



Soil Fauna-Indicators of Ungrazed Versus Grazed Grassland Ecosystems in Romania



Abstract
1. Introduction
- To analyze the structure of soil fauna communities from two grassland types (ungrazed vs. intense grazed);
- To measure environmental variables that are characteristic of the investigated ecosystems;
- To establish any correlations between the type of grassland management, environmental variables, and characteristic structures of soil fauna communities.
2. Material and Methods
2.1. Study Area
2.2. Soil Fauna Samples
2.3. Environmental Variables
2.4. Data Analysis
3. Results
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Carlier, L.; Rotar, I.; Vlahova, M.; Vidican, R. Importance and Functions of Grasslands. Not. Bot. Horti Agrobot. Cluj-Napoca 2009, 37, 25–30. [Google Scholar]
- Kachler, J.; Benra, F.; Bolliger, R.; Isaac, R.; Bonn, A.; Felipe-Lucia, M.R. Can we have it all? The role of grassland conservation in supporting forage production and plant diversity. Landsc. Ecol. 2023, 38, 4451–4465. [Google Scholar] [CrossRef]
- Pärtel, M.; Bruun, H.H.; Sammul, M. Biodiversity in temperate European grasslands: Origin and conservation. Grassl. Sci. Eur. 2005, 10, 1–14. [Google Scholar]
- European Commision. Pressure Reported by Member States Under Article 17 Reporting (Period 2013–2018). Conservation Status of Habitat Types and Species: Datasets from Article 17, Habitats Directive 92/43/EEC Reporting. 2020. Available online: https://www.eea.europa.eu/data-and-maps/data/article-17-database-habitats-directive-92-43-eec-2/article-17-2020-dataset (accessed on 20 December 2024).
- Habel, J.C.; Dengler, J.; Janišová, M.; Török, P.; Wellstein, C.; Wiezik, M. European grassland ecosystems:threatened hotspots of biodiversity. Biodiv. Conserv. 2013, 22, 2131–2138. [Google Scholar] [CrossRef]
- Tavşanoğlu, Ç.; Bernardi, R. Old-growth grasslands of Central Anatolia (Türkiye) require better conservation and management. Environ. Conserv. 2024, 51, 242–244. [Google Scholar] [CrossRef]
- Nita, A.; Hartel, T.; Manolache, S.; Ciocanea, C.M.; Miu, I.V.; Rozylowicz, L. Who is researching biodiversity hotspots in Eastern Europe? A case study on the grasslands in Romania. PLoS ONE 2019, 14, e0217638. [Google Scholar] [CrossRef]
- Török, P.; Dembicz, I.; Dajic-Stevanovic, Z.; Kuzemko, A. Grasslands of Eastern Europe. Encyclopedia of World’s Biomes; Elsevier: Amsterdam, The Netherlands, 2020; pp. 703–713. [Google Scholar]
- Potapov, A.M.; Beaulieu, F.; Birkhofer, K.; Bluhm, S.L.; Degtyarev, M.I.; Devetter, M.; Goncharov, A.A.; Klarner, B.; Korobushkin, D.I.; Liebke, D.F.; et al. Feeding habits and multifunctional classification of soil-associated consumers from protists to vertebrates. Biol. Rev. 2022, 97, 1057–1117. [Google Scholar] [CrossRef] [PubMed]
- Angst, G.; Potapov, A.; Joly, F.X.; Angst, S.; Frouz, J.; Ganault, P.; Eisenhauer, N. Conceptualizing soil fauna effects on labile and stabilized soil organic matter. Nat. Commun. 2024, 15, 5005. [Google Scholar] [CrossRef]
- Bünemann, E.K.; Bongiorno, G.; Bai, Z.; Creamer, R.E.; De Deyn, G.; de Goede, R.; Fleskens, L.; Geissen, V.; Kuyper, T.W.; Mäder, P.; et al. Soil quality—A critical review. Soil Biol. Biochem. 2018, 120, 105–125. [Google Scholar] [CrossRef]
- Cole, L.; Mark, A.; Bradford, M.A.; Shaw, P.J.A.; Bardgett, R.D. The abundance, richness and functional role of soil meso- and macrofauna in temperate grassland—A case study. Appl. Soil Ecol. 2006, 33, 186–198. [Google Scholar] [CrossRef]
- Menta, C. Soil Fauna Diversity—Function, Soil Degradation, Biological Indices, Soil Restoration. In Biodiversity Conservation and Utilization in a Diverse World; Lameed, G.A., Ed.; IntechOpen: London, UK, 2012; pp. 59–94. [Google Scholar]
- Coleman, D.C.; Wall, D.H. Soil Fauna: Occurrence, Biodiversity, and Roles in Ecosystem Function. In Soil Microbiology, Ecology and Biochemistry, 4th ed.; Eldor, A.P., Ed.; Academic Press: Cambridge, MA, USA, 2015; pp. 1–41. [Google Scholar]
