

Investigating Endemic Alpine Communities of Papaver corona-sancti-stephani and Cerastium lerchenfeldianum in the Southern Carpathians



Abstract
1. Introduction
2. Materials and Methods
2.1. Area of Study and Sampling Sites
2.2. Field Methods
2.3. Data Analysis
3. Results
- H (Hemicryptophytes): perennial plants with regenerating shoots at ground level, protected by leaves or snow, adapted to cold or extreme climates.
- Ch (Chamaephytes): low, woody or semi-woody plants, with shoots close to the ground, adapted to dry and exposed environments. These plants, being low, are characteristic of extreme environments.
- Th (Therophytes): annual plants that survive the unfavorable season as seeds, characteristic of disturbed areas.
- G (Geophytes): Perennial plants that keep their regenerating organs underground (bulbs, tubers, rhizomes), adapted to variable conditions.The Kruskal–Wallis test indicated that the number of species belonging to each bioform (Ch, Th, H, and G) did not differ significantly among the three massifs.
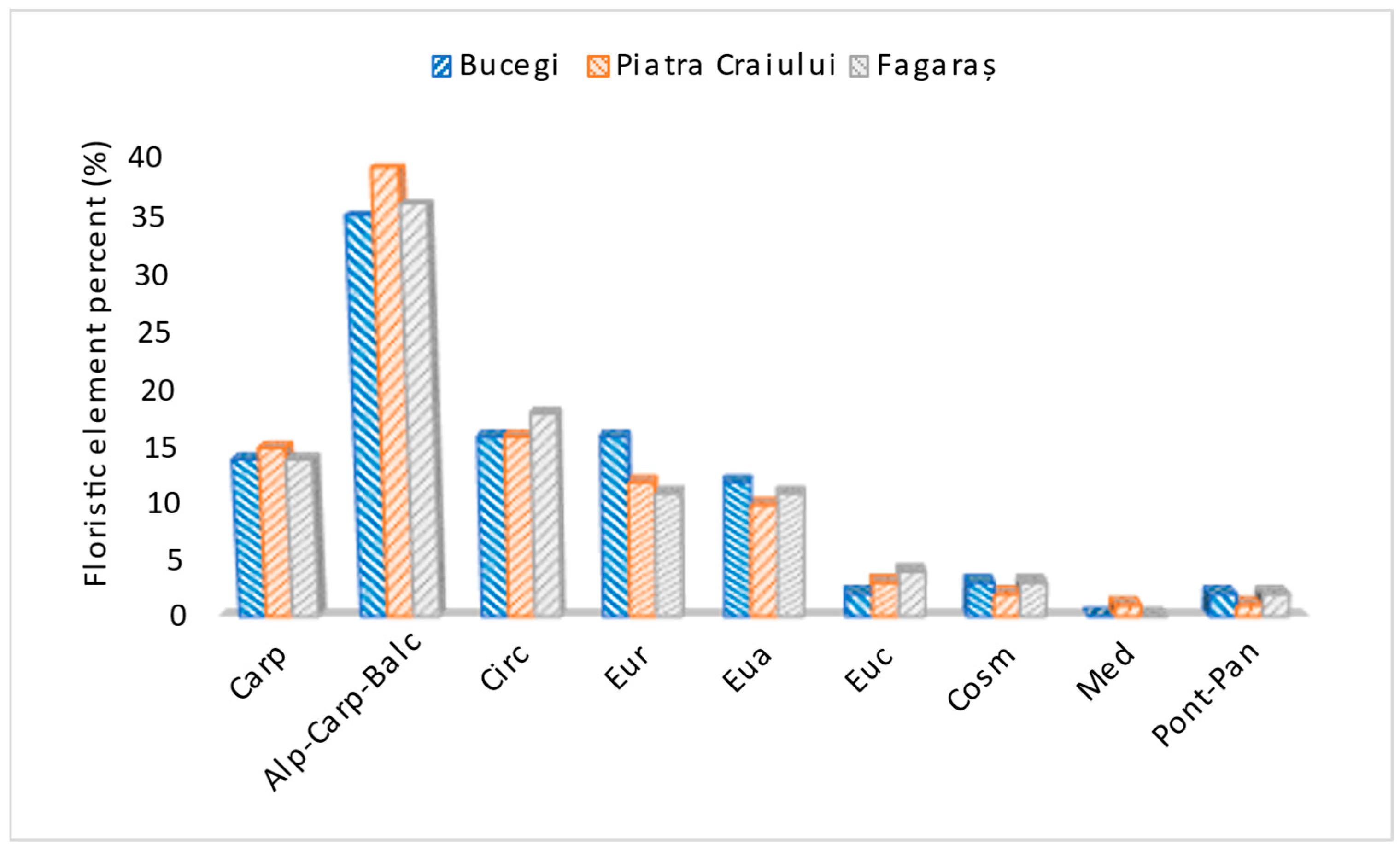
- Carp (Carpathian): Endemic species, with restricted distribution or specific to the Carpathian Mountain range.
