
Exploring Ice Cave Biodiversity in Northeastern Italy

Abstract
1. Introduction
2. Materials and Methods
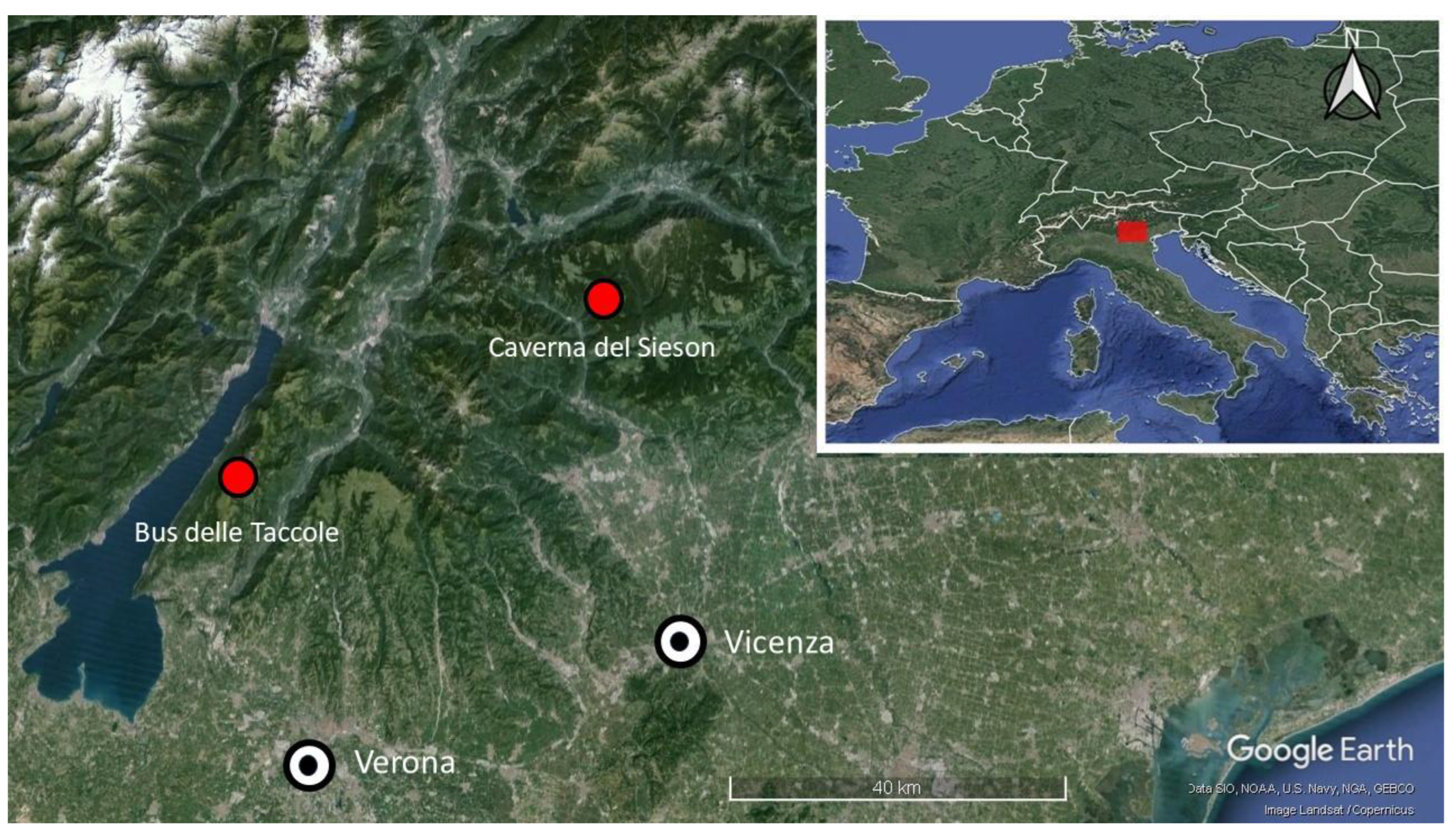
2.1. Study Area
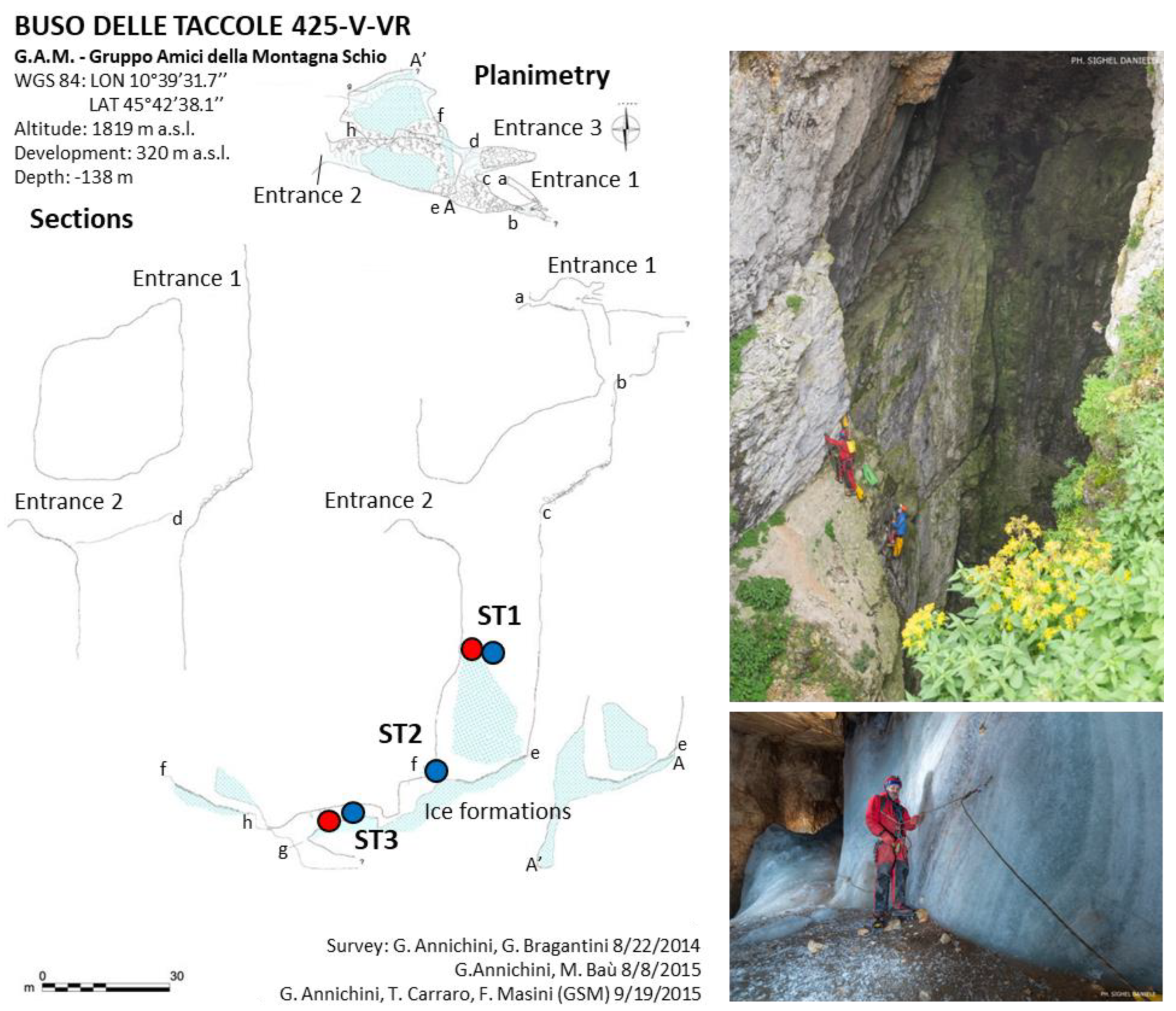
2.1.1. Bus Delle Taccole
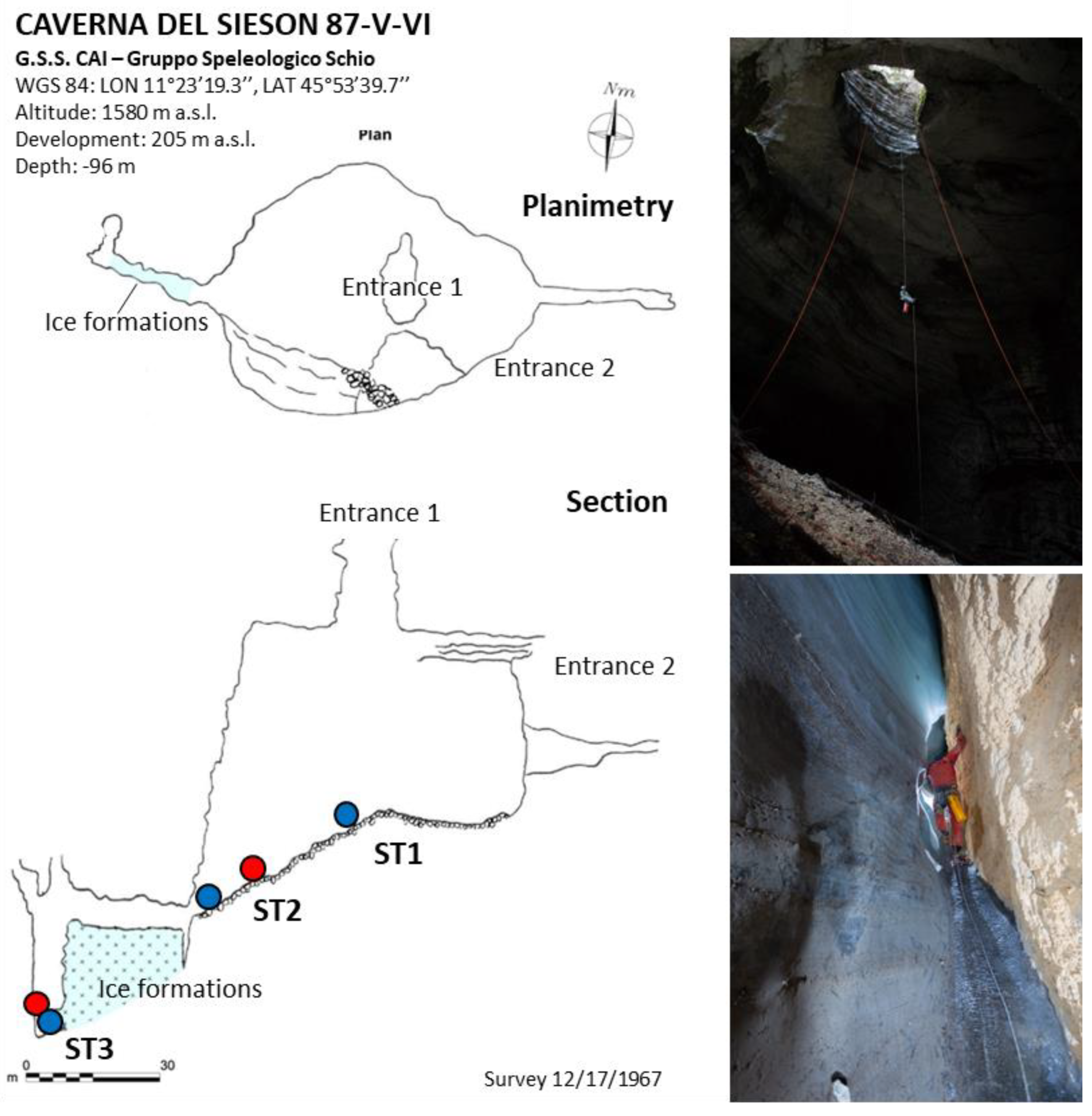
2.1.2. Caverna del Sieson
2.2. Specimens Sampling
2.3. Temperature Measurement
2.4. Ice Sampling and Isotopic Analyses
2.5. Data Analysis
