
Allosaurus europaeus (Theropoda: Allosauroidea) Revisited and Taxonomy of the Genus



Abstract
1. Introduction
2. Geological Context

- Praia de Vale de Frades north (top of Praia Azul Mb.). The fossil locality altitude varies between −1 and 10 m relative to sea level. The faunal list is:
- Transgressive layer (last of Praia Azul Mb.), dominated by the bivalves Jurassicorbula edwardi (Sharpe, 1850), with some Isognomon lusitanicus and fragments of Plesiochelidae turtles, and charcoal fragments.
- Allosaurus europaeus Mateus et al., 2006 (type specimen ML415), from the gray sandstone channel, 2–3 m below the guide transgressive layer.
- Camarasauridae sacral neural spine fragment (ML specimen, previously under ML415b).
- Vale de Frades (top of the cliff, Santa Rita Mb.), base of the sandstone channel at the road, cut at the coordinates 39°16′31″ N, 9°20′00″ W (~36 m alt.).
- Draconyx loureiroi Mateus and Antunes, 2003 (type specimen).
- Plesiochelyidae indet (specimen ML).
- Praia de Vale de Frades south (new locality) is a terrestrial sandstone channel at 39°16′22″ N, 9°20′08″ W (3 m alt.), with Tracheophytes in situ roots and Lycopodiopsida megaspores (Santa Rita Mb.).
3. Materials and Methods
4. Systematic Paleontology
| Dinosauria Owen, 1842 |
| Saurischia Seeley, 1887 |
| Theropoda Marsh, 1881 |
| Neotheropoda Bakker, 1986 |
| Tetanurae Gauthier, 1986 |
| Avetheropoda Paul, 1988 |
| Carnosauria Huene, 1920 |
| Allosauroidea Currie and Zhao, 1994 |
| Allosauridae Marsh, 1878 |
| Allosaurus Marsh, 1877 |
| Allosaurus europaeus Mateus et al., 2006 |
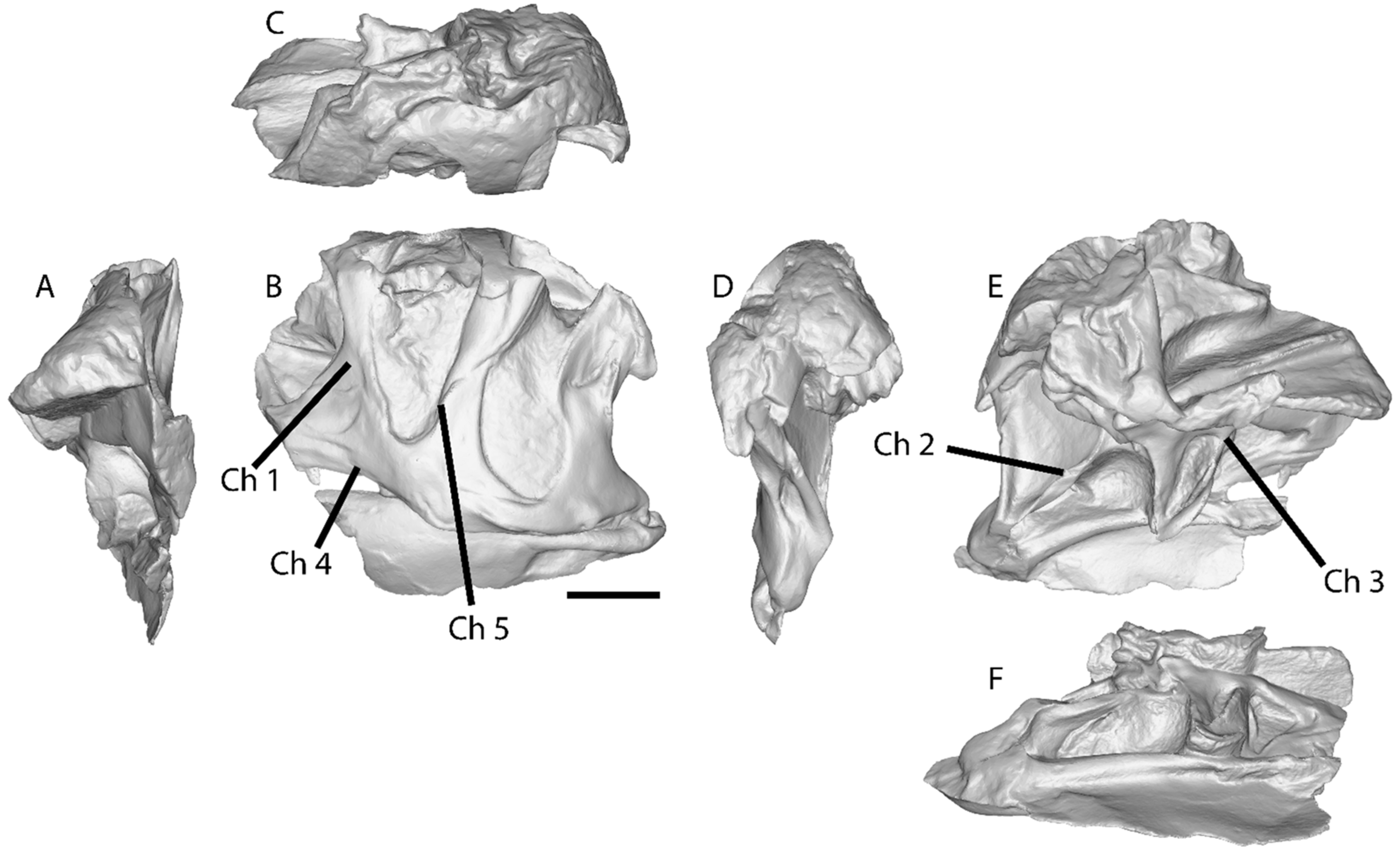
- Lacrimal anterior projections contacting nasally, posteriorly, ventrally, and dorsally (i.e., the lateroposterior tip of the nasal projection projects ventrally to the lacrimal dorsoanterior projection; Ch. 1).
- Lacrimal ventral ramus, lateroanterior vertical projection, restricted to the dorsal half of the antorbital fenestra (aof), and not continuous to the medial vertical ridge that borders the aof (Ch. 3).
- Pterygoid contact with the quadrate ventral shelf of the pterygoid flange, with participation of the pterygoid in the fold that constitutes the shelf (Ch. 6; see Hendrickx et al., 2015 [32] for quadrate topology).
- Pterygoid contact with the ectopterygoid, with prominent ventral projection and anterior notch (Ch. 7).
- The maxilla ventroposterior end is step-like (nearly vertical contact; Ch. 8).
- Postorbital, ventral termination of ventral ramus close to the ventral margin of the orbit and ventral to squamosal–quadratojugal contact (plesiomorphic, shared with Sinraptor, but is a local autapomorphy different from other Allosaurus; Ch. 10).
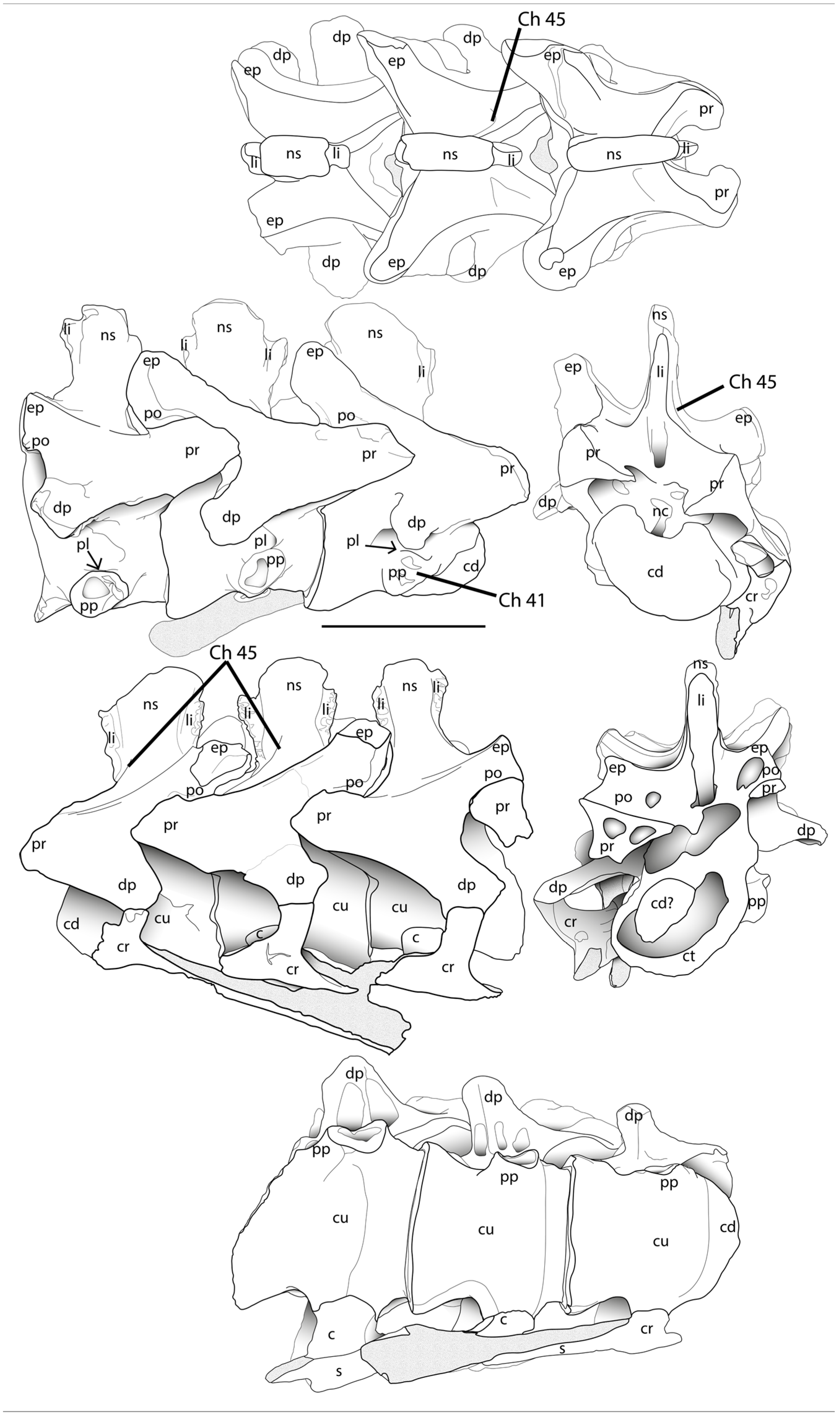
- Cervical vertebrae, position of parapophyses on the centrum in the middle (Ch. 43; shared with Neovenator; Ch. 41).
- Fourth and fifth cervical vertebrae, latero-anterior base of the neural spine with oblique-to-vertical subtle accessory lamina at the posterior end of the spinoprezygapophyseal lamina (best seen in lateral–dorsal views; Ch. 45).
- Shape of postorbital contact with jugal sigmoidal bone (Ch. 46).