- Manu, M.; Băncilă, R.I.; Mountford, O.J.; Onete, M. Soil Invertebrate Communities as Indicator of Ecological Conservation Status of Some Fertilised Grasslands from Romania. Diversity 2022, 14, 1031. [Google Scholar] [CrossRef]
- Manu, M.; Băncilă, R.I.; Onete, M. Effect of Grazing Management on Predator Soil Mite Communities (Acari: Mesotigmata) in Some Subalpine Grasslands from the Făgăraş Mountains—Romania. Insects 2023, 14, 626. [Google Scholar] [CrossRef]
- Fusco, T.; Fortini, L.; Casale, F.; Di Giulio, A. Assessing soil quality of Italian Western Alps protected areas by QBS-ar: Impact of management and habitat type on soil microarthropods. Environ. Monit. Assess. 2023, 195, 1287. [Google Scholar] [CrossRef] [PubMed]
- Menta, C.; Leoni, A.; Gardi, C.; Conti, F.D. Are grasslands important habitats for soil microarthropod conservation? Biodivers. Conserv. 2011, 20, 1073–1087. [Google Scholar] [CrossRef]
- Menta, C.; Conti, F.D.; Pinto, S. Microarthropods biodiversity in natural, seminatural and cultivated soils—QBS-ar approach. Appl. Soil Ecol. 2017, 123, 740–743. [Google Scholar] [CrossRef]
- Potapov, A.M.; Goncharov, A.A.; Semenina, E.E.; Korotkevich, A.Y.; Tsurikov, S.M.; Rozanova, O.L.; Anichkin, A.E.; Zuev, A.G.; Samoylova, E.S.; Semenyuk, I.I.; et al. Arthropods in the subsoil: Abundance and vertical distribution as related to soil organic matter, microbial biomass and plant roots. Eur. J. Soil Biol. 2017, 82, 88–97. [Google Scholar] [CrossRef]
- Mioulet, C.; Schrama, M.; Berg, M.P.; Hannula, S.E. Comparison of metrics to reveal the role of soil fauna in soil health assessment in peat meadow restoration. Eur. J. Soil Sci. 2024, 75, e70018. [Google Scholar] [CrossRef]
- Triquet, C.; Perennes, M.; Séchaud, R.; van der Meer, M.; Fabian, Y.; Jeanneret, P. What evidence exists on the effect of the main European lowland crop and grassland management practices on biodiversity indicator species groups? A systematic map. Environ. Evid. 2024, 13, 20. [Google Scholar] [CrossRef]
- Miller, J.J.; Battigelli, J.P.; Willms, W.D. Grazing Protection Influences Soil Mesofauna in Ungrazed and Grazed Riparian and Upland Pastures. Rangel. Ecol. Manag. 2014, 67, 429–434. [Google Scholar] [CrossRef]
- Siebert, J.; Thakur, M.P.; Reitz, T.; Schaedler, M.; Schulz, E.; Yin, R.; Weigelt, A.; Eisenhauer, N. Extensive grassland-use sustains high levels of soil biological activity, but does not alleviate detrimental climate change effects. In Resilience in Complex Socio-Ecological Systems; Advances in Ecological Research; Academic Press: Cambridge, MA, USA, 2019; Volume 60, pp. 25–58. [Google Scholar]
- Johnston, A.S.A.; Sibly, R.M. Multiple environmental controls explain global patterns in soil animal communities. Oecologia 2020, 192, 1047–1056. [Google Scholar] [CrossRef]
- Manu, M.; Băncilă, R.I.; Mountford, O.J.; Maruşca, T.; Blaj, V.A.; Onete, M. Soil Mite (Acari: Mesostigmata) Communities and Their Relationship with Some Environmental Variables in Experimental Grasslands from Bucegi Mountains in Romania. Insects 2022, 13, 285. [Google Scholar] [CrossRef]
- Báldi, A. Using higher taxa as surrogates of species richness: A study based on 3700 Coleoptera, Diptera, and Acari species in Central-Hungarian reserves. Basic Appl. Ecol. 2003, 4, 589–593. [Google Scholar] [CrossRef]
- Rosser, N. Shortcuts in biodiversity research: What determines the performance of higher taxa as surrogates for species? Ecol. Evol. 2017, 7, 2595–2603. [Google Scholar] [CrossRef]
- Popovici, I.; Ciobanu, M. Diversity and distribution of nematode communities in grasslands from Romania in relation to vegetation and soil characteristics. Appl. Soil Ecol. 2000, 14, 27–36. [Google Scholar] [CrossRef]
- Korsós, Z.; Lazányi, E. Millipedes (Diplopoda) of Maramureş (Romania). Stud. Univ. Vasile Goldiş Arad Ser. Ştiinţ. Vieţii. 2008, 18, 199–209. [Google Scholar]
- Boros, G.; Dózsa-Farkas, K. The Enchytraeid fauna of Romania. Soil Org. 2015, 87, 137–147. [Google Scholar]