- Alp-Carp-Balc (Alpine-Carpathian-Balkan): Species with a common distribution in the Alps, Carpathians and Balkans.
- Circ (Circumpolar): Species distributed in cold or temperate.
- Eur (European): Species with a distribution restricted to Europe.
- Eua (Eurasian): Species distributed in both Europe and Asia.
- Euc (Europeo-Centrale): Species characteristic of Central Europe.
- Cosm (Cosmopolitan): Globally distributed species, adapted to different ecological conditions and found on most continents.
- Med (Mediterraneans): Species characteristic of the Mediterranean region, adapted to the warm and dry climates.
- Pont-Pan (Pontic-Pannonian): Species in the Pontic (around the Black Sea) and Pannonian (Pannonian Plain) regions.
4. Discussion
Conservation
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Conflicts of Interest
Abbreviations
| B | Bucegi Massif |
| PC | Piatra Craiului Massif |
| F | Făgăraș Massif |
References
- Körner, C. Mountain biodiversity, its causes and function: An overview. Mt. Biodivers. 2024, 33, 11–17. [Google Scholar]
- Leuschner, C.; Ellenberg, H. Vegetation of the alpine and nival belts. In Ecology of Central European Non-Forest Vegetation: Coastal to Alpine, Natural to Man-Made Habitats: Vegetation Ecology of Central Europe, Volume II; Springer: Cham, Switzerland, 2017; pp. 271–431. [Google Scholar]
- Varga, Z. Extra-Mediterranean refugia, post-glacial vegetation history and area dynamics in Eastern Central Europe. In Relict Species: Phylogeography and Conservation Biology; Habel, J.C., Assmann, T., Eds.; Springer: Berlin/Heidelberg, Germany, 2009; pp. 57–87. [Google Scholar]
- Zheng, G.; Gu, J.; Zhao, W.; Zhang, Y.; Guan, Z.; Lei, M.; He, C. Spatial, Geographical, Climatic, and Edaphic Influences on Moss Community Structure: A Case Study from Qinhuangdao, China. Forests 2024, 15, 424. [Google Scholar] [CrossRef]
- Stewart, J.R.; Lister, A.M.; Barnes, I.; Dalén, L. Refugia revisited: Individualistic responses of species in space and time. Proc. R. Soc. B Biol. Sci. 2010, 277, 661–671. [Google Scholar] [CrossRef]
- Pils, G. Systematics, distribution, and karyology of the Festuca violacea Group (Poaceae) in the Eastern Alps. Plant Syst. Evol. 1980, 136, 73–124. [Google Scholar] [CrossRef]
- Kropf, M.; Comes, H.P.; Kadereit, J.W. Long-distance dispersal vs. vicariance: The origin and genetic diversity of alpine plants in the Spanish Sierra Nevada. New Phytol. 2006, 172, 169–184. [Google Scholar] [CrossRef]
- Parisod, C. Plant speciation in the face of recurrent climate changes in the Alps. Alp. Bot. 2022, 132, 21–28. [Google Scholar] [CrossRef]
- Vintsek, L.; Klichowska, E.; Nowak, A.; Nobis, M. Genetic differentiation, demographic history and distribution models of high alpine endemic vicariants outline the response of species to predicted climate changes in a Central Asian biodiversity hotspot. Ecol. Indic. 2022, 144, 109419. [Google Scholar] [CrossRef]
- The World Checklist of Vascular Plants (WCVP). Available online: www.gbif.org (accessed on 20 November 2024).
