
Exploring the Orchid Flora of Montenegro: Ten Newly Identified Taxa



Abstract
:1. Introduction
2. Materials and Methods
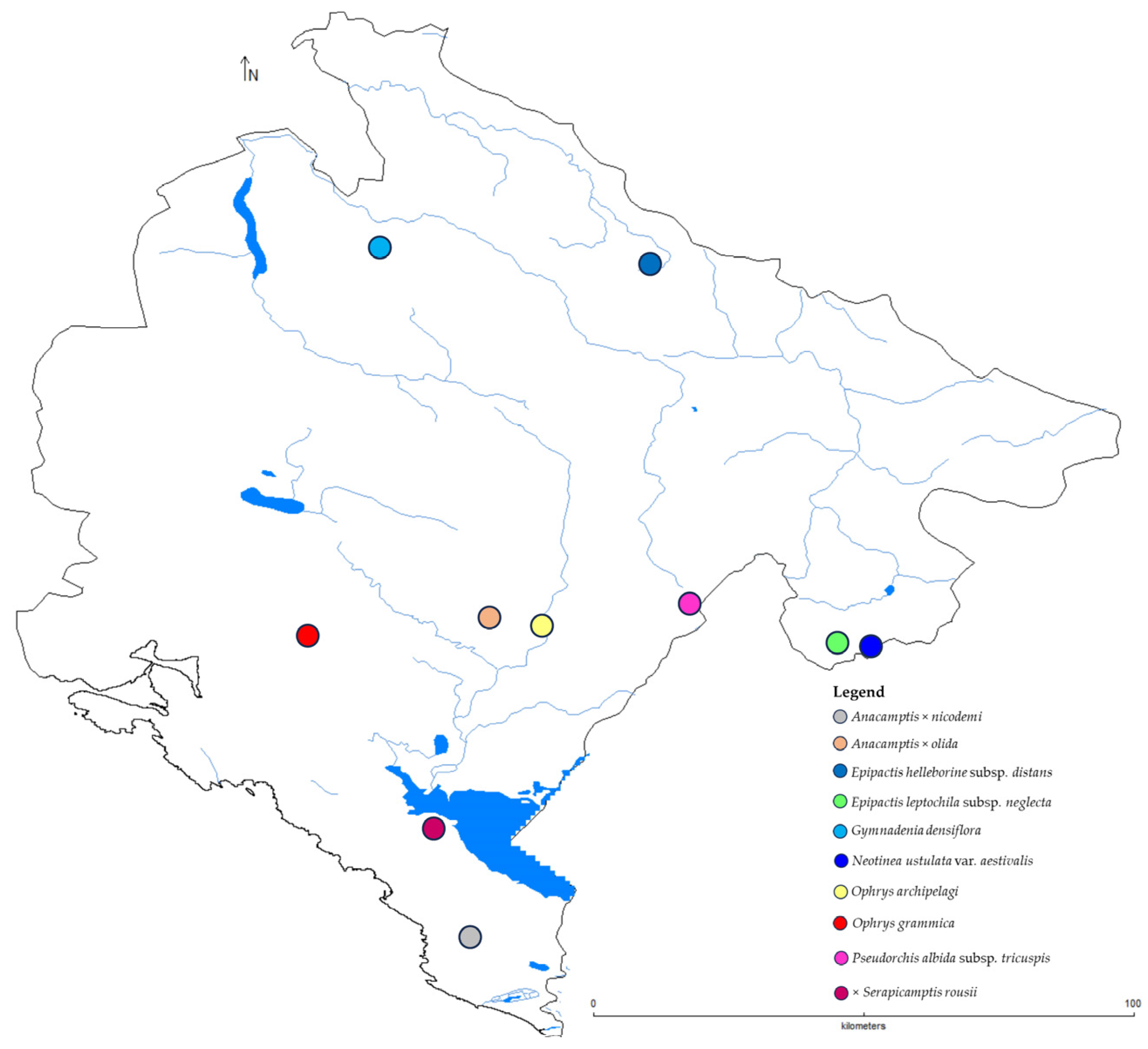
3. Results
3.1. Species New to Montenegro
- Gymnadenia densiflora (Wahlenb.) A. Dietr., Allg. Gartenzeit. 7(22): 170 (1839)
- Ophrys archipelagi Gölz and H. R. Reinhard, Mitteilungsbl. Arbeitskreis Heimische Orchid. Baden-Württemberg 18(4): 731 (1986)
- Ophrys grammica (B. Willing and E. Willing) Devillers-Tersch. and Devillers, Naturalistes Belges 72(3): 101 (1991)
3.2. Subspecies New to Montenegro
- Epipactis helleborine (L.) Crantz subsp. distans (Arv.-Touv.) R. Engel and P. Quentin, Orchidophile (Asnières) 124: 205 (1996)
- Epipactis leptochila (Godfery) Godfery subsp. neglecta Kümpel, Mitt. Arbeitskreises Heimische Orchideen 11: 30 (1982)
- Pseudorchis albida (L.) Á. Löve and D. Löve subsp. tricuspis (Beck) E. Klein, Phyton 40(1): 143 (2000)
3.3. Variety New to Montenegro
- Neotinea ustulata (L.) R. M. Bateman, Pridgeon and M. W. Chase var. aestivalis (Kümpel) Tali, M. F. Fay and R. M. Bateman, Biol. J. Linn. Soc. 87(1): 23 (2006)
3.4. Hybrids New to Montenegro
- Anacamptis ×nicodemi (Cirillo ex Ten.) B. Bock, Bull. Soc. Bot. Centre-Ouest 42: 267 (2012)
- Anacamptis ×olida (Bréb.) H. Kretzschmar, Eccarius and H. Dietr., Orchid Gen. Anacamptis Orchis Neotinea 428 (2007)
- ×Serapicamptis rousii (Dupuy) J. M. H. Shaw, Orchid Rev. Suppl., 113 (1264): 20 (2005)
4. Discussion
4.1. New Species to Montenegro’s Flora
- Gymnadenia densiflora
- Ophrys archipelagi
- Ophrys grammica
4.2. New Subspecies to Montenegroˈs Flora
- Epipactis helleborine subsp. distans
- Epipactis leptochila subsp. neglecta
- Pseudorchis albida subsp. tricuspis
4.3. New Variety to Montenegroˈs Flora
- Neotinea ustulata var. aestivalis
4.4. New Hybrids to Montenegroˈs Flora
- Anacamptis ×nicodemi
- Anacamptis ×olida
- ×Serapicamptis rousii
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Appendix A

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Taxa | Herbarium Labels |
|---|---|
| Gymnadenia densiflora | Durmitor Mt., Jablan Lake, N43.16753°, E19.06601°, MGRS 34TCN42, 1835 m asl, subalpine meadow, limestones, 10 July 2021, coll. et det. B. Radak (BUNS 22242) |
| Ophrys archipelagi | Morača Gorge, Duga, near the bridge across the Morača river, N42.54113°, E19.33718°, MGRS 34TCN63, 107 m asl, highly degraded Quercus trojana woods, limestone, 11 April 2021, coll. B. Radak, det. B. Radak, J. Peškanov (BUNS 22238) |
| Ophrys grammica | Kopitnik Mt., Velestovo, near the road to Čevo, N42.52758°, E18.94766°, MGRS 34TCN31, highly degraded Quercus pubescens woods, limestone, 30 April 2022, coll. B. Radak, det. B. Radak, J. Peškanov (BUNS 22239) |
| Epipactis helleborine subsp. distans | Stožer Mt., Ponikvice, N43.14202°, E19.51986°, MGRS 34TCN79, 1226 m asl, mountain hay meadow, limestones, 27 July 2022, coll. et det. B. Radak (BUNS 22246) |
