
Carabid Beetles under the Influence of Megafires in Pine and Secondary Forests of Central European Russia



Abstract
1. Introduction
2. Materials and Methods
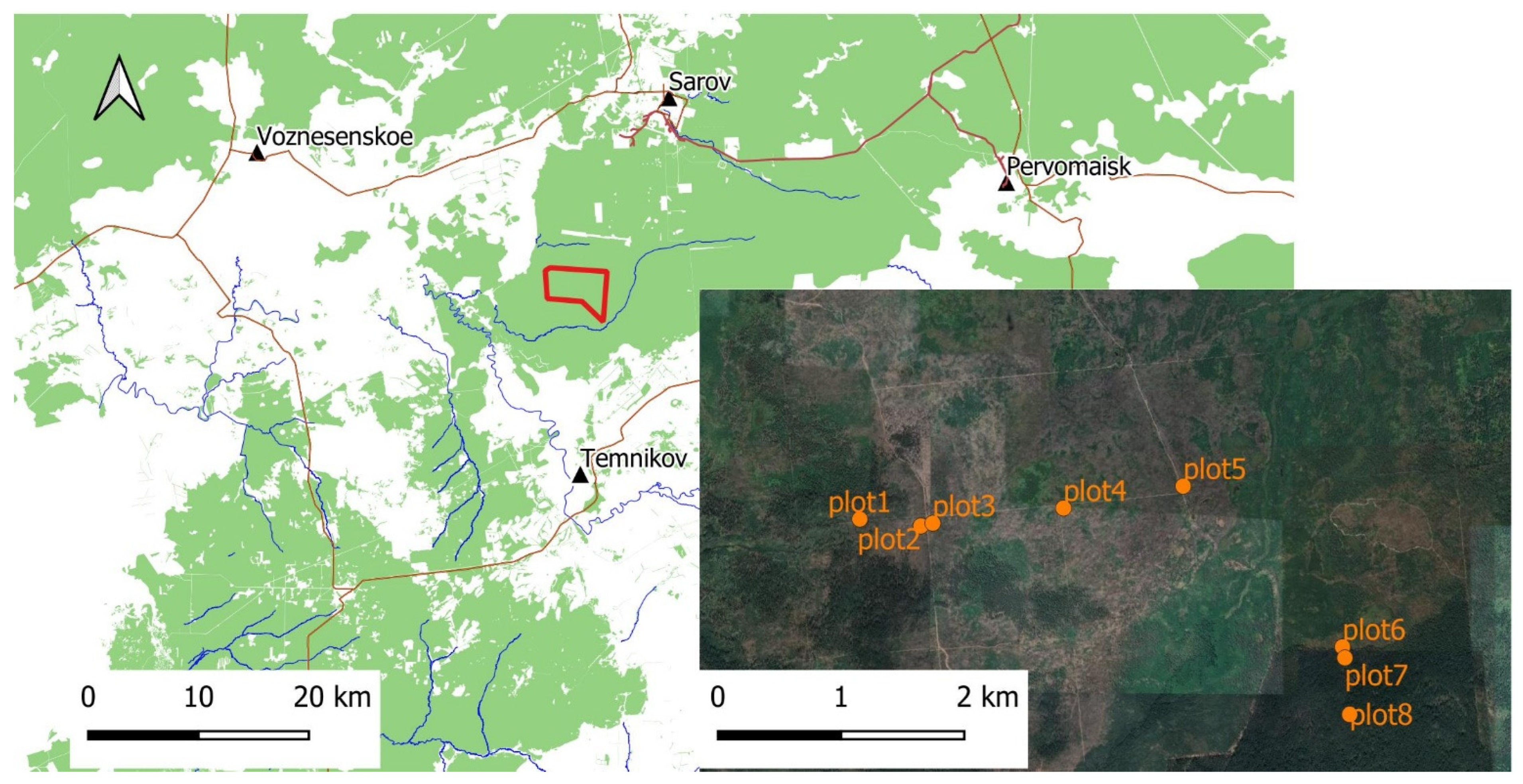
2.1. Study Area
2.2. Sampling Procedures
2.3. Data Analysis
3. Results
3.1. Species Composition and Structure
3.2. Changes in the First Year after the Megafire
3.3. Changes in the Second Year after the Megafire
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Conflicts of Interest
References
- Granström, A. Fire management for biodiversity in the European boreal forest. Scand. J. For. Res. 2001, 16 (Suppl. 3), 62–69. [Google Scholar] [CrossRef]
- Driscoll, D.A.; Lindenmayer, D.B.; Bennett, A.F.; Bode, M.; Bradstock, R.A.; Cary, G.J.; York, A. Fire management for biodiversity conservation: Key research questions and our capacity to answer them. Biol. Conserv. 2010, 143, 1928–1939. [Google Scholar] [CrossRef]
- He, T.; Lamont, B.B.; Pausas, J.G. Fire as a key driver of Earth’s biodiversity. Biol. Rev. 2019, 94, 1983–2010. [Google Scholar] [CrossRef] [PubMed]
- Kharitonova, A.O.; Kharitonova, T.I. The effect of landscape pattern on the 2010 wildfire spread in the Mordovia State Nature Reserve, Russia. Nat. Conserv. Res. 2021, 6, 29–41. [Google Scholar] [CrossRef]
- Tiberio, F.C.S.; Xavier, R.O.; Dodonov, P.; Silva Matos, D.M. Fire has short-term negative effects on a super-dominant native fern, Pteridium arachnoideum (Dennstaedtiaceae), in a Brazilian savanna. Nat. Conserv. Res. 2022, 7, 15–25. [Google Scholar] [CrossRef]
- Atutova, Z.V. Post-fire restoration of pine forests in the Badary area, Tunkinskiy National Park, Russia. Nat. Conserv. Res. 2023, 8, 22–32. [Google Scholar] [CrossRef]
- Viljur, M.L.; Abella, S.R.; Adámek, M.; Alencar, J.B.R.; Barber, N.A.; Beudert, B.; Burkle, L.A.; Cagnolo, L.; Campos, B.R.; Chao, A.; et al. The effect of natural disturbances on forest biodiversity: An ecological synthesis. Biol. Rev. 2022, 97, 1930–1947. [Google Scholar] [CrossRef]
- Mahmoodzadeh, A.; Eisapoor, S.S.; Mirghiasy, S.A. Biodiversity in the third millennium. Sci. Rep. Life Sci. 2022, 3, 1–14. [Google Scholar] [CrossRef]
- Certini, G.; Moya, D.; Lucas-Borja, M.E.; Mastrolonardo, G. The impact of fire on soil-dwelling biota: A review. For. Ecol. Manag. 2021, 488, 118989. [Google Scholar] [CrossRef]
- El Khayati, M.; Chergui, B.; Barranco, P.; Fahd, S.; Ruiz, J.L.; Taheri, A.; Santos, X. Assessing the Response of Different Soil Arthropod Communities to Fire: A Case Study from Northwestern Africa. Fire 2023, 6, 206. [Google Scholar] [CrossRef]
- Vilkova, V.V.; Kazeev, K.S.; Privizentseva, D.A.; Nizhelsky, M.S.; Kolesnikov, S.I. Activity in post-pyrogenic soils in the Utrish State Nature Reserve (Russia) in the early succession stages. Nat. Conserv. Res. 2023, 8, 10–23. [Google Scholar] [CrossRef]
- Vilkova, V.V.; Kazeev, K.S.; Nizhelskiy, M.S.; Kolesnikov, S.I.; Kozun, Y.S. Changes in soil properties of xerophytic forests in Southern Russia after anthropogenic impact. Nat. Conserv. Res. 2024, 9, 61–72. [Google Scholar] [CrossRef]
- Bouderbala, I.; Labadie, G.; Béland, J.M.; Tremblay, J.A.; Boulanger, Y.; Hébert, C.; Desrosiers, P.; Allard, A.; Fortin, D. Long-term effect of forest harvesting on boreal species assemblages under climate change. PLoS Clim. 2023, 2, e0000179. [Google Scholar] [CrossRef]
- Mansoor, S.; Farooq, I.; Kachroo, M.M.; Mahmoud, A.E.D.; Fawzy, M.; Popescu, S.M.; Ahmad, P. Elevation in wildfire frequencies with respect to the climate change. J. Environ. Manag. 2022, 301, 113769. [Google Scholar] [CrossRef] [PubMed]
- Shashkov, M.P.; Bobrovsky, M.V.; Shanin, V.N.; Khanina, L.G.; Grabarnik, P.Y.; Stamenov, M.N.; Ivanova, N.V. Data on 30-year stand dynamics in an old-growth broad-leaved forest in the Kaluzhskie Zaseki State Nature Reserve, Russia. Nat. Conserv. Res. 2022, 7 (Suppl. 1), 24–37. [Google Scholar] [CrossRef]
- Gandhi, K.J.; Spence, J.R.; Langor, D.W.; Morgantini, L.E. Fire residuals as habitat reserves for epigaeic beetles (Coleoptera: Carabidae and Staphylinidae). Biol. Conserv. 2001, 102, 131–141. [Google Scholar] [CrossRef]
- Saint-Germain, M.; Larrivée, M.; Drapeau, P.; Fahrig, L.; Buddle, C.M. Short-term response of ground beetles (Coleoptera: Carabidae) to fire and logging in a spruce-dominated boreal landscape. For. Ecol. Manag. 2005, 212, 118–126. [Google Scholar] [CrossRef]
- Gongalsky, K.; Midtgaard, F.; Overgaard, H. Effects of prescribed forest burning on carabid beetles (Coleoptera: Carabidae): A case study in south-eastern Norway. Entomol. Fenn. 2006, 17, 325–333. [Google Scholar] [CrossRef]
