0.96/0.63/0.78 | 1.1/0.79/0.86 |
| Pedipalp | L | 4.86 | 6.05 | 5.98 | 4.76 | 5.98 |
| Femur | L/W | 1.31/0.43 | 1.75/0.55 | 1.62/0.51 | 1.16/0.51 | 1.6/0.44 |
| Patella | L/W | 1.48/0.53 | 1.87/0.78 | 1.69/0.7 | 1.53/0.57 | 1.86/0.63 |
| Chela | L | 2.07 | 2.43 | 2.67 | 2.07 | 2.52 |
| Manus | W/D | 0.43/0.45 | 0.60/0.63 | 0.58/0.63 | 0.39/0.45 | 0.49/0.54 |
| Movable finger | L | 1.50 | 2.00 | 1.8 | 1.44 | 1.93 |
| Pectine | L/W | 1.59/0.40 | 1.76/0.35 | 1.62/0.37 | 1.31/0.32 | 1.66/0.32 |
| Genital Operculum | L/W | 0.32/0.47 | 0.47/0.62 | 0.45/0.57 | 0.29/0.57 | 0.32/0.48 |
| Total | L | 12.1 | 16.97 | 15.79 | 11.38 | 13.53 |
| Pectinal teeth count | PTC | 16/17 | 18/16 | 16/16 | 16/15 | - |

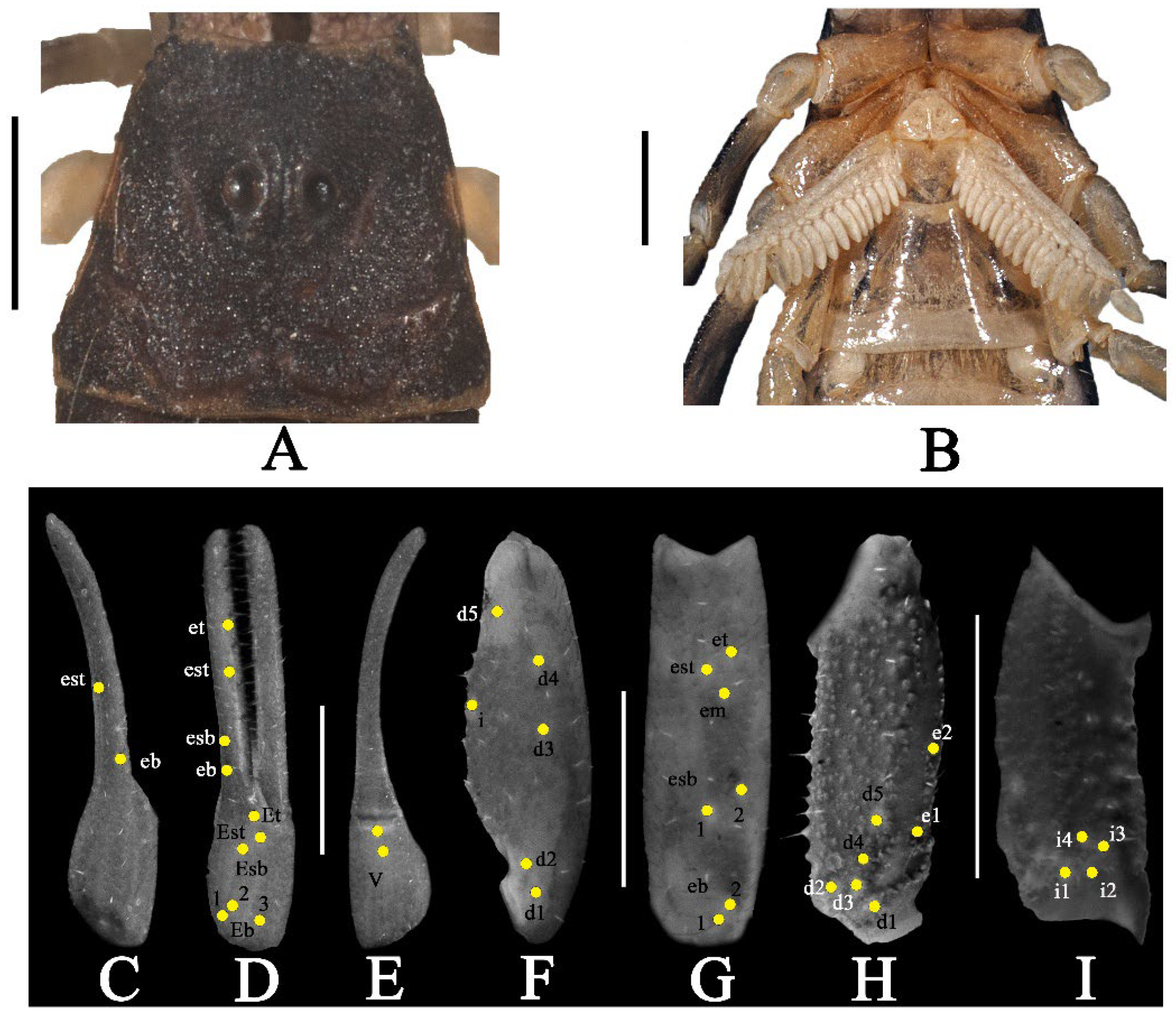

3.4. Charmus dakshini sp. nov.
urn:lsid:zoobank.org:pub:4DD30BD9-355F-4837-9053-C51489896E88
| BNHS SC-404 (♂) | IES-624 (♂) | IES-626 (♂) | IES-627 (♂) | |||
|---|---|---|---|---|---|---|
| Holotype | Paratype | Paratype | Paratype | |||
| Carapace | L/W | 2.15/1.84 | 2.09/1.75 | 2.01/1.77 | 2.09/1.74 | |
| Mesosoma | L | 3.68 | 3.61 | 3.18 | 3.63 | |
| Tergite VII | L/W | 1.06/1.80 | 1.01/1.9 | 0.92/1.85 | 1.04/1.8 | |
| Metasoma and telson | L | 9.46 | 8.93 | 9.50 | 5.53 | |
| Segment I | L/W/D | 1.01/1.36/0.93 | 1.08/1.37/1.18 | 1.1/1.35/0.94 | 1.07/1.31/0.9 | |
| Segment II | L/W/D | 1.05/1.33/0.94 | 1.11/1.36/1.25 | 1.17/1.3/1.08 | 1.04/1.31/0.98 | |
| Segment III | L/W/D | 1.31/1.38/1.06 | 1.29/1.37/1.33 | 1.37/1.34/1.13 | 1.24/1.3/1.08 | |
| Segment IV | L/W/D | 1.62/1.36/1.04 | 1.59/1.35/1.35 | 1.62/1.35/1.15 | - | |
| Segment V | L/W/D | 2.41/1.35/1.07 | 1.87/1.33/1.25 | 2.14/1.35/1.16 | - | |
| Telson | L/W/D | 1.15/0.79/0.95 | 1.27/0.97/0.94 | 1.32/0.84/0.92 | 1.35/0.94/0.98 | |
| Pedipalp | L | 6.97 | 7.88 | 6.48 | 6.76 | |
| Femur | L/W | 1.81/0.58 | 2.03/0.64 | 1.8/0.57 | 1.68/0.64 | |
| Patella | L/W | 2.19/0.79 | 2.37/0.92 | 2.16/0.76 | 2.12/0.89 | |
| Chela | L | 2.97 | 3.48 | 2.52 | 2.96 | |
| Manus | W/D | 0.62/0.70 | 0.70/0.62 | 0.61/0.64 | 0.64/0.74 | |
| Movable finger | L | 1.90 | 2.02 | 1.88 | 2.36 | |