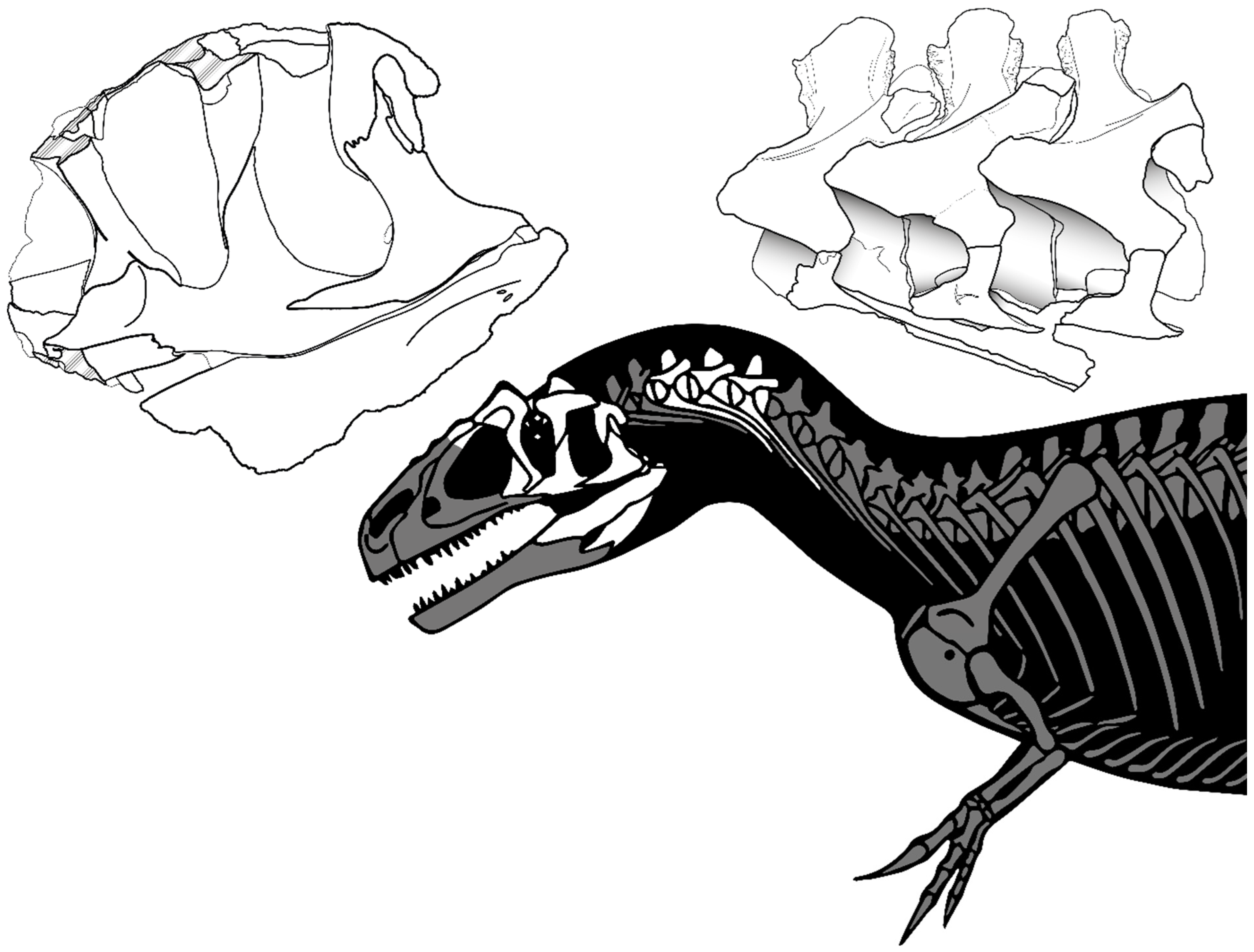
5. Description
5.1. General Description of the Skull and Mandible
5.1.1. Major Cranial Fenestrae, Foramina, and Fossae

5.1.2. Bones of the Dermatocranium




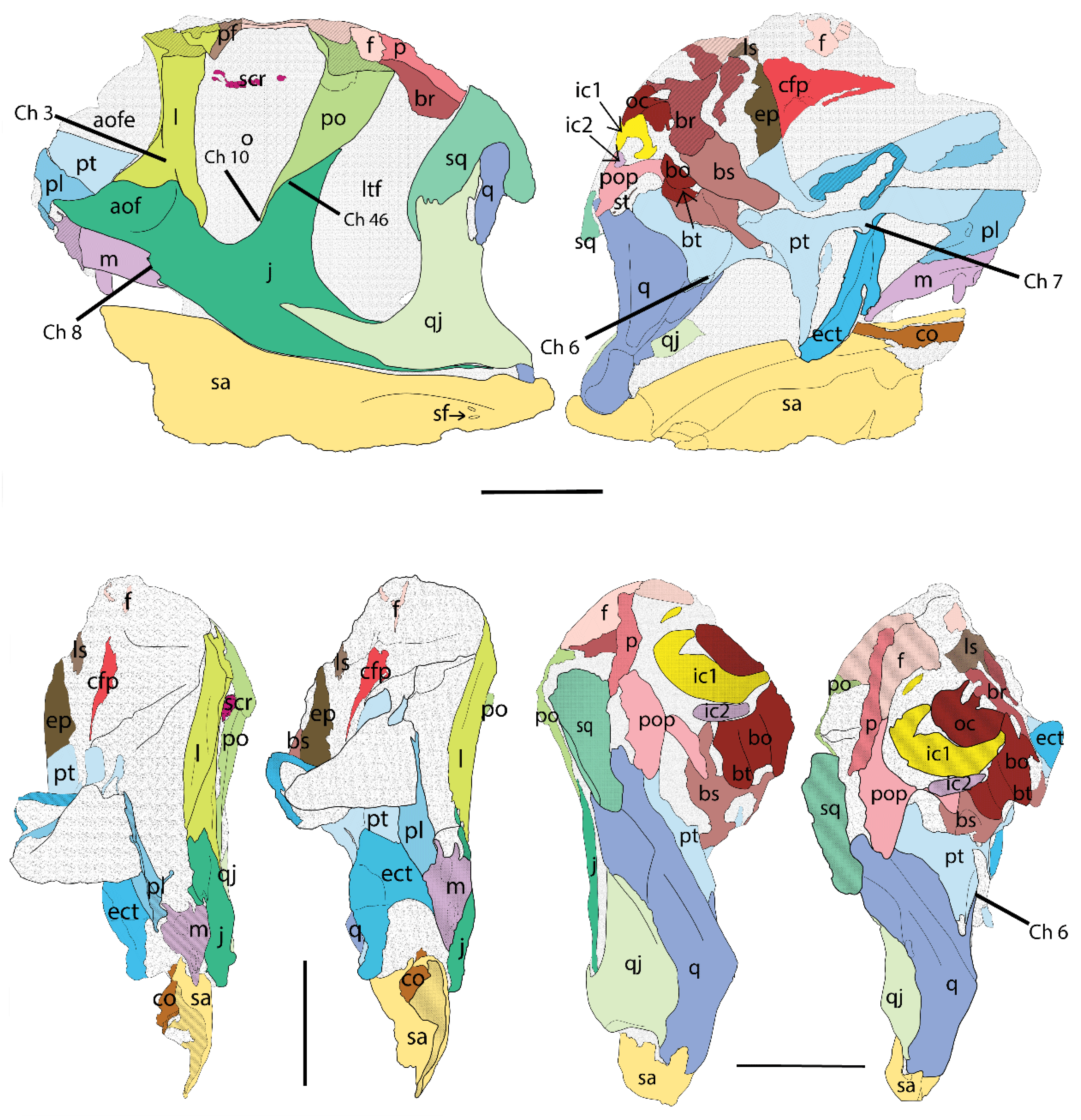
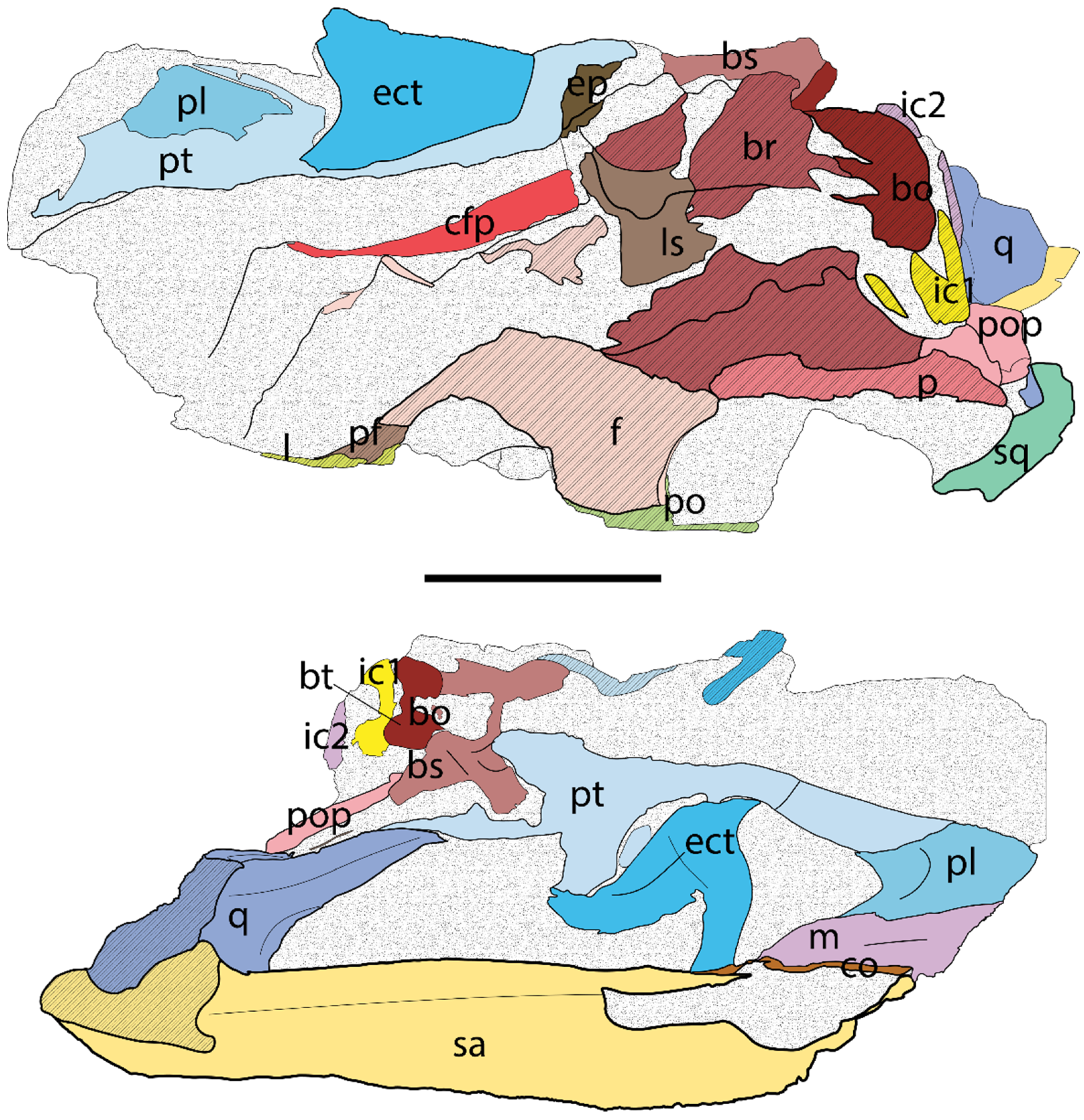


5.1.3. Bones of the Chondrocranium
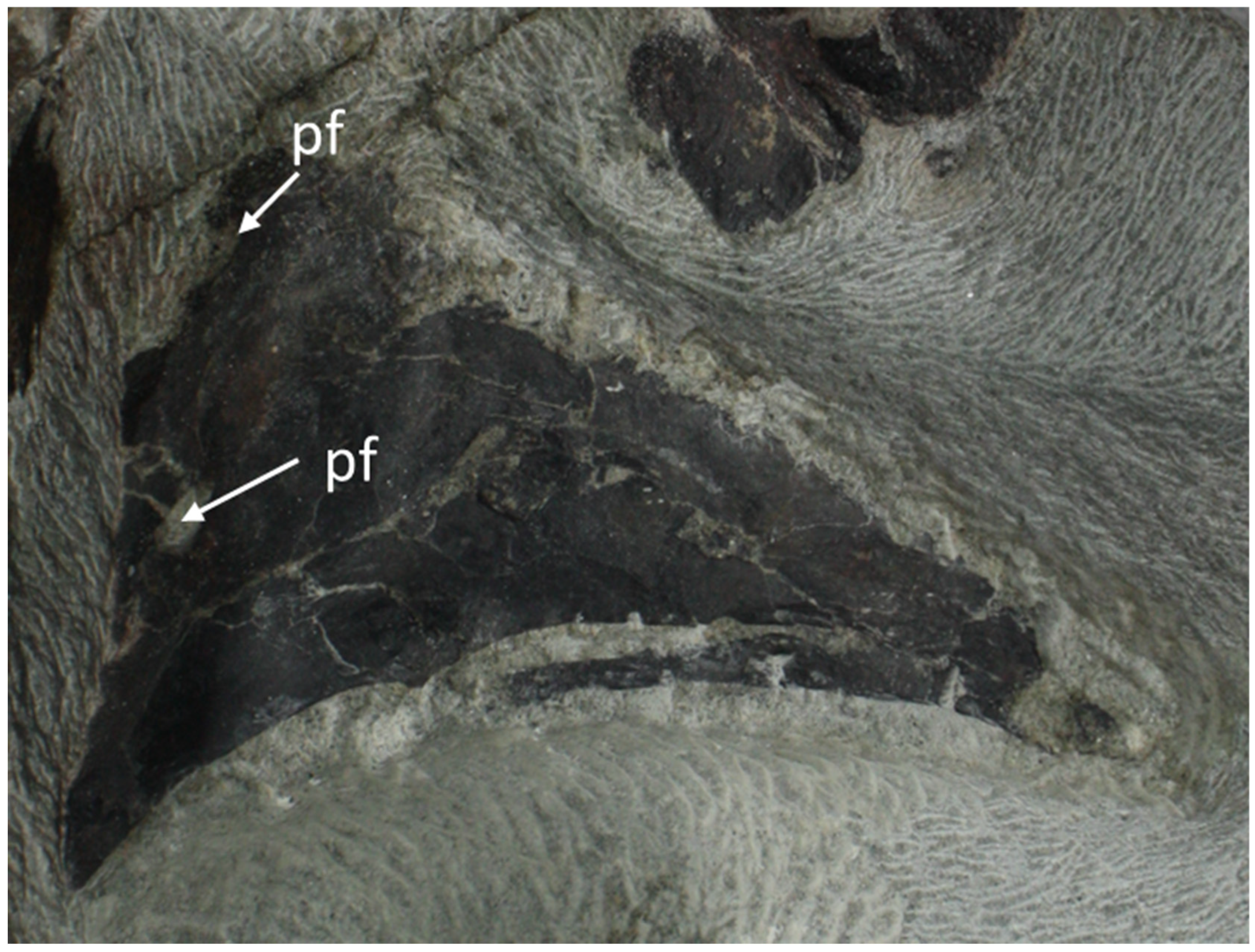
5.1.4. Bones of the Splanchnocranium


5.1.5. Dermal Bones of the Lower Jaw