| Epipactis leptochila subsp. neglecta | Prokletije Mts., Vusanje, Savino Oko, N42.51170°, E19.83481°, MGRS 34TDN04, 1036 m asl, subalpine Fagus sylvatica stand, limestones and dolomitic limestones, 22 July 2022, coll. et det. B. Radak (BUNS 22245) |
| Pseudorchis albida subsp. tricuspis | Kučke Mts., Katun Ljakovića, N42.58190°, E19.59019°, MGRS 34TCN84, 1764 m asl., limestones, 04 July 2021, coll. J. Lazarevic, det. B. Radak (BUNS 22244) |
| Neotinea ustulata var. aestivalis | Prokletije Mts., Ćaf Bora, MGRS 34TDN09, meadows, 23 July 2014, coll. S. Ivković, L. Horvat, det. B. Radak (BUNS 22243) |
| Anacamptis ×nicodemi | Krute-Velje Selo, N42.02357°, E19.21719°, MGRS 34TCM52, 205 m asl, pasture, limestones, 07 April 2021, coll. B. Radak, J. Beloica, det. B. Radak, J. Peškanov (BUNS 22240) |
| Anacamptis ×olida | Kopiljsko polje, N42.55350°, E19.24961°, MGRS 34TCN56, 601 m asl, rocky grasslands, limestones, 16 May 2021, coll. B. Bokić, M. Rat, det. B. Radak, J. Peškanov (BUNS 22241) |
| ×Serapicamptis rousii | Skadar Lake, Gornja Seoca-Marstijepovići, N42.20085°, E19.16018°, MGRS 34TCM48, 408 m asl, road cracks, limestones, 16 May 2021, coll. B. Radak, det. B. Radak, J. Peškanov (BUNS 22247) |
References
- Stevanović, V.; Vasić, V. Biodiverzitet Jugoslavije sa Pregledom Vrsta od Međunarodnog Značaja; Ecolibri, Biološki Fakultet: Belgrade, Serbia, 1995. [Google Scholar]
- Stešević, D.; Caković, D. Catalogue of Vascular Flora of Montenegro; Karaman, G., Ed.; Crnogorska Akademija Nauka i Umjetnosti: Podgorica, Montenegro, 2013; Volume 1. [Google Scholar]
- Pulević, V. Botanists and Montenegro, Special ed.; Natural History Museum of Montenegro: Podgorica, Montenegro, 2006. [Google Scholar]
- Rohlena, J. Conspectus florae montenegriane. Preslia 1941, 20–21, 3–506. [Google Scholar]
- Caković, D.; Stešević, D. Catalogue of Vascular Flora of Montenegro; Karaman, G., Ed.; Crnogorska Akademija Nauka i Umjetnosti: Podgorica, Montenegro, 2021; Volume 2. [Google Scholar]
- Horvatić, S. Analitička Flora Jugoslavije 1; Institut za Botaniku Sveučilišta u Zagrebu: Zagreb, Yugoslavia, 1967. [Google Scholar]
- Trinajstić, I. Analitička Flora Jugoslavije 1, 2; Institut za Botaniku Sveučilišta u Zagrebu: Zagreb, Yugoslavia, 1973. [Google Scholar]
- Pulević, V. Građa za Vaskularnu Floru Crne Gore: Dopuna “Conspectus Florae Montenegrinae”; Republički Zavod za Zaštitu Prirode Crne Gore: Podgorica, Montenegro, 2005. [Google Scholar]
- Stešević, D.; Caković, D.; Snežana, V.; Biberdžić, V. Contribution to the vascular flora of Montenegro (Supplementum to the Material for vascular flora of Montenegro). Nat. Montenegrina 2008, 7, 463–480. [Google Scholar]
- Christenhusz, M.J.M.; Byng, J.W. The number of known plants species in the world and its annual increase. Phytotaxa 2016, 261, 201–217. [Google Scholar] [CrossRef]