| Pectine | L/W | 1.98/0.4 | 2.07/0.41 | 1.91/0.38 | 1.22/0.44 | |
| Genital Operculum | L/W | 0.38/0.7 | 0.32/0.53 | 0.37/0.55 | 0.32/0.34 | |
| Total | L | 15.29 | 14.63 | 14.69 | 11.25 | |
| Pectinal teeth count | PTC | 18/18 | 18/18 | 18/19 | 18/19 | |
| IES-385 (♂) | IES-621 (♀) | IES-622 (♀) | IES-623 (♀) | IES-386 (♀) | ||
| Paratype | Paratype | Paratype | Paratype | Paratype | ||
| Carapace | L/W | 2.01/1.88 | 2.04/1.76 | 2.01/1.78 | 1.95/1.7 | 2.23/1.66 |
| Mesosoma | L | 4.36 | 4.40 | 3.15 | 3.34 | 4.07 |
| Tergite VII | L/W | 1.19/1.80 | 1.1/1.73 | 0.86/1.99 | 0.94/1.78 | 1.13/2.02 |
| Metasoma and telson | L | 9.01 | 9.50 | 8.40 | 8.56 | 10.05 |
| Segment I | L/W/D | 0.84/1.31/1.19 | 1.02/1.28/1.02 | 0.91/1.18/1.28 | 0.82/1.26/1.23 | 1.03/1.44.1.33 |
| Segment II | L/W/D | 1.06/1.22/1.13 | 1.15/1.25/1.04 | 1.01/1.29/1.29 | 0.98/1.26/1.16 | 1.22/1.43/1.3 |
| Segment III | L/W/D | 1.4/1.23/1.17 | 1.34/1.28/1.04 | 1.16/1.26/1.31 | 1.15/1.23/1.31 | 1.38/1.4/1.35 |
| Segment IV | L/W/D | 1.53/1.23/1.16 | 1.66/1.38/1.04 | 1.37/1.33/1.35 | 1.49/1.26/1.3 | 1.66/1.42/1.33 |
| Segment V | L/W/D | 2.25/1.28/1.12 | 2.1/1.36/1.04 | 2.15/1.35/1.21 | 2.17/1.28/1.25 | 2.70/1.57/1.26 |
| Telson | L/W/D | 1.09/0.77/0.9 | 1.41/0.85/1.00 | 1.2/0.85/0.99 | 1.21/0.77/0.94 | 1.18/0.98/1.21 |
| Pedipalp | L | 6.77 | 7.09 | 6.64 | 6.42 | 7.37 |
| Femur | L/W | 1.65/0.51 | 1.85/0.59 | 1.76/0.54 | 1.67/0.54 | 1.99/0.64 |
| Patella | L/W | 2.14/0.68 | 2.21/0.79 | 2.10/0.72 | 1.95/0.75 | 2.28/0.92 |
| Chela | L | 2.98 | 3.03 | 2.78 | 2.80 | 3.10 |
| Manus | W/D | 0.54/0.58 | 0.62/0.59 | 0.61/0.58 | 0.6/0.61 | 0.73/0.71 |
| Movable finger | L | 1.76 | 2.15 | 2.16 | 2.10 | 2.47 |
| Pectine | L/W | 1.86/0.37 | 2.01/0.41 | 1.74/0.31 | 1.82/0.32 | 2.05/0.47 |
| Genital Operculum | L/W | 0.37/0.53 | 0.44/0.64 | 0.41/0.58 | 0.37/0.61 | 0.37/0.68 |
| Total | L | 15.38 | 15.94 | 13.56 | 13.85 | 16.35 |
| Pectinal teeth count | PTC | - | 18/19 | 18/18 | 17/18 | 18/18 |
| IES-618 (♂) | IES-619 (♂) | IES-620 (♀) | IES-401 (♀) | ||