3. Results
3.1. Subterranean Fauna
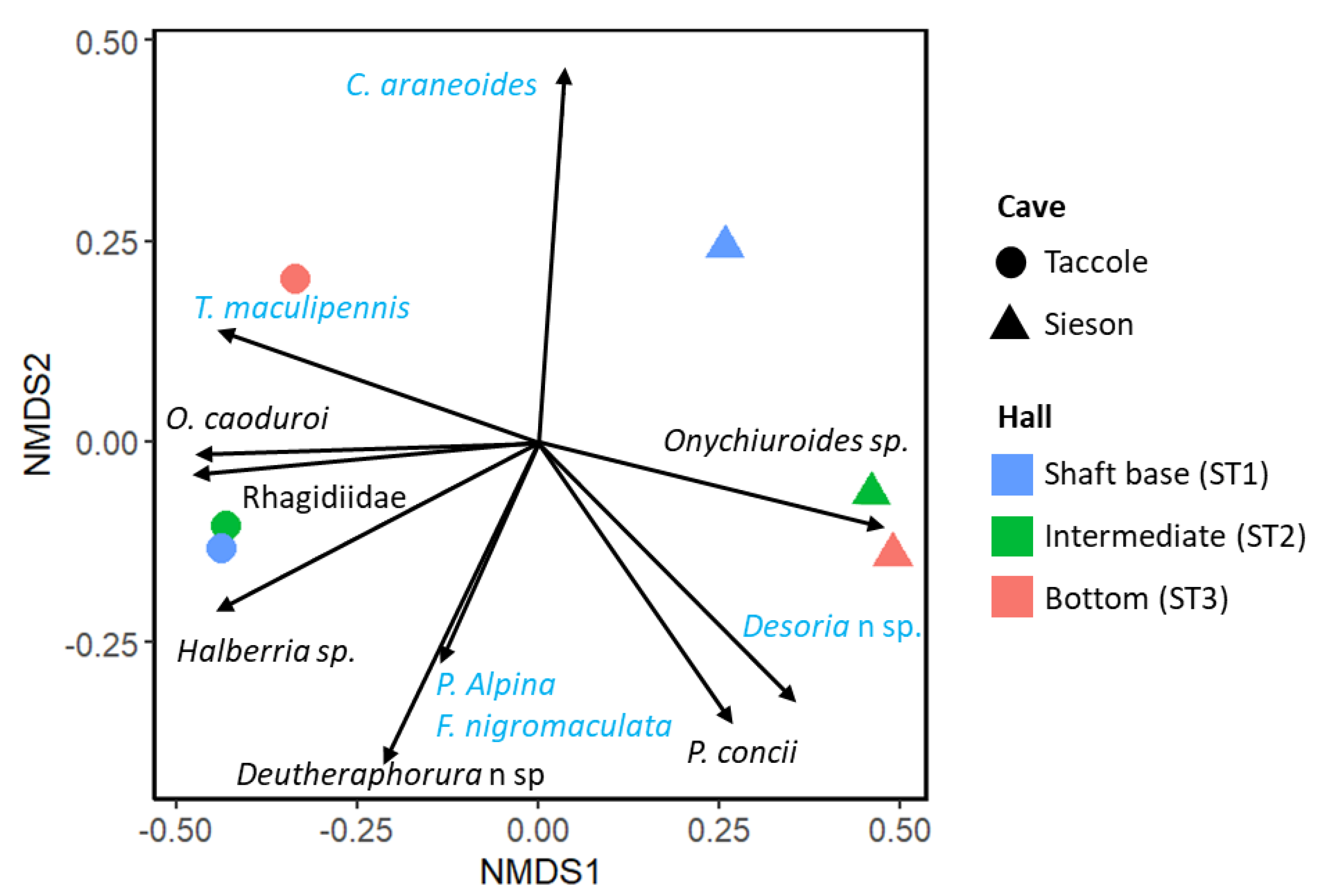
3.2. Distinct Cave Communities
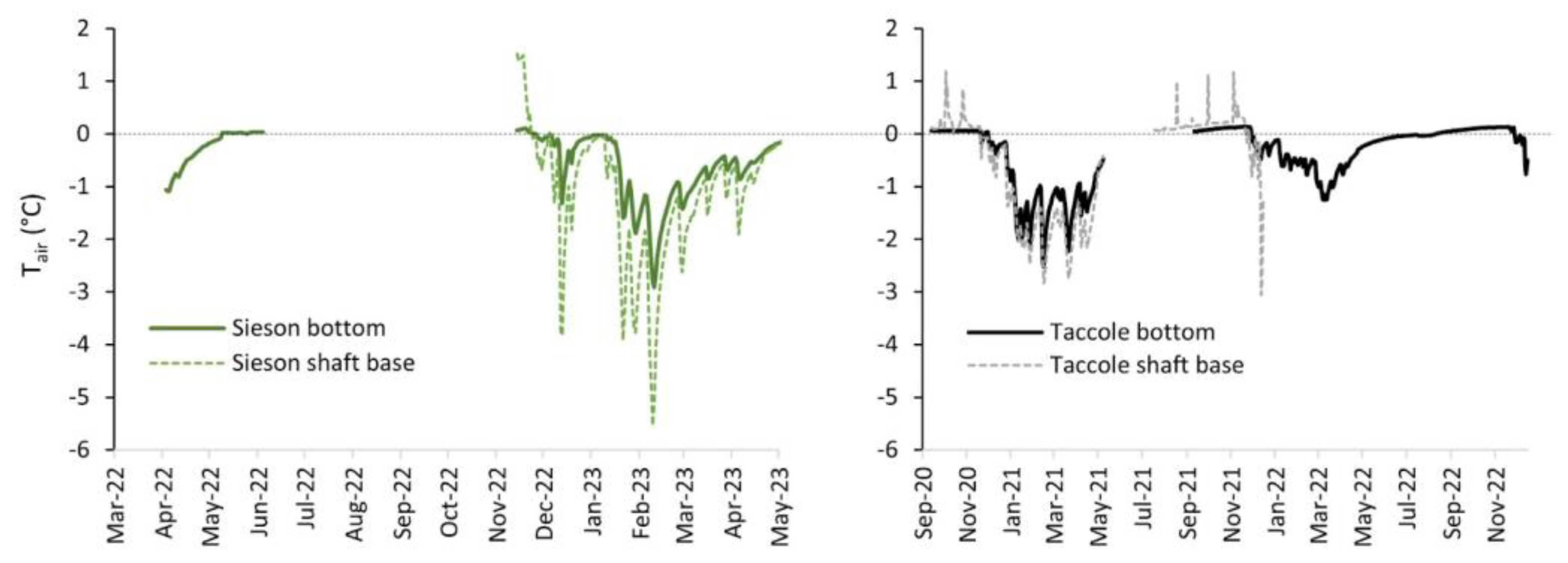
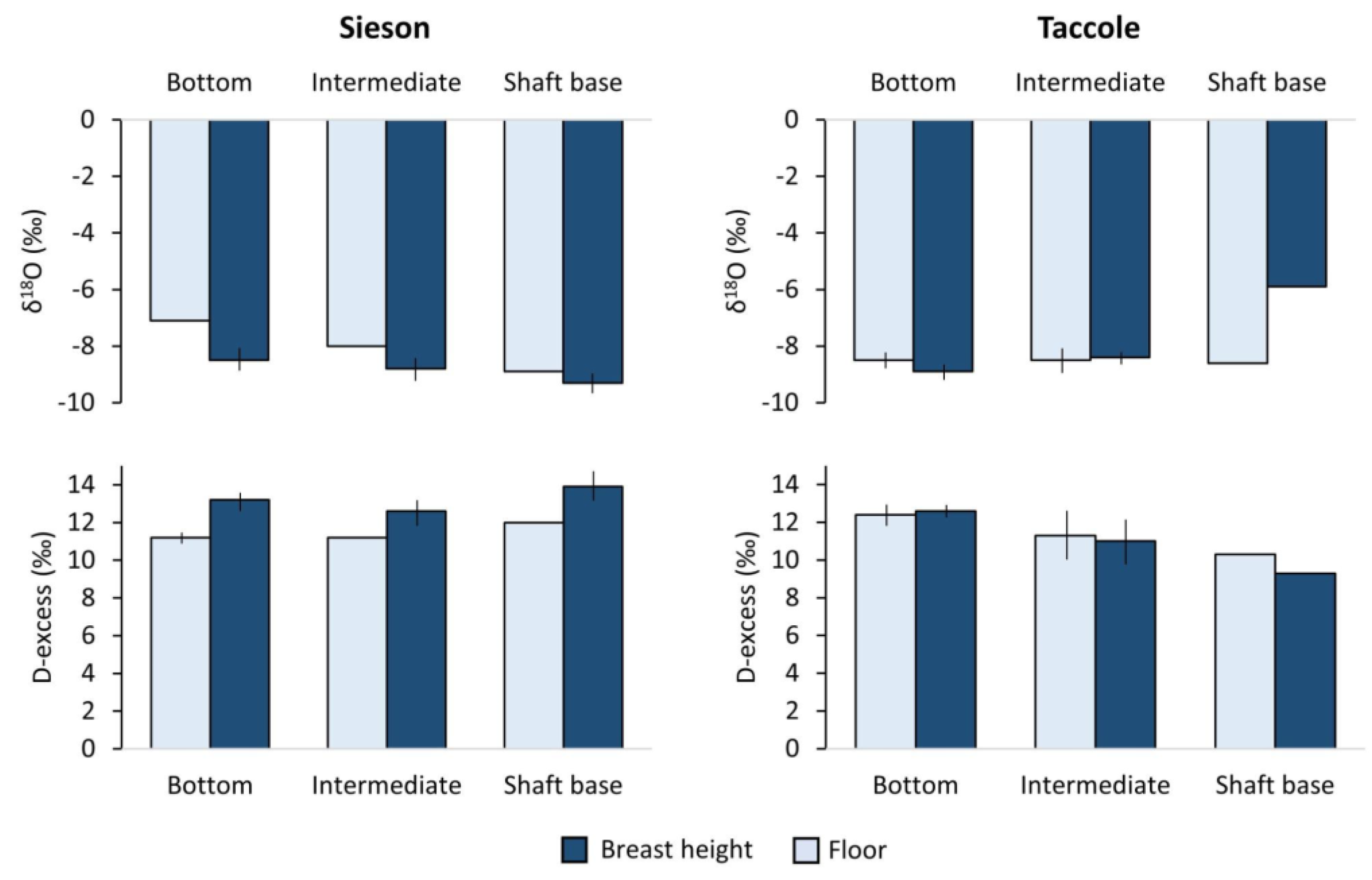
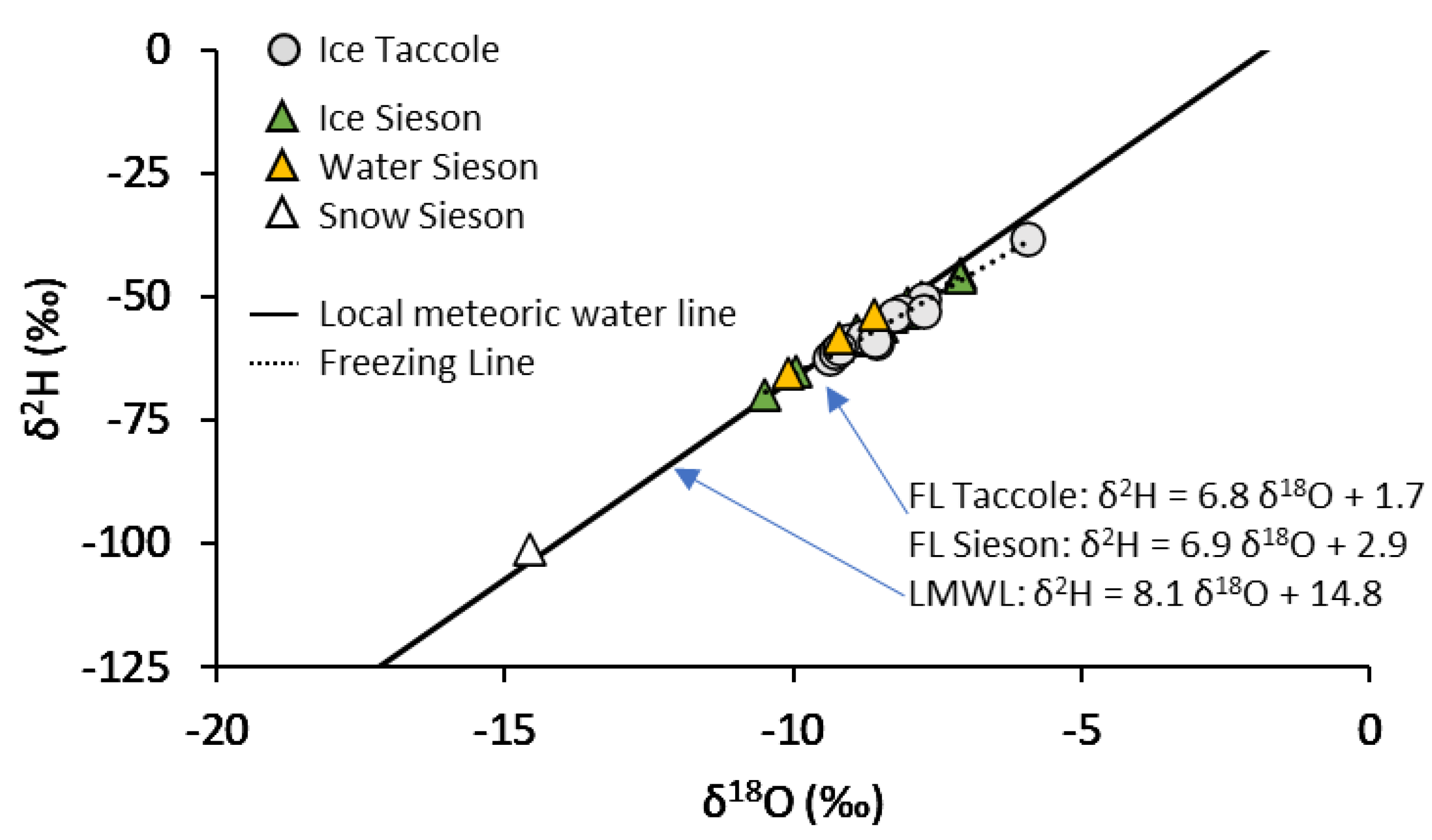
3.3. Environmental and Isotopic Conditions
4. Discussion
4.1. Ice Cave Fauna Composition
4.2. Relation to Environmental Variables
4.3. Heterogeneous Isotopic Fingerprint of Cave Ice