- Pridgeon, A.M.; Cribb, P.; Chase, M.W.; Rasmussen, F.N. Genera Orchidacearum: Volume 1: Apostasioideae and Cypripedioideae; Oxford University Press: Oxford, UK, 2009; Volume 1. [Google Scholar]
- Delforge, P. Orchids of Europe, North Africa and the Middle East; A&C Black Publishers Ltd.: London, UK, 2006. [Google Scholar]
- Turco, A.; Albano, A.; Medagli, P.; D’Emerico, S.; Wagensommer, R.P. Orchidaceae in Puglia (Italy): Consistency, Distribution, and Conservation. Plants 2023, 12, 2223. [Google Scholar] [CrossRef]
- Fay, M. British and Irish Orchids in a Changing World. Curtis Bot. Mag. 2015, 32, 3–23. [Google Scholar] [CrossRef]
- Kretzschmar, H.; Eccarius, W.; Dietrich, H. The Orchid Genera Anacamptis, Orchis and Neotinea: Phylogeny, Taxonomy, Morphology, Biology, Distribution, Ecology and Hybridisation; EchinoMedia: Burgel, Germany, 2007. [Google Scholar]
- Antonopoulos, Z.; Tsiftsis, S. Atlas of the Greek Orchids; Mediterraneo Editions: Rethymno, Greece, 2017. [Google Scholar]
- CITES Checklist of CITES Species. Available online: https://checklist.cites.org/ (accessed on 12 January 2025).
- Radak, B.Đ.; Vlku, A.Z.; Peškanov, J.M.; Matevski, V.S.; Anačkov, G.T. Morphological characterization of three natural hybrid orchid taxa, new for Serbia, Montenegro and North Macedonia. Arch. Biol. Sci. 2019, 71, 596–607. [Google Scholar] [CrossRef]
- Molnár, A.; Óvári, M.; Kis, S.; Bak, H.; Fekete, R. Data to the distribution and ecology of Ophrys tetraloniae WP Teschner (Orchidaceae). Ecol. Montenegrina 2025, 81, 37–42. [Google Scholar] [CrossRef]
- Verhart, F. Ophrys helenae, a new species for the flora of Montenegro and its distribution on the Balkan peninsula. J. Eur. Orch. 2024, 56, 209–218. [Google Scholar]
- Radak, B.; Bokić, B.; Miljković, P.; Peškanov, J.; Vlku, A.; Beloica, J.; Anačkov, G. Towards the checklist of the orchids of Montenegro. In Proceedings of the 6th Congress of Ecologists of the Republic of North Macedonia, Ohrid, North Macedonia, 15–18 October 2022; p. 94. [Google Scholar]
- Arvet–Touvet, C. Essai Sur L’espèce et Les Variétés Principalement Dans les Plantes; Prudhomme: Grenoble, France, 1872. [Google Scholar]
- Vlčko, J.; Dítě, D.; Kolník, M. Orchids of Slovakia; ZO SZOPK Orchidea: Zvolen, Slovakia, 2003. [Google Scholar]
- Kreutz, C.A.J.; Fateryga, A.V. Two taxa of the genus Epipactis Zinn (Orchidaceae) new for the flora of Ukraine. Ukr. Bot. J. 2012, 69, 713–716. [Google Scholar]
- Dolinar, B. Kukavičevke v Sloveniji; Pipinova Knjiga: Podsmreka, Slovenia, 2015. [Google Scholar]
- Krajnc, A.U.; Ivanuš, A.; Luthar, Z.; Lipovšek, M. Morphological Variability and Taxonomic Concepts of Broad-Leaved Helleborine Ingroup Epipactis helleborine (L.) Crantz. Folia Biol. Geol. 2020, 61, 97–125. [Google Scholar] [CrossRef]
- Šabanović, E.; Djordjević, V.; Milanović, Đ.; Boškailo, A.; Šarić, Š.; Huseinović, S.; Randjelović, V. Checklist of the Orchidaceae of Bosnia and Herzegovina. Phyton 2021, 61, 83–95. [Google Scholar] [CrossRef]