|---|---|---|---|---|---|
| Paratype | Paratype | Paratype | Paratype | ||
| Carapace | L/W | 2.22/1.85 | 1.96/1.64 | 2.03/1.73 | 2.1/2.06 |
| Mesosoma | L | 3.96 | 3.20 | 3.69 | 3.27 |
| Tergite VII | L/W | 1.05/2.06 | 0.9/1.71 | 1/1.79 | 0.95/1.93 |
| Metasoma and telson | L | 9.93 | 9.09 | 9.53 | 7.88 |
| Segment I | L/W/D | 1.25/1.47/1.22 | 1.13/1.18/1.05 | 1.01/1.24/1.23 | 1.13/1.32/1.22 |
| Segment II | L/W/D | 1.28/1.41/1.23 | 1.17/1.16/1.06 | 1.2/1.14/1.2 | 1.41/1.29/1.22 |
| Segment III | L/W/D | 1.46/1.39/1.27 | 1.24/1.17/1.05 | 1.36/1.2/1.24 | 1.47/1.32/1.21 |
| Segment IV | L/W/D | 1.52/1.42/1.22 | 1.38/1.2/1.05 | 1.58/1.24/1.2 | 1.61/1.3/1.22 |
| Segment V | L/W/D | 2.38/1.46/1.23 | 2.18/1.19/1.08 | 2.34/1.27/1.14 | 2.26/1.32/1.21 |
| Telson | L/W/D | 1.29/0.84/1.03 | 1.21/0.77/0.91 | 1.26/0.77/0.93 | - |
| Pedipalp | L | 7.23 | 5.69 | 7.14 | 6.86 |
| Femur | L/W | 1.80/0.58 | 1.53/0.51 | 1.69/0.55 | 1.63/0.56 |
| Patella | L/W | 2.25/0.85 | 1.78/0.68 | 2.35/0.75 | 2.10/0.76 |
| Chela | L | 3.18 | 2.38 | 3.10 | 3.13 |
| Manus | W/D | 0.64/0.65 | 0.53/0.56 | 0.59/0.58 | 0.63/0.63 |
| Movable finger | L | 2.00 | 2.15 | 2.14 | 2.10 |
| Pectine | L/W | 2.07/0.41 | 1.79/0.40 | 1.7/0.33 | 1.97/0.42 |
| Genital Operculum | L/W | 0.37/0.57 | 0.38/0.58 | 0.35/0.54 | 0.35/0.52 |
| Total | L | 16.11 | 14.25 | 15.25 | 13.25 |
| Pectinal teeth count | PTC | 18/17 | 19/19 | 16/17 | 19/19 |

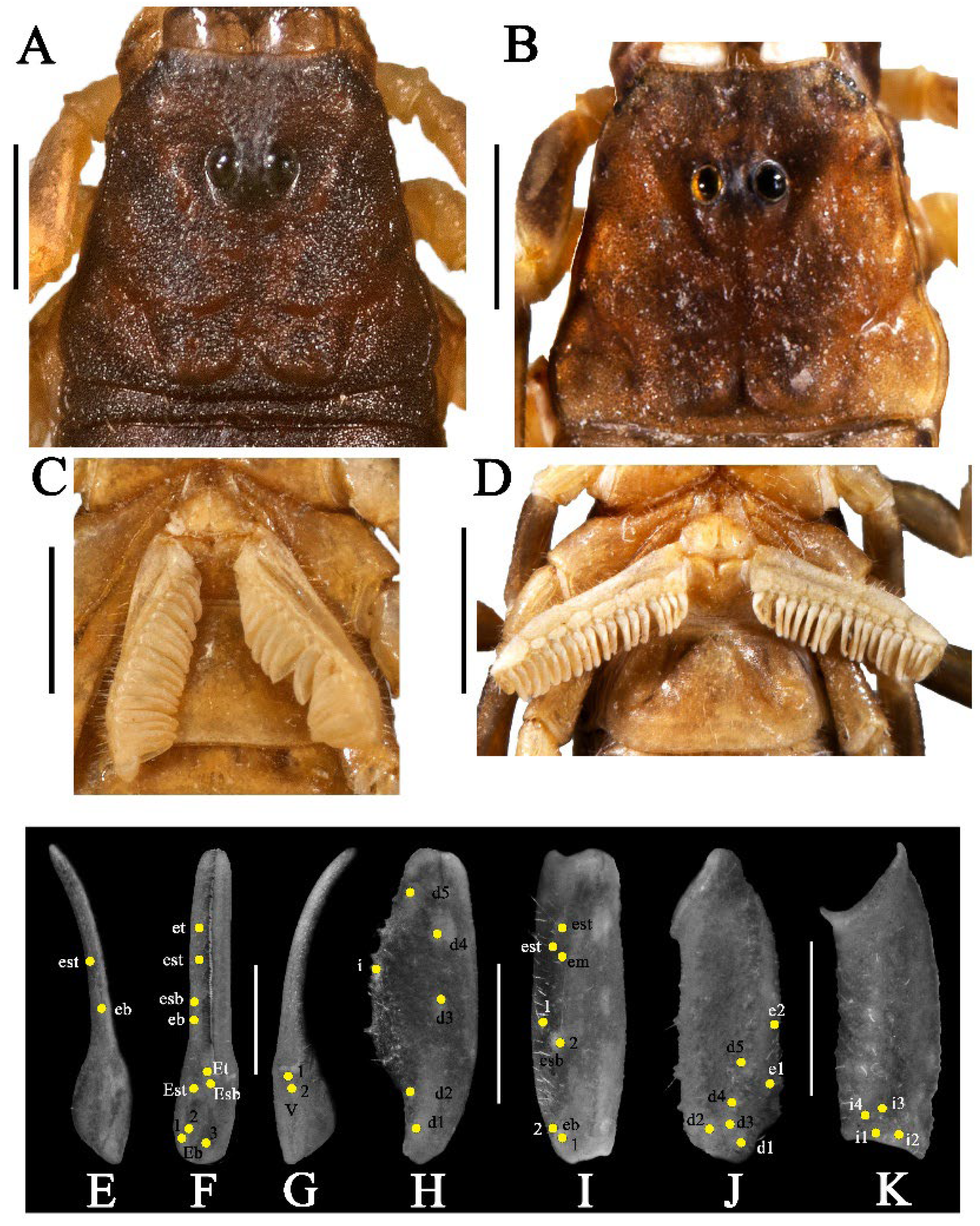

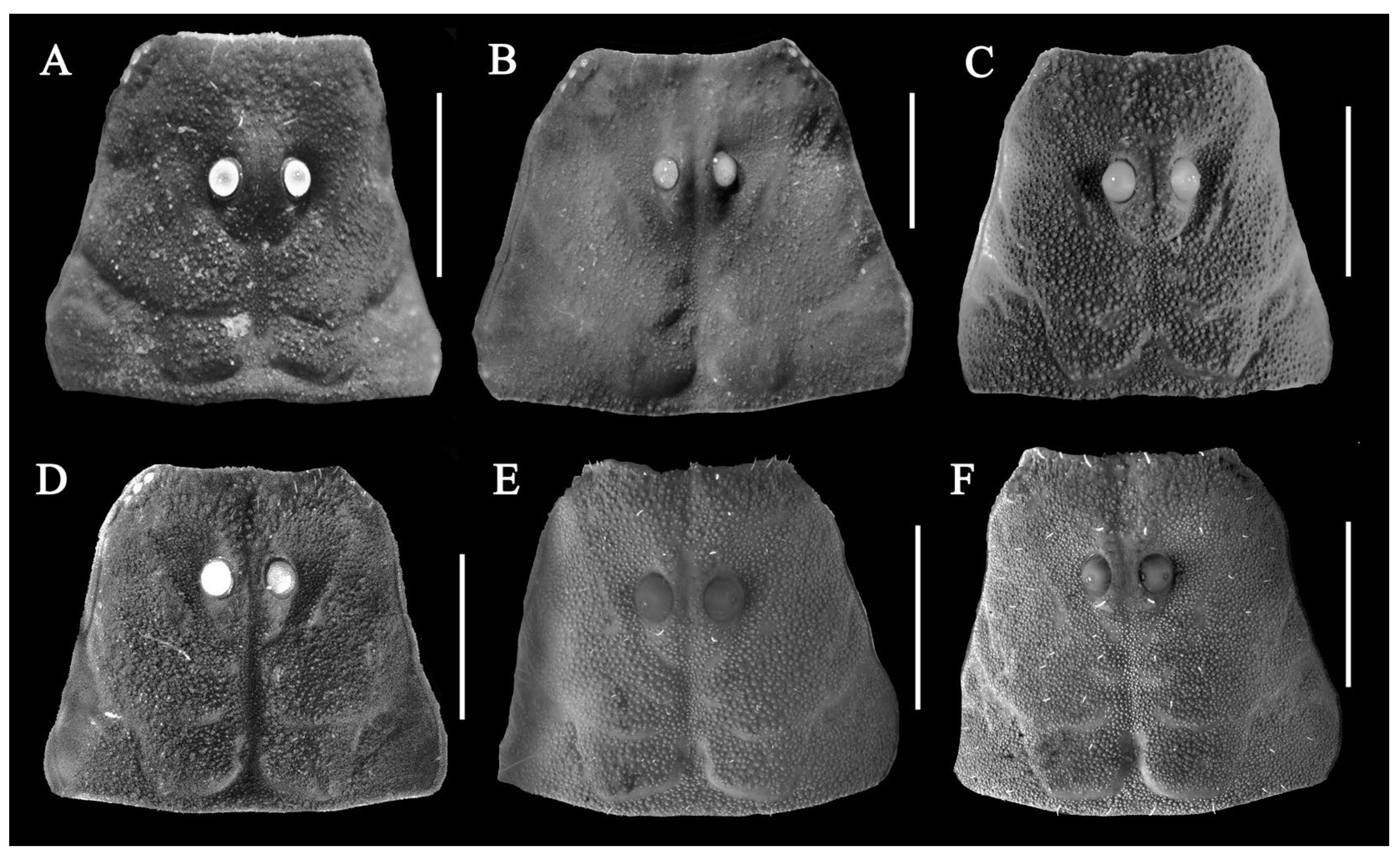
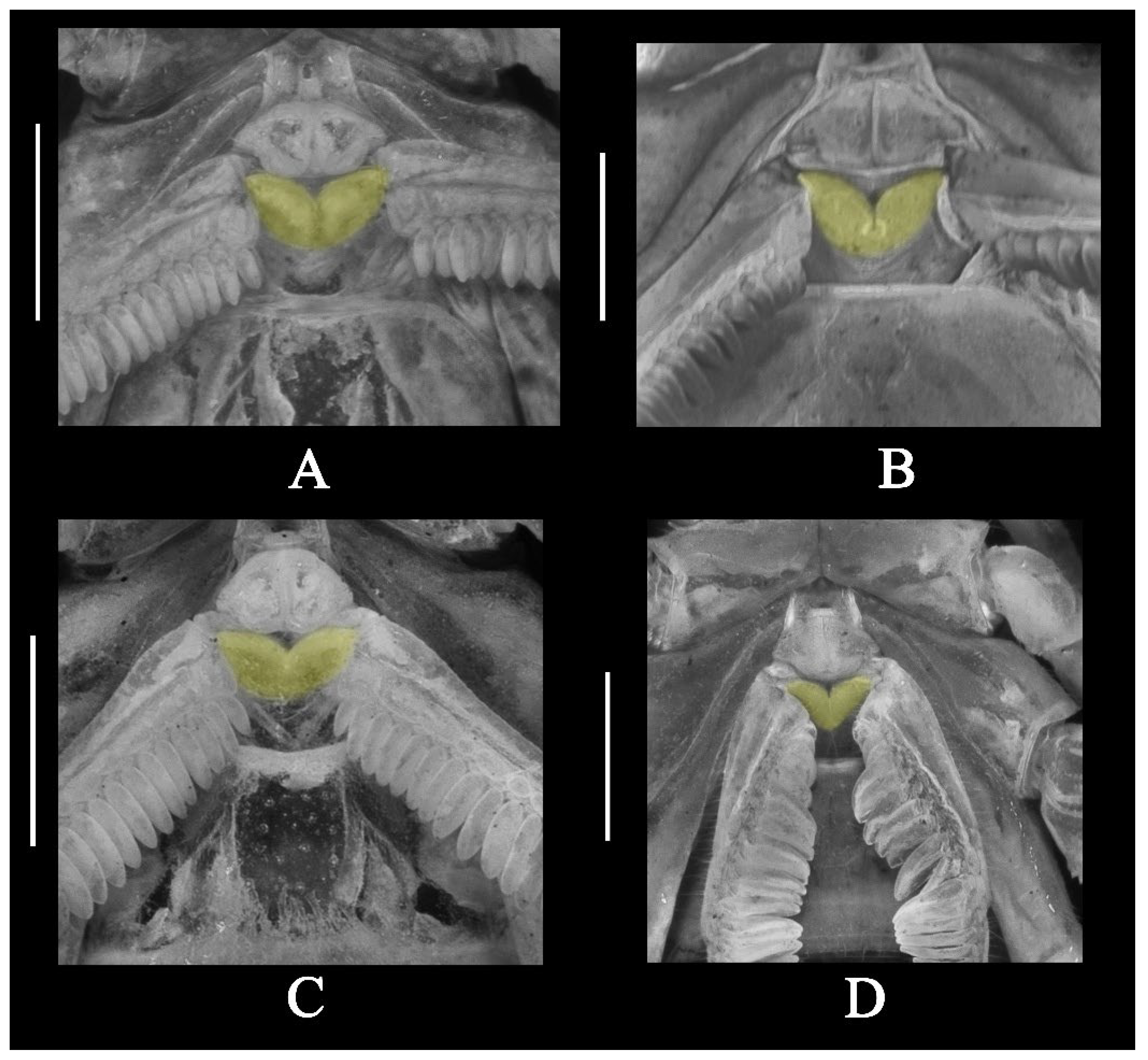
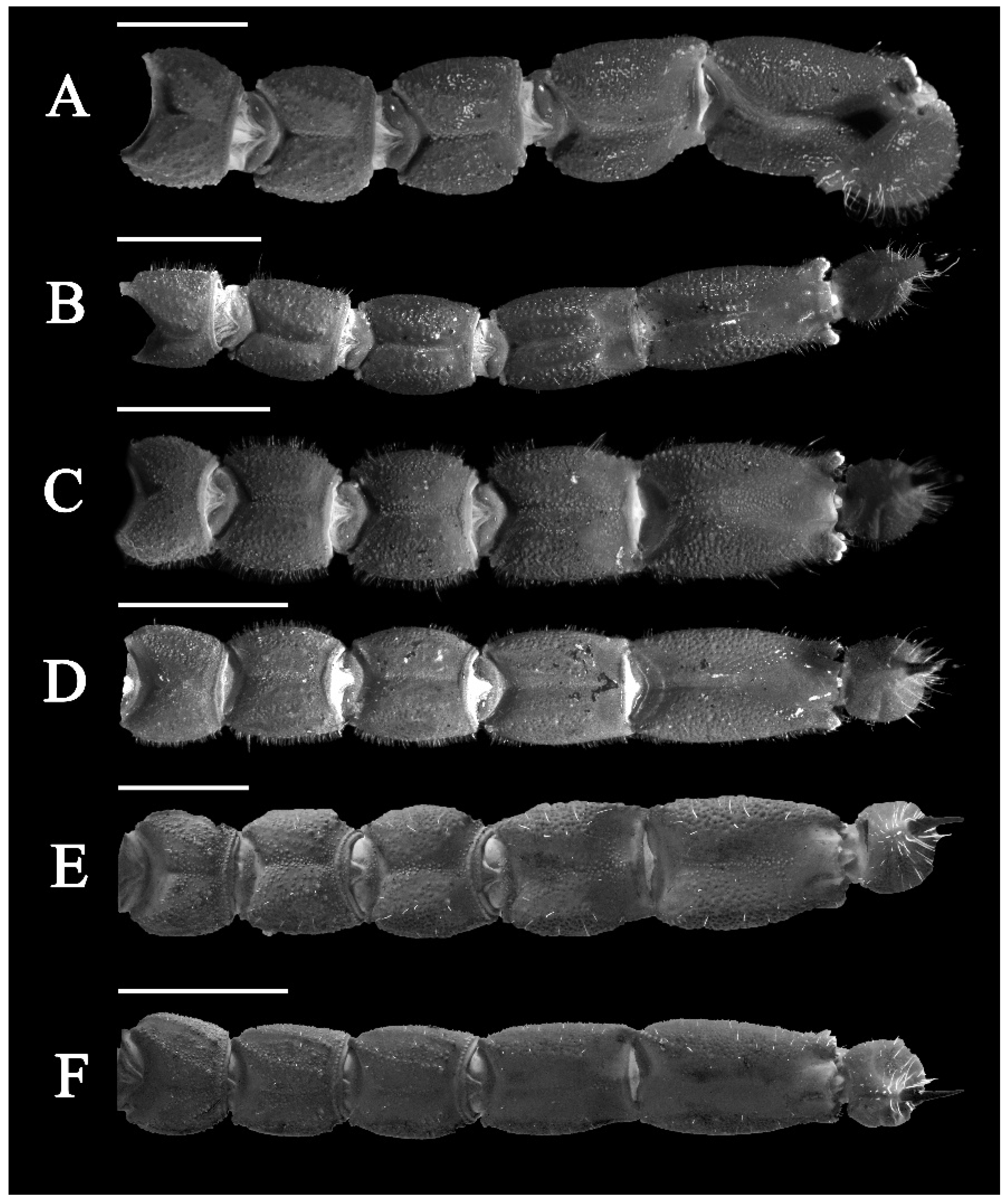
3.5. Affinities (Figure 2, Figure 3, Figure 5, Figure 6, Figure 8, Figure 9, Figure 10, Figure 12, Figure 13, Figure 15, Figure 16, Figure 17, Figure 18, Figure 19 and Figure 20)
- Surface of carapace finely and densely granular in C. sinhagadensis (Figure 6A and Figure 15B) and C. dakshini sp. nov. (Figure 13A and Figure 15D) as opposed to surface with mixed and dense granulation in C. laneus (Figure 3A and Figure 15A,E), C. brignolii (Figure 9A and Figure 15C) and C. saradieli (Figure 15F).