- Călugăr, A. Qualitative and quantitative studies upon the edaphic microarthropods fauna in some grassland ecosystems from Moldavian Plain. Universitatea de Ştiinţe Agricole şi Medicină Veterinară Iaşi (Romania). Studii şi Comunicări Compl. Muz. Şt. Nat. Ion Borcea Bacău 2006, 21, 230–231. [Google Scholar]
- Călugăr, A. Researches on the edaphic mesofauna from some agro-ecosystems from North-Eastern part of Romania. Rom. Biol. Sci. 2007, 5, 92–100. [Google Scholar]
- Călugăr, A. Researches on the edaphic mesofauna from some grassland ecosystems from the inferior section of Prut riverside (Romania). Lucr. Stiinţ. Univ. Agron. Ion Ionescu de la Brad Iaşi Ser. Agron. 2010, 53, 84–88. [Google Scholar]
- Călugăr, A.; Ivan, O. Saline grasslands, a suitable environment for maintaining the functional relationships of the edaphic mesofauna? Lucr. Stiinţ. Univ. Agron. Ion Ionescu de la Brad Iaşi Ser. Agron. 2018, 61, 123–129. [Google Scholar]
- Băncilă, R.I.; Cogălniceanu, D.; Manu, M.; Plăiaş̧u, R.; Stănescu, F.; Memedemin, D.; Skolka, M.; Tofan, L.; Lăcătuşu, A. A Multi-Community Approach in Biodiversity Assessment of a Peat Bog in the Southern Carpathians (Romania) and Implications for Conservation. Environ. Entomol. 2023, 52, 217–229. [Google Scholar] [CrossRef] [PubMed]
- Manu, M.; Bîrsan, C.C.; Mountford, O.; Lăcătuşu, A.R.; Onete, M. Preliminary study on soil fauna as a tool for monitoring of the “Springs Complex of Corbii Ciungi” protected area—Romania. Sci. Papers Ser. D Anim. Sci. 2020, 63, 272–280. [Google Scholar]
- Onete, M.; Zaharia, D.; Nicoară, G.R.; Manu, M. Studii Privind Aprecierea Valorii Pastorale şi a Capacităţii de Păşunat în Unele Pajişti Din Zona Sud-Vestică a Masivului Făgăraş; Editura Ars Docendi, Universitatea Bucureşti: Bucureşti, Romania, 2021; pp. 1–124. [Google Scholar]
- Florea, N.; Munteanu, I. Sistemul Român de Taxonomie a Solurilor (SRTS-2003); Editura ESTFALIA: Bucureşti, Romania, 2003; pp. 1–182. [Google Scholar]
- Balogh, J. The Oribatid Genera of the World; Akademiai Kiado: Budapest, Hungary, 1972; pp. 1–188. [Google Scholar]
- Gîdei, P.; Popescu, I.E. Îndrumator Pentru Cunoas, terea Coleopterelor; Editura Pim: Iaşi, Romania, 2009; pp. 5–419. [Google Scholar]
- Krantz, G.W.; Walter, D.E. A Manual of Acarology, 3rd ed.; Texas Tech University Press: Lubbock, TX, USA, 2009; pp. 1–807. [Google Scholar]
- Ceuca, T. Diplopoda. In Determinatorul Ilustrat al Florei si Faunei Romaniei—Mediul Terestru; Godeanu, S.P., Ed.; Vasile Goldiş University Press: Arad, România, 2010; Volume 3, pp. 290–300. [Google Scholar]
- Platnick, N.I. World Spider Catalog. World Spider Catalog. Version 19.5. Natural History Museum Bern. 2018. Available online: http://wsc.nmbe.ch (accessed on 5 February 2022).
- Brussaard, L.; Behan-Pelletier, V.M.; Bignell, D.E.; Brown, V.K.; Didde, W.; Folgaria, P.; Fragoso, C.; Freckman, D.W.; Gupta, V.V.S.R.; Hattori, T.; et al. Biodiversity and ecosystem functioning in soil. Ambio. J. Hum. Environ. 1997, 26, 563–570. [Google Scholar]
- Chahartaghi, M.; Langel, R.; Scheu, S.; Ruess, L. Feeding guilds in Collembola based on nitrogen stable isotope ratios. Soil Biol. Biochem. 2005, 37, 1718–1725. [Google Scholar] [CrossRef]
- The R Project for Statistical Computing. Available online: http://www.R-project.org/ (accessed on 10 December 2024).
- Wickham, H. ggplot2: Elegant Graphics for Data Analysis; Springer: New York, NY, USA, 2016; pp. 1–276. [Google Scholar]
- Zuur, A.F.; Ieno, E.N.; Walker, N.J.; Saveliev, A.A.; Smith, G.M. Mixed Effects Models and Extensions in Ecology with R; Springer: New York, NY, USA, 2009; pp. 1–574. [Google Scholar]
- Kindt, R.; Coe, R. Tree Diversity Analysis: A Manual and Software for Common Statistical Methods for Ecological and Biodiversity Studies; World Agroforestry Centre (ICRAF): Nairobi, Kenya, 2005; pp. 1–196. [Google Scholar]
- Bates, D.; Maechler, M.; Bolker, B.; Walker, S. Fitting Linear Mixed-Effects Models Using lme4. J. Stat. Softw. 2015, 67, 1–48. [Google Scholar] [CrossRef]
- Bolker, B. GLMM FAQ. Available online: http://bbolker.github.io/mixedmodels-misc/glmmFAQ.html (accessed on 15 June 2022).