- Đorđević, V.; Jakovljević, K.; Stevanović, V. Three taxa of Epipactis (Orchidaceae-Epidendroideae) new for the flora of Serbia. Phyton 2016, 56, 77–89. [Google Scholar] [CrossRef]
- Bartolucci, F.; Peruzzi, L.; Galasso, G.; Alessandrini, A.; Ardenghi, N.M.G.; Bacchetta, G.; Banfi, E.; Barberis, G.; Bernardo, L.; Bouvet, D. A second update to the checklist of the vascular flora native to Italy. Plant Biosyst. Int. J. Deal. Asp. Plant Biol. 2024, 158, 219–296. [Google Scholar] [CrossRef]
- Kühn, R.; Pedersen, H.; Cribb, P. Field Guide to the Orchids of Europe and the Mediterranean; Kew Publishing: London, UK, 2019. [Google Scholar]
- Sramkó, G.; Paun, O.; Brandrud, M.K.; Laczkó, L.; Molnár, A.; Bateman, R.M. Iterative allogamy–autogamy transitions drive actual and incipient speciation during the ongoing evolutionary radiation within the orchid genus Epipactis (Orchidaceae). Ann. Bot. 2019, 124, 481–497. [Google Scholar] [CrossRef]
- Bateman, R.M. Implications of next-generation sequencing for the systematics and evolution of the terrestrial orchid genus Epipactis, with particular reference to the British Isles. Kew Bull. 2020, 75, 4. [Google Scholar] [CrossRef]
- Łobas, Z.; Khapugin, A.; Żołubak, E.; Jakubska-Busse, A. The Epipactis helleborine group (Orchidaceae): An overview of recent taxonomic changes, with an updated list of currently accepted taxa. Plants 2021, 10, 1839. [Google Scholar] [CrossRef]
- Flora Croatica Database. Available online: https://hirc.botanic.hr/fcd (accessed on 10 January 2025).
- Đorđević, V. Flora Orhideja (Orchidaceae) Zapadne Srbije; Srpska Akademija Nauka i Umetnosti: Belgrade, Serbia, 2021. [Google Scholar]
- Đorđević, V. Orhideje Nacionalnog Parka Tara; JP Nacionalni Park Tara: Bajina Bašta, Serbia, 2022. [Google Scholar]
- Plants of the World Online. Available online: https://powo.science.kew.org/ (accessed on 10 January 2025).
- Kranjčev, R. Hrvatske Orhideje; Agencija za Komercijalnu Djelatnost: Zagreb, Croatia, 2005. [Google Scholar]
- Molnár, V.A. Atlas of Hungarian Orchids; Kossuth Kiadó: Budapest, Hungary, 2011. [Google Scholar]
- Krajnc, A.U.; Lipovšek, M.; Luthar, Z.; Ivanuš, A.; Miljuš, S.; Bohanec, B.; Šiško, M. Taxonomic analysis of certain taxa of Epipactis in Slovenia. Perspect. Plant Ecol. EVolume Syst. 2022, 56, 125674. [Google Scholar] [CrossRef]
- Wahlenberg, G. Utkast till Gottlands flora. Kongl. Vetensk. Acad. Nya Handl. 1806, 27, 68. [Google Scholar]
- Gustafsson, S.; Lönn, M. Genetic differentiation and habitat preference of flowering-time variants within Gymnadenia conopsea. Heredity 2003, 91, 284–292. [Google Scholar] [CrossRef]
- Lang, D. Britain’s Orchids: A Guide to the Identification and Ecology of the Wild Orchids of Britain and Ireland; WILDGuides Ltd.: Hampshire, UK, 2004. [Google Scholar]
- Dietrich, A. Flora des Königreichs Preussen. Allgemaine Gartenzeitung 1839, 7, 170. [Google Scholar]