- Anterior margin of carapace finely crenulated in the median portion in C. laneus (Figure 3A and Figure 15A,E), C. brignolii (Figure 9A and Figure 15C), C. saradieli (Figure 15F), and C. dakshini sp. nov. (Figure 13A and Figure 15D) as opposed to smooth throughout in C. sinhagadensis (Figure 6A and Figure 15B).
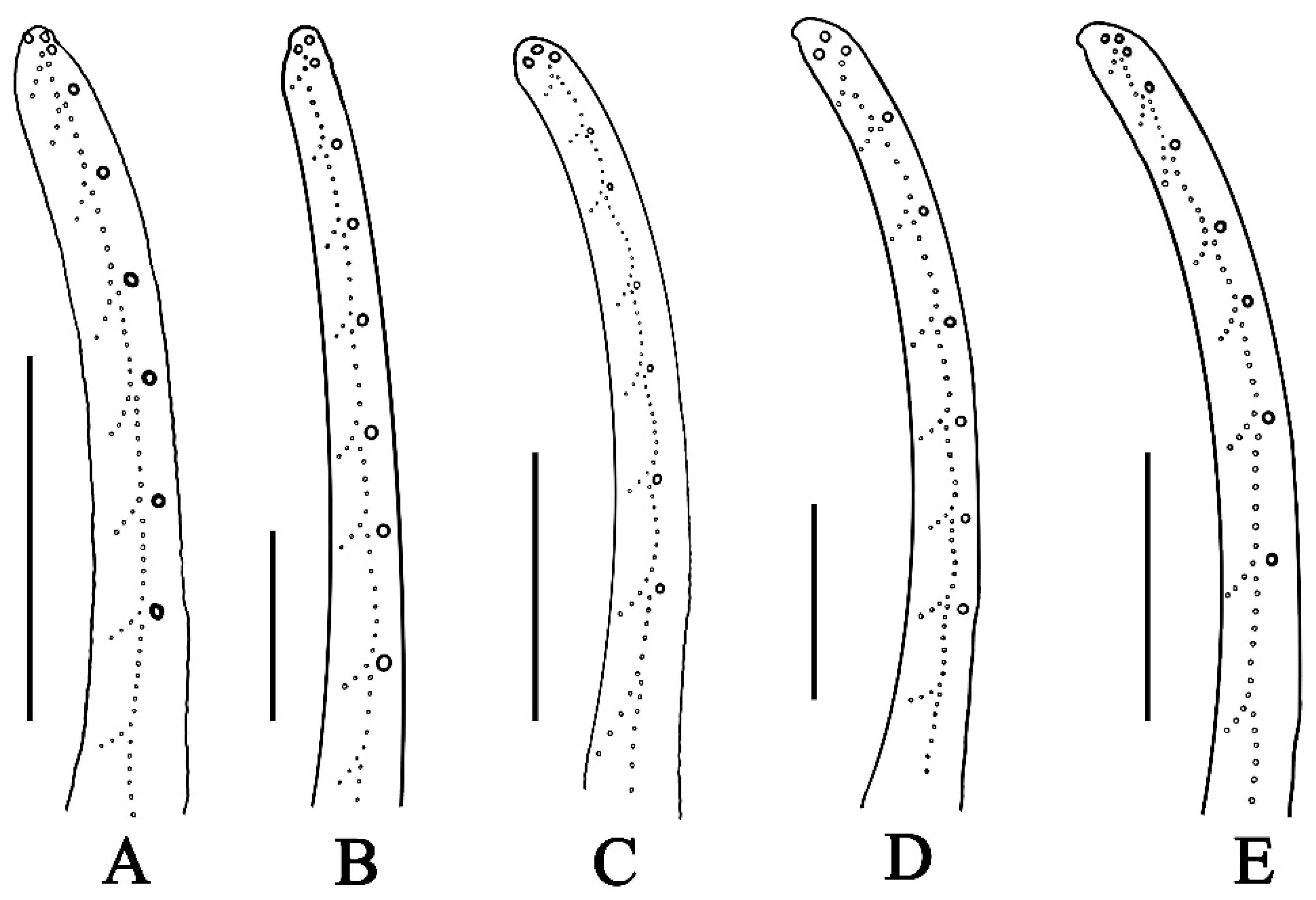
- Ventral submedian carina of metasomal segment (Figure 19) I–III weakly developed in C. dakshini sp. nov. as opposed to moderately developed in C. sinhagadensis and strongly developed in C. laneus C. brignolii and C. saradieli.
- Metasomal segment V acarinate (Figure 17, Figure 18, Figure 19 and Figure 20) in C. dakshini sp. nov. as opposed to segment with a pair of weakly developed dorsal lateral carina present on the proximal portion in C. saradieli, moderately developed and present on the proximal portion in C. sinhagadensis and strongly developed and present on the proximal portion in C. laneus and C. brignolii.
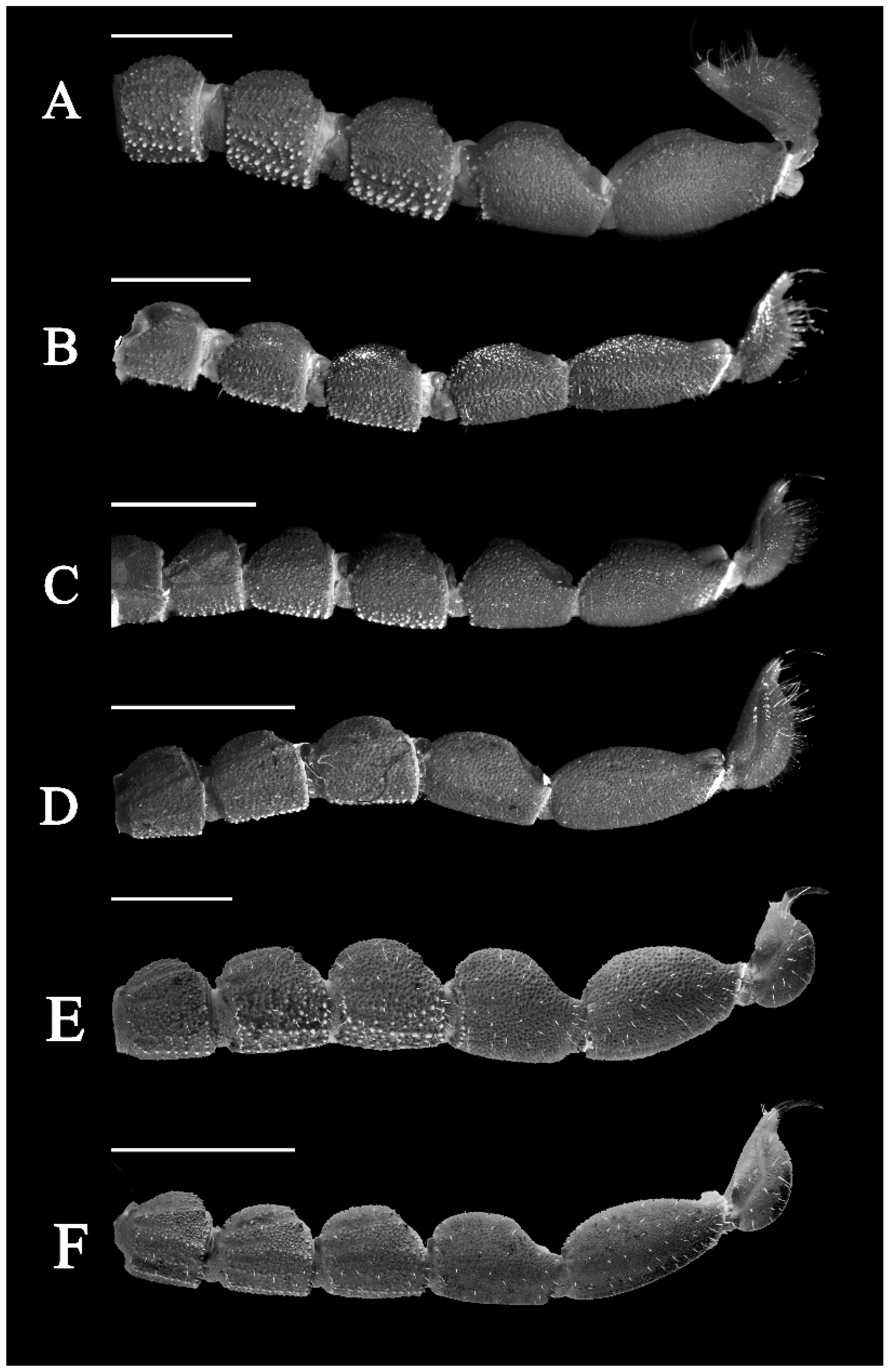
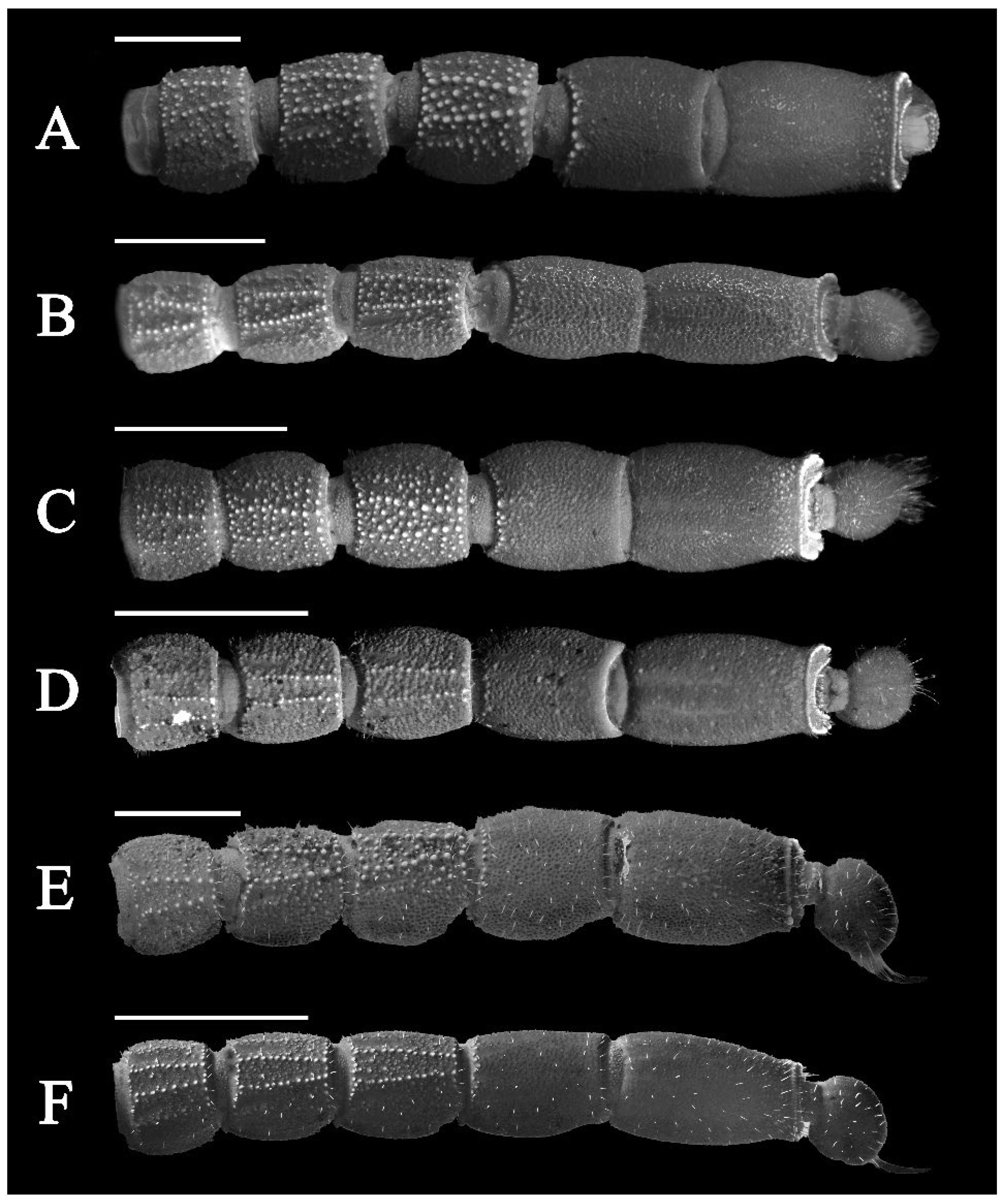

4. Results
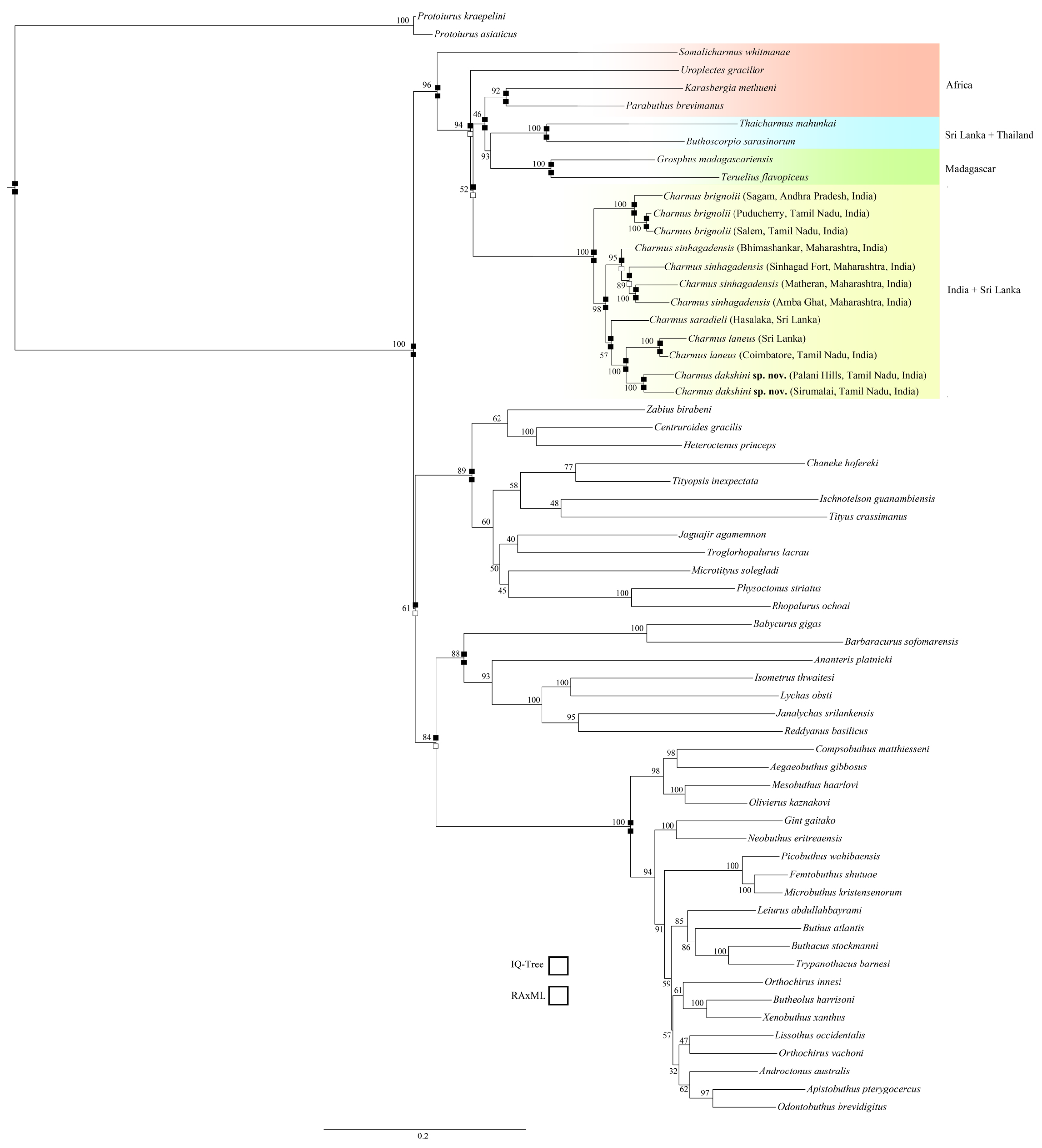
4.1. Molecular Phylogeny of Charmus Based on Sanger-Sequenced Data (Figure 21 and Figure S1)
4.2. Comparisons of p-Distance with Closest Related Congeners (Tables S1–S3)
4.3. Molecular Phylogenetics Based on the UCE Data (Figure 22)

5. Discussion
5.1. Congruence Between Morphology, Genetic Divergence, Phylogeny and Geography
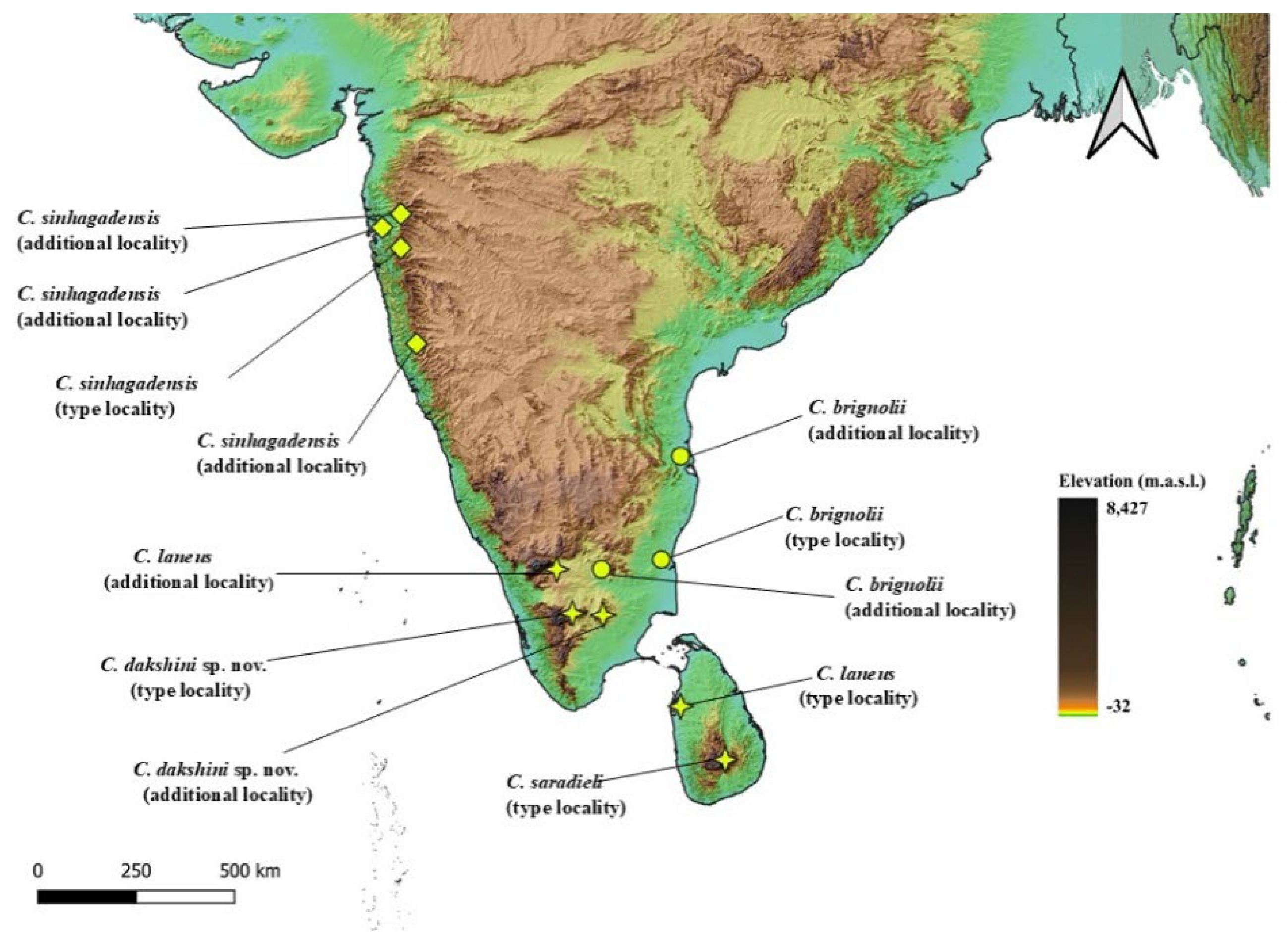
5.2. Morphological Difference Between Lowland and Highland Species
5.3. Topological Differences Between Phylogenies
5.4. Biogeographic Considerations