- Coldea, G.; Cristea, V. Description of the plant associations distinguished in the Retezat National Park. In The Vascular Plant Communities of the Retezat National Park (Southern Carpathians); Springer: Cham, Switzerland, 2022; pp. 85–245. [Google Scholar]
- Mucina, L.; Bültmann, H.; Dierßen, K.; Theurillat, J.; Raus, T.; Čarni, A.; Šumberová, K.; Willner, W.; Dengler, J.; García, R.G. Vegetation of Europe: Hierarchical floristic classification system of vascular plant, bryophyte, lichen, and algal communities. Appl. Veg. Sci. 2016, 19, 3–264. [Google Scholar] [CrossRef]
- Biţă-Nicolae, C.D. The screes vegetation of the higher basin of the Prahova river. Rom. J. Biol.-Plant Biol. 2009, 54, 157–172. [Google Scholar]
- Rodwell, J.S.; Schaminée, J.H.J.; Mucina, L.; Pignatti, S.; Dring, J.; Moss, D. The diversity of European vegetation. An overview of phytosociological alliances and their relationships to EUNIS habitats. Rep. EC-LNV 2002, 54, 168. [Google Scholar]
- Grabherr, G.; Gottfried, M.; Pauli, H. Climate change impacts in alpine environments. Geogr. Compass 2010, 4, 1133–1153. [Google Scholar] [CrossRef]
- Llambi, L.D.; Gámez, L.E.; Pelayo, R.; Azócar, C.J.; Torres, J.E.; Márquez, N.J.; Berdugo, M.B.; Cuesta, F.; Ramirez, L.A. Species, Growth Form, and Biogeographic Diversity of Summit Vegetation along an Elevation Gradient in the Tropical Andes: A Baseline for Climate Change Monitoring. J. Mt. Sci. 2022, 19, 3441–3457. [Google Scholar] [CrossRef]
- Squires, V.R.; Dengler, J.; Hua, L.; Feng, H. Grasslands of the Mediterranean Basin and the Middle East and their Management. In Grasslands of the World; CRC Press: Boca Raton, FL, USA, 2018; pp. 103–126. [Google Scholar]
- Aalto, J.; le Roux, P.C.; Luoto, M. Vegetation Mediates Soil Temperature and Moisture in Arctic-Alpine Environments. Arct. Antarct. Alp. Res. 2013, 45, 429–439. [Google Scholar] [CrossRef]
- Valachovic, M. Papaverion tatrici, a vicarious alliance of alpine limestone-scree communities in the Western Carpathians. Biol.-Bratisl. 1995, 50, 377–390. [Google Scholar]
- Onaca, A.; Ardelean, F.; Urdea, P.; Magori, B. Southern Carpathian rock glaciers: Inventory, distribution and environmental controlling factors. Geomorphology 2017, 293, 391–404. [Google Scholar] [CrossRef]
- Shijin, W.; Yuande, Y.; Yanjun, C. Global snow-and ice-related disaster risk: A review. Nat. Hazards Rev. 2022, 23, 03122002. [Google Scholar] [CrossRef]
- Google Earth Pro. 7.3.6. Available online: https://www.google.com/earth/about/versions/#earth-pro (accessed on 15 December 2024).