- Bateman, R.; Pridgeon, A.M.; Chase, M. Phylogenetics of Subtribe Orchidinae (Orchidoideae, Orchidaceae) Based on Nuclear ITS Sequences. 2. Infrageneric Relationships and Reclassification to Achieve Monophyly of Orchis Sensu Stricto. Lindleyana 1997, 12, 113–141. [Google Scholar]
- Bateman, R.M.; Hollingsworth, P.M.; Preston, J.; Yi-Bo, L.U.O.; Pridgeon, A.M.; Chase, M.W. Molecular phylogenetics and evolution of Orchidinae and selected Habenariinae (Orchidaceae). Bot. J. Linn. Soc. 2003, 142, 1–40. [Google Scholar] [CrossRef]
- Marhold, K.; Jongepierová, I.; Krahulcová, A.; Kucera, J. Morphological and karyological differentiation of Gymnadenia densiflora and G. conopsea in the Czech Republic and Slovakia. Preslia 2005, 77, 159–176. [Google Scholar]
- Bateman, R.M.; Rudall, P.J.; Denholm, I. In situ morphometric survey elucidates the evolutionary systematics of the orchid genus Gymnadenia in the British Isles. Syst. Biodivers. 2021, 19, 571–600. [Google Scholar] [CrossRef]
- Kümpel, H. Über eine spätblühende Orchis ustulata-Sippe. Haussknechtia 1988, 4, 23–24. [Google Scholar]
- Kümpel, H.; Mrkvicka, A.C. Untersuchungen Zur Abtrennung Der Orchis ustulata L. subsp. Aestivalis (Kümpel) Kümpel & Mrkvicka. Jahresber. Naturwisschaften Ver. Wuppertal 1990, 22, 306–324. [Google Scholar]
- Molnár, A. Magyarország Orchideáinak Atlasza; Kossuth Kiadó: Budapest, Hungary, 2012. [Google Scholar]
- Martel, C.; Rakosy, D.; Dötterl, S.; Johnson, S.D.; Ayasse, M.; Paulus, H.F.; Nilsson, L.A.; Mejlon, H.; Jersáková, J. Specialization for tachinid fly pollination in the phenologically divergent varieties of the orchid Neotinea ustulata. Front. Ecol. EVolume 2021, 9, 659176. [Google Scholar] [CrossRef]
- Trávníček, P.; Chumová, Z.; Záveská, E.; Hanzlíčková, J.; Kupková, L.; Kučera, J.; Gbúrová Štubňová, E.; Rejlová, L.; Mandáková, T.; Ponert, J. Integrative study of genotypic and phenotypic diversity in the Eurasian orchid genus Neotinea. Front. Plant Sci. 2021, 12, 734240. [Google Scholar] [CrossRef]
- Paulus, H.F. Pollination biology of two phenological forms of Neotinea ustulata (Orchidaceae) in Austria, with field experiments to clarify the biological significance of the plants’“burnt tips”. Acta ZooBot Austria 2022, 158, 149–175. [Google Scholar]
- Haraštová-Sobotková, M.; Jersáková, J.; Kindlmann, P.; Čurn, L. Morphometric and genetic divergence among populations of Neotinea ustulata (Orchidaceae) with different flowering phenologies. Folia Geobot. 2005, 40, 385–405. [Google Scholar] [CrossRef]
- Tali, K.; Fay, M.F.; Bateman, R.M. Little genetic differentiation across Europe between early-flowering and late-flowering populations of the rapidly declining orchid Neotinea ustulata. Biol. J. Linn. Soc. 2006, 87, 13–25. [Google Scholar] [CrossRef]
- Nikolić, T.; Topić, J. Crvena Knjiga Vaskularne Flore Hrvatske; Ministarstvo Kulture, Državni Zavod za Zaštitu Prirode: Zagreb, Croatia, 2005.