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Perera, S.J.; Suranjanfernando, R.H.S. Physiography, Climate, and Historical Biogeography of Sri Lanka in Making a Biodiversity Hotspot. In Biodiversity Hotspots of the Western Ghats and Sri Lanka; Apple Academic Press: Palm Bay, FL, USA, 2022; pp. 405–445. [Google Scholar]
- Dissanayake, C.B.; Chandrajith, R. Sri Lanka–Madagascar Gondwana Linkage: Evidence for a Pan-African Mineral Belt. J. Geol. 1999, 107, 223–235. [Google Scholar] [CrossRef]
- Deepak, V.; Karanth, P. Aridification Driven Diversification of Fan-Throated Lizards from the Indian Subcontinent. Mol. Phylogenet. Evol. 2018, 120, 53–62. [Google Scholar] [CrossRef] [PubMed]
- Gorin, V.A.; Solovyeva, E.N.; Hasan, M.; Okamiya, H.; Karunarathna, D.M.S.S.; Pawangkhanant, P.; de Silva, A.; Juthong, W.; Milto, K.D.; Nguyen, L.T.; et al. A little frog leaps a long way: Compounded colonizations of the Indian Subcontinent discovered in the tiny Oriental frog genus Microhyla (Amphibia: Microhylidae). PeerJ 2020, 8, e9411. [Google Scholar] [CrossRef] [PubMed]
- Prendini, L.; Weygoldt, P.; Wheeler, W.C. Systematics of the Damon variegatus group of African whip spiders (Chelicerata: Amblypygi): Evidence from behavior, morphology, and DNA. Org. Divers. Evol. 2005, 5, 203–236. [Google Scholar] [CrossRef]
- Seiter, M.; Lerma, A.; Král, J.; Sember, A.; Divišová, K.; Palacios-Vargas, J.; Colmenares, P.A.; Loria, S.F.; Prendini, L. Cryptic diversity in the whip spider genus Paraphrynus (Amblypygi: Phrynidae): Integrating morphology, karyotype, and DNA. Arthropod Syst. Phylogeny 2020, 78, 265–285. [Google Scholar]
- Schramm, F.D.; Valdez-Mondragón, A.; Prendini, L. Volcanism and palaeoclimate change drive diversification of the world’s largest whip spider (Amblypygi). Mol. Ecol. 2021, 12, 2872–2890. [Google Scholar] [CrossRef]