- Mason, S.C., Jr.; Shirey, V.; Waite, E.S.; Gallagher, M.R.; Skowronski, N.S. Exploring Prescribed Fire Severity Effects on Ground Beetle (Coleoptera: Carabidae) Taxonomic and Functional Community Composition. Fire 2023, 6, 366. [Google Scholar] [CrossRef]
- Zumr, V.; Remeš, J.; Nakládal, O. Short-Term Response of Ground Beetles (Coleoptera: Carabidae) to Fire in Formerly Managed Coniferous Forest in Central Europe. Fire 2024, 7, 76. [Google Scholar] [CrossRef]
- Holliday, N.J. Species responses of carabid beetles (Coleoptera: Carabidae) during post-fire regeneration of boreal forest. Can. Entomol. 1991, 123, 1369–1389. [Google Scholar] [CrossRef]
- Fernández, M.F.; Costas, J.S. Recolonization of a burnt pine forest (Pinus pinaster) by Carabidae (Coleoptera). Eur. J. Soil Biol. 2004, 40, 47–53. [Google Scholar] [CrossRef]
- Gongalsky, K.B.; Wikars, L.-O.; Persson, T. Ground beetle (Coleoptera: Carabidae) responses to a forest wildfire in northern Europe. Russ. Entomol. J. 2008, 17, 273–282. [Google Scholar]
- Mason, S.C., Jr.; Shirey, V.; Ponisio, L.C.; Gelhaus, J.K. Responses from bees, butterflies, and ground beetles to different fire and site characteristics: A global meta-analysis. Biol. Conserv. 2021, 261, 109265. [Google Scholar] [CrossRef]
- Belluz, V.; Langor, D.W.; Niemelä, J.K.; He, F.; Spence, J.R. Long-term responses of ground beetles (Coleoptera: Carabidae) to clear-cutting and wildfire in lodgepole pine stands of western Alberta, Canada. Can. Entomol. 2022, 154, e41. [Google Scholar] [CrossRef]
- Gongalsky, K.B. Perfugia as a mechanism for the recovery of soil fauna after ecosystem disturbances. Russ. J. Ecosyst. Ecol. 2017, 2. [Google Scholar] [CrossRef]
- Paquin, P. Carabid beetle (Coleoptera: Carabidae) diversity in the black spruce succession of eastern Canada. Biol. Conserv. 2008, 141, 261–275. [Google Scholar] [CrossRef]
- Lassau, S.A.; Hochuli, D.F.; Cassis, G.; Reid, C.A. Effects of habitat complexity on forest beetle diversity: Do functional groups respond consistently? Divers. Distrib. 2005, 11, 73–82. [Google Scholar] [CrossRef]
- Toïgo, M.; Paillet, Y.; Noblecourt, T.; Soldati, F.; Gosselin, F.; Dauffy-Richard, E. Does forest management abandonment matter more than habitat characteristics for ground beetles? Biol. Conserv. 2013, 157, 215–224. [Google Scholar] [CrossRef]
- Mordkovich, V.G.; Berezina, O.G. Effect of fire on the pedobiont communities of a birch-aspen grove in the southern forest-steppe of West Siberia. Euroasian Entomol. J. 2009, 8, 279–283. [Google Scholar]
- Samu, F.; Kádár, F.; Ónodi, G.; Kertész, M.; Szirányi, A.; Szita, É.; Fetykó, K.; Neidert, D.; Botos, E.; Altbäcker, V. Differential ecological responses of two generalist arthropod groups, spiders and carabid beetles (Araneae, Carabidae), to the effects of wildfire. Community Ecol. 2010, 11, 129–139. [Google Scholar] [CrossRef]
- Matalin, A.V.; Trushitsina, O.S.; Makarov, K.V. Influence of different types of wildfire on the community structure of ground beetles (Coleoptera, Carabidae) in pine forests of the Meshchera Lowlands. In Proceedings of the 18th European Carabidologist Meeting, Rennes, France, 25–29 September 2017; p. 87. [Google Scholar]
- Ruchin, A.B.; Alekseev, S.K.; Khapugin, A.A. Post-fire fauna of carabid beetles (Coleoptera, Carabidae) in forests of the Mordovia State Nature Reserve (Russia). Nat. Conserv. Res. 2019, 4 (Suppl. 1), 11–20. [Google Scholar] [CrossRef]
- Ruchin, A.B. The selected insect families and their seasonal dynamics in the Mordovia State nature reserve in the burned areas of 2021. J. Wildl. Biodivers. 2024, 8, 17–38. [Google Scholar] [CrossRef]
- Khapugin, A.A.; Vargot, E.V.; Chugunov, G.G. Vegetation recovery in fire-damaged forests: A case study at the southern boundary of the taiga zone. For. Stud. 2016, 64, 39–50. [Google Scholar] [CrossRef]
- Koch, K. Die Käfer Mitteleuropas: Ökologie. Bd. 1; Goecke & Evers: Keltern, Germany, 1989; 440p. [Google Scholar]
- Lindroth, C.H. Ground Beetles (Carabidae) of Fennoscandia: A Zoogeographic Study: Part 1. Specific Knowledge regarding the Species; Intercept Ltd.: Andover, UK, 1992; pp. xxviii + 630. [Google Scholar]
- Ribera, I.; Foster, G.N.; Downie, I.S.; McCracken, D.I.; Abernethy, V.J. A comparative study of the morphology and life traits of Scottish ground beetles (Coleoptera, Carabidae). Ann. Zool. Fenn. 1999, 36, 21–37. [Google Scholar]
- Aleksanov, V.V.; Alekseev, S.K. Inventory of the Ground Beetles (Coleoptera, Carabidae) of Kaluga Urban Okrug; Ministry of Natural Resources and Ecology of the Kaluga Oblast: Izhevsk, Russia, 2019; 278p.