- Nikolić, T.; Milović, M.; Bogdanović, S.; Jasprica, N. Endemi u Hrvatskoj Flori; Alfa: Zagreb, Croatia, 2015. [Google Scholar]
- Pignatti, S.; Guarino, R.; La Rosa, M. Flora d’Italia, 2nd ed.; Edagricole: Bologna, Italy, 2017; Volume 1. [Google Scholar]
- Pedersen, H.Æ.; Faurholdt, N. Ophrys: The Bee Orchids of Europe; Royal Botanic Gardens Kew: London, UK, 2007. [Google Scholar]
- Willing, B.; Willing, E. Beitrag zur Orchideenflora NW Griechenlands—Kartierungsergebnisse 1984/85. Mitt. Bl. Arbeitskr. Heim. Orch. Baden-Württ. 1985, 17, 508–628. [Google Scholar]
- Antonopoulos, Z. The Bee Orchids of Greece. The Genus Ophrys; Mediterraneo Editions: Rethymno, Greece, 2009. [Google Scholar]
- Niketic, M.; Tomović, G.; Perić, R.; Zlatković, B.; Anačkov, G.; Djordjević, V.; Jogan, N.; Radak, B.; Duraki, Š.; Stanković, M.; et al. Material on the Annotated Checklist of Vascular Flora of Serbia. Nomenclatural, taxonomic and floristic notes I. Bull. Nat. Hist. Mus. Belgr. 2018, 11, 101–180. [Google Scholar] [CrossRef]
- Peškanov, J.; Radak, B.; Vlku, A.; Anačkov, G. Uskrsnuće Ophrys grammica—Morfološka karakterizacija spornog taksona orhideja. In Proceedings of the 2. Simpozijum “Treći Vek Botanike u Vojvodini”, Novi Sad, Serbia, 6 October 2023; Matica Srpska: Novi Sad, Serbia, 2023; pp. 66–68. [Google Scholar]
- Landwehr, J. Wilde Orchideeën van Europa; Vereniging tot Behoud van Natuurmonumenten in Nederland: Amsterdam, The Netherlands, 1977. [Google Scholar]
- Jersáková, J.; Malinová, T.; Jerábková, K.; Dötterl, S. Biological Flora of the British Isles: Pseudorchis albida (L.) Á. & D. Löve. J. Ecol. 2011, 99, 1282–1298. [Google Scholar] [CrossRef]
- Harrap, S. A Pocket Guide to the Orchids of Britain and Ireland; Bloomsbury Publishing: London, UK, 2016. [Google Scholar]
- Klein, E. Pseudorchis albida subsp. tricuspis (Beck) Klein stat. nov.; eine weitgehend ubersehene, calcicole, alpisch-boreale Sippe (Orchidaceae-Orchideae). Phyton 2000, 40, 141–159. [Google Scholar]
- Dimopoulos, P.; Raus, T.; Bergmeier, E.; Constantinidis, T.; Iatrou, G.; Kokkini, S.; Strid, A.; Tzanoudakis, D. Vascular plants of Greece: An annotated checklist. Supplement. Willdenowia 2016, 46, 301–347. [Google Scholar] [CrossRef]
- Anghelescu, N.E.D.G.; Kertész, H.; Constantin, N.; Simon-Gruița, A.; Duță Cornescu, G.; Pojoga, M.D.; Georgescu, M.I.; Petra, S.A.; Toma, F. New intergeneric orchid hybrid found in Romania× Pseudorhiza nieschalkii (Senghas) PF Hunt nothosubsp. siculorum H. Kertész & N. Anghelescu, 2020. PLoS ONE 2021, 16, e0241733. [Google Scholar] [CrossRef]
- Bateman, R.M.; Rudall, P.J.; Denholm, I. Morphometric comparison of British Pseudorchis albida with Icelandic P. straminea (orchidaceae: Orchidinae). New J. Bot. 2017, 7, 78–93. [Google Scholar] [CrossRef]
- Index Herbariorum. Available online: https://sweetgum.nybg.org/science/ih/ (accessed on 10 January 2025).
- Marković, M. The Geographical Areas of the Socialist Federal Republic of Yugoslavia; Zavod za Udžbenike i Nastavna Sredstva Srbije: Belgrade, Serbia, 1970. [Google Scholar]
- Chytrý, M.; Tichý, L.; Hennekens, S.M.; Knollová, I.; Janssen, J.A.M.; Rodwell, J.S.; Peterka, T.; Marcenò, C.; Landucci, F.; Danihelka, J. EUNIS Habitat Classification: Expert system, characteristic species combinations and distribution maps of European habitats. Appl. Veg. Sci. 2020, 23, 648–675. [Google Scholar] [CrossRef]
- Mirković, M.; Živaljević, M.; Đokić, V.; Perović, Z.; Kalezić, M.; Pajović, M. Geological Map of Southern Socialist Republic of Montenegro, 1:200,000; The Republic Self-Managing Community of Interest for Geological Exploration of Montenegro: Titograd, Yugoslavia, 1985. [Google Scholar]
- Jacquemyn, H.; Hutchings, M.J. Biological Flora of the British Isles: Spiranthes spiralis (L.) Chevall. J. Ecol. 2010, 98, 1253–1267. [Google Scholar] [CrossRef]