- Fox, J.; Hong, J. Effect Displays in R for Multinomial and Proportional-Odds Logit Models: Extensions to the effects Package. J. Stat. Softw. 2009, 32, 1–24. [Google Scholar] [CrossRef]
- Oksanen, J.F.; Blanchet, G.; Friendly, M.; Kindt, R.; Legendre, P.; McGlinn, D.; Minchin, P.R.; O’Hara, R.B.; Simpson, G.L.; Solymos, P.; et al. Vegan: Community Ecology Package. R Package Version 2.5-7. Available online: https://CRAN.R-project.org/package=vegan (accessed on 20 June 2022).
- Manu, M.; Lăcătuşu, A.; Bodescu, F.; Nicoară, R.; Chiriac, L.S.; Onete, M. Are habitats with Campanula romanica Săvul. preferred by the soil fauna? Sci. Papers Ser. D Anim. Sci. 2023, 46, 105–112. [Google Scholar]
- Curry, J.P. The invertebrate fauna of grassland and its influence on productivity. The composition of the fauna. Grass Forage Sci. 1987, 42, 103–120. [Google Scholar] [CrossRef]
- Schrama, M.; Casper, W.; Quist, C.W.; de Groot, A.; Cieraad, E.; Ashworth, D.; Laros, I.; Hestbjerg Hansen, L.; Leff, J.; Fierer, N.; et al. Cessation of grazing causes biodiversity loss and homogenization of soil food webs. Proc. R. Soc. B. 2023, 290, 20231345. [Google Scholar] [CrossRef]
- Arnott, A.; Riddell, G.; Emmerson, M.; Caruso, T.; Reid, N. Upland grassland habitats and agri-environment schemes change soil microarthropod abundance. J. Appl. Ecol. 2021, 58, 2256–2265. [Google Scholar] [CrossRef]