- Sharova, I.K. Life Forms of Ground Beetles (Coleoptera, Carabidae); Nauka: Moscow, Russia, 1981; 24p. [Google Scholar]
- Hengeveld, R. Polyphagy, oligophagy and food specialization in ground beetles (Coleoptera: Carabidae). Neth. J. Zool. 1980, 30, 564–584. [Google Scholar] [CrossRef]
- Talarico, F.; Giglio, A.; Pizzolotto, R.; Brandmayr, P. A synthesis of feeding habits and reproduction rhythm in Italian seed-feeding ground beetles (Coleoptera: Carabidae). EJE 2016, 113, 325–336. [Google Scholar] [CrossRef]
- Matalin, A.V. The life cycles of grounds beetles (Coleoptera, Carabidae) in West Palaearctic. Ph.D. Thesis, Moscow State Pedagogical University, Moscow, Russia, 2011; 549p. [Google Scholar]
- Hammer, Ø.; Harper, D.A.; Ryan, P.D. PAST: Paleontological Statistics Software Package for Education and Data Analysis. Palaeontol. Electron. 2001, 4, 9. [Google Scholar]
- R Core Team. R: A Language and Environment for Statistical Computing; R Foundation for Statistical Computing: Vienna, Austria, 2022; Available online: https://www.R-project.org/ (accessed on 20 June 2024).
- Oksanen, J. Vegan: Ecological Diversity. Available online: http://cran.r-project.org/web/packages/vegan/vignettes/diversity-vegan.pdf (accessed on 1 October 2022).
- Fedorenko, D.N. The fauna of ground-beetles (Coleoptera, Carabidae) of Moscow Province. In Insects of the Moscow Region: Problems of Cadastre and Protection; Nauka: Moscow, Russia, 1988; pp. 20–46. [Google Scholar]
- Skłodowski, J.; Garbalińska, P. Ground beetle (Coleoptera, Carabidae) assemblages inhabiting Scots pine stands of Puszcza Piska Forest: Six-year responses to a tornado impact. In ZooKeys, Carabid Beetles as Bioindicators: Biogeographical, Ecological and Environmental Studies; Kotze, D.J., Assmann, T., Noordijk, J., Turin, H., Vermeulen, R., Eds.; Pensoft Publishers: Moscow, Russia, 2011; Volume 100, pp. 371–392. [Google Scholar] [CrossRef]
- Alekseev, S.K.; Ruchin, A.B. Fauna and abundance of ground beetle (Coleoptera, Carabidae) in pine forests. Entomol. Appl. Sci. Lett. 2020, 7, 1–9. [Google Scholar]
- Cook, W.M.; Holt, R.D. Fire frequency and mosaic burning effects on a tallgrass prairie ground beetle assemblage. Biodivers. Conserv. 2006, 15, 2301–2323. [Google Scholar] [CrossRef]
- Jacobs, J.M.; Bergeron, J.A.C.; Work, T.T.; Spence, J.R. Low intensity surface fire instigates movement by adults of Calosoma frigidum (Coleoptera, Carabidae). ZooKeys 2011, 147, 641–649. [Google Scholar] [CrossRef] [PubMed]
- Ariza, G.M.; Jácome, J.; Kotze, D.J. Carabid beetles of tropical dry forests display traits that cope with a harsh environment. Int. J. Trop. Insect Sci. 2021, 41, 3011–3021. [Google Scholar] [CrossRef]
- Ruchin, A.B.; Egorov, L.V.; MacGowan, I.; Makarkin, V.N.; Antropov, A.V.; Gornostaev, N.G.; Khapugin, A.A.; Dvořák, L.; Esin, M.N. Post-fire insect fauna explored by crown fermental traps in forests of the European Russia. Sci. Rep. 2021, 11, 21334. [Google Scholar] [CrossRef] [PubMed]
- Moretti, M.; Duelli, P.; Obrist, M.K. Biodiversity and resilience of arthropod communities after fire disturbance in temperate forests. Oecologia 2006, 149, 312–327. [Google Scholar] [CrossRef] [PubMed]
- Gonganlsky, K.B.; Wikars, L.-O.; Persson, T. Dynamics of pyrophilous carabids in a burned pine forest in Central Sweden. Baltic, J. Coleopterol. 2003, 3, 107–111. [Google Scholar]
- Pradella, C.; Wermelinger, B.; Obrist, M.K.; Duelli, P.; Moretti, M. On the occurrence of five pyrophilous beetle species in the Swiss Central Alps (Leuk, Canton Valais). Mitteilungen Schweiz. Entomol. Ges. 2010, 83, 187–197. [Google Scholar]
- Bell, A.J.; Calladine, K.S.; Wardle, D.A.; Phillips, I.D. Rapid colonization of the post-burn environment improves egg survival in pyrophilic ground beetles. Ecosphere 2022, 13, e4213. [Google Scholar] [CrossRef]