- Karsch, F. Skorpionologische Beiträge I. und II. Mitt. Münch. Entomol. Ver. 1879, 3, 6–22, 97–136. [Google Scholar]
- Hirst, S. Description of a new Indian Scorpion (Charmus indicus sp. n.). Ann. Mag. Nat. Hist. 1915, 15, 224–225. [Google Scholar] [CrossRef]
- Sreenivasa-Reddy, R.P. Contribution à la connaissance des Scorpions de l’Inde. I. Charmus indicus Hirst, 1915 (Fam. Buthidae). Bull. Mus. Natl. Hist. 1966, 38, 247–256. [Google Scholar]
- Tikader, B.K.; Bastawade, D.B. The Fauna of India; Zoological Survey of India: Calcutta, India, 1983; Volume 3, pp. 146–152. [Google Scholar]
- Lourenço, W.R. Taxonomic considerations about the genus Charmus Karsch, 1879 with description of a new species to India (Scorpiones: Buthidae). Mem. Soc. Entomol. 2000, 78, 295–303. [Google Scholar]
- Lourenço, W.R. Further taxonomic considerations about the genus Charmus Karsch, 1879 (Scorpiones, Buthidae), with the description of a new species from Sri Lanka. Entomol. Mitt. Zool. Mus. Hamb. 2002, 14, 17–25. [Google Scholar]
- Kovarik, F.; Lowe, G.; Ranawana, K.B.; Hoferek, D.; Jayarathne, V.A.S.; Pliskova, J. Scorpions of Sri Lanka (Arachnida, Scorpiones: Buthidae, Chaerilidae, Scorpionidae) with Description of Four New Species of the Genera Charmus Karsch, 1879 and Reddyanus Vachon, 1972, stat. n. Euscorpius 2016, 220, 1–133. [Google Scholar] [CrossRef]
- International Commission on Zoological Nomenclature (ICZN). International Code of Zoological Nomenclature; Online Edition; International Commission on Zoological Nomenclature (ICZN): Singapore, 2000; Available online: https://www.iczn.org/the-code/the-international-code-of-zoological-nomenclature/the-code-online/ (accessed on 7 December 2024).