- Braun-Blanquet, J. Pflanzensoziologie: Grundzüge der Vegetationskunde, 3rd ed.; Springer: Vienna, Austria; New York, NY, USA, 1964; 631p. [Google Scholar]
- World Flora Online. Available online: http://wfoplantlist.org/ (accessed on 20 November 2024).
- Biță-Nicolae, C.; Sanda, V. Cormophlora of Romania: Spontaneous and Cultivated Cormophytes in Romania; Lambert Academic Publishing: Saarbrucken, Germany, 2011. [Google Scholar]
- Ellenberg, H.H. Vegetation Ecology of Central Europe; Cambridge University Press: Cambridge, UK, 1988. [Google Scholar]
- Past 4—The Past of the Future. Available online: https://www.nhm.uio.no/english/research/resources/past/ (accessed on 20 December 2024).
- RStudio Team. RStudio Desktop 2024.12.0-467: Integrated Development for; RStudio, PBC: Boston, MA, USA, 2020; Available online: https://posit.co/ (accessed on 12 February 2025).
- Oksanen, J.; Simpson, G.; Blanchet, F.; Kindt, R.; Legendre, P.; Minchin, P.; O’Hara, R.; Solymos, P.; Stevens, M.; Szoecs, E.; et al. Vegan: Community Ecology Package. R package version 2.6-10, 2025. Available online: https://CRAN.R-project.org/package=vegan (accessed on 12 February 2025).
- Bacaro, G.; Gioria, M.; Ricotta, C. Testing for differences in beta diversity from plot-to-plot dissimilarities. Ecol. Res. 2012, 27, 285–292. [Google Scholar] [CrossRef]
- Clarke, K.; Somerfield, P.; Chapman, M.G. On resemblance measures for ecological studies, including taxonomic dissimilarities and a zero-adjusted Bray-Curtis coefficient for denuded assemblages. J. Exp. Mar. Biol. Ecol. 2006, 330, 55–80. [Google Scholar] [CrossRef]
- Magurran, A.E. Measuring Biological Diversity; Blackwell Publishing: Oxford, UK, 2004; 132p. [Google Scholar]
- Somerfield, P.; Clarke, K.; Warwick, R. Simpson Index. In Encyclopedia of Ecology; Elsevier: Amsterdam, The Netherlands, 2008; pp. 3252–3255. [Google Scholar]
- Lande, R. Statistics and Partitioning of Species Diversity and Similarity among Multiple Communities. Oikos 1996, 76, 5–13. [Google Scholar] [CrossRef]
- Aeschimann, D.; Rasolofo, N.; Theurillat, J.P. Analyse de la flore des Alpes. 2: Biodiversité et chorologie. Candollea 2011, 66, 225–253. [Google Scholar] [CrossRef]
- Biță-Nicolae, C.; Florescu, L.I.; Purice, D.; Kaya, O. Riparian woody plant communities in the Romanian Carpathians: Species diversity and community structure of Salix and Hippophaë communities. Ecol. Evol. 2024, 14, e11361. [Google Scholar] [CrossRef] [PubMed]