- Bell, A.J. Like moths to a flame: A review of what we know about pyrophilic insects. For. Ecol. Manag. 2023, 528, 120629. [Google Scholar] [CrossRef]
- Süda, I.; Voolma, K.; Õunap, H. Short-term monitoring of fire-adapted Coleoptera in burnt pine forest of northern Estonia. Acta Biol. Univ. Daugavp. 2009, 9, 43–48. [Google Scholar]
- Wikars, L.O. Clear-cutting before burning prevents establishment of the fire-adapted Agonum quadripunctatum (Coleoptera: Carabidae). Ann. Zool. Fenn. 1995, 32, 375–384. [Google Scholar]
- Saint-Germain, M.; Drapeau, P.; Buddle, C.M. Persistence of pyrophilous insects in fire-driven boreal forests: Population dynamics in burned and unburned habitats. Divers. Distrib. 2008, 14, 713–720. [Google Scholar] [CrossRef]
- Niemelä, J.; Haila, Y.; Halme, E.; Lahti, T.; Pajunen, T.; Punttila, P. The distribution of carabid beetles in fragments of old coniferous taiga and adjacent managed forest. Ann. Zool. Fenn. 1988, 25, 107–119. [Google Scholar]
- Wallin, H. Spatial and temporal distribution of some abundance carabid beetles (Col., Carabidae) in cereal fields and adjacent habitats. Pedobiologia 1985, 28, 19–34. [Google Scholar]
- Wallin, H.; Ekbom, B.S. Movements of carabid beetles (Coleoptera: Carabidae) inhabiting cereal fields: A field tracing study. Oecologia 1988, 77, 39–43. [Google Scholar] [CrossRef]
- Zhang, J.; Drummond, F.A.; Liebman, M.; Hartke, A. Phenology and dispersal of Harpalus rufipes De Geer (Coleoptera: Carabidae) in agroecosystems in Maine. J. Agric. Entomol. 1997, 14, 171–186. [Google Scholar]
- Matalin, A.V. Evolution of biennial life cycles in ground beetles (Coleoptera, Carabidae) of the Western Palaearctic. In Proceedings of the XIII European Carabidologist Meeting, Blagoevgrad, Bulgaria, 20–24 August 2008; Back to the roots and back to the future. pp. 259–284. [Google Scholar]
- Davies, C.E.; Moss, D.; Hill, M.O. EUNIS habitat classification revised 2004. Report to: European Environment Agency-European Topic Centre on Nature Protection and Biodiversity. Estuar. Coast. Shelf Sci. 2004, 62, 127–143. [Google Scholar]
- Skłodowski, J. Multi-phase recovery of carabid assemblages during 19 years of secondary succession in forest stands disturbed by windstorm without salvage logging in northern Poland. Sci. Total Environ. 2023, 862, 160763. [Google Scholar] [CrossRef] [PubMed]

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Plots | Description |
|---|---|
| 1 | This plot was burned in 2010. However, it was not affected by the fire in 2021. It is located 500 m from the edge of the megafire. It has a significant amount of deadwood, dry pines and birches. Young birch trees grow well. Shrubs are represented mainly by raspberries. The herbaceous layer is sparse due to a large amount of deadwood. The litter is small from birch fall. There are areas on the ground surface that are devoid of vegetation. |
| 2 | The 2021 fire boundary. This plot is similar to the previous plot in vegetation, deadwood, and dead trees. However, the plot is situated at a distance of 10 m from the edge of the 2021 fire. |
| 3 | The 2021 fire boundary. It is located in a very close proximity to plot 2. It was burned in 2010 and 2021. The plot is situated at a distance of 10 m away from the edge of the fire and deep within the burned area. In 2021, the area was completely burned out with vegetation and plant remains. There are a few dry bushes left. Revegetation began in late 2022. Perennial grasses and annual cereals have appeared. Shrubs and trees that were not completely destroyed have sprouted new shrubs and young shoots. In 2023, the projective cover of the herbaceous layer continued to increase. Nevertheless, there are numerous areas that remain devoid of vegetation. |