- Wong, E.S.; Dahlem, G.A.; Stamper, T.I.; DeBry, R.W. Discordance between Morphological Species Identification and mtDNA Phylogeny in the Flesh Fly Genus Ravinia (Diptera: Sarcophagidae). Invertebr. Syst. 2015, 29, 1–11. [Google Scholar] [CrossRef]
- Andriaholinirina, N.; Fausser, J.L.; Roos, C.; Zinner, D.; Thalmann, U.; Rabarivola, C.; Ravoarimanana, I.; Ganzhorn, J.U.; Meier, B.; Hilgartner, R.; et al. Molecular Phylogeny and Taxonomic Revision of the Sportive Lemurs (Lepilemur, Primates). BMC Evol. Biol. 2006, 6, 1–13. [Google Scholar] [CrossRef]
- Cook, L.G.; Edwards, R.D.; Crisp, M.D.; Hardy, N.B. Need Morphology Always Be Required for New Species Descriptions? Invertebr. Syst. 2010, 24, 322–326. [Google Scholar] [CrossRef]
- Gullan, P.J.; Downie, D.A.; Steffan, S.A. A New Pest Species of the Mealybug Genus Ferrisia Fullaway (Hemiptera: Pseudococcidae) from the United States. Ann. Entomol. Soc. Am. 2003, 96, 723–737. [Google Scholar] [CrossRef]
- Sharma, P.K.; Fernández, R.M.N.; Esposito, L.A.; González-Santillán, E.; Monod, L. Phylogenomic Resolution of Scorpions Reveals Multilevel Discordance with Morphological Phylogenetic Signal. Proc. R. Soc. B Biol. Sci. 2015, 282, 20142953. [Google Scholar] [CrossRef]
- Santibáñez-López, C.E.; Ojanguren-Affilastro, A.A.; Sharma, P.P. Another One Bites the Dust: Taxonomic Sampling of a Key Genus in Phylogenomic Datasets Reveals More Non-Monophyletic Groups in Traditional Scorpion Classification. Invertebr. Syst. 2020, 34, 133–143. [Google Scholar] [CrossRef]
- Štundlová, J.; Šťáhlavský, F.; Opatova, V.; Stundl, J.; Kovařík, F.; Dolejš, P.; Šmíd, J. Molecular Data Do Not Support the Traditional Morphology-Based Groupings in the Scorpion Family Buthidae (Arachnida: Scorpiones). Mol. Phylogenet. Evol. 2022, 173, 107511. [Google Scholar] [CrossRef]
- Svenson, G.J.; Whiting, M.F. Reconstructing the origins of praying mantises (Dictyoptera: Mantodea): The roles of Gondwanan vicariance and morphological convergence. Cladistics 2009, 25, 468–514. [Google Scholar] [CrossRef]
- Hendrixson, E.; Bond, J.E. Evaluating the efficacy of continuous quantitative characters for reconstructing the phylogeny of a morphologically homogeneous spider taxon (Araneae, Mygalomorphae, Antrodiaetidae, Antrodiaetus). Mol. Phylogenet. Evol. 2009, 53, 300–313. [Google Scholar] [CrossRef] [PubMed]
- Ortiz, D.; Francke, O.F.; Bond, J.E. A tangle of forms and phylogeny: Extensive morphological homoplasy and molecular clock heterogeneity in Bonnetina and related tarantulas. Mol. Phylogenet. Evol. 2018, 127, 55–73. [Google Scholar] [CrossRef] [PubMed]
- Opatova, V.; Hamilton, C.A.; Hedin, M.; Montes De Oca, L.; Král, J.; Bond, J.E. Phylogenetic systematics and evolution of the spider infraorder Mygalomorphae using genomic scale data. Syst. Biol. 2020, 69, 671707. [Google Scholar] [CrossRef] [PubMed]
- Masonick, P.K.; Knyshov, A.; Gordon, E.R.L.; Forero, D.; Hwang, W.S.; Hoey-Chamberlain, R.; Bush, T.; Castillo, S.; Hernandez, M.; Ramirez, J.; et al. A revised classification of the assassin bugs (Hemiptera: Heteroptera: Reduviidae) based on combined analysis of phylogenomic and morphological data. Syst. Entomol. 2025, 50, 102–138. [Google Scholar] [CrossRef]
- Sharma, P.P.; Kaluziak, S.T.; Pérez-Porro, A.R.; González, V.L.; Hormiga, G.; Wheeler, W.C.; Giribet, G. Phylogenomic interrogation of Arachnida reveals systemic conflicts in phylogenetic signal. Mol. Biol. Evol. 2014, 31, 2963–2984. [Google Scholar] [CrossRef]
- Agnarsson, I.; Coddington, J.A.; Kuntner, M. Systematics: Progress in the study of spider diversity and evolution. In Spider Research in the 21st Century: Trends and Perspectives; Penney, D., Ed.; Siri Scientific Press: Greater Manchester, UK, 2013; pp. 58–111. [Google Scholar]
- Bond, J.E.; Garrison, N.L.; Hamilton, C.A.; Godwin, R.L.; Hedin, M.; Agnarsson, I. Phylogenomics resolves a spider backbone phylogeny and rejects a prevailing paradigm for orb web evolution. Curr. Biol. 2014, 24, 1765–1771. [Google Scholar] [CrossRef] [PubMed]
- Fernández, R.; Hormiga, G.; Giribet, G. Phylogenomic analysis of spiders reveals nonmonophyly of orb weavers. Curr. Biol. 2014, 24, 1772–1777. [Google Scholar] [CrossRef]
- Godwin, R.L.; Opatova, V.; Garrison, N.L.; Hamilton, C.A.; Bond, J.E. Phylogeny of a cosmopolitan family of morphologically conserved trapdoor spiders (Mygalomorphae, Ctenizidae) using Anchored Hybrid Enrichment, with a description of the family Halonoproctidae Pocock 1901. Mol. Phylogenet. Evol. 2018, 126, 303–313. [Google Scholar] [CrossRef]
- Hedin, M.; Derkarabetian, S.; Ramírez, M.J.; Vink, C.; Bond, J.E. Phylogenomic reclassification of the world’s most venomous spiders (Mygalomorphae, Atracinae), with implications for venom evolution. Sci. Rep. 2018, 8, 1636. [Google Scholar] [CrossRef]
- Wood, H.M.; González, V.L.; Lloyd, M.; Coddington, J.; Scharff, N. Next-generation Museum genomics: Phylogenetic relationships among palpimanoid spiders using sequence capture techniques (Araneae: Palpimanoidea). Mol. Phylogenet. Evol. 2018, 127, 907–918. [Google Scholar] [CrossRef]
- Kulkarni, S.; Kallal, B.; Wood, H.; Dimitrov, D.; Giribet, G.; Hormiga, G. Interrogating genomic-scale data to resolve recalcitrant nodes in the spider tree of life. Mol. Biol. Evol. 2020, 38, 891–903. [Google Scholar] [CrossRef] [PubMed]
- Faircloth, B.C.; McCormack, J.E.; Crawford, N.G.; Harvey, M.G.; Brumfield, R.T.; Glenn, T.C. Ultraconserved elements anchor thousands of genetic markers spanning multiple evolutionary timescales. Syst. Biol. 2012, 61, 717–726. [Google Scholar] [CrossRef] [PubMed]
- Vachon, M. Études des Caractères Utilisés pour Classer les Familles et les Genres de Scorpions (Arachnides). 1. La Trichobothriotaxie en Arachnologie. Sigles Trichobothriaux et Types de Trichobothriotaxie chez les Scorpions. Bull. Mus. Natl. Hist. Nat. 1974, 104, 857–958. [Google Scholar]
- Francke, O.F. Scorpions of the Genus Diplocentrus from Oaxaca, Mexico (Scorpionida, Diplocentridae). J. Arachnol. 1977, 4, 145–200. [Google Scholar]
- González-Santillán, E.; Prendini, L. Redefinition and Generic Revision of the North American Vaejovid Scorpion Subfamily Syntropinae Kraepelin, 1905, with Descriptions of Six New Genera. Bull. Am. Mus. Nat. Hist. 2013, 382, 1–71. [Google Scholar] [CrossRef]
- Hjelle, J.T. Anatomy and morphology. In The Biology of Scorpions; Polis, G.A., Cloudsley-Thompson, J., Eds.; Stanford University Press: Stanford, CA, USA, 1990; pp. 9–63. [Google Scholar]
- Loria, S.F.; Prendini, L. Homology of the Lateral Eyes of Scorpiones: A Six-Ocellus Model. PLoS ONE 2014, 9, 1–30. [Google Scholar] [CrossRef]
- Volschenk, E.S. A New Technique for Examining Surface Morphosculpture of Scorpions. J. Arachnol. 2005, 33, 820–825. [Google Scholar] [CrossRef]
- Stahnke, H.L. Scorpion Nomenclature and Mensuration. Entomol. News 1970, 81, 297–316. [Google Scholar]
- Starrett, J.; Derkarabetian, S.; Hedin, M.; Bryson, R.W., Jr.; McCormack, J.E.; Faircloth, B.C. High phylogenetic utility of an ultraconserved element probe set designed for Arachnida. Mol. Ecol. Resour. 2017, 17, 812–823. [Google Scholar] [CrossRef]
- Santibáñez-López, C.E.; Aharon, S.; Ballesteros, J.A.; Gainett, G.; Baker, C.M.; González-Santillán, E.; Harvey, M.S.; Hassan, M.K.; Abu-Almaaty, A.H.; Aldeyarbi, S.M.; et al. Phylogenomics of scorpions reveal a co-diversification of scorpion mammalian predators and mammal-specific sodium channel toxins. Syst. Biol. 2022, 71, 1281–1289. [Google Scholar] [CrossRef]
- Sulakhe, S.; Deshpande, S.; Gowande, G.; Dandekar, N.; Ketkar, M. Arboreal Gems: Resurrection of Isometrus sankeriensis Tikader & Bastawade, 1983 and Descriptions of Two New Species of Isometrus Ehrenberg, 1828 (Scorpiones: Buthidae) from the Western Ghats, India. Eur. J. Taxon. 2022, 811, 1–50. [Google Scholar] [CrossRef]
- Folmer, O.; Black, M.; Hoeh, W.; Lutz, R.; Vrijenhoek, R. DNA primers for amplification of mitochondrial cytochrome c oxidase subunit I from diverse metazoan invertebrates. Mol. Mar. Biol. Biotechnol. 1994, 3, 294–299. [Google Scholar] [CrossRef] [PubMed]