- Di Biase, L.; Pace, L.; Mantoni, C.; Fattorini, S. Variations in Plant Richness, Biogeographical Composition, and Life Forms along an Elevational Gradient in a Mediterranean Mountain. Plants 2021, 10, 2090. [Google Scholar] [CrossRef] [PubMed]
- Bhattarai, K.R.; Vetaas, O.R. Variation in plant species richness of different life forms along a subtropical elevation gradient in the Himalayas, east Nepal. Glob. Ecol. Biogeogr. 2003, 12, 327–340. [Google Scholar] [CrossRef]
- Giménez, E.; Melendo, M.; Valle, F.; Gómez-Mercado, F.; Cano, E. Endemic flora biodiversity in the south of the Iberian Peninsula: Altitudinal distribution, life forms and dispersal modes. Biodivers. Conserv. 2004, 13, 2641–2660. [Google Scholar] [CrossRef]
- Mota, G.S.; Luz, G.R.; Mota, N.M.; Coutinho, E.S.; Veloso, M.D.D.M.; Fernandes, G.W.; Nunes, Y.R.F. Changes in species composition, vegetation structure, and life forms along an altitudinal gradient of rupestrian grasslands in south-eastern Brazil. Flora 2018, 238, 32–42. [Google Scholar] [CrossRef]
- Bita-Nicolae, C.; Yildiz, F.; Kaya, O. Exploring the biodiversity and conservation value of alpine grasslands in the Bucegi Massif, Romanian Carpathians. Sustainability 2023, 15, 12643. [Google Scholar] [CrossRef]
- Médail, F.; Verlaque, R. Ecological characteristics and rarity of endemic plants from southeast France and Corsica: Implications for biodiversity conservation. Biol. Conserv. 1997, 80, 269–281. [Google Scholar] [CrossRef]
- Fenu, G.; Fois, M.; Cañadas, E.M.; Bacchetta, G. Using endemic-plant distribution, geology and geomorphology in biogeography: The case of Sardinia (Mediterranean Basin). Syst. Biodivers. 2014, 12, 181–193. [Google Scholar] [CrossRef]
- Kullman, L. A richer, greener and smaller alpine world: Review and projection of warming-induced plant cover change in the Swedish Scandes. AMBIO 2010, 39, 159–169. [Google Scholar] [CrossRef]
- Escolar, C.; Martínez, I.; Bowker, M.A.; Maestre, F.T. Warming reduces the growth and diversity of biological soil crusts in a semi-arid environment: Implications for ecosystem structure and functioning. Philos. Trans. R. Soc. B Biol. Sci. 2012, 367, 3087–3099. [Google Scholar] [CrossRef]