| 4 | This plot is similar to plot 3. However, the plot is situated at a distance of 1000 m away from the edge of the fire and deep within the burned area. The rate of overgrowth of herbaceous vegetation is slower than in plot 3. The plot is characterized by extensive sandy areas devoid of vegetation. |
| 5 | This plot is similar to plot 3. However, the plot is situated at a distance of 2000 m away from the edge of the fire and deep within the burned area. The rate of overgrowth of herbaceous vegetation is even slower than in plot 3. The plot is characterised by extensive sandy areas devoid of vegetation. |
| 6 | The 2021 fire boundary. It was burned in 2010 and 2021. However, the plot is bordered by a living forest area that has not been affected by fire for at least 200 years. The plot is situated at a distance of 10 m away from the edge of the fire and deep within the burned area (20 m from plot 7). The area burned in 2021. Deadwood, birch, shrubs, and herbaceous layer partially burned. Deadwood, birch wood, shrubs, and herbaceous layer partially burned. At least half of the deadwood and dense dry birch undergrowth remained. |
| 7 | This plot is control. A section of the forest that has not been affected by fire for at least 200 years. The plot is situated at a distance of 10 m away from the edge of the 2010 and 2021 fires. Old-growth mixed forest of pine, birch with admixture of linden, rowan, euonymus, bird cherry (in the second layer). The litter is well defined, thick and the herbaceous layer is sparse. |
| 8 | This plot is similar to plot 7. However, it is situated 500 m away from the edge of the 2010 and 2021 fires. Old-growth mixed forest of pine, birch with admixture of linden, rowan, euonymus, bird cherry (in the second layer). The litter is well defined, thick and the herbaceous layer is sparse. |
| Species | Specimens | Number of Plots | ||
|---|---|---|---|---|
| 2022 | 2023 | 2022 | 2023 | |
| Agonum gracile Sturm, 1824 | 2 | 0 | 1 | 0 |
| Agonum gracilipes (Duftschmid, 1812) | 1 | 1 | 1 | 1 |
| Amara aenea (De Geer, 1774) * | 7 | 4 | 5 | 3 |
| Amara apricaria (Paykull, 1790) * | 0 | 5 | 0 | 2 |
| Amara aulica (Panzer, 1796) * | 2 | 1 | 1 | 1 |
| Amara bifrons (Gyllenhal, 1810) ** | 28 | 96 | 7 | 8 |
| Amara brunnea (Gyllenhal, 1810) ** | 381 | 234 | 7 | 7 |
| Amara communis (Panzer, 1797) * | 37 | 280 | 6 | 7 |
| Amara consularis (Duftschmid, 1812) * | 12 | 130 | 4 | 7 |
| Amara convexior Stephens, 1828 * | 0 | 17 | 0 | 4 |
| Amara eurynota (Panzer, 1796) * | 1 | 7 | 1 | 4 |
| Amara famelica C.C.A. Zimmermann, 1832 * | 4 | 2 | 1 | 2 |
| Amara familiaris (Duftschmid, 1812) * | 1 | 6 | 1 | 3 |
| Amara fulva (O.F. Müller, 1776) * | 1 | 7 | 1 | 3 |
| Amara ingenua (Duftschmid, 1812) * | 8 | 10 | 1 | 4 |
| Amara lunicollis Schiødte, 1837 * | 0 | 10 | 0 | 4 |
| Amara majuscula (Chaudoir, 1850) * | 2 | 1 | 2 | 1 |
| Amara municipalis (Duftschmid, 1812) * | 1 | 0 | 1 | 0 |
| Amara nitida Sturm, 1825 ** | 0 | 11 | 0 | 3 |
| Amara ovata (Fabricius, 1792) * | 10 | 1 | 4 | 1 |
| Amara plebeja (Gyllenhal, 1810) * | 1 | 2 | 1 | 1 |
| Amara praetermissa (C.R. Sahlberg, 1827) ** | 0 | 12 | 0 | 2 |
| Amara similata (Gyllenhal, 1810) * | 4 | 4 | 3 | 3 |
| Amara tibialis (Paykull, 1798) * | 23 | 29 | 5 | 4 |
| Anisodactylus nemorivagus (Duftschmid, 1812) * | 4 | 11 | 2 | 5 |
| Anisodactylus signatus (Panzer, 1796) * | 2 | 0 | 2 | 0 |
| Asaphidion flavipes (Linnaeus, 1761) | 0 | 1 | 0 | 1 |
| Badister bullatus (Schrank, 1798) | 1 | 1 | 1 | 1 |
| Badister sodalis (Duftschmid, 1812) | 0 | 1 | 0 | 1 |
| Bembidion biguttatum (Fabricius, 1779) | 2 | 1 | 1 | 1 |
| Bembidion lampros (Herbst, 1784) | 5 | 22 | 2 | 2 |
| Bembidion properans (Stephens, 1828) | 0 | 1 | 0 | 1 |
| Bembidion quadrimaculatum (Linnaeus, 1761) | 0 | 3 | 0 | 2 |