- Souza, R.; Hartzell, S.; Ferraz, A.P.F.; de Almeida, A.Q.; de Sousa Lima, R.S.; Antonino, A.C.D.; de Souza, E.S. Dynamics of soil penetration resistance in water-controlled environments. Soil Till. Res. 2021, 205, 104768. [Google Scholar] [CrossRef]
- Aupic-Samain, A.; Baldy, V.; Delcourt, N.; Krogh, P.H.; Gauquelin, T.; Fernandez, C.; Santonja, M. Water availability rather than temperature control soil fauna community structure and prey–predator interactions. Funct. Ecol. 2020, 35, 1550–1559. [Google Scholar] [CrossRef]
- Amossé, J.; Dózsa-Farkas, K.; Boros, G.; Rochat, G.; Sandoz, G.; Fournier, B.; Mitchell, E.A.D.; Le Bayon, R.C. Patterns of earthworm, enchytraeid and nematode diversity and community structure in urban soils of different ages. Eur. J. Soil Biol. 2016, 73, 46–58. [Google Scholar] [CrossRef]
- Bogyó, D.; Magura, T.; Nagy, D.D.; Tóthmérész, B. Distribution of millipedes (Myriapoda, Diplopoda) along a forest interior—forest edge—grassland habitat complex. ZooKeys 2015, 510, 181–195. [Google Scholar] [CrossRef]
- Heděnec, P.; Jiménez, J.J.; Moradi, J.; Domene, X.; Hackenberger, D.; Barot, S.; Frossard, A.; Oktaba, L.; Filser, J.; Kindlmann, P.; et al. Global distribution of soil fauna functional groups and their estimated litter consumption across biomes. Sci. Rep. 2022, 12, 17362. [Google Scholar] [CrossRef]