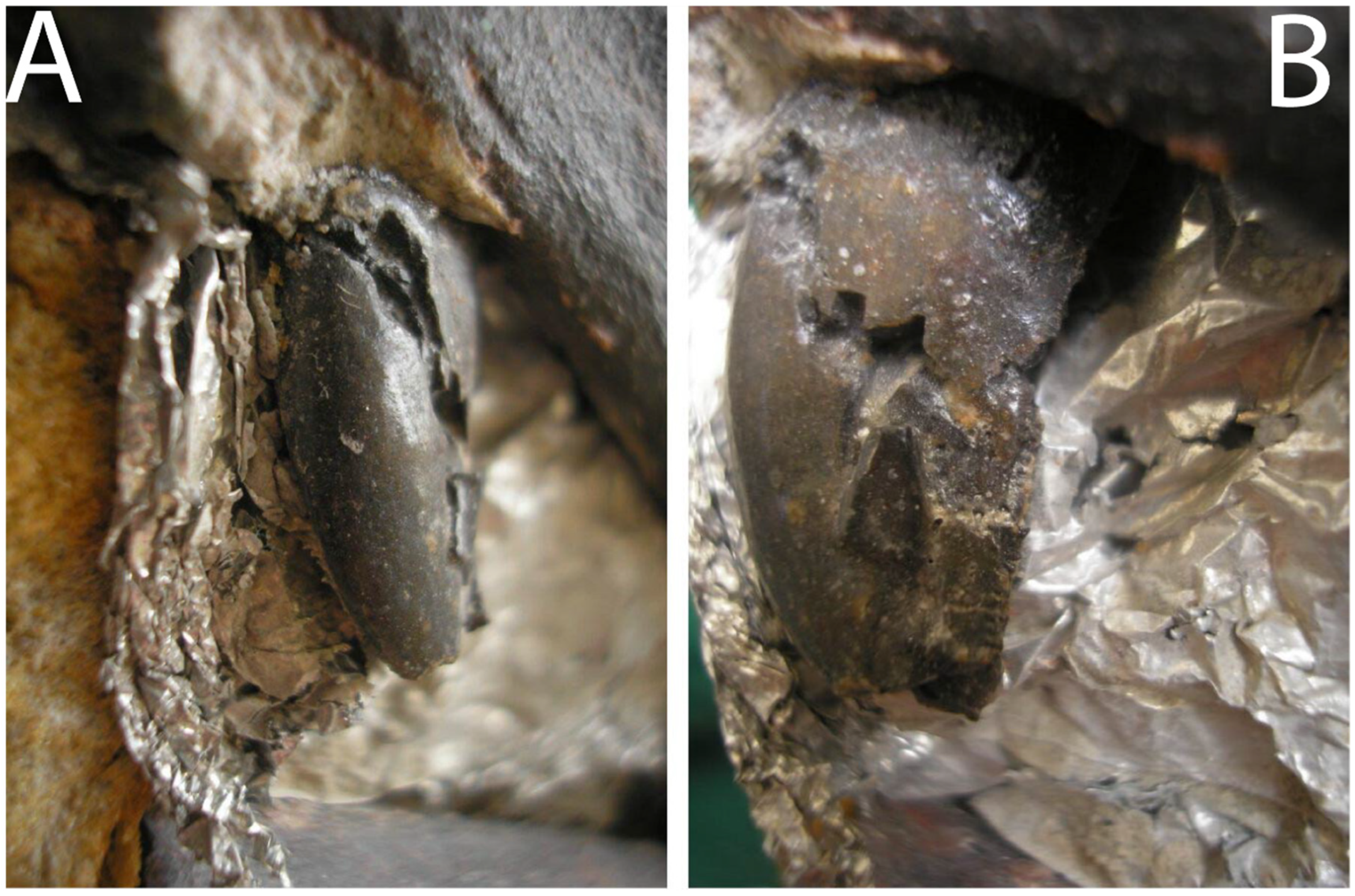
5.1.6. Dentition
5.2. Description of the Cervical Vertebrae and Ribs

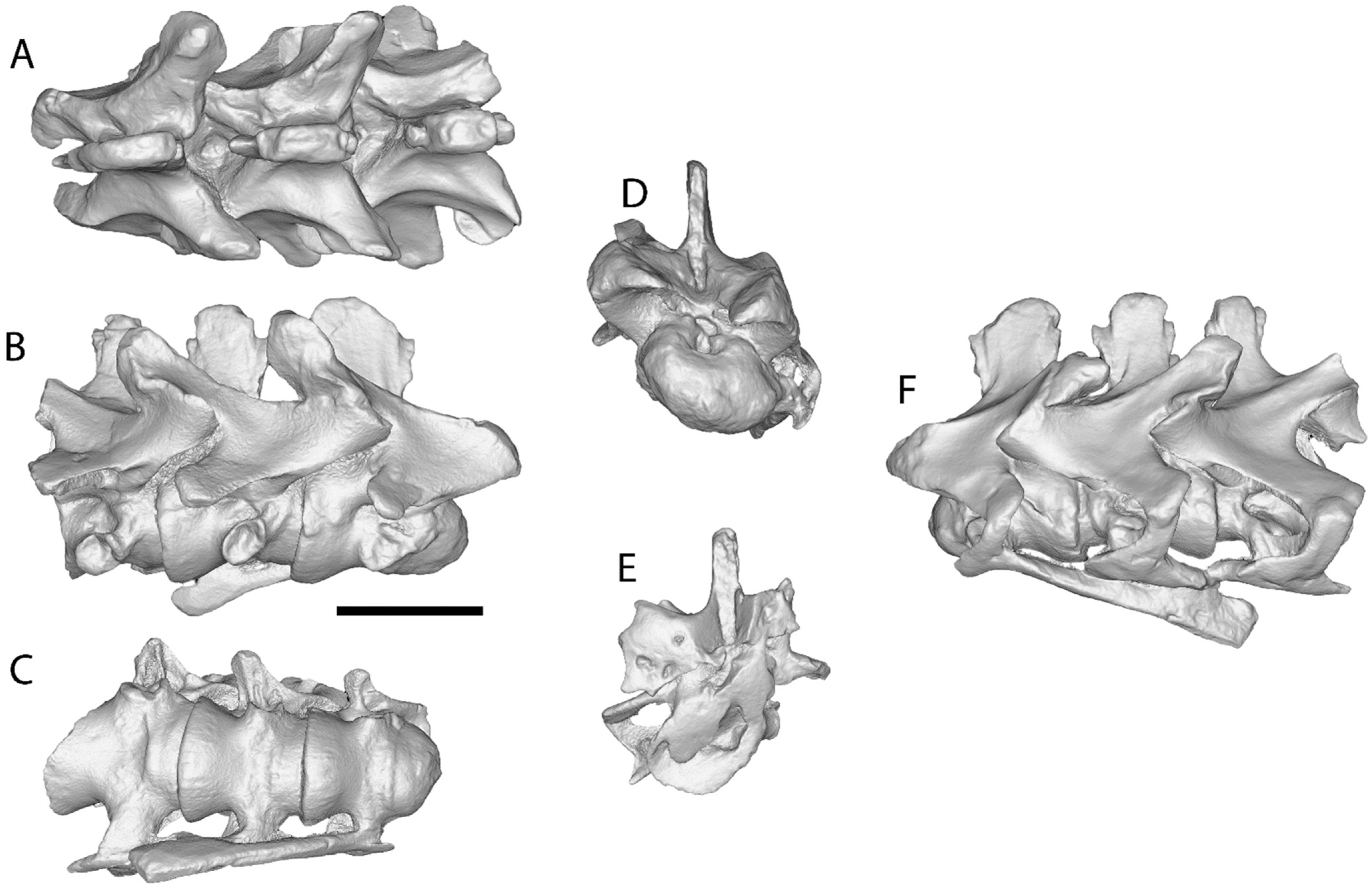



6. Discussion
6.1. Validity of Other Allosaurus and Allosaurus-like Species
- Allosaurus fragilis Marsh 1878
- Allosaurus lucasi Dalman 2014 (nomen dubium)
- Labrosaurus ferox Marsh, 1884 (Figure 24)

- “Allosaurus” tendagurensis Janensch, 1925 (incerta sedis)
- Allosaurus lucaris (Marsh 1878), originally Labrosaurus.