| Bradycellus caucasicus (Chaudoir, 1846) * | 3 | 7 | 2 | 2 |
| Broscus cephalotes (Linnaeus, 1758) | 2 | 0 | 2 | 0 |
| Calathus ambiguus (Paykull, 1790) | 0 | 1 | 0 | 1 |
| Calathus erratus (C.R. Sahlberg, 1827) | 13 | 59 | 5 | 7 |
| Calathus fuscipes (Goeze, 1777) | 0 | 3 | 0 | 3 |
| Calathus melanocephalus (Linnaeus, 1758) | 4 | 8 | 4 | 5 |
| Calathus micropterus (Duftschmid, 1812) | 65 | 140 | 4 | 4 |
| Calosoma inquisitor (Linnaeus, 1758) | 4 | 0 | 3 | 0 |
| Calosoma investigator (Illiger, 1798) | 0 | 4 | 0 | 2 |
| Carabus cancellatus Illiger, 1798 | 4 | 2 | 3 | 2 |
| Carabus convexus Fabricius, 1775 | 7 | 8 | 2 | 4 |
| Carabus arcensis baschkiricus Breuning, 1932 | 572 | 370 | 5 | 7 |
| Carabus clathratus Linnaeus, 1761 | 0 | 1 | 0 | 1 |
| Carabus coriaceus Linnaeus, 1758 | 4 | 3 | 1 | 3 |
| Carabus estreicheri Fischer von Waldheim, 1820 | 0 | 1 | 0 | 1 |
| Carabus glabratus Paykull, 1790 | 330 | 351 | 5 | 5 |
| Carabus granulatus Linnaeus, 1758 | 3 | 9 | 2 | 2 |
| Carabus hortensis Linnaeus, 1758 | 3 | 4 | 2 | 2 |
| Carabus schoenherri Fischer von Waldheim, 1820 | 1 | 1 | 1 | 1 |
| Cicindela campestris Linnaeus, 1758 | 10 | 12 | 2 | 2 |
| Cicindela hybrida Linnaeus, 1758 | 1 | 0 | 1 | 0 |
| Cicindela sylvatica Linnaeus, 1758 | 6 | 11 | 2 | 1 |
| Cychrus caraboides (Linnaeus, 1758) | 2 | 1 | 1 | 1 |
| Cymindis angularis Gyllenhal, 1810 | 1 | 3 | 1 | 3 |
| Dolichus halensis (Schaller, 1783) | 1 | 1 | 1 | 1 |
| Harpalus affinis (Schrank, 1781) * | 14 | 29 | 4 | 5 |
| Harpalus anxius (Duftschmid, 1812) * | 1 | 9 | 1 | 3 |
| Harpalus calceatus (Duftschmid, 1812) * | 0 | 4 | 0 | 2 |
| Harpalus distinguendus (Duftschmid, 1812) * | 3 | 42 | 3 | 5 |
| Harpalus flavescens (Piller & Mitterpacher, 1783) * | 1 | 1 | 1 | 1 |
| Harpalus griseus (Panzer, 1796) * | 7 | 26 | 3 | 6 |
| Harpalus hirtipes (Panzer, 1796) * | 0 | 3 | 0 | 3 |
| Harpalus laevipes Zetterstedt, 1828 * | 45 | 66 | 5 | 5 |
| Harpalus latus (Linnaeus, 1758) * | 57 | 136 | 7 | 8 |
| Harpalus luteicornis (Duftschmid, 1812) * | 0 | 5 | 0 | 3 |
| Harpalus progrediens Schauberger, 1922 * | 7 | 13 | 2 | 5 |
| Harpalus pumilus Sturm, 1818 * | 8 | 14 | 2 | 4 |
| Harpalus rubripes (Duftschmid, 1812) * | 43 | 51 | 7 | 7 |
| Harpalus rufipes (De Geer, 1774) * | 774 | 536 | 8 | 7 |
| Harpalus signaticornis (Duftschmid, 1812) * | 1 | 12 | 1 | 3 |
| Harpalus smaragdinus (Duftschmid, 1812) * | 31 | 148 | 6 | 6 |
| Harpalus tardus (Panzer, 1796) * | 60 | 197 | 6 | 8 |
| Harpalus xanthopus winkleri Schauberger, 1923 * | 11 | 15 | 4 | 4 |
| Lebia chlorocephala (J.J. Hoffmann, 1803) | 1 | 0 | 1 | 0 |
| Lebia cruxminor (Linnaeus, 1758) | 0 | 1 | 0 | 1 |
| Leistus ferrugineus (Linnaeus, 1758) | 0 | 1 | 0 | 1 |
| Licinus depressus (Paykull, 1790) | 2 | 3 | 2 | 2 |
| Masoreus wetterhallii (Gyllenhal, 1813) | 2 | 10 | 2 | 3 |
| Microlestes minutulus (Goeze, 1777) | 0 | 1 | 0 | 1 |
| Notiophilus aquaticus (Linnaeus, 1758) | 5 | 0 | 2 | 0 |
| Notiophilus germinyi Fauvel, 1863 | 0 | 36 | 0 | 6 |
| Notiophilus palustris (Duftschmid, 1812) | 6 | 33 | 2 | 7 |
| Olisthopus rotundatus (Paykull, 1790) | 0 | 1 | 0 | 1 |
| Ophonus rufibarbis (Fabricius, 1792) ** | 1 | 1 | 1 | 1 |
| Ophonus stictus Stephens, 1828 ** | 1 | 0 | 1 | 0 |
| Oxypselaphus obscurus (Herbst, 1784) | 2 | 3 | 1 | 1 |
| Panagaeus bipustulatus (Fabricius, 1775) | 2 | 6 | 1 | 2 |
| Paradromius linearis (G.-A. Olivier, 1795) | 1 | 0 | 1 | 0 |
| Patrobus atrorufus (Strøm, 1768) | 1 | 0 | 1 | 0 |
| Poecilus cupreus (Linnaeus, 1758) | 6 | 2 | 2 | 2 |
| Poecilus lepidus (Leske, 1785) | 214 | 339 | 7 | 6 |
| Poecilus versicolor (Sturm, 1824) | 118 | 78 | 7 | 7 |
| Pterostichus melanarius (Illiger, 1798) | 14 | 11 | 3 | 2 |
| Pterostichus minor (Gyllenhal, 1827) | 0 | 3 | 0 | 2 |
| Pterostichus niger (Schaller, 1783) | 169 | 250 | 4 | 5 |