- Seeber, J.; Seeber, G.U.H.; Kössler, W.; Langel, R.; Scheu, S.; Meyer, E. Abundance and trophic structure of macro-decomposers on alpine pastureland (Central Alps, Tyrol): Effects of abandonment of pasturing. Pedobiologia 2005, 49, 221–228. [Google Scholar] [CrossRef]
- Ríos-Díaz, C.L.; Moreno, C.E.; Ortega-Martínez, I.J.; Zuria, I.; Escobar, F.; Castellanos, I. Sheep herding in small grasslands promotes dung beetle diversity in a mountain forest landscape. J. Insect. Conserv. 2021, 25, 13–26. [Google Scholar] [CrossRef]
- Feketeová, Z.; Mangová, B.; Cierniková, M. The Soil Chemical Properties Influencing the Oribatid Mite (Acari; Oribatida) Abundance and Diversity in Coal Ash Basin Vicinage. Appl. Sci. 2021, 11, 3537. [Google Scholar] [CrossRef]

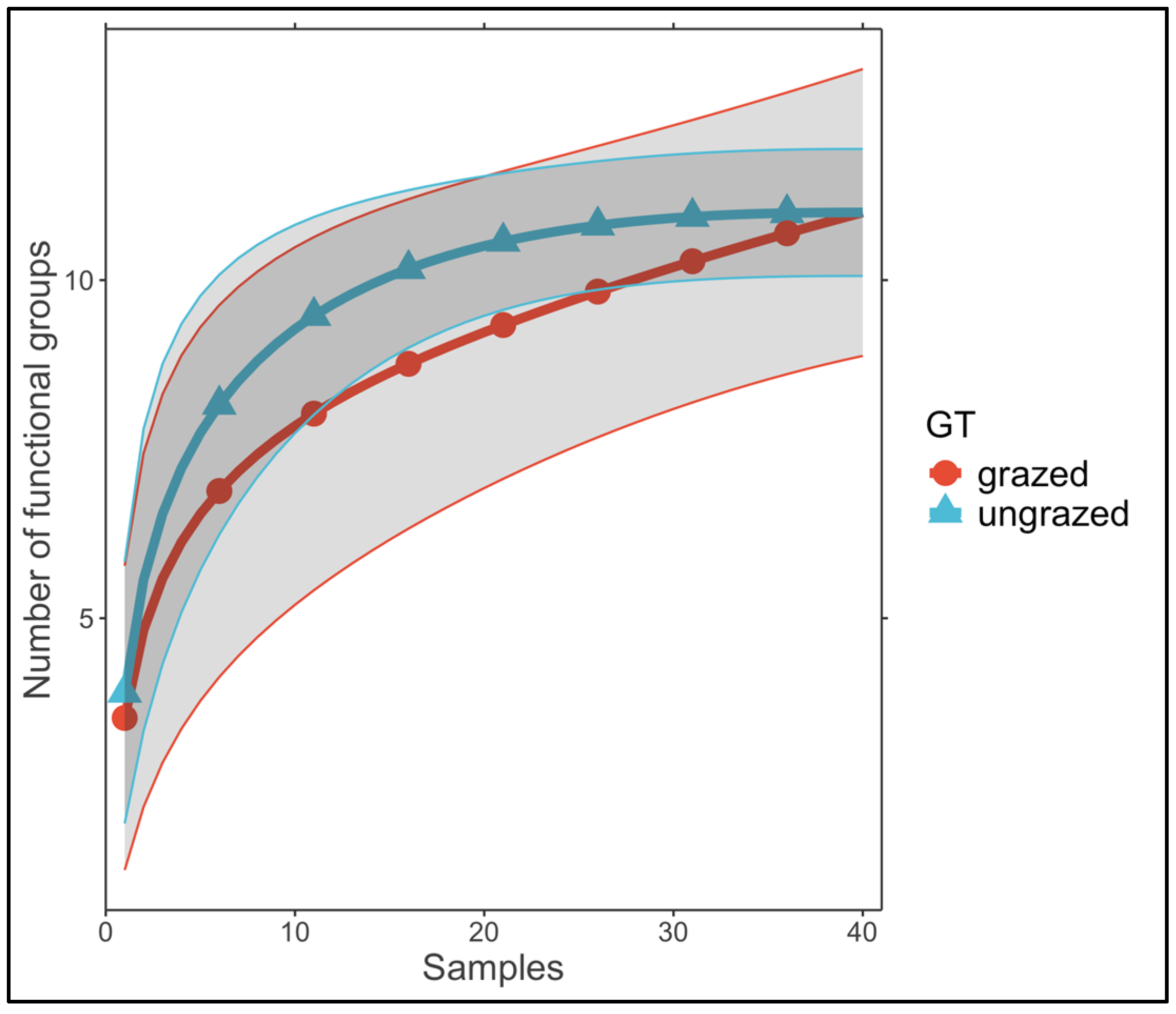
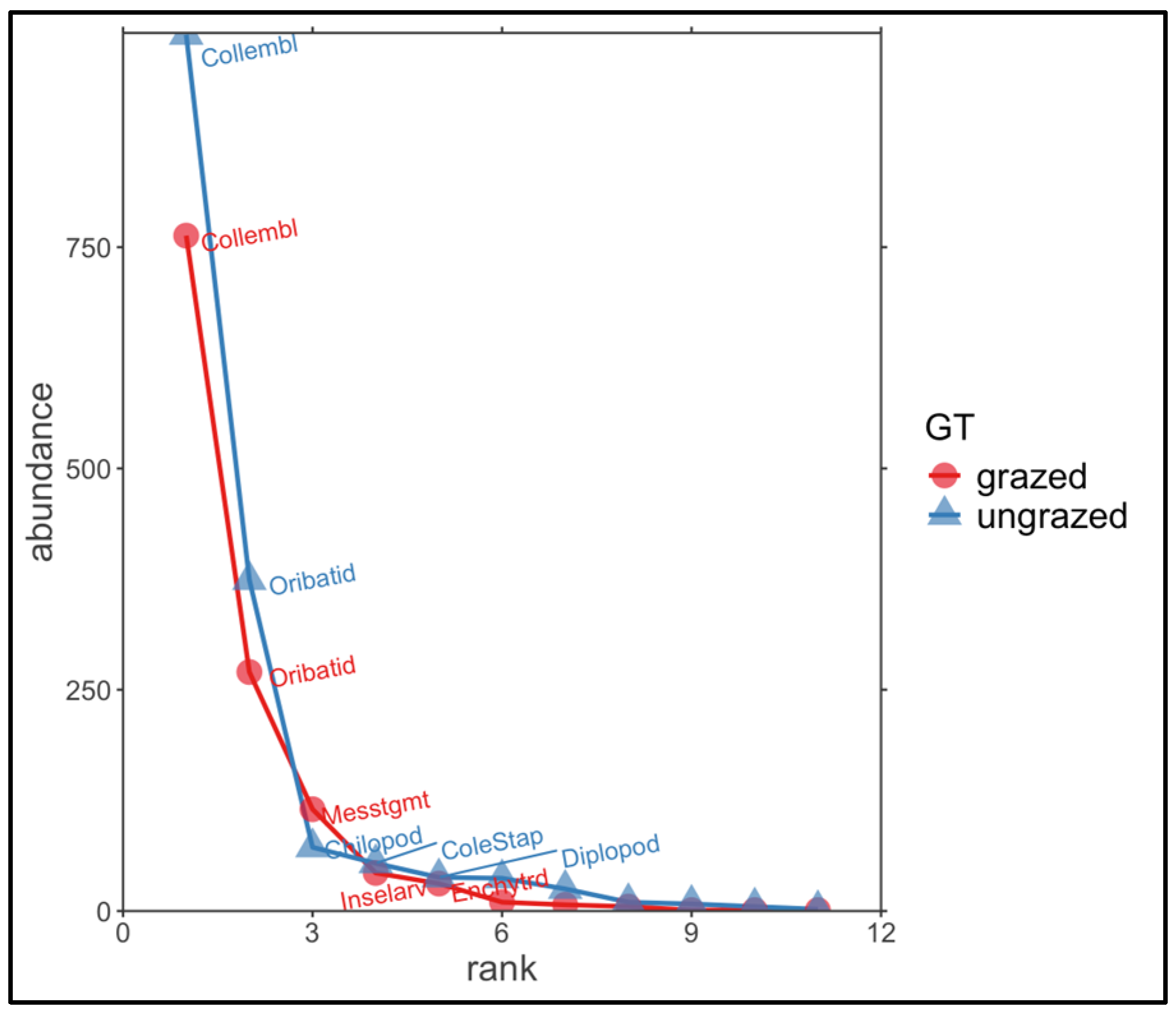

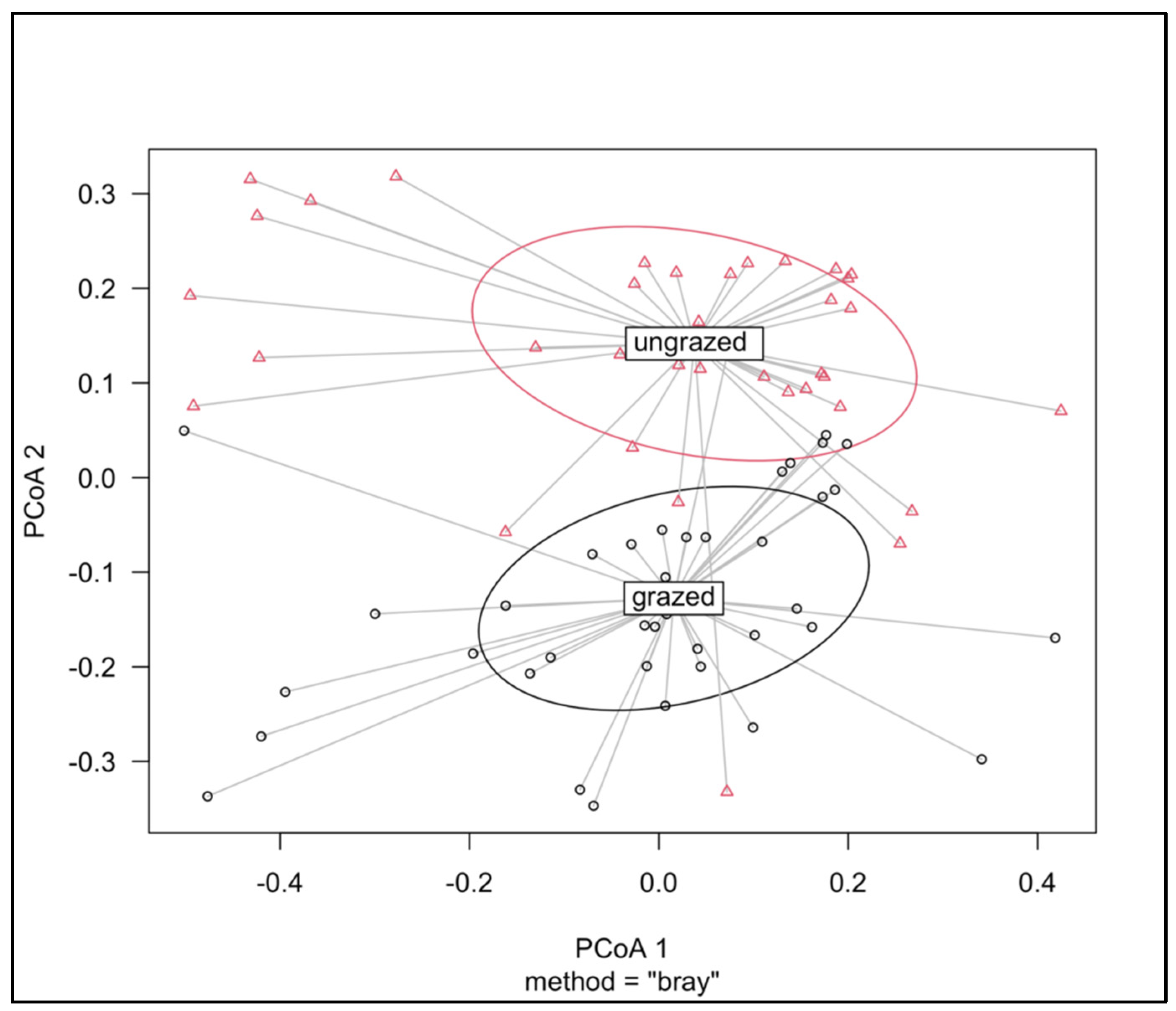

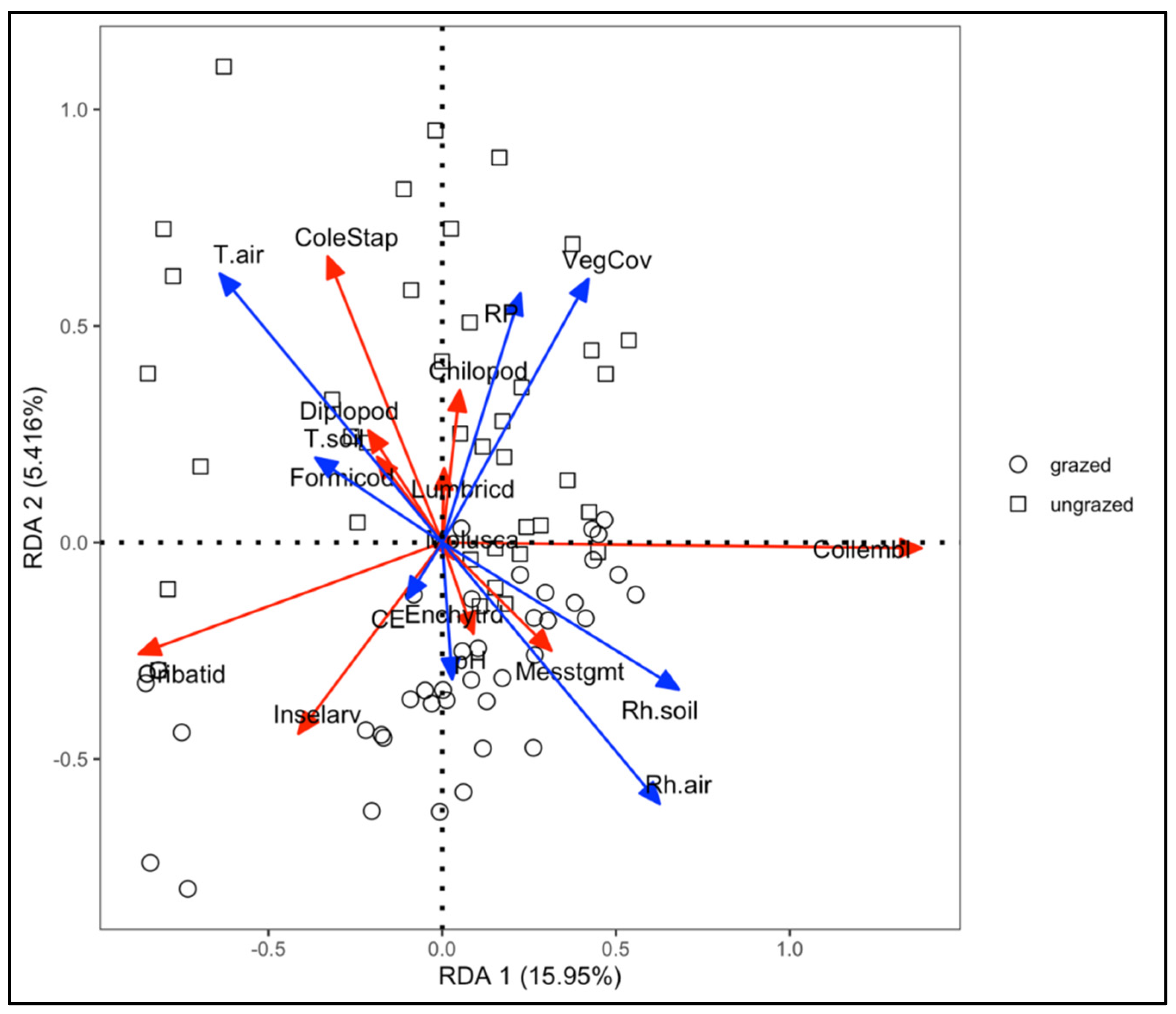

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Ungrazed | Tair (°C) | Rhair (%) | Tsoil (°C) | Rhsoil (%) | RP (MPa) | pH | CE (μS/cm) | VegCov (%) |
|---|---|---|---|---|---|---|---|---|
| Cun | 18.8 ± 0.08 | 77.8 ± 1.81 | 14.37 ± 0.22 | 77.13 ± 4.13 | 1.95 ± 0.3 | 4.58 ± 0.45 | 191.2 ± 27.10 | 99.5 ± 0.5 |
| Sun | 17.47 ± 0.46 | 85.5 ± 1.50 | 14.87 ± 1.46 | 73.18 ± 4.69 | 0.92 ± 0.15 | 4.67 ± 0.08 | 63.93 ± 18.79 | 99.5 ± 1.58 |