- Radak, B.; Hristovski, S.; Matevski, V.S.; Anačkov, G. New orchid taxa for North Macedonia. In Proceedings of the 6th Congress of Ecologists of the Republic of North Macedonia, Ohrid, North Macedonia, 15–18 October 2022; p. 82. [Google Scholar]
- Radak, B.; Vlku, A.; Peškanov, J.; Bokić, B.; Anačkov, G. Orhideje u Srbiji 2: Prilozi Kritičkoj Listi Vrsta Vaskularne Flore Srbije 1. In Proceedings of the 2. Simpozijum “Treći Vek Botanike u Vojvodini”, Novi Sad, Serbia, 6 October 2023; pp. 72–73. [Google Scholar]
- Paucâ, A. Epipactis Zinn. In Flora Reipublicae Socialisticae Romania XII; Nyárády, I., Ed.; Academia Reipublicae Socialisticae Romania: Bucharest, Romania, 1972; pp. 757–761. [Google Scholar]
- Stornov, N. Салепс, Орхидеје—Orchidaceae Lindl. In Flora na Narodna Republika Bulgariia II; Jordanov, D., Velcev, V., Eds.; Academiae Scientiarum Bulgaricae: Sofia, Bulgaria, 1964; pp. 349–399. [Google Scholar]
- Dimitrov, D. Conspectus of the Bulgarian Vascular Flora; Bulgarian-Swiss Biodiversity Conservation Programme: Sofia, Bulgaria, 2002. [Google Scholar]
- Kull, T.; Selgis, U.; Pecina, M.V.; Metsare, M.; Ilves, A.; Tali, K.; Sepp, K.; Kull, K.; Shefferson, R.P. Factors influencing IUCN threat levels to orchids across Europe on the basis of national red lists. Ecol. EVolume 2016, 6, 6245–6265. [Google Scholar] [CrossRef]
- Gölz, P.; Reinhard, H.R. Orchideen in Jugoslawien. Mitt. Bl. Arbeitskr. Heim. Orch. 1986, 18, 689–827. [Google Scholar]
- Jeričević, M.; Jeričević, N. Orhideje na Otoku Korčuli; Adriatika—Društvo za Istraživanje i Očuvanje Prirodoslovne Raznolikosti Mediterana: Korčula, Croatia, 2024. [Google Scholar]
- Milanović, Đ.; Caković, D.; Hadžiablahović, S.; Vuksanović, S.; Mačić, V.; Stešević, D.; Stanišić-Vujačić, M.; Biberdžić, V.; Lakušić, D. Priručnik za Identifikaciju Tipova Staništa Crne Gore od Značaja za Evropsku Uniju sa Obrađenim Glavnim Indikatorskim Vrstama; Agencija za Zaštitu Životne Sredine Crne Gore; Univerzitet u Banjoj Luci: Banja Luka, Bosnia and Herzegovina, 2021. [Google Scholar]
- Orsenigo, S.; Fenu, G.; Gargano, D.; Montagnani, C.; Abeli, T.; Alessandrini, A.; Bacchetta, G.; Bartolucci, F.; Carta, A.; Castello, M. Red list of threatened vascular plants in Italy. Plant Biosyst. Int. J. Deal. Asp. Plant Biol. 2021, 155, 310–335. [Google Scholar] [CrossRef]
- Conti, F.; Manzi, A.; Pedrotti, F. Liste Rosse Regionali Delle Piante d’italia.; WWF Italia: Camerino, Italy, 1997. [Google Scholar]
- Pravilnik o Izmjenama i Dopunama Pravilnika o Uređivanju Šuma; Narodne Novine: Zagreb, Croatia, 2008; Volume 73, pp. 1–20.
- Peškanov, J.; Radak, B.; Vlku, A.; Anačkov, G. Ophrys grammica—nova vrsta orhideja za floru Srbije. In Proceedings of the 2. simpozijum “Treći Vek Botanike u Vojvodini”, Novi Sad, Serbia, 6 October 2023; Matica Srpska: Novi Sad, Serbia, 2023; pp. 68–70. [Google Scholar]
- AHO-Bayern e.V. Einblicke in Die Gattung Epipactis. Available online: https://www.aho-bayern.de/epipactis/fs_epipactis_1.html (accessed on 12 January 2025).
- Rempicci, M.; Buono, S.; Gransinigh, E.; Magrini, S. Epipactis helleborine subsp. orbicularis, a new orchid for the flora of Latium (Central Italy). J. Eur. Orchid. 2015, 47, 71. [Google Scholar]
- Bilz, M.; Kell, S.P.; Maxted, N.; Lansdown, R. V European Red List of Vascular Plants; Publications Office of the European Union: Luxembourg, 2011. [Google Scholar]
- Antonopoulos, Z.; Tsiftsis, S. Epipactis purpurata Sm. and Epipactis leptochila (Godf.) Godf. subsp. neglecta Kümpel (Orchidaceae), two new Epipactis taxa for the flora of Greece. Ber. Arbeitskreis. Heim. Orchid 2012, 29, 81–99. [Google Scholar]
- Ardelean, C.; Boceanu, B.; Nicolin, A.L. Epipactis muelleri Godfery and Epipactis neglecta (Kümpel) Kümpel (Orchidaceae) new species for the Romanian flora. In Proceedings of the International Conference on Life Sciences, Timisoara, Romania, 24–25 May 2018; pp. 54–60. [Google Scholar]
- Petrova, A.S.; Venkova, D.Y. Epipactis leptochila (Orchidaceae): A new species for the Bulgarian flora. Phytol. Balc. 2006, 12, 75–78. [Google Scholar]