| Pterostichus nigrita (Paykull, 1790) | 28 | 19 | 3 | 4 |
| Pterostichus oblongopunctatus (Fabricius, 1787) | 158 | 137 | 4 | 3 |
| Pterostichus quadrifoveolatus Letzner, 1852 | 600 | 486 | 6 | 5 |
| Pterostichus rhaeticus Heer, 1837 | 3 | 0 | 2 | 0 |
| Pterostichus strenuus (Panzer, 1796) | 13 | 15 | 4 | 4 |
| Pterostichus uralensis (Motschulsky, 1850) | 1 | 2 | 1 | 2 |
| Sericoda quadripunctata (De Geer, 1774) | 3 | 4 | 1 | 1 |
| Syntomus foveatus (Geoffroy, 1785) | 0 | 2 | 0 | 1 |
| Synuchus vivalis (Illiger, 1798) | 2 | 10 | 1 | 3 |
| Tachyta nana (Gyllenhal, 1810) | 0 | 3 | 0 | 2 |
| Total specimens | 3992 | 4675 | - | - |
| Species number | 83 | 96 | - | - |
| Variable | Plots | |||||||
|---|---|---|---|---|---|---|---|---|
| 1 | 2 | 3 | 4 | 5 | 6 | 7 | 8 | |
| Species number | 37 40 | 29 42 | 18 28 | 19 42 | 13 28 | 44 54 | 42 38 | 28 37 |
| Total abundance, ex. per 100 trap days | 180.9 203.5 | 170.4 334.8 | 54.3 172.2 | 109.3 127.3 | 31.5 55.1 | 746.9 504.0 | 511.7 337.4 | 659.3 626.8 |
| Shannon index | 2.447 2.763 | 2.452 2.574 | 1.749 2.386 | 1.448 2.602 | 1.754 2.468 | 1.737 2.338 | 2.523 2.802 | 1.701 2.289 |
| Simpson_1-D | 0.836 0.900 | 0.859 0.861 | 0.643 0.847 | 0.533 0.826 | 0.727 0.857 | 0.691 0.758 | 0.882 0.916 | 0.732 0.857 |
| Berger-Parker | 0.348 0.216 | 0.290 0.306 | 0.591 0.314 | 0.678 0.393 | 0.490 0.266 | 0.474 0.470 | 0.210 0.157 | 0.420 0.219 |
| Abundance of some groups and species, ex. per 100 trap days | ||||||||
| Macropterous | 101.9 132.3 | 116 222.2 | 49.4 150.0 | 101.2 118.2 | 24.1 53.0 | 668.5 431.3 | 263.6 138.9 | 146.3 219.2 |
| Brachypterous | 14.8 11.1 | 3.7 4.5 | 1.2 1.0 | 3.7 3.0 | 1.9 1.5 | 435.8 296.5 | 234.0 172.2 | 488.3 365.7 |
| Omnivorous | 63.6 106.1 | 89.5 195.5 | 45.7 139.9 | 92.6 107.1 | 19.1 43.4 | 258.6 131.8 | 150.6 114.6 | 30.9 150.5 |
| Herbivorous | 25.3 14.1 | 22.8 20.7 | 1.2 3.0 | 2.5 2.5 | 0.0 0.5 | 4.3 10.1 | 67.9 13.1 | 112.3 60.6 |
| A. brunnea | 25.3 9.1 | 22.8 20.7 | 1.2 3 | 2.5 1.5 | 0 0 | 4.3 10.1 | 66.7 13.1 | 112.3 60.6 |
| A. communis | 7.4 0.5 | 3.1 4 | 1.2 2 | 1.2 1 | 0 0 | 0 11.1 | 1.2 25.8 | 8.6 97 |
| C. arcensis | 1.2 0.5 | 0 1.5 | 0.6 0.5 | 0 0.5 | 0 0 | 1.2 1 | 72.8 49 | 277.2 133.8 |
| C. glabratus | 6.2 2.5 | 1.9 0.5 | 0 0 | 0 0 | 0 0 | 3.1 7.6 | 32.1 29.3 | 160.5 137.4 |
| H. rufipes | 10.5 18.2 | 30.2 50 | 32.1 54 | 74.1 50 | 15.4 13.6 | 206.2 52 | 107.4 32.8 | 1.9 0 |
| P. lepidus | 63 43.9 | 49.4 102.5 | 4.3 20.7 | 7.4 1.5 | 5.6 0 | 1.9 2 | 0.6 0 | 0 0.5 |
| P. niger | 0 5.1 | 0.6 0 | 0 0 | 0 0 | 0 0.5 | 58 31.8 | 13.6 21.2 | 32.1 67.7 |
| P. oblongopunctatus | 0.6 0 | 0 0 | 0 0 | 0 0 | 0 0 | 4.3 3 | 81.5 53 | 11.1 13.1 |
| P. quadrifoveolatus | 0 1.5 | 0 0 | 0.6 0 | 3.7 0 | 0.6 0.5 | 354.3 236.9 | 10.5 5.6 | 0.6 1 |
| S. quadripunctata | 0 0 | 0 0 | 0 0 | 0 0 | 0 0 | 1.9 2.0 | 0 0 | 0 0 |
| Variable | Variant | Df | SS | MS | F | Pr (>F) |
|---|---|---|---|---|---|---|
| Total abundance | Burned_2010 | 1 | 205,304 | 205,304 | 2.829 | 0.153 |
| Burned_2021 | 1 | 4780 | 4780 | 0.066 | 0.808 | |
| Residuals | 5 | 362,868 | 72,574 | |||
| Species number | Burned_2010 | 1 | 104.2 | 104.2 | 0.733 | 0.431 |
| Burned_2021 | 1 | 120.3 | 120.3 | 0.846 | 0.400 | |
| Residuals | 5 | 711.0 | 142.2 | |||
| Macropterous | Burned_2010 | 1 | 1183 | 1183 | 0.020 | 0.892 |
| Burned_2021 | 1 | 13,832 | 13,832 | 0.239 | 0.646 | |
| Residuals | 5 | 289,411 | 57,882 | |||
| Brachypterous | Burned_2010 | 1 | 450,700 | 450,700 | 5.012 | 0.0753 |
| Burned_2021 | 1 | 631 | 631 | 0.007 | 0.9365 | |
| Residuals | 5 | 449,631 | 89,926 | |||
| Omnivorous | Burned_2010 | 1 | 25 | 25 | 0.003 | 0.958 |
| Burned_2021 | 1 | 1006 | 1006 | 0.119 | 0.744 | |
| Residuals | 5 | 42,154 | 8431 | |||
| Herbivorous | Burned_2010 | 1 | 9784 | 9784 | 48.874 | 0.0009 |
| Burned_2021 | 1 | 649 | 649 | 3.244 | 0.1316 | |