| Gun | 30.68 ± 0.76 | 70.7 ± 3.49 | 27.07 ± 1.67 | 42.77 ± 5.03 | 1.42 ± 0.17 | 3.488 ± 0.35 | 150.79 ± 36.92 | 99 ± 2.10 |
| Vun | 31.24 ± 1.53 | 63.3 ± 4.00 | 19.71 ± 1.69 | 56.24 ± 8.42 | 1.62 ± 0.28 | 4.713 ± 0.95 | 31.1 ± 29.60 | 96.5 ± 7.47 |
| Average | 24.55 ± 3.71 | 74.33 ± 4.75 | 19.00 ± 2.94 | 62.33 ± 7.94 | 1.48 ± 0.21 | 4.36 ± 0.29 | 109.26 ± 37.19 | 98.63 ± 2.14 |
| p | <0.001 | <0.001 | <0.001 | <0.001 | <0.001 | 0.055 | <0.001 | 0.301 |
| Grazed | Tair (°C) | Rhair (%) | Tsoil (°C) | Rhsoil (%) | RP (MPa) | pH | CE (μS/cm) | VegCov (%) |
|---|---|---|---|---|---|---|---|---|
| Cg | 16.6 ± 0.47 | 87.8 ± 1.81 | 16.17 ± 0.46 | 86.13 ± 4.32 | 1.25 ± 0.30 | 4.36 ± 0.45 | 94.3 ± 11.6 | 93.5 ± 3.37 |
| Sg | 18 ± 1.25 | 88.5 ± 1.35 | 20.39 ± 3.30 | 78.5 ± 4.30 | 0.92 ± 0.27 | 4.29 ± 0.17 | 55.86 ± 37.68 | 96 ± 5.67 |
| Gg | 28.41 ± 1.08 | 70.3 ± 4.32 | 23.68 ± 2.99 | 43.43 ± 7.91 | 1.19 ± 0.29 | 4 ± 0.41 | 73.3 ± 46.58 | 89 ± 9.66 |
| Vg | 27.73 ± 1.14 | 88 ± 1.94 | 18.09 ± 2.47 | 64.23 ± 6.12 | 1.08 ± 0.15 | 4.72 ± 0.19 | 59.75 ± 25.42 | 86.5 ± 17.32 |
| Average | 20.18 ± 2.75 | 83.65 ± 4.52 | 19.58 ± 1.61 | 68.07 ± 9.38 | 1.11 ± 0.07 | 4.34 ± 0.14 | 70.80 ± 8.67 | 91.25 ± 2.14 |
| p | <0.001 | <0.001 | <0.001 | <0.001 | 0.032 | <0.001 | 0.574 | 0.188 |