- Apatodon mirus Marsh, 1877 Nomen dubium (Dinosauria incertae sedis)
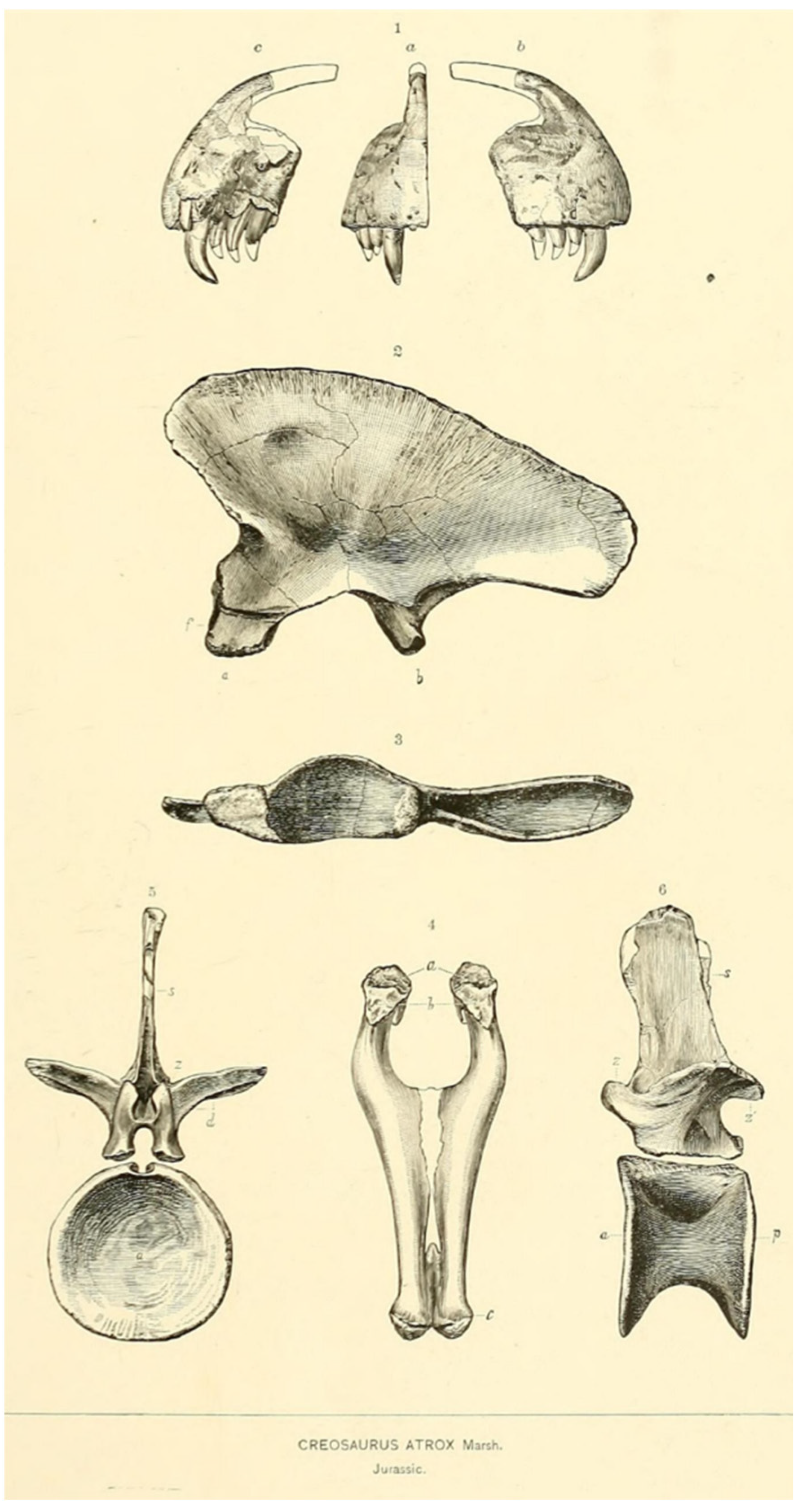
- Epanterias amplexus Cope, 1878
- Laelaps trihedrodon (Cope, 1877) Nomen dubium
- Antrodemus valens (Leidy, 1870) Leidy, 1873 (originally Poekilopleuron)
- Camptonotus amplus Marsh, 1879
- Saurophaganax maximus (Chure, 1995), originally nomen nudum Saurophagus maximus Stovall, 1941
6.2. Ontogeny and Size Comparisons
6.3. Taphonomy
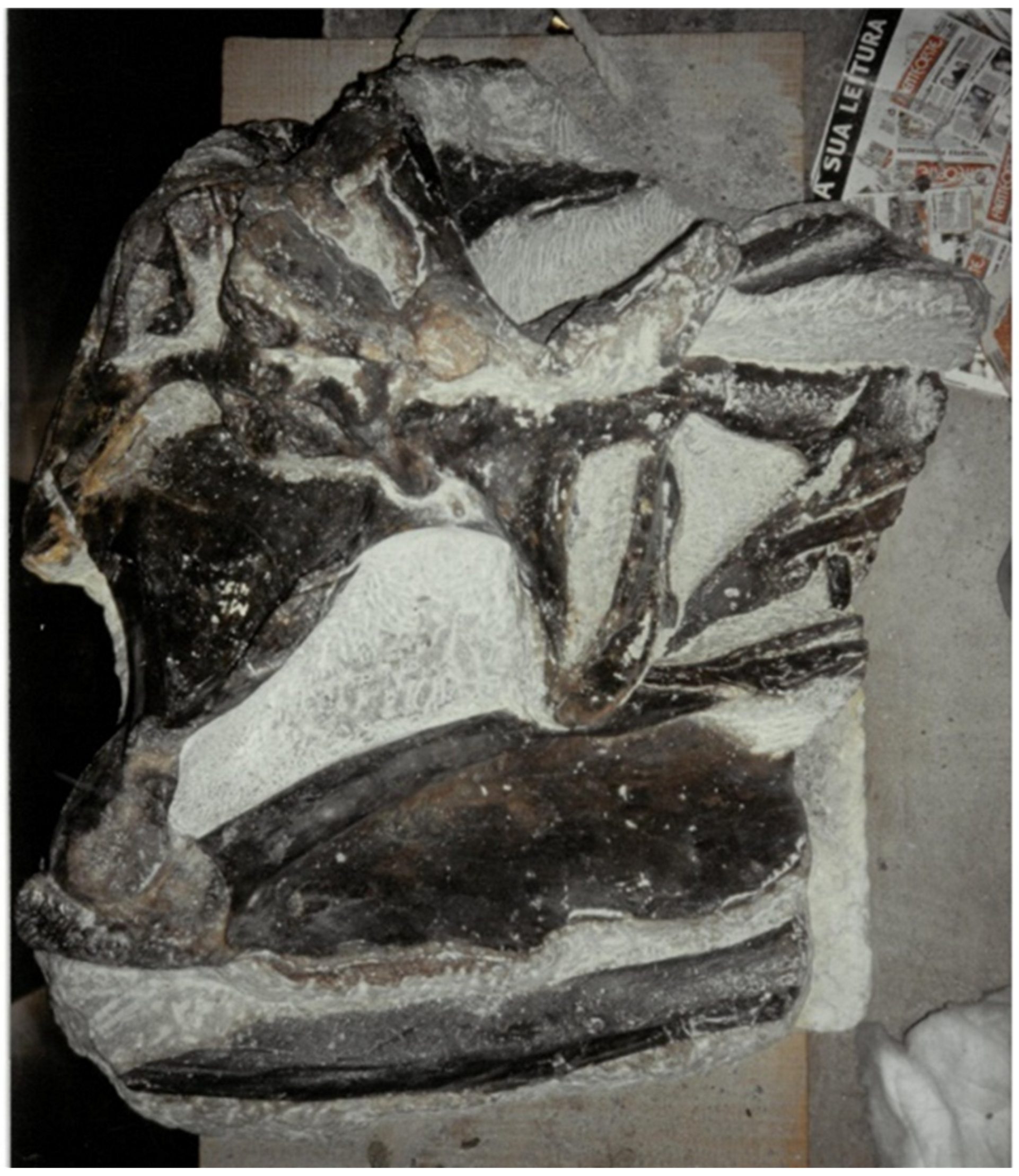
6.4. Comparisons of Allosaurus europaeus ML415
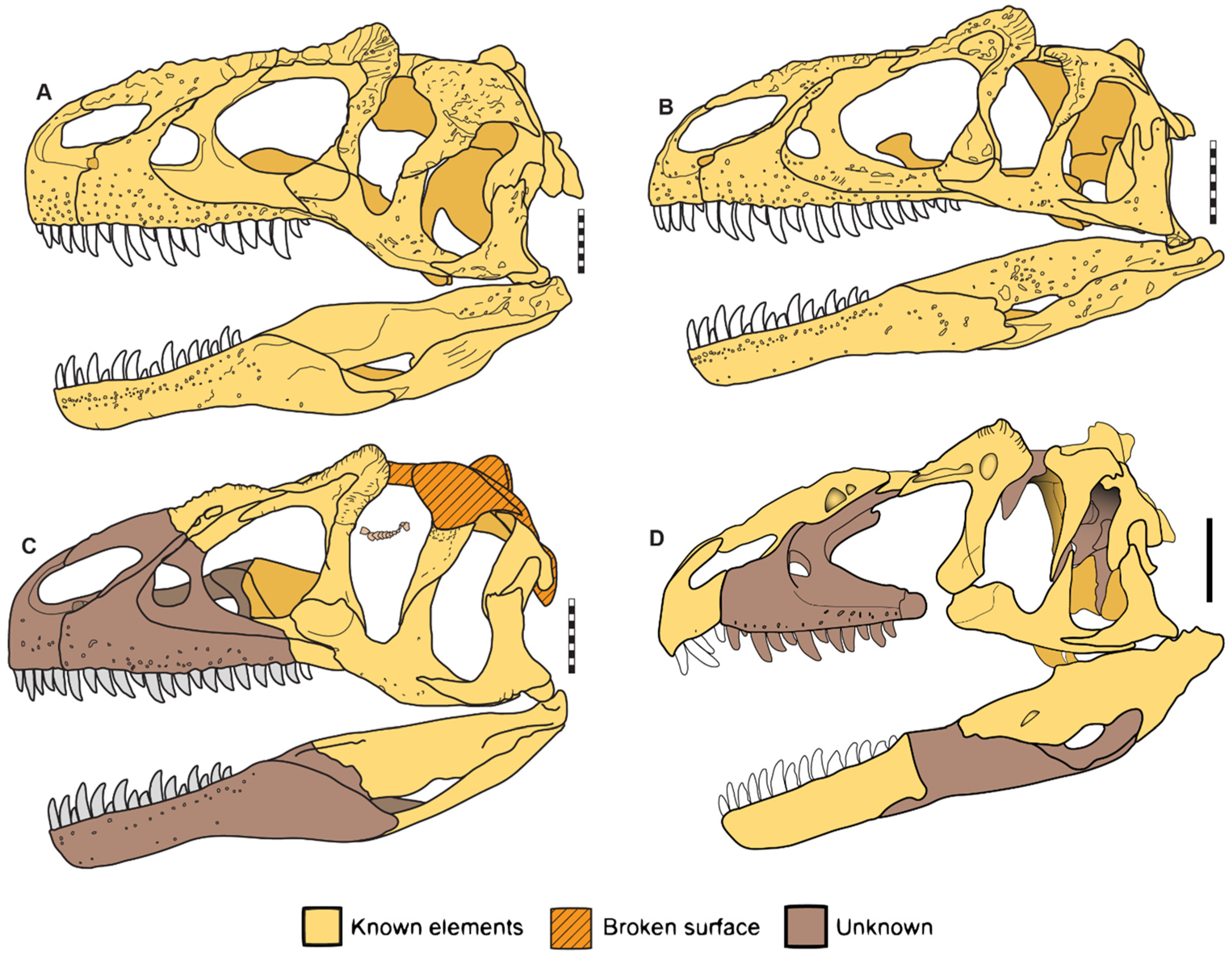
7. Phylogeny
7.1. General Phylogenetic Analysis
- ML_415
- ‘Allosaurus_jimmadseni’
- Ch. 7: Maxilla, promaxillary fenestra, lateral exposure is present, but completely obscured by the lateral lamina of the ascending ramus in A. fragilis and A. jimmadseni and unknown in A. europaeus (Ch. 7 from Eddy and Clarke, 2011 [30]).