| Residuals | 5 | 1001 | 200 | |||
| A. brunnea | Burned_2010 | 1 | 9635 | 9635 | 45.596 | 0.0011 |
| Burned_2021 | 1 | 649 | 649 | 3.073 | 0.1400 | |
| Residuals | 5 | 1057 | 211 | |||
| H. rufipes | Burned_2010 | 1 | 69 | 69 | 0.012 | 0.916 |
| Burned_2021 | 1 | 5055 | 5055 | 0.897 | 0.387 | |
| Residuals | 5 | 28,169 | 5634 | |||
| H. smaragdinus | Burned_2010 | 1 | 15.26 | 15.257 | 2.800 | 0.155 |
| Burned_2021 | 1 | 12.7 | 12.701 | 2.331 | 0.187 | |
| Residuals | 5 | 27.24 | 5.449 | |||
| C. arcensis | Burned_2010 | 1 | 45,668 | 45,668 | 10.94 | 0.0213 |
| Burned_2021 | 1 | 0 | 0 | 0.00 | 0.9979 | |
| Residuals | 5 | 20,875 | 4175 | |||
| C. glabratus | Burned_2010 | 1 | 13,380 | 13,380 | 8.100 | 0.036 |
| Burned_2021 | 1 | 14 | 14 | 0.008 | 0.930 | |
| Residuals | 5 | 8259 | 1652 | |||
| P. lepidus | Burned_2010 | 1 | 700 | 700 | 32.21 | 0.0024 |
| Burned_2021 | 1 | 3521 | 3521 | 161.98 | 0.0001 | |
| Residuals | 5 | 109 | 22 | |||
| P. niger | Burned_2010 | 1 | 256.1 | 256.1 | 0.475 | 0.521 |
| Burned_2021 | 1 | 268.8 | 268.8 | 0.498 | 0.512 | |
| Residuals | 5 | 2696.8 | 539.4 | |||
| P. oblongopunctatus | Burned_2010 | 1 | 3101.7 | 3101.7 | 6.228 | 0.0548 |
| Burned_2021 | 1 | 0.8 | 0.8 | 0.002 | 0.9697 | |
| Residuals | 5 | 2490.2 | 498.0 | |||
| P. quadrifoveolatus | Burned_2010 | 1 | 4426 | 4426 | 0.237 | 0.647 |
| Burned_2021 | 1 | 10,756 | 10,756 | 0.576 | 0.482 | |
| Residuals | 5 | 93,340 | 18,668 | |||
| S. quadripunctata | Burned_2010 | 1 | 0.1429 | 0.1429 | 0.278 | 0.621 |
| Burned_2021 | 1 | 0.2858 | 0.2858 | 0.556 | 0.490 | |
| Residuals | 5 | 2.2 | 0.5144 |
| Year | Factor | Df | SS | R2 | F | Pr (>F) |
|---|---|---|---|---|---|---|
| 2022 | Burned_2010 | 1 | 0.665 | 0.308 | 3.325 | 0.003 |
| Burned_2021 | 1 | 0.494 | 0.229 | 2.468 | 0.028 | |
| Residual | 5 | 1.001 | 0.463 | |||
| Total | 7 | 2.160 | 1.000 | |||
| 2023 | Burned_2010 | 1 | 0.678 | 0.359 | 3.736 | 0.005 |
| Burned_2021 | 1 | 0.301 | 0.160 | 1.658 | 0.129 | |
| Residual | 5 | 0.907 | 0.481 | |||
| Total | 7 | 1.886 | 1.000 |
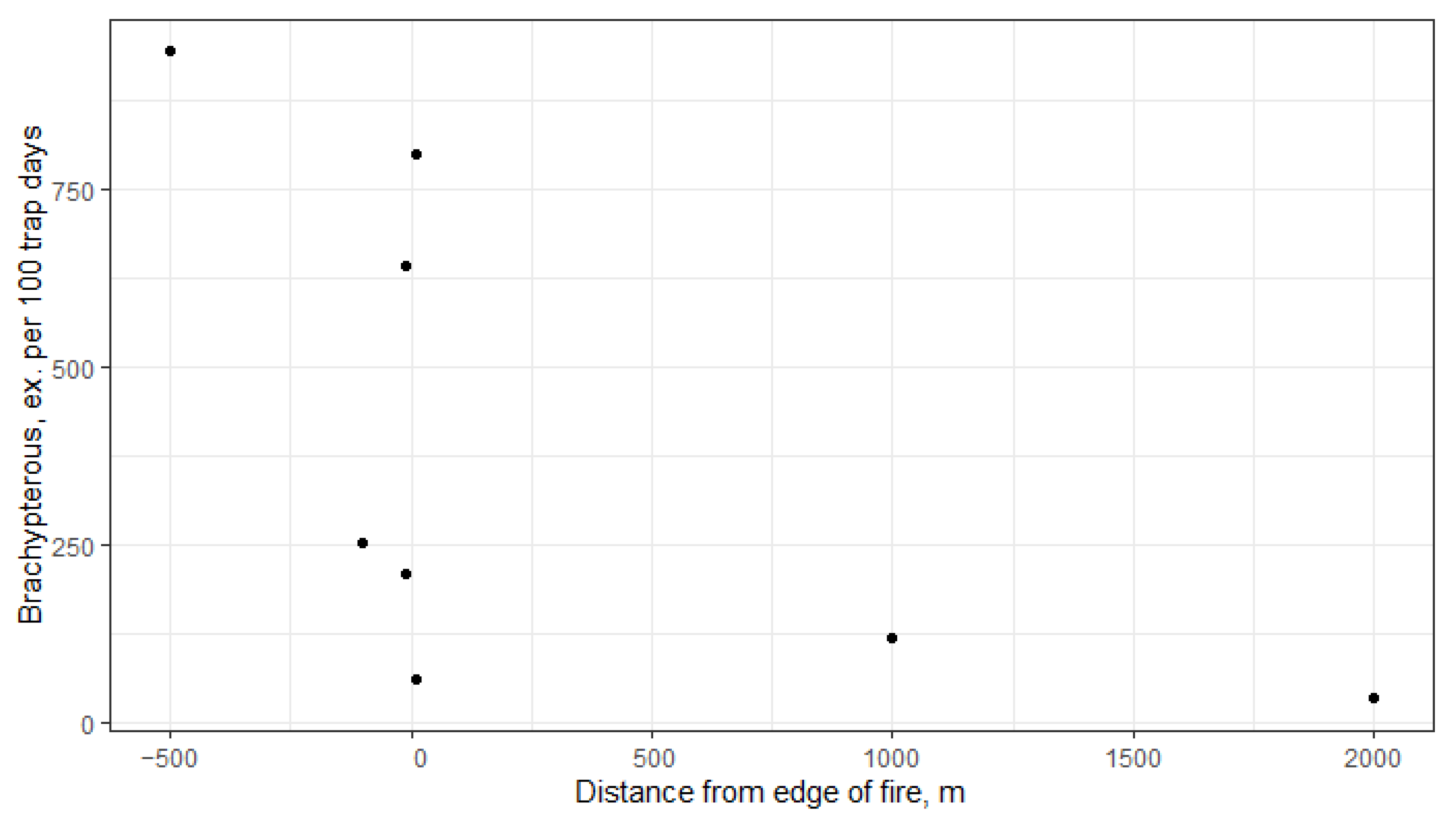
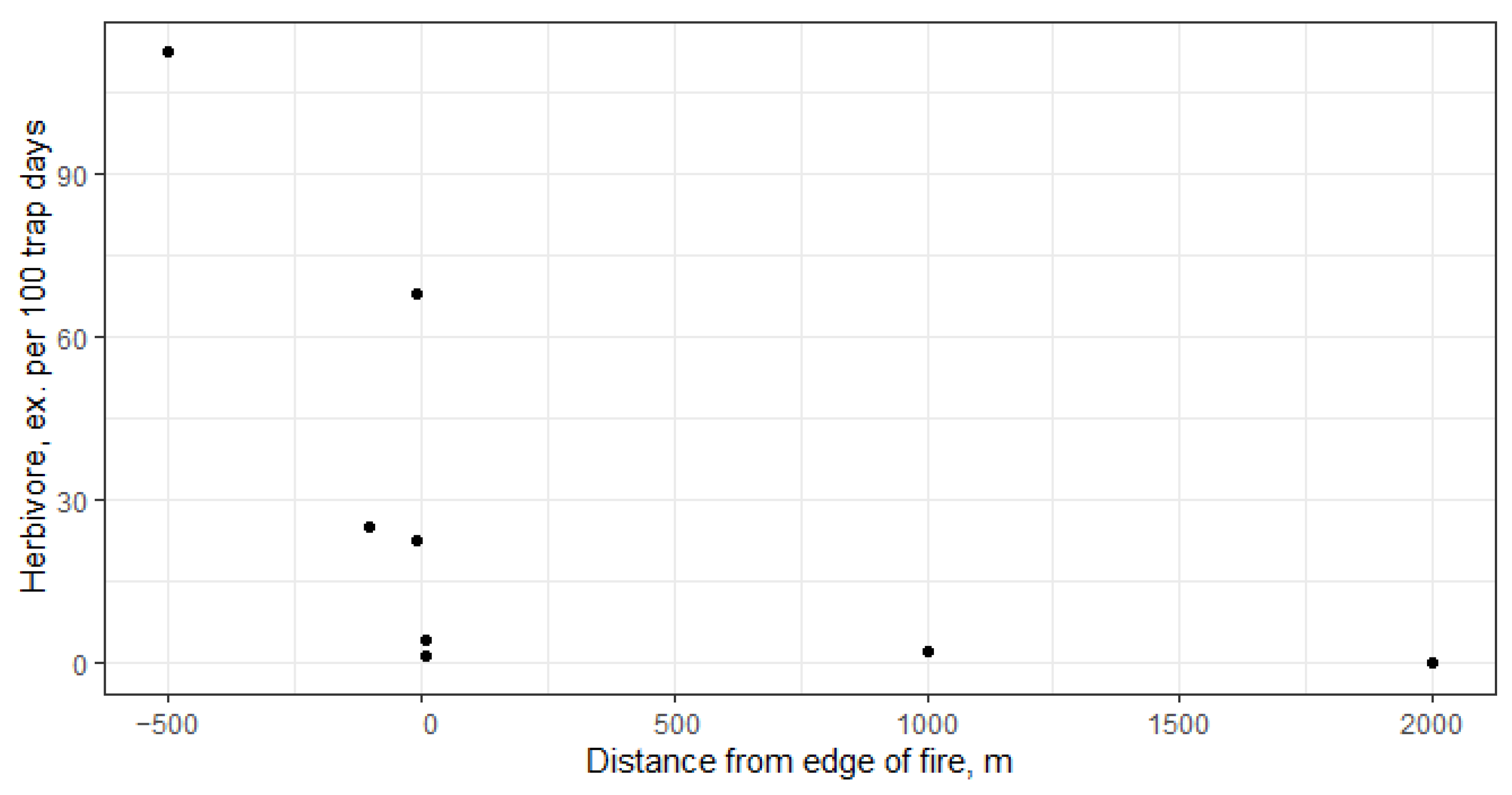
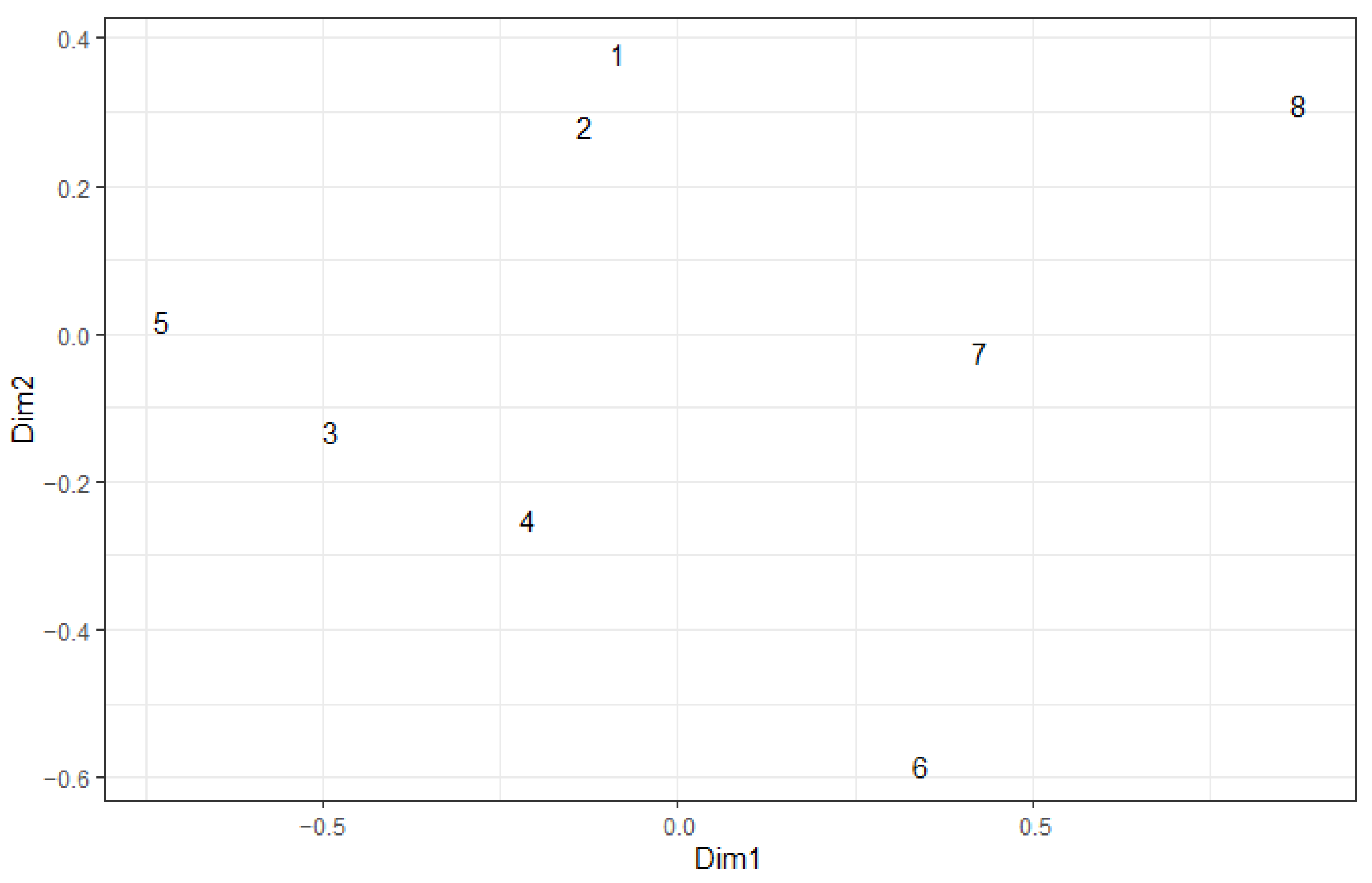
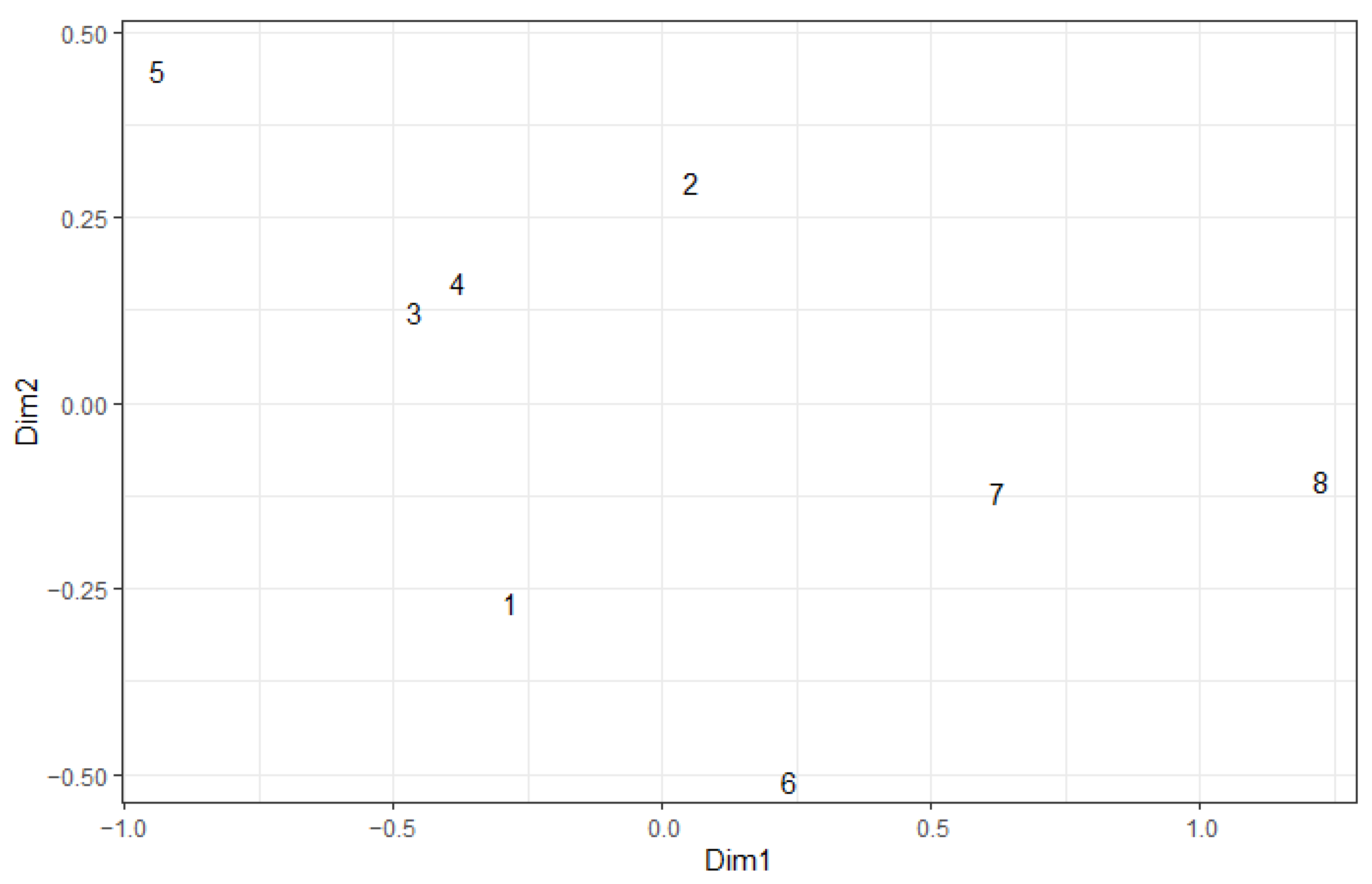
| Year | Variable | Dim1 | Dim2 | r2 | Pr (>r) |
|---|---|---|---|---|---|
| 2022 | Brachypterous | 0.964 | −0.265 | 0.935 | 0.001 |
| Herbivore | 0.753 | 0.658 | 0.877 | 0.003 | |
| Distance from edge of fire | −0.932 | −0.363 | 0.540 | 0.136 | |
| 2023 | Brachypterous | 0.993 | −0.121 | 0.887 | 0.007 |
| Herbivore | 0.861 | 0.509 | 0.747 | 0.013 | |
| Distance from edge of fire | −0.589 | 0.808 | 0.641 | 0.078 |
| Df | SS | MS | F | Pr (>F) | |
|---|---|---|---|---|---|
| Year | 1 | 0.002 | 0.002 | 0.287 | 0.6005 |
| Residuals | 14 | 0.104 | 0.007 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Aleksanov, V.V.; Alekseev, S.K.; Ruchin, A.B.; Esin, M.N. Carabid Beetles under the Influence of Megafires in Pine and Secondary Forests of Central European Russia. Diversity 2024, 16, 370. https://doi.org/10.3390/d16070370
Aleksanov VV, Alekseev SK, Ruchin AB, Esin MN. Carabid Beetles under the Influence of Megafires in Pine and Secondary Forests of Central European Russia. Diversity. 2024; 16(7):370. https://doi.org/10.3390/d16070370
Chicago/Turabian StyleAleksanov, Victor V., Sergei K. Alekseev, Alexander B. Ruchin, and Mikhail N. Esin. 2024. "Carabid Beetles under the Influence of Megafires in Pine and Secondary Forests of Central European Russia" Diversity 16, no. 7: 370. https://doi.org/10.3390/d16070370
APA StyleAleksanov, V. V., Alekseev, S. K., Ruchin, A. B., & Esin, M. N. (2024). Carabid Beetles under the Influence of Megafires in Pine and Secondary Forests of Central European Russia. Diversity, 16(7), 370. https://doi.org/10.3390/d16070370