- Buri, A.; Grand, S.; Yashiro, E.; Adatte, T.; Spangenberg, J.E.; Pinto-Figueroa, E.; Verrecchia, E.; Guisan, A. What are the most crucial soil variables for predicting the distribution of mountain plant species? A comprehensive study in the Swiss Alps. J. Biogeogr. 2020, 47, 1143–1153. [Google Scholar] [CrossRef]
- Huston, M.A.; McBride, A.C. Evaluating the relative strengths of biotic versus abiotic controls on ecosystem processes. In Biodiversity and Ecosystem Functioning: Synthesis and Perspectives; Loreau, M., Naeem, S., Inchausti, P., Eds.; Oxford University Press: Oxford, UK, 2002; pp. 47–60. [Google Scholar]
- Callaway, R.M.; Walker, L.R. Competition and facilitation: A synthetic approach to interactions in plant communities. Ecology 1997, 78, 1958–1965. [Google Scholar] [CrossRef]
- Tarnocai, C.; Canadell, J.G.; Schuur, E.A.; Kuhry, P.; Mazhitova, G.; Zimov, S. Soil organic carbon pools in the northern circumpolar permafrost region. Glob. Biogeochem. Cycles 2009, 23, GB2023. [Google Scholar] [CrossRef]
- Larcher, W.; Kainmüller, C.; Wagner, J. Survival types of high mountain plants under extreme temperatures. Flora-Morphol. Distrib. Funct. Ecol. Plants 2010, 205, 3–18. [Google Scholar] [CrossRef]
- Sturm, M.; Racine, C.; Tape, K. Increasing shrub abundance in the Arctic. Nature 2001, 411, 546–547. [Google Scholar] [CrossRef]
- Chapin, F.S., III; Zavaleta, E.; Eviner, V.; Naylor, R.; Vitousek, P.; Reynolds, H.; Hooper, D.; Lavorel, S.; Sala, O.; Hobbie, S.; et al. Consequences of changing biodiversity. Nature 2000, 405, 234–242. [Google Scholar] [CrossRef]
- Anisimov, O.; Vaughan, D.G.; Callaghan, T.V.; Furgal, C.; Marchant, H.J.; Prowse, T.D.; Vilhjalmsson, H.; Walsh, J.E. Polar Regions (Arctic and Antarctic). In Climate Change 2007—Impacts, Adaptations and Vulnerability; Parry, M., Canziani, O., Palutikof, J., van der Linden, P., Hanson, C., Eds.; Cambridge University Press: Cambridge, UK, 2007; pp. 653–685. [Google Scholar]
- Epstein, H.E.; Raynolds, M.K.; Walker, D.A.; Bhatt, U.S.; Tucker, C.J.; Pinzon, J.E. Dynamics of aboveground phytomass of the circumpolar Arctic tundra during the past three decades. Environ. Res. Lett. 2012, 7, 015506. [Google Scholar] [CrossRef]
- Körner, C.; Hiltbrunner, E. Why is the alpine flora comparatively robust against climatic warming? Diversity 2021, 13, 383. [Google Scholar] [CrossRef]
- Sumner, E.E.; Venn, S.E. Thermal tolerance and growth responses to in situ soil water reductions among alpine plants. Plant Ecol. Divers. 2022, 15, 297–308. [Google Scholar] [CrossRef]