- Ch. 33: Jugal bone, lateral view, relative heights of quadratojugal prongs, the dorsal prong is shorter or equal in height in all species of Allosaurus (Ch. 33 from Eddy and Clarke, 2011 [30]).
- Ch. 36: Jugal bone, lateral view, accessory pneumatization of the antorbital fossa is absent or shallow, shared across Allosaurus species (Ch. 36 from Eddy and Clarke, 2011 [30]).
- Ch. 47: Prefrontal bone, medial view, shape of frontal bone articular surface is triangular in A. fragilis and A. jimmadseni, unknown in A. europaeus (Ch. 47 from Eddy and Clarke, 2011 [30]).
- Ch. 55: Parietal bone, posteriorly placed, knob-like dorsal projection, form is absent or very low (0) in A. fragilis and A. jimmadseni, and is unknown in A. europaeus (Ch. 55 from Eddy and Clarke, 2011 [30]).
- Ch. 84: Palatine bone, pneumatic recess, form is absent or small in all Allosaurus species (Ch. 84 from Eddy and Clarke, 2011 [30]).
- Ch. 185: Lacrimal bone, horn morphology has a triangular horn across all Allosaurus species (Ch. 185, 47 from Carrano et al., 2012 [2]).
- “Allosaurus_europaeus”
- “Allosaurus_jimmadseni”
7.2. Discussion of Phylogenetic Characters
7.3. New Restricted Phylogenetic Analysis for Allosaurus (Table 1)
- Maxila, lateral aspect, medioventral wall of the maxillary antorbital fossa: smooth (0); a row of neurovascular foramina (1) (Ch. 36).
- Maxilla: straight posteroventral jugal ramus of maxilla, where it articulates with jugal bone (0); curved (1) (Ch. 37).
- Laterodorsal margin of nasal bone “pinched” into the low crest, continuous from premaxilla to lacrimal bone: absent (0); present (1) (Ch. 38).
- Posterior portion of the dorsal surface of nasal bone, cup-shaped, producing a median peak in the region of nasofrontal contact: absent (0); present (1) (Ch. 39).
- Lacrimal horns: absent or reduced (0); pronounced (as in Allosaurus fragilis [12]) (1); very pronounced (20% or more in the lacrimal bone height, in lateral view) (2) (Ch. 40).
- Jugal bone with a relatively ventral margin: straight-to-slightly-curved outline in dorsal view (0); curved sinosoidal (1) (Ch. 41).
- Lacrimal anterior projections contacting the lacrimal bone portion overlapping nasal bone dorsally: Contacts posteriorly, ventrally, and dorsally (i.e., the lateroposterior tip of the nasal bone projects ventral to the lacrimal dorsoanterior projection). It is seen in both A. europaeus and USNM 4734 specimens, it is unknown in Allosaurus of Andrés since the publish lacrimal bone is disarticulated, and in other specimens, the lacrimal bone contacts only posteriorly and ventrally to nasal bone—NEW (Ch. 1).
- Lacrimal vacuity is single in Sinraptor dongi [36], A. europaeus, and DINO 2560 and clearly divided by a vertical strut in Allosaurus of Andrés and in USNM 4734. A. jimmadseni share both states. This character may not be significant since Carpenter, 2010 [33], states that this characteristic is within the intra-specific variation—NEW (Ch. 2).
- Lacrimal ventral ramus lateroanterior vertical projection is restricted to the dorsal half of the antorbital fenestra (aof) and not continuous to the medial vertical ridge that borders the aof in A. europaeus. In pictures published of Allosaurus of Andrés lacrimal bone, it is hard to observe, but it seems similar to ML415, so it is considered that these share the same characteristic. Nevertheless, with further detailed study of the Andrés specimen, this may be subject to change. For the other specimens of Allosaurus and Sinraptor dongi [36], this is continuous. In Neovenator salerii, the lacrimal bone is not reported. If proven to be true in Portuguese taxa, it may represent an autapomorphy—NEW (Ch. 3).
- Nasal and dorsal ramus of maxilla: posterior end (dorsal portion in lateral view, without the skull roof) in DINO 2560, and the nasal bone and maxilla dorsal ramus end are at the same level or close, whereas on the other taxa, the nasal bone end is clearly posterior to the maxilla dorsal ramus—NEW (Ch. 4).
- Nasal pneumatic foramina size in USNM 4734: the anterior foramen is larger than the posterior, but in A. europaeus and A. jimmadseni, the posterior foramen is larger than the anterior, while in DINO 2560 it is not applicable (no foramina present, single foramen, or two of the same size, score “?”)—NEW (Ch. 5).
- Pterygoid bone contact with the quadrate ventral shelf of the pterygoid flange (see Hendricks et al., 2015 [32], for topology) is unique and autapomorphic in A. europaeus due to the participation of the pterygoid bone in the fold that constitutes the shelf—NEW (Ch. 6).
- Pterygoid bone contact with the ectopterygoid is autapomorphic in A. europaeus, with prominent ventral projection and anterior notch—NEW (Ch. 7).
- Maxilla ventroposterior end is autapomorphic in A. europaeus, with a step-like, nearly vertical contact—NEW (Ch. 8).
- Dentary, the posterior end is bifurcated in A. jimmadseni, potentially autapomorphic for this taxon, whereas in DINO 2560, it is single pointed, but it is not known in Neovenator salerii and other specimens of Allosaurus—NEW (Ch. 42).
- Nasal bone proportions (total length/maximum width measured at the anterior-most end of prefrontal) are 9 or less in Sinraptor dongi and in A. fragilis (USNM 4734) but 10 or more in A. jimmadseni and in DINO 2560. In A. europaeus it is unknown, and in Andrés, the nasal bone is not reported—NEW (Ch. 43).
- The angular dorsoposterior rim (contact with surangular bone)/minimum depth at mid-length is 5 or more (1) in A. europaeus and A. jimmadseni. This condition is unknown in USNM 4734 since it is not preserved, and in the Andrés specimen, the angular bone is not reported. In Sinraptor dongi and in DINO 2560, it is 4 or less (0)—NEW (Ch. 44).
- For the 4th and 5th cervical vertebrae, the latero-anterior base of the neural spine with oblique-to-vertical subtle accessory lamina at the posterior end of the spinoprezygapophyseal lamina (best seen in lateral–dorsal views) is autapomorphically present in ML415 but not in other taxa, and it is not reported in the Andrés specimen. (1)—NEW (Ch. 45).
- The shape of postorbital bone contact with the jugal bone is sigmoidal, with a clear, curved outline, whereas in A. fragilis (USNM 4734 and DINO 2560) and A. jimmadseni (types), it is straighter—NEW (Ch. 46).

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Sinraptor dongi | Neovenator salerii | Allosaurus europaeus ML415 | Allosaurus fragilis USNM4734 | Allosaurus jimmadseni types | Allosaurus fragilis DINO 2560 | Allosaurus Andrés | |
|---|---|---|---|---|---|---|---|
| 1. Lacrimal bone anterior projections contacting nasal lacrimal bone contacts only posteriorly and ventral to nasal (0); lacrimal bone portion overlapping nasal dorsally: contacts posteriorly, ventrally, and dorsally (i.e., the lateroposterior tip of the nasal bone projects ventral to the lacrimal dorsoanterior projection) (1). NEW | 0 | ? | 1 | ? | 0 | 0 | ? |
| 2. Lacrimal vacuity absent (0); single (1); clearly divided by a vertical strut (2). NEW | 0 | ? | 1 | 2 | 1/2 | 1 | 2 |
| 3. Lacrimal ventral ramus lateroanterior vertical projection: continuous to the medial vertical ridge that borders the aof (0); restricted to the dorsal half of the aof and not continuous to the medial vertical ridge that borders the aof (1). NEW | 0 | ? | 1 | 0 | 0 | 0 | 1 |
| 4. Nasal and dorsal ramus of maxilla, posterior end (dorsal portion in lateral view, without the skull roof) and nasal end clearly posterior to maxilla dorsal ramus (0); nasal and maxilla dorsal ramus end at the same level or close (1). NEW | 0 | 0 | 0 | ? | 0 | 1 | ? |
| 5. Nasal pneumatic foramina size: anterior foramen larger than posterior (0); posterior foramen larger than anterior (1); scored “?” when not applicable (no foramina present, single foramen, or two of the same size). NEW | 0 | ? | 1 | 0 | 1 | ? | ? |
| 6. Pterygoid bone contact with quadrate ventral shelf of pterygoid flange (see Hendricks et al., 2015 [32], for topology): no participation of pterygoid bone (0); with participation of pterygoid bone in the fold that constitutes the shelf (1). NEW | ? | ? | 1 | ? | 0 | 0 | ? |
| 7. Pterygoid bone contact with the ecpt: continuous (no obvious ventral projections) (0); with prominent ventral projection and anterior notch (1). NEW | 0 | ? | 1 | ? | 0 | 0 | ? |
| 8. Maxilla ventroposterior end: acute/tapering (ventroposteriorly inclined) (0); step-like (nearly vertical contact) (1). NEW | 0 | 0 | 1 | 0 | 0 | 0 | ? |
| 9. Cervical vertebrae, posterior articular face of mid-cervical centra, width: approximately as broad as tall (0); at least 20% broader than tall (1). (Ch. 110 [22]: p. 62, modified from [20]: p. 53) | 0 | 0 | 1 | 0 | 0 | 0 | ? |