- Simon, C.; Frati, F.; Beckenbach, A.; Crespi, B.; Liu, H.; Flook, P. Evolution, weighting, and phylogenetic utility of mitochondrial gene sequences and a compilation of conserved polymerase chain reaction primers. Ann. Entomol. Soc. Am. 1994, 87, 651–701. [Google Scholar] [CrossRef]
- Giribet, G.; Carranza, S.; Baguñà, J.; Riutort, M.; Ribera, C. First molecular evidence for the existence of a Tardigrada + Arthropoda clade. Mol. Biol. Evol. 1996, 13, 76–84. [Google Scholar] [CrossRef]
- Barrett, R.D.H.; Hebert, P.D.N. Identifying spiders through DNA barcodes. Can. J. Zool. 2005, 83, 481–491. [Google Scholar] [CrossRef]
- Benson, D.A.; Cavanaugh, M.; Clark, K.; Karsch-Mizrachi, I.; Lipman, D.J.; Ostell, J.; Sayers, E.W. GenBank. Nucleic Acids Res. 2017, 45, D37–D42. [Google Scholar] [CrossRef]
- Kumar, S.; Stecher, G.; Tamura, K. MEGA7: Molecular Evolutionary Genetics Analysis Version 7.0 for Bigger Datasets. Mol. Biol. Evol. 2016, 33, 1870–1874. [Google Scholar] [CrossRef]
- Edgar, R.C. MUSCLE: Multiple Sequence Alignment with High Accuracy and High Throughput. Nucleic Acids Res. 2004, 32, 1792–1797. [Google Scholar] [CrossRef]
- Nguyen, L.T.; Schmidt, H.A.; Von Haeseler, A.; Minh, B.Q. IQ-TREE: A Fast and Effective Stochastic Algorithm for Estimating Maximum-Likelihood Phylogenies. Mol. Biol. Evol. 2015, 32, 268–274. [Google Scholar] [CrossRef]
- Edler, D.; Klein, J.; Antonelli, A.; Silvestro, D. raxmlGUI 2.0: A graphical interface and toolkit for phylogenetic analyses using RAxML. Methods Ecol. Evol. 2021, 12, 373–377. [Google Scholar] [CrossRef]
- Kalyaanamoorthy, S.; Minh, B.Q.; Wong, T.K.; Von Haeseler, A.; Jermiin, L.S. ModelFinder: Fast Model Selection for Accurate Phylogenetic Estimates. Nat. Methods 2017, 14, 587–589. [Google Scholar] [CrossRef] [PubMed]
- Hoang, D.T.; Chernomor, O.; Von Haeseler, A.; Minh, B.Q.; Vinh, L.S. UFBoot2: Improving the Ultrafast Bootstrap Approximation. Mol. Biol. Evol. 2018, 35, 518–522. [Google Scholar] [CrossRef] [PubMed]
- Minh, B.Q.; Schmidt, H.A.; Chernomor, O.; Schrempf, D.; Woodhams, M.D.; Von Haeseler, A.; Lanfear, R. IQ-TREE 2: New Models and Efficient Methods for Phylogenetic Inference in the Genomic Era. Mol. Biol. Evol. 2020, 37, 1530–1534. [Google Scholar] [CrossRef] [PubMed]
- Lanfear, R.; Calcott, B.; Ho, S.Y.W.; Guindon, S. PartitionFinder: Combined Selection of Partitioning Schemes and Substitution Models for Phylogenetic Analyses. Mol. Biol. Evol. 2012, 29, 1695–1701. [Google Scholar] [CrossRef]
- Schwarz, G. Estimating the Dimension of a Model. Ann. Stat. 1978, 6, 461–464. [Google Scholar] [CrossRef]
- Suchan, T.; Pitteloud, C.; Gerasimova, N.S.; Kostikova, A.; Schmid, S.; Arrigo, N.; Pajkovic, M.; Ronikier, M.; Alvarez, N. Hybridization Capture Using RAD Probes (hyRAD), a New Tool for Performing Genomic Analyses on Collection Specimens. PLoS ONE 2016, 11, e0151651. [Google Scholar] [CrossRef]
- Meyer, M.; Kircher, M. Illumina Sequencing Library Preparation for Highly Multiplexed Target Capture and Sequencing. Cold Spring Harb. Protoc. 2010, 2010, pdb-prot5448. [Google Scholar] [CrossRef]
- Faircloth, B.C. Identifying conserved genomic elements and designing universal bait sets to enrich them. Methods Ecol. Evol. 2017, 8, 1103–1112. [Google Scholar] [CrossRef]
- Grabherr, M.G.; Haas, B.J.; Yassour, M.; Levin, J.Z.; Thompson, D.A.; Amit, I.; Adiconis, X. Trinity: Reconstructing a Full-Length Transcriptome Without a Genome from RNA-Seq Data. Nat. Biotechnol. 2011, 29, 644. [Google Scholar] [CrossRef]
- Bankevich, A.; Nurk, S.; Antipov, D.; Gurevich, A.A.; Dvorkin, M.; Kulikov, A.S.; Lesin, V.M.; Nikolenko, S.I.; Pham, S.; Prjibelski, A.D.; et al. SPAdes: A new genome assembly algorithm and its applications to single-cell sequencing. J. Comput. Biol. 2012, 19, 455–477. [Google Scholar] [CrossRef]
- Faircloth, B.C. PHYLUCE is a software package for the analysis of conserved genomic loci. Bioinformatics 2016, 32, 786–788. [Google Scholar] [CrossRef] [PubMed]
- Katoh, K.; Standley, D.M. MAFFT Multiple Sequence Alignment Software Version 7: Improvements in Performance and Usability. Mol. Biol. Evol. 2013, 30, 772–780. [Google Scholar] [CrossRef]
- Castresana, J. Selection of Conserved Blocks from Multiple Alignments for Their Use in Phylogenetic Analysis. Mol. Biol. Evol. 2000, 17, 540–552. [Google Scholar] [CrossRef] [PubMed]
- Talavera, G.; Casteresana, J. Improvement of phylogenies after removing divergent and ambiguously aligned blocks from protein sequence alignments. Syst. Biol. 2007, 56, 564–577. [Google Scholar] [CrossRef] [PubMed]
- Stamatakis, A.; Hoover, P.; Rougemont, J. A Rapid Bootstrap Algorithm for the RAxML Web Servers. Syst. Biol. 2008, 57, 758–771. [Google Scholar] [CrossRef]
- Bello, E.; Chen, Y.; Alleyne, M. Staying Dry and Clean: An Insect’s Guide to Hydrophobicity. Insects 2023, 14, 42. [Google Scholar] [CrossRef]
- Wenzel, R.N. Resistance of Solid Surfaces to Wetting by Water. Ind. Eng. Chem. 1936, 28, 988–994. [Google Scholar] [CrossRef]
- Cassie, A.B.D.; Baxter, S. Wettability of Porous Surfaces. Trans. Faraday Soc. 1944, 40, 546–551. [Google Scholar] [CrossRef]
- Mani, M.S. (Ed.) Ecology and Biogeography in India, 3rd ed.; Dr. W. Junk, B.V., Publishers: The Hague, The Netherlands, 1974; pp. 154–196. [Google Scholar]
- Aitchison, J.C.; Ali, J.R.; Davis, A.M. When and where did India and Asia collide? J. Geophys. Res. Solid Earth 2007, 112, B05423. [Google Scholar] [CrossRef]
- Conti, E.; Eriksson, T.; Schönenberger, J.; Sytsma, K.J.; Baum, D.A. Early Tertiary Out-of-India dispersal of Crypteroniaceae: Evidence from phylogeny and molecular dating. Evolution 2002, 56, 1931–1942. [Google Scholar]
- Bossuyt, F.; Brown, R.M.; Hillis, D.M.; Cannatella, D.C.; Milinkovitch, M.C. Phylogeny and biogeography of a cosmopolitan frog radiation: Late Cretaceous diversification resulted in continent-scale endemism in the family Ranidae. Syst. Biol. 2006, 55, 579–594. [Google Scholar] [CrossRef] [PubMed]
- Joshi, J.; Karanth, P. Did southern Western Ghats of peninsular India serve as refugia for its endemic biota during the Cretaceous volcanism? Ecol. Evol. 2013, 3, 3275–3282. [Google Scholar] [CrossRef] [PubMed]
- Loria, S.F.; Prendini, L. Out of India, thrice: Diversification of Asian forest scorpions reveals three colonizations of Southeast Asia. Sci. Rep. 2020, 10, 22301. [Google Scholar] [CrossRef] [PubMed]
- Bharti, D.K.; Edgecombe, G.D.; Karanth, K.P.; Joshi, J. Spatial patterns of phylogenetic diversity and endemism in the Western Ghats, India: A case study using ancient predatory arthropods. Ecol. Evol. 2021, 11, 16499–16513. [Google Scholar] [CrossRef]

Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Joshi, M.; Deshpande, S.; Ukale, S.; Gowande, G.; Bilat, J.; Kovařík, F.; Mottaz, H.; Šťáhlavský, F.; Bastawade, D.; Monod, L.; et al. Systematic Revision of the Genus Charmus Karsch, 1879 (Scorpiones: Buthidae), and Assessment of Its Phylogenetic Position Within Buthidae C. L. Koch, 1837 Using Ultraconserved Elements. Diversity 2025, 17, 354. https://doi.org/10.3390/d17050354
Joshi M, Deshpande S, Ukale S, Gowande G, Bilat J, Kovařík F, Mottaz H, Šťáhlavský F, Bastawade D, Monod L, et al. Systematic Revision of the Genus Charmus Karsch, 1879 (Scorpiones: Buthidae), and Assessment of Its Phylogenetic Position Within Buthidae C. L. Koch, 1837 Using Ultraconserved Elements. Diversity. 2025; 17(5):354. https://doi.org/10.3390/d17050354
Chicago/Turabian StyleJoshi, Mihir, Shubhankar Deshpande, Sajiri Ukale, Gaurang Gowande, Julia Bilat, František Kovařík, Hélène Mottaz, František Šťáhlavský, Deshabhushan Bastawade, Lionel Monod, and et al. 2025. "Systematic Revision of the Genus Charmus Karsch, 1879 (Scorpiones: Buthidae), and Assessment of Its Phylogenetic Position Within Buthidae C. L. Koch, 1837 Using Ultraconserved Elements" Diversity 17, no. 5: 354. https://doi.org/10.3390/d17050354
APA StyleJoshi, M., Deshpande, S., Ukale, S., Gowande, G., Bilat, J., Kovařík, F., Mottaz, H., Šťáhlavský, F., Bastawade, D., Monod, L., & Sulakhe, S. (2025). Systematic Revision of the Genus Charmus Karsch, 1879 (Scorpiones: Buthidae), and Assessment of Its Phylogenetic Position Within Buthidae C. L. Koch, 1837 Using Ultraconserved Elements. Diversity, 17(5), 354. https://doi.org/10.3390/d17050354