- Alexander, J.M.; Lembrechts, J.J.; Cavieres, L.A.; Daehler, C.C.; Haider, S.; Kueffer, C.; Liu, G.; McDougall, K.L.; Milbau, A.; Pauchard, A.; et al. Plant Invasions into Mountains and Alpine Ecosystems: Current Status and Future Challenges. Alp. Bot. 2016, 126, 89–103. [Google Scholar] [CrossRef]
- Hou, X.; Liu, S.; Zhao, S.; Dong, S.; Sun, Y.; Beazley, R. The alpine meadow around the mining areas on the Qinghai-Tibetan Plateau will degenerate as a result of the change of dominant species under the disturbance of open-pit mining. Environ. Pollut. 2019, 254, 113111. [Google Scholar] [CrossRef] [PubMed]
- Steiger, R.; Knowles, N.; Pöll, K.; Rutty, M. Impacts of climate change on mountain tourism: A review. J. Sustain. Tour. 2024, 32, 1984–2017. [Google Scholar] [CrossRef]
- Chapin III, F.S.; Sturm, M.; Serreze, M.C.; McFadden, J.P.; Key, J.R.; Lloyd, A.H.; Welker, J.M. Role of land-surface changes in Arctic summer warming. Science 2005, 310, 657–660. [Google Scholar] [CrossRef] [PubMed]
- Tape, K.; Sturm, M.; Racine, C. The evidence for shrub expansion in Northern Alaska and the Pan-Arctic. Glob. Change Biol. 2006, 12, 686–702. [Google Scholar] [CrossRef]
- Callaghan, T.V.; Björn, L.O.; Chernov, Y.; Chapin, T.; Christensen, T.; Huntley, B.; Ims, R.A.; Johansson, M.; Jolly, D.; Jonasson, S.; et al. Biodiversity, Distributions and Adaptations of Arctic Species in the Context of Environmental Change. AMBIO 2004, 33, 404–417. [Google Scholar] [CrossRef] [PubMed]
- Haskell, D.E.; Flaspohler, D.J.; Webster, C.R.; Meyer, M.W. Variation in soil temperature, moisture, and plant growth with the addition of downed woody material on lakeshore restoration sites. Restor. Ecol. 2012, 20, 113–121. [Google Scholar] [CrossRef]
- Leng, X.; Cui, J.; Zhang, S.; Zhang, W.; Liu, Y.; Liu, S.; An, S. Differential water uptake among plant species in humid alpine meadows. J. Veg. Sci. 2013, 24, 138–147. [Google Scholar] [CrossRef]
- Li, C.; Peng, F.; Xue, X.; You, Q.; Lai, C.; Zhang, W.; Cheng, Y. Productivity and quality of alpine grassland vary with soil water availability under experimental warming. Front. Plant Sci. 2018, 9, 1790. [Google Scholar] [CrossRef]
- Nagy, L.; Grabherr, G. The Biology of Alpine Habitats; Oxford University Press: Oxford, UK, 2009. [Google Scholar]