| 10. Postorbital, ventral termination of ventral ramus: close to ventral margin of the orbit and ventral to squamosal–quadratojugal bone contact (0); dorsal to ventral margin of orbit and at same height or dorsal to squamosal–quadratojugal bone contact (1). (Ch. 41, modified from [21]: p. 7) | 0 | ? | 0 | 1 | 1 | 1 | ? |
| 11. Nasal, development of dorsolateral surfaces: none, nasals low and dorsally convex (0); pronounced dorsolateral rims, sometimes with lateral crests (1); tall, parasagittal crests (2); inflated and forming a hollow midline crest (3). (Ch. 40) | 0 | 1 | 2 | 1 | 2 | 1 | ? |
| 12. Maxilla, promaxillary fenestra, lateral exposure: absent, no fenestra (0); partially or fully exposed (1); present, but completely obscured by the lateral lamina of the ascending ramus (2). (Ch. 7, modified from [22]: p. 6; ordered) | 1 | 0 | ? | ? | 2 | 2 | ? |
| 13. Maxilla, curvature of posterior ramus: straight (0); curved, ventral deflection does not surpass ventral margin of largest maxillary tooth (1); curved, ventral deflection equal to or surpasses ventral margin of largest maxillary tooth (2). (Ch. 15, modified from [12]: p. 32; ordered) | 0 | ? | ? | ? | 1 | 1 | ? |
| 14. Lacrimal bone, suborbital process along posterior margin of ventral ramus: absent (0); present (1). (Ch. 30, modified from [12]: p. 79) | 1 | ? | 0 | 0 | 1 | 0 | 0 |
| 15. Jugal bone, lateral view, relative heights of quadratojugal bone prongs: dorsal prong shorter or equal in height (0); taller (1) than ventral prong. (Ch. 33) | 1 | ? | 0 | 0 | 0 | 0 | ? |
| 16. Jugal bone, lateral view, accessory pneumatization of the antorbital fossa: absent or shallow (0); extensive, invaginated recess (1). (Ch. 36, modified from [22]: p. 28; [21]: p. 12) | 1 | ? | 0 | 0 | 0 | 0 | ? |
| 17. Prefrontal, medial view, shape of frontal bone articular surface: triangular (0); rounded (1). (Ch. 47). | 1 | ? | ? | ? | 0 | 0 | ? |
| 18. Braincase, supraoccipital, width of dorsal expansion: less than twice the width (0); more than twice the width (1) of the foramen magnum. (Ch. 62, wording modified from [13]: p. 97; [45]: p. 13). | 1 | ? | ? | 0 | 0 | 0 | 0 |
| 19. Braincase, angle between occipital condyle and basal tubera: perpendicular, at or near 90° (0); acute (1). (Ch. 64, wording modified from [13]: p. 99; [45]: p. 7). | 1 | ? | 0 | 0 | 0 | 0 | 0 |
| 20. Braincase, shape of occipital condyle: subspherical (0); dorsoventrally compressed (1). (Ch. 65, wording modified from [45]: p. 15) | 1 | ? | 0 | 0 | 0 | 0 | 0 |
| 21. Braincase, form of trigeminal foramen (cranial nerve V) exit: single (0); fully split (1). (Ch. 69, wording modified from [13]: p. 108) | 0 | ? | ? | ? | 1 | ? | ? |
| 22. Pterygoid, medial view, fossae penetrating the quadrate and ectopterygoid rami: absent (0); present (1). (Ch. 77) | 1 | ? | 0 | ? | 0 | 0 | ? |
| 23. Pterygoid bone, medial view, angle of medial process with respect to angle of vomeropalatine ramus: parallel or subparallel (0); rotated dorsally by at least 30° (1). (Ch. 78) | 1 | ? | 0 | ? | 0 | 0 | ? |
| 24. Palatine bone, pneumatic recess, form: absent or small foramen (0); large fossa with one or more foramina (1). (Ch. 84) | 1 | 1 | 0 | ? | 0 | 0 | ? |
| 25. External mandibular fenestra, lateral view, maximum anteroposterior length of fenestra relative to length of mandible: greater than 15% (0); less than 15% (1). (Ch. 87, modified from [21]: p. 38) | 0 | ? | ? | 1 | 1 | 1 | ? |
| 26. Splenial bone, medial view, mylohyoid foramen: present, completely enclosed by splenial (0); present, opens anteroventrally (1); absent (2). (Ch. 94, modified from [12]: p. 239) | 0 | ? | ? | ? | 1 | 1 | ? |
| 27. Angular bone, posterior termination: posterior or ventral to surangular foramen (0); anterior to surangular foramen (1). (Ch. 95, wording modified from [12]: p. 233) | 1 | ? | 0 | 0 | 0 | 0 | ? |
| 28. Surangular bone, dorsoventral depth over the external mandibular fenestra: less than half the depth of the mandible (0); more than half the depth of the mandible (1). (Ch. 96, [22]: p. 56, modified from [20]: p. 47). | 0 | ? | 1 | 1 | 1 | 1 | ? |
| 29. Prearticular bone, mylohyoid foramen: absent or fails to perforate anteroventral margin of prearticular (0); present, foramen height less than half the dorsoventral height of prearticular above foramen midline (1); (Ch. 99; ordered) | 0 | ? | ? | 1 | 1 | 1 | ? |
| 30. Anterior caudal vertebrae, ventral surface: shallow groove (0); distinct sunken groove (1); robust ventral ridge (2). (Ch. 127, wording modified from [9]: p. 120; [10]: p. 128) | 1 | 2 | ? | ? | ? | 0 | ? |
| 31. Scapulocoracoid, notch between scapular acromion process and coracoid bone: absent (0); present (1). (Ch. 141, [22]: p. 94; wording modified from [1]: p. 68) | 0 | ? | ? | 1 | ? | 1 | ? |
| 32. Pubis, pubic boot size relative to pubic length: less than 50% (0); 50–60% (1); greater than 60% (2). (Ch. 157, [22]: p. 80; wording modified from [20]: p. 51; ordered) | 0 | 0 | ? | 1 | ? | 1 | 1 |
| 33. Tibia, lateral condyle: confluent with cnemial crest anteriorly in proximal view (0); strongly offset from cnemial crest by incisura tibialis (1). (Ch. 166, [9]: p. 204; [10]: p. 200) | 0 | ? | ? | 1 | ? | 1 | ? |
| 34. Maxila, lateral aspect, medioventral wall of the maxillary antorbital fossa: smooth (0); a row of neurovascular foramina (1). | 0 | ? | ? | ? | 1 | 0 | ? |
| 35. Maxilla posteroventral jugal ramus of maxilla, where it articulates with jugal bone: straight (0); curved (1). | 0 | 0 | 1 | 1 | 0 | 1 | ? |
| 36. Nasal, laterodorsal margin of nasal bone pinched into a low crest, continuous from premaxilla to lacrimal bone: absent (0); present (1). | 0 | ? | 1 | 0 | 1 | 0 | ? |
| 37. Posterior portion of dorsal surface of nasal bone, cup-shaped, producing a median peak in the region of nasofrontal contact (Chure and Loewen 2020: pp. 20 and 22): absent (0); present (1). | 0 | 0 | ? | 0 | 1 | 0 | ? |
| 38. Lacrimal horns (Chure and Loewen 2020: pp. 20 and 22): absent or reduced (0); pronounced (as in Allosaurus fragilis Madsen 1976) (1); very pronounced (20% or more the lacrimal bone height, in lateral view) (2). | 0 | ? | 2 | 1 | 2 | 1 | 2 |
| 39. Jugal bone with relatively ventral margin: straight-to-slightly-curved outline in dorsal view (0); curved sinosoidal (1). | 0 | ? | 1 | 1 | 0 | 1 | ? |
| 40. Surangular bone, number of posterior surangular foramina: one (0);. two (1) (Ch. 132, Carrano et al., 2012, [2]) | 1 | ? | 1 | 0 | 0 | 1 | ? |
| 41. Cervical vertebrae, position of parapophysis on centrum: anterior (0); middle (1). (Ch. 173, Carrano et al., 2012, [2]) | 0 | 1 | 1 | 0 | 0 | 0 | ? |
| 42. Dentary, posterior end bifurcated (0); single pointed (1). NEW | 0 | ? | ? | ? | 0 | 1 | ? |
| 43. Nasal proportions (total length/maximum width measured at the anterior-most end of prefrontal): 9 or less (0); 10 or more (1). NEW | 0 | 1 | ? | 0 | 1 | 1 | ? |
| 44. Angular: dorsoposterior rim (contact with surangular bone)/mininum depth at mid-length: 4 or less (0); 5 or more (1). NEW | 0 | ? | 1 | ? | 1 | 0 | ? |
| 45. 4th and 5th cervical vertebrae, latero-anterior base of the neural spine with oblique-to-vertical subtle accessory lamina at the posteror end of the spinoprezygapophyseal lamina (best seen in lateral–dorsal views): absent (0); present (1). NEW | 0 | 0 | 1 | 0 | ? | 0 | ? |
| 46. Postorbital contact with jugal bone: straight (0); sigmoidal (1). NEW | 0 | ? | 1 | 0 | 0 | 0 | ? |
- xread
- 46 7
- Sinraptor_dongi 00000?0000010111111101110010010000000001000000
- Neovenator_salerii ???0???00?10???????????1?????2?0??0?0???1?1?0?
- Allosaurus_europaeus_ML415 11101111102??000??00?000??01??????11?2111??111
- Allosaurus_fragilis_USNM4734 ?20?0??0011??000?000????1?011?111?1001100?0?00
- Allosaurus_jimmadseni__types_ 0[12]001000012210000000100011011????10112000011?0
- Allosaurus_dino2560 0101?000011210000000?0001101101110100111011000
- Allosaurus_andres ?21??????????0???000???????????1?????2????????
- ;proc /;
- ;
8. Phylogenetic Results and Discussion
- Neovenator: Ch. 41 (Bootstrap clade support value: 40)
- Allosaurus genus: Ch. 24 and 32 (Bootstrap clade support value: 26)
- Allosaurus_fragilis USNM4734: Ch. 40 and 43
- A.jimmadseni + A. europaeus: Ch. 5, 11, 36, and 44 (Bootstrap clade support value: 21)
- Allosaurus_jimmadseni_(types): Ch. 40
- Allosaurus europaeus ML415: Ch. 3 (Ch. 1, 3, 6, 7, 8, 41, and 46 if OTU Andrés is removed) (Bootstrap clade support value: 14).
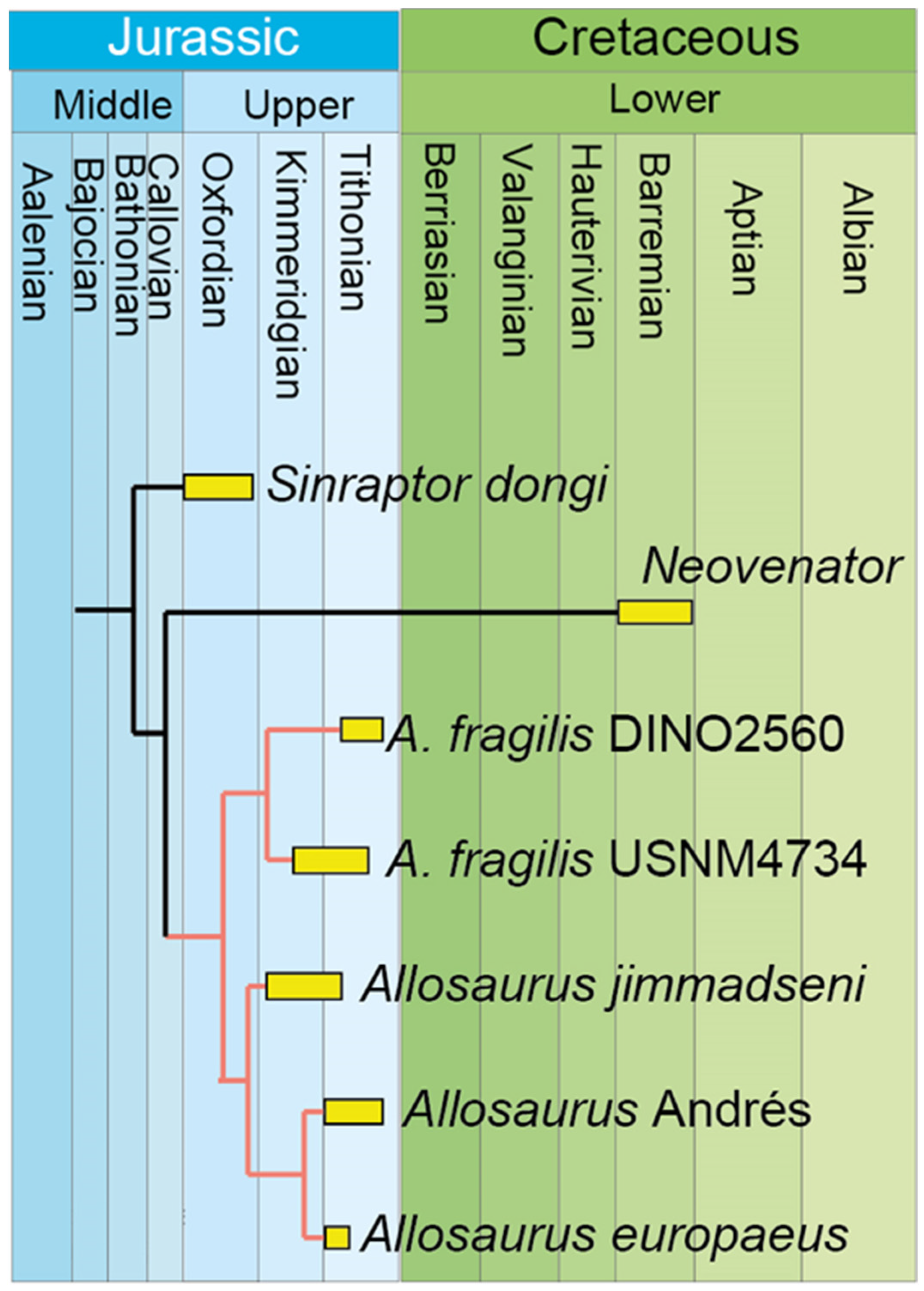
9. Cleaning the Allosaurus Wastebasket in Europe and Asia
10. Paleobiogeography and Biochronology
11. Conclusions
- The specimen of theropod dinosaur ML415, holotype of Allosaurus europaeus Mateus et al., 2006, is composed of cranial and cervical bones, with revealed additional elements, including the atlas-axis, coronoid bone, new dental remains, and rib fragments.