| Abundance | Richness | H′ | |||||
|---|---|---|---|---|---|---|---|
| Df | χ2 | p | χ2 | p | χ2 | p | |
| GT | 1 | 0.091 | 0.762 | 0.135 | 0.712 | 0.015 | 0.901 |
| T soil | 1 | 3.140 | 0.076 | 0.003 | 0.953 | 0.061 | 0.804 |
| Rh soil | 1 | 1.079 | 0.298 | 0.076 | 0.781 | 3.509 | 0.061 |
| RP | 1 | 48.279 | <0.001 | 0.788 | 0.374 | 3.363 | 0.066 |
| pH | 1 | 16.724 | <0.001 | 0.348 | 0.765 | 1.516 | 0.218 |
| CE | 1 | 1.383 | 0.239 | 0.088 | 0.766 | 0.239 | 0.624 |
| VegCov | 1 | 12.463 | 0.001 | 2.097 | 0.147 | 2.287 | 0.130 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Manu, M.; Băncilă, R.I.; Onete, M. Soil Fauna-Indicators of Ungrazed Versus Grazed Grassland Ecosystems in Romania. Diversity 2025, 17, 323. https://doi.org/10.3390/d17050323
Manu M, Băncilă RI, Onete M. Soil Fauna-Indicators of Ungrazed Versus Grazed Grassland Ecosystems in Romania. Diversity. 2025; 17(5):323. https://doi.org/10.3390/d17050323
Chicago/Turabian StyleManu, Minodora, Raluca Ioana Băncilă, and Marilena Onete. 2025. "Soil Fauna-Indicators of Ungrazed Versus Grazed Grassland Ecosystems in Romania" Diversity 17, no. 5: 323. https://doi.org/10.3390/d17050323
APA StyleManu, M., Băncilă, R. I., & Onete, M. (2025). Soil Fauna-Indicators of Ungrazed Versus Grazed Grassland Ecosystems in Romania. Diversity, 17(5), 323. https://doi.org/10.3390/d17050323