- Su, X.; Han, W.; Liu, G.; Zhang, Y.; Lu, H. Substantial gaps between the protection of biodiversity hotspots in alpine grasslands and the effectiveness of protected areas on the Qinghai-Tibetan Plateau, China. Agric. Ecosyst. Environ. 2019, 278, 15–23. [Google Scholar] [CrossRef]
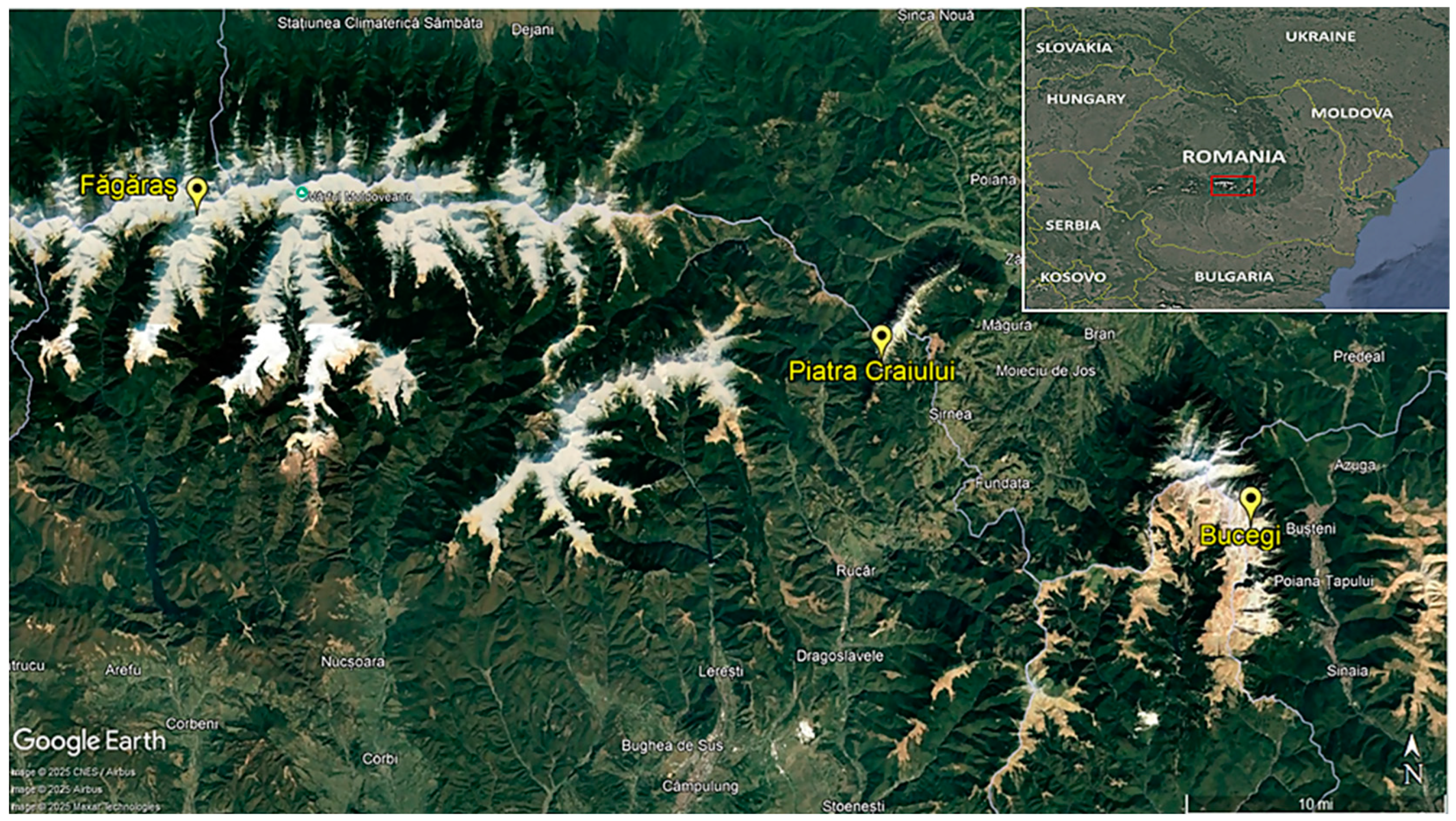




{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| B | F | PC | |
|---|---|---|---|
| Th | 6.818182 | 6.666667 | 7.407407 |
| H | 72.72727 | 80 | 75.92593 |
| Ch | 20.45455 | 6.666667 | 14.81481 |
| G | 0 | 6.666667 | 1.851852 |
| B | F | PC | |
|---|---|---|---|
| B | 0.2361 | 0.0006 | |
| F | 0.1867 | 0.0003 | |
| PC | 0.617 | 0.82 |
| Taxon | Average Dissim. | Contrib. % | Cumulative % | Mean B | Mean F | Mean PC |
|---|---|---|---|---|---|---|
| C. lerchenfeldianum | 26.03 | 31.68 | 31.68 | 11.9 | 24.2 | 0.143 |
| P. corona-sancti-stephani | 20.38 | 24.8 | 56.47 | 12.5 | 20.6 | 3 |
| Eigenvalue | % Variance | PCA Loadings | |||||
|---|---|---|---|---|---|---|---|
| M | N | R | L | T | |||
| PC 1 | 2.85192 | 54.412 | 0.44796 | 0.41877 | 0.47796 | 0.42954 | 0.45936 |
| PC 2 | 0.84244 | 16.073 | −0.25715 | 0.54711 | −0.69562 | 0.36361 | 0.13578 |
| PC 3 | 0.60274 | 11.5 | 0.45362 | 0.58114 | −0.07595 | −0.56146 | −0.36811 |
| PC 4 | 0.48017 | 9.1612 | −0.4458 | 0.31695 | 0.45728 | 0.31633 | −0.62581 |
| PC 5 | 0.464091 | 8.8544 | 0.57332 | −0.29516 | −0.26979 | 0.51767 | −0.49337 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Biță-Nicolae, C.; Mogîldea, D.; Pop, O.G. Investigating Endemic Alpine Communities of Papaver corona-sancti-stephani and Cerastium lerchenfeldianum in the Southern Carpathians. Diversity 2025, 17, 283. https://doi.org/10.3390/d17040283
Biță-Nicolae C, Mogîldea D, Pop OG. Investigating Endemic Alpine Communities of Papaver corona-sancti-stephani and Cerastium lerchenfeldianum in the Southern Carpathians. Diversity. 2025; 17(4):283. https://doi.org/10.3390/d17040283
Chicago/Turabian StyleBiță-Nicolae, Claudia, Daniela Mogîldea, and Oliviu G. Pop. 2025. "Investigating Endemic Alpine Communities of Papaver corona-sancti-stephani and Cerastium lerchenfeldianum in the Southern Carpathians" Diversity 17, no. 4: 283. https://doi.org/10.3390/d17040283
APA StyleBiță-Nicolae, C., Mogîldea, D., & Pop, O. G. (2025). Investigating Endemic Alpine Communities of Papaver corona-sancti-stephani and Cerastium lerchenfeldianum in the Southern Carpathians. Diversity, 17(4), 283. https://doi.org/10.3390/d17040283