- A specimen-based analysis of Allosaurus confirmed a monophyletic genus, composed of three valid species: A. fragilis (USNM 4734 as neotype), A. europaeus (ML415), and A. jimmadseni.
- Allosaurus atrox, A. amplus, and A. lucasi holotypes do not have distinct autapomorphies and, therefore, are considered nomina dubia.
- A single parsimonious tree, with 58 steps, placed A. europaeus ML415 and the Andrés specimen MNHNUL/AND.001 as sister taxa, supporting that the Andrés specimen is A. europaeus.
- A. jimmadseni is closer to both Portuguese specimens than it is to A. fragilis. This confirms the validity of the species A. europaeus.
- The origin of the Allosaurus genus appears to be rooted in North America. The dispersal of Allosaurus into Europe (no later than Late Kimmeridgian) was after the phylogenetic split between A. jimmadseni and A. europaeus, and before the split between A. europaeus ML 451 and Andrés MNHNUL/AND.001.
- The previously reported Allosaurus sp. from the Cenomanian of Japan is attributed to Segnosaurus sp. The Allosaurus genus is restricted to the United States (Late Kimmeridgian to Late Tithonian), Portugal (Late Kimmeridgian to Early Tithonian), and Germany (Late Kimmeridgian).
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Rauhut, O.W.M.; Pol, D. Probable Basal Allosauroid from the Early Middle Jurassic Cañadón Asfalto Formation of Argentina Highlights Phylogenetic Uncertainty in Tetanuran Theropod Dinosaurs. Sci. Rep. 2019, 9, 18826. [Google Scholar] [CrossRef]
- Carrano, M.T.; Benson, R.B.J.; Sampson, S.D. The Phylogeny of Tetanurae (Dinosauria: Theropoda). J. Syst. Palaeontol. 2012, 10, 211–300. [Google Scholar] [CrossRef]
- Chure, D.J.; Loewen, M.A. Cranial Anatomy of Allosaurus jimmadseni, a New Species from the Lower Part of the Morrison Formation (Upper Jurassic) of Western North America. PeerJ 2020, 2020, e7803. [Google Scholar] [CrossRef] [PubMed]
- Pérez-Moreno, B.P.; Chure, D.J.; Pires, C.; Marques Da Silva, C.; Dos Santos, V.; Dantas, P.; Póvoas, L.; Cachão, M.; Sanz, J.L.; Galopim De Carvalho, A.M. On the Presence of Allosaurus fragilis (Theropoda: Carnosauria) in the Upper Jurassic of Portugal: First Evidence of an Intercontinental Dinosaur Species. J. Geol. Soc. 1999, 156, 449–452. [Google Scholar] [CrossRef]
- Malafaia, E.; Dantas, P.; Ortega, F.; Escaso, F. Nuevos Restos de Allosaurus fragilis (Theropoda: Carnosauria) Del Yacimiento de Andrés (Jurásico Superior; Centro-Oeste de Portugal). In Proceedings of the V Encuentro de Jóvenes Investigadores en PaleontologíaAt, Cuenca, Spain, 18–21 April 2007. [Google Scholar]
- Malafaia, E.; Ortega, F.; Escaso, F.; Dantas, P.; Pimentel, N.; Gasulla, J.M.; Ribeiro, B.; Barriga, F.; Sanz, J.L. Vertebrate Fauna at the Allosaurus Fossil-Site of Andrés (Upper Jurassic), Pombal, Portugal. J. Iber. Geol. 2010, 36, 193–204. [Google Scholar] [CrossRef]
- Rauhut, O.W.M.; Fechner, R. Early Development of the Facial Region in a Non-Avian Theropod Dinosaur. Proc. R. Soc. B Biol. Sci. 2005, 272, 1179–1183. [Google Scholar] [CrossRef]
- Malafaia, E.; Ortega, F.; Escaso, F.; Silva, B.; Ramalheiro, G.; Dantas, P.; Moniz, C.; Barriga, F. Análisis Preliminar de Un Nuevo Ejemplar de Allosaurus Del Grupo Lourinhã (Jurásico Superior de Torres Vedras, Portugal) A Preliminary Account of a New Allosaurus Individual from the Lourinhã Group (Upper Jurassic of Torres Vedras, Portugal). Actas De Las IV Jorn. Int. Sobre Paleontol. De Dinosaur. Y Su Entorno Salas De Los Infantes 2008, 243–251. [Google Scholar]
- Malafaia, E.; Mocho, P.; Escaso, F.; Ortega, F. A New Carcharodontosaurian Theropod from the Lusitanian Basin: Evidence of Allosauroid Sympatry in the European Late Jurassic. J. Vertebr. Paléontol. 2020, 40, e1768106. [Google Scholar] [CrossRef]
- Mateus, O.; Walen, A.; Antunes, M.T. The Large Theropod Fauna of the Lourinhã Formation (Portugal) and Its Similarity to That of the Morrison Formation, With a Description of a New Species of Allosaurus. Paleontol. Geol. Up. Jurass. Morrison Form. Bull. 2006, 36, 123. [Google Scholar]
- Evers, S.W.; Foth, C.; Rauhut, O.W.M. Notes on the Cheek Region of the Late Jurassic Theropod Dinosaur Allosaurus. PeerJ 2020, 2020, e8493. [Google Scholar] [CrossRef] [PubMed]
- Madsen, J.H. Allosaurus fragilis: A Revised Osteology Utah Department of Natural Resources, 1993rd ed.; Utah Geological Survey: Salt Lake City, UT, USA, 1976; Volume 109.
- Escaso, F.; Ortega, F.; Dantas, P.; Malafaia, E.; Pimentel, N.L.; Pereda-Suberbiola, X.; Sanz, J.L.; Kullberg, J.C.; Kullberg, M.C.; Barriga, F. New Evidence of Shared Dinosaur across Upper Jurassic Proto-North Atlantic: Stegosaurus from Portugal. Naturwissenschaften 2006, 94, 367–374. [Google Scholar] [CrossRef]
- Costa, F.; Mateus, O. Dacentrurine Stegosaurs (Dinosauria): A New Specimen of Miragaia longicollum from the Late Jurassic of Portugal Resolves Taxonomical Validity and Shows the Occurrence of the Clade in North America. PLoS ONE 2019, 14, e0224263. [Google Scholar] [CrossRef]
- Mateus, O.; Dinis, J.; Cunha, P.P. The Lourinhã Formation: The Upper Jurassic to Lower Most Cretaceous of the Lusitanian Basin, Portugal – Landscapes Where Dinosaurs Walked. Ciências Terra-Earth Sci. J. 2017, 19, 75–97. [Google Scholar] [CrossRef]
- Gowland, S.; Taylor, A.M.; Martinius, A.W. Integrated Sedimentology and Ichnology of Late Jurassic Fluvial Point-Bars—Facies Architecture and Colonization Styles (Lourinhã Formation, Lusitanian Basin, Western Portugal). Sedimentology 2018, 65, 400–430. [Google Scholar] [CrossRef]
- Taylor, A.M.; Gowland, S.; Leary, S.; Keogh, K.J.; Martinius, A.W. Stratigraphical Correlation of the Late Jurassic Lourinhã Formation in the Consolação Sub-Basin (Lusitanian Basin), Portugal. Geol. J. 2014, 49, 143–162. [Google Scholar] [CrossRef]
- Mateus, O.; Milàn, J. Ichnological Evidence for Giant Ornithopod Dinosaurs in the Upper Jurassic Lourinhã Formation, Portugal. Oryctos 2008, 8, 47–52. [Google Scholar]
- Milán, J.; Mateus, O. Fra Strandbred Til Museum På Syv Dage–Historien Om et Gigantisk Dinosaur Fodspor. Varv–Bl. Med Ældste Nyheder 2003, 8–14. [Google Scholar] [CrossRef]
- Mateus, O.; Milàn, J.; Romano, M.; Whyte, M.A. New Finds of Stegosaur Tracks from the Upper Jurassic Lourinhã Formation, Portugal. Acta Palaeontol. Pol. 2011, 56, 651–658. [Google Scholar] [CrossRef]
- Guillaume, A.R.D.; Costa, F.; Mateus, O. Stegosaur Tracks from the Upper Jurassic of Portugal: New Occurrences and Perspectives. Ciências Terra/Earth Sci. J. 2022, 20, 37–60. [Google Scholar] [CrossRef]
- Lapparent, A.F.; Zbyszewski, G. Les Dinosauriens Du Portugal. Mémoires Des Serv. Géologiques Du Portugal. 1957, 2, 1–63. [Google Scholar]
- Bonaparte, J.; Mateus, O. A New Diplodocid, Dinheirosaurus Iourinhanensis Gen. et Sp. Nov., from the Late Jurassic Beds of Portugal. Rev. Del Mus. Argent. De Cienc. Nat. “Bernardino Rivadavia” E Inst. Nac. De Investig. De Las Cienc. Nat. 1999, 5, 13. [Google Scholar]
- Tschopp, E.; Mateus, O.; Benson, R.B.J. A Specimen-Level Phylogenetic Analysis and Taxonomic Revision of Diplodocidae (Dinosauria, Sauropoda). PeerJ 2015, 2015, e857. [Google Scholar] [CrossRef] [PubMed]
- Mateus, O. Lourinhanosaurus antunesi, a new upper Jurassic allosauroid (Dinosauria: Theropoda) from Lourinhã, Portugal. Memórias Acad. Ciências Lisb. 1998, 37, 111–124. [Google Scholar]
- Mateus, O.; Maidment, S.C.R.; Christiansen, N.A. A New Long-Necked ‘Sauropod-Mimic’ Stegosaur and the Evolution of the Plated Dinosaurs. Proc. R. Soc. B Biol. Sci. 2009, 276, 1815. [Google Scholar] [CrossRef]
- Ramalho, M.M.R. Contribution à L` Étude Micropaleontologique et Stratigraphique Du Jurassique Supérieur et Du Crétacé Inférieur Des Environs de Lisbonne (Portugal). Memórias Serviços Geológicos Port. 1971, 19, 1–212. [Google Scholar]
- Mateus, O.; Telles Antunes, M. Draconyx loureiroi, a New Camptosauridae (Dinosauria, Ornithopoda) from the Late Jurassic of Lourinhã, Portugal. Ann. Paléontologie 2001, 87, 61–73. [Google Scholar] [CrossRef]
- Rotatori, F.M.; Moreno-Azanza, M.; Mateus, O. Reappraisal and New Material of the Holotype of Draconyx loureiroi (Ornithischia: Iguanodontia) Provide Insights on the Tempo and Modo of Evolution of Thumb-Spiked Dinosaurs. Zoöl. J. Linn. Soc. 2022, 195, 125–156. [Google Scholar] [CrossRef]
- Eddy, D.R.; Clarke, J.A. New Information on the Cranial Anatomy of Acrocanthosaurus atokensis and Its Implications for the Phylogeny of Allosauroidea (Dinosauria: Theropoda). PLoS ONE 2011, 6, e17932. [Google Scholar] [CrossRef] [PubMed]
- Goloboff, P.A.; Morales, M.E. TNT Version 1.6, with a Graphical Interface for MacOS and Linux, Including New Routines in Parallel. Cladistics 2023, 39, 144–153. [Google Scholar] [CrossRef]
- Hendrickx, C.; Araújo, R.; Mateus, O. The Non-Avian Theropod Quadrate I: Standardized Terminology with an Overview of the Anatomy and Function. PeerJ 2015, 2015, e1245. [Google Scholar] [CrossRef] [PubMed]