- Barina, Z. Distribution Atlas of Vascular Plants in Albania; Magyar Természettudományi Múzeum: Budapest, Hungary, 2017. [Google Scholar]
- Reinhammar, L.-G. Systematics of Pseudorchis albida sl (Orchidaceae) in Europe and North America. Bot. J. Linn. Soc. 1998, 126, 363–382. [Google Scholar] [CrossRef]
- Tali, K.; Kull, T. Highly variable flowering time in Orchis ustulata (Orchidaceae): Consequences for population dynamics. Nord. J. Bot. 2001, 21, 457–466. [Google Scholar] [CrossRef]
- Verhart, F. Contribution to the knowledge of spatial distribution of Croatian orchids, results of fieldwork 2014-2019. Glas. Hrvat. Bot. Društva 2022, 10, 69–79. [Google Scholar] [CrossRef]
- Kreutz, C.A.J. Neue Erkentnisse zu den Orchideen Rumäniens. Ber. Arbeitskreis. Heim. Orchid. 2014, 31, 82–146. [Google Scholar]
- Tali, K.; Foley, M.J.Y.; Kull, T. Biological Flora of the British Isles No. 232 List Br. Vasc. Pl. (1958) 642, 4 Orchis ustulata L. J. Ecol. 2004, 92, 174. [Google Scholar] [CrossRef]
- Štípková, Z.; Kindlmann, P. Orchid Extinction over the Last 150 Years in the Czech Republic. Diversity 2021, 13, 78. [Google Scholar] [CrossRef]
- Štípková, Z.; Tsiftsis, S.; Kindlmann, P. How did the agricultural policy during the communist period affect the decline in orchid biodiversity in central and eastern Europe? Glob. Ecol. Conserv. 2021, 26, e01498. [Google Scholar] [CrossRef]
- Kazlauskas, M.; Taura, L.; Gudžinskas, Z. Current state of critically endangered Neotinea ustulata (Orchidaceae) in Lithuania and report on a new record of the species. Botanica 2022, 28, 91–101. [Google Scholar] [CrossRef]
- Wolfram, F.; Jakely, D. Orchidaceae Juss. In Exkursionsflora für Istrien; Rottensteiner, W.K., Ed.; Verlag des Naturwissenschaftlichen Vereins für Kärnten: Klagenfurt, Austria, 2014; pp. 616–637. [Google Scholar]
- Tomović, G.; Sabovljevic, M.; Djordjević, V.; Krdžić, S.; Niketic, M.; Šovran, S.; Knežević, A.; Szűcs, P.; Stoykov, D.; Ștefănuț, M.-M.; et al. New records and noteworthy data of plants, algae and fungi in SE Europe and adjacent regions, 16. Bot. Serbica 2024, 48, 93–104. [Google Scholar] [CrossRef]
- De Angelli, N.; Anghelescu, D. Orchids of Romania, 1st ed.; Rotolito Romania SA: Pantelimon, Romania, 2020. [Google Scholar]
- Dimopoulos, P.; Raus, T.; Bergmeier, E.; Constantinidis, T.; Iatrou, G.; Kokkini, S.; Strid, A.; Tzanoudakis, D. Vascular Plants of Greece: An Annotated Checklist; Botanic Garden and Botanical Museum: Berlin, Germany, 2013. [Google Scholar]
- Borovečki-Voska, L.; Horvatić, B. Orhidejski hibridi (Orchidaceae) na otoku Krku. Glas. Hrvat. Bot. Društva 2020, 8, 78–87. [Google Scholar] [CrossRef]
- Anghelescu, N.; Balogh, L.; Balogh, M.; Kigyossy, N.; Dulugeac, R.; Kertész, H.; Maalouf, R.; Georgescu, M.; Petra, S.; Toma, F. An Overview of Recently Discovered Intra and Inter-Generic Orchid Hybrids as New Additions to Romanian Flora. Sci. Pap. Ser. B Hortic. 2023, 6, 417–429. [Google Scholar]
- Vladimirov, V.; Aybeke, M.; Tan, K. New floristic records in the Balkans: 37. Phytol. Balc. 2018, 24, 397–461. [Google Scholar]





Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Radak, B.Đ.; Peškanov, J.M.; Bokić, B.S.; Anačkov, G.T. Exploring the Orchid Flora of Montenegro: Ten Newly Identified Taxa. Diversity 2025, 17, 337. https://doi.org/10.3390/d17050337
Radak BĐ, Peškanov JM, Bokić BS, Anačkov GT. Exploring the Orchid Flora of Montenegro: Ten Newly Identified Taxa. Diversity. 2025; 17(5):337. https://doi.org/10.3390/d17050337
Chicago/Turabian StyleRadak, Boris Đoka, Jovan Milan Peškanov, Bojana Slavenko Bokić, and Goran Timča Anačkov. 2025. "Exploring the Orchid Flora of Montenegro: Ten Newly Identified Taxa" Diversity 17, no. 5: 337. https://doi.org/10.3390/d17050337
APA StyleRadak, B. Đ., Peškanov, J. M., Bokić, B. S., & Anačkov, G. T. (2025). Exploring the Orchid Flora of Montenegro: Ten Newly Identified Taxa. Diversity, 17(5), 337. https://doi.org/10.3390/d17050337