- Carpenter, K. Variation in a Population of Theropoda (Dinosauria): Allosaurus from the Cleveland-Lloyd Quarry (Upper Jurassic), Utah, USA. Paléontol. Res. 2010, 14, 250–259. [Google Scholar] [CrossRef]
- Chure, D.J. A New Species of Allosaurus from the Morrison Formation of Dinosaur National Monument (UT-CO) and a Revision of the Theropod Family Allosauridae; Columbia University: New York, NY, USA, 2000. [Google Scholar]
- Gilmore, C.W. Osteology of the Carnivorous Dinosauria in the United States National Museum, with Special Reference to the Genera Antrodemus (Allosaurus) and Ceratosaurus. Bull. U.S. Natl. Mus. 1920, I–XI, 1–159. [Google Scholar] [CrossRef]
- Currie, P.J.; Zhao, X.-J. A New Carnosaur (Dinosauria, Theropoda) from the Jurassic of Xinjiang, People’s Republic of China. Can. J. Earth Sci. 2011, 30, 2037–2081. [Google Scholar] [CrossRef]
- McClelland, B.K. Anatomy and Kinesis of the Allosaurus Skull. Ph.D. Thesis, Texas Tech University, Lubbock, TX, USA, 1990. [Google Scholar]
- Paul, G.S.; Carpenter, K. Case 3506 Allosaurus Marsh, 1877 (Dinosauria, Theropoda): Proposed Conservation of Usage by Designation of a Neotype for Its Type Species Allosaurus fragilis Marsh, 1877. Bull. Zoöl. Nomencl. 2010, 67, 53–56. [Google Scholar] [CrossRef]
- Paul, G.S. Predatory Dinosaurs of the World; A New York Academy of Sciences Book; Touchstone: New York, NY, USA, 1988; ISBN 0-671-61946-2. [Google Scholar]
- Dalman, S.G. Osteology of a Large Allosauroid Theropod from the Upper Jurassic (Tithonian) Morrison Formation of Colorado, USA. Vol. Jurassica 2014, XII, 69–106. [Google Scholar]
- Carrano, M.T.; Loewen, M.A.; Evers, S.W. Comment (Case 3506)—Conservation of Allosaurus Marsh, 1877 (Dinosauria, Theropoda): Additional Data in Support of the Proposed Neotype for Its Type Species Allosaurus fragilis Marsh, 1877. Bull. Zool. Nomencl. 2018, 75, 59–64. [Google Scholar] [CrossRef]
- Smith, D.K. Patterns of Size-Related Variation within Allosaurus. J. Vertebr. Paléontol. 2010, 19, 402–403. [Google Scholar] [CrossRef]
- Rauhut, O.W.M. Post-Cranial Remains of ‘Coelurosaurs’ (Dinosauria, Theropoda) from the Late Jurassic of Tanzania. Geol. Mag. 2005, 142, 97–107. [Google Scholar] [CrossRef]
- Rauhut, O.W.M. Theropod Dinosaurs from the Late Jurassic of Tendaguru (Tanzania). Spec. Pap. Palaeontol. 2011, 86, 195–239. [Google Scholar] [CrossRef]
- Galton, P.M.; Carpenter, K.; Dalman, S.G. The Holotype Pes of the Morrison Dinosaur Camptosaurus amplus Marsh, 1879 (Upper Jurassic, Western USA)—Is It Camptosaurus, Sauropoda or Allosaurus? Neues Jahrb. Für Geol. Und Paläontologie Abh. 2015, 275, 317–335. [Google Scholar] [CrossRef] [PubMed]
- Chure, D.J. A Reassessment of the Gigantic Theropod Saurophaganax maximus from the Morrison Formation (Upper Jurassic) of Oklahoma, USA. Sixth Symp. Mesoz. Terr. Ecosyst. Biota 1995, 6, 103–106. [Google Scholar]
- Hone, D.W.E.; Farke, A.A.; Wedel, M.J. Ontogeny and the Fossil Record: What, If Anything, Is an Adult Dinosaur? Biol. Lett. 2016, 12, 20150947. [Google Scholar] [CrossRef]
- Madsen, J.H., Jr.; Welles, S.P. Ceratosaurus (Dinosauria, Theropoda) a Revised Osteology; Stringfellow, J., Ed.; Utah Geological Survey: Salt Lake City, UT, USA, 2000; Volume 2, ISBN 1557913803.
- Hendrickx, C.; Mateus, O. Torvosaurus gurneyi n. Sp., the Largest Terrestrial Predator from Europe, and a Proposed Terminology of the Maxilla Anatomy in Nonavian Theropods. PLoS ONE 2014, 9, e88905. [Google Scholar] [CrossRef]
- McIntosh, J.S. Annotated Catalogue of the Dinosaurs (Reptilia, Archosauria) in the Collections of Carnegie Museum of Natural History. Bull. Carnegie Mus. Nat. Hist. 1981, 18, 1–67. [Google Scholar] [CrossRef]
- Mojon, P.-O. Dinosauriens Éocretacés Des Faciès Purbeckiens (Berriasien Inférieur) Du Jura Méridional (S.-E. de La France). Arch. Sci. Compte Rendu Seances Soc. 2001, 54, 1–5. [Google Scholar] [CrossRef]
- Blows, W.T. Reptiles On The Rocks; Isle of Wight Museum Publication: Newport, UK, 1978; pp. 1–60. [Google Scholar]
- Gerke, O.; Wings, O. Multivariate and Cladistic Analyses of Isolated Teeth Reveal Sympatry of Theropod Dinosaurs in the Late Jurassic of Northern Germany. PLoS ONE 2016, 11, e0158334. [Google Scholar] [CrossRef] [PubMed]
- Diedrich, C. Upper Jurassic Tidal Flat Megatracksites of Germany—Coastal Dinosaur Migration Highways between European Islands, and a Review of the Dinosaur Footprints. Paleobiodivers Paleoenviron 2011, 91, 129–155. [Google Scholar] [CrossRef]
- Tamura, M.; Okazaki, Y.; Ikegami, N. Occurrence of Carnosaurian and Herbivorous Dinosaurs from Upper Formation of Mifune Group, Japan. Mem. Fac. Educ. Kumamoto Univ. 1991, 40, 31–45. [Google Scholar]
- Zanno, L.E. A Taxonomic and Phylogenetic Re-Evaluation of Therizinosauria (Dinosauria: Maniraptora). J. Syst. Palaeontol. 2010, 8, 503–543. [Google Scholar] [CrossRef]
- Ikegami, N. A Therizinosaurid Dinosaur from the Upper Cretaceous Mifune Group in Kyushu, Japan. J. Vertebr. Paleontol. 2005, 25, 73A. [Google Scholar]
- Gradstein, F.; Gale, A.; Kopaevich, L.; Waskowska, A.; Grigelis, A.; Glinskikh, L.; Görög, Á. The Planktonic Foraminifera of the Jurassic. Part II: Stratigraphy, Palaeoecology and Palaeobiogeography. Swiss J. Palaeontol. 2017, 136, 259–271. [Google Scholar] [CrossRef]
- Wilhem, C. Maps of the Callovian and Tithonian Paleogeography of the Caribbean, Atlantic, and Tethyan Realms: Facies and Environments. Geol. Soc. Am. Digit. Map Chart Ser. 2014, 17, 1–9. [Google Scholar] [CrossRef]
- Francischini, H.; Sales, M.A.F.; Dentzien–Dias, P.; Schultz, C.L. The Presence of Ankylosaur Tracks in the Guará Formation (Brazil) and Remarks on the Spatial and Temporal Distribution of Late Jurassic Dinosaurs. Ichnos 2018, 25, 177–191. [Google Scholar] [CrossRef]
- Mateus, O. Late Jurassic Dinosaurs from the Morrison Formation (USA), the Lourinhã and Alcobaca Formations (Portugal), and the Tendaguru Beds (Tanzania): A Comparison. New Mex. Mus. Nat. Hist. Sci. Bull. 2006, 36, 223–231. [Google Scholar]
- Turner, C.E.; Peterson, F. Biostratigraphy of Dinosaurs in the Upper Jurassic Morrison Formation of the Western Interior, U.S.A. In Vertebrate Paleontology in Utah; Gillette, D.D., Ed.; Utah Geological Survey: Salt Lake City, UT, USA, 1999; Volume 99, pp. 77–102. [Google Scholar]
- Evanoff, E.; Carpenter, K. History, Sedimentology, and Taphonomy of Felch Quarry 1 and Associated Sandbodies, Morrison Formation, Garden Park, Colorado. Mod. Geol. 1998, 22, 145–169. [Google Scholar]
- Carpenter, K. Vertebrate Biostratigraphy of the Morrison Formation near Canon City, Colorado. In Modern Geology; Overseas Publishers Association: Geneva, Switzerland, 1998; Volume 23, p. 418. [Google Scholar]
- Kirkland, J.; DeBlieux, D.D.; Hunt-Foster, R.K.; Foster, J.R.; Trujillo, K.; Finzel, E. The Morrison Formation and Its Bounding Strata on the Western Side of the Blanding Basin, San Juan County, Utah. Geol. Intermt. West 2020, 7, 137–195. [Google Scholar] [CrossRef]
- Gates, T.A. The Late Jurassic Cleveland-Lloyd Dinosaur Quarry as a Drought-Induced Assemblage. Palaios 2005, 20, 363–375. [Google Scholar] [CrossRef]
- Maidment, S.C.R. Diversity through Time and Space in the Upper Jurassic Morrison Formation, Western U.S.A. J Vertebr Paleontol 2023, 43, 5. [Google Scholar] [CrossRef]
- Mocho, P.; Royo-Torres, R.; Escaso, F.; Malafaia, E.; de Miguel Chaves, C.; Narváez, I.; Pérez-García, A.; Pimentel, N.; Silva, B.C.; Ortega, F. Upper Jurassic Sauropod Record in the Lusitanian Basin (Portugal): Geographical and Lithostratigraphical Distribution. Palaeontol. Electron. 2017, 20, 1–50. [Google Scholar] [CrossRef] [PubMed]
- Vavilov, N.I. Origin and Geography of Cultivated Crops; Cambridge University Press: London, UK, 1992; ISBN 0521111595. [Google Scholar]
- Ward, L.F. The Mesozoic Flora of Portugal Compared with That of the United States. New Ser. 1895, 1, 337–346. [Google Scholar] [CrossRef] [PubMed]
- Galton, P.M. The Ornithopod Dinosaur Dryosaurus and a Laurasia–Gondwanaland Connection in the Upper Jurassic. Nature 1977, 268, 230–232. [Google Scholar] [CrossRef]
- Brikiatis, L. Late Mesozoic North Atlantic Land Bridges. Earth-Sci. Rev. 2016, 159, 47–57. [Google Scholar] [CrossRef]
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Burigo, A.; Mateus, O. Allosaurus europaeus (Theropoda: Allosauroidea) Revisited and Taxonomy of the Genus. Diversity 2025, 17, 29. https://doi.org/10.3390/d17010029
Burigo A, Mateus O. Allosaurus europaeus (Theropoda: Allosauroidea) Revisited and Taxonomy of the Genus. Diversity. 2025; 17(1):29. https://doi.org/10.3390/d17010029
Chicago/Turabian StyleBurigo, André, and Octávio Mateus. 2025. "Allosaurus europaeus (Theropoda: Allosauroidea) Revisited and Taxonomy of the Genus" Diversity 17, no. 1: 29. https://doi.org/10.3390/d17010029
APA StyleBurigo, A., & Mateus, O. (2025). Allosaurus europaeus (Theropoda: Allosauroidea) Revisited and Taxonomy of the Genus. Diversity, 17(1), 29. https://doi.org/10.3390/d17010029