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Intergovernmental Panel on Climate Change [IPCC]. Special Report on the Ocean and Cryosphere in a Changing Climate; IPCC: Paris, France, 2019. [Google Scholar]
- Luetscher, M.; Jeannin, P.-Y. A process-based classification of alpine ice caves. Theor. Appl. Karstol. 2007, 17, 5–10. [Google Scholar]
- Kern, Z.; Fórizs, I.; Pavuza, R.; Molnár, M.; Nagy, B. Isotope Hydrological Studies of the Perennial Ice deposit of Saarhalle, Mammuthöhle, Dachstein Mts, Austria. Cryosphere 2011, 5, 291–298. [Google Scholar] [CrossRef]
- Perşoiu, A.; Onac, B.P.; Wynn, J.G.; Bojar, A.-V.; Holmgren, K. Stable isotope behavior during cave ice formation by water freezing in Scărişoara Ice Cave, Romania. J. Geophys. Res. 2011, 116, D02111. [Google Scholar] [CrossRef]
- Bădăluţă, C.-A.; Perşoiu, A.; Ionita, M.; Piotrowska, N. Stable Isotopes in Cave Ice Suggest Summer Temperatures in East-central Europe Are Linked to Atlantic Multidecadal Oscillation Variability. Clim. Past 2020, 16, 2445–2458. [Google Scholar] [CrossRef]
- Smith, D.F.; Lyons, W.B.; Welch, S.A.; Zorn, M.; Tičar, J.; Lipar, M.; Carey, A.E. The Chemistry of Cave Ice: Two Examples from Slovenia. Environ. Eng. Geosci. 2023, 29, 291–307. [Google Scholar]
- Holmlund, P.; Onac, B.P.; Hansson, M.; Holmgren, K.; Mörth, M.; Nyman, M.; Perşoiu, A. Assessing the palaeoclimate potential of cave glaciers: The example of the Scǎrişoara ice cave (Romania). Geogr. Ann. A 2005, 87, 193–201. [Google Scholar] [CrossRef]
- Perşoiu, A.; Buzjak, N.; Onaca, A.; Pennos, C.; Sotiriadis, Y.; Ionita, M.; Zachariadis, S.; Styllas, M.; Kosutnik, J.; Hegyi, A.; et al. Record summer rains in 2019 led to massive loss of surface and cave ice in SE Europe. Cryosphere 2021, 15, 2383–2399. [Google Scholar] [CrossRef]
- Colucci, R.R.; Fontana, D.; Forte, E.; Potleca, M.; Guglielmin, M. Response of ice caves to weather extremes in the southeastern Alps, Europe. Geomorphology 2016, 261, 1–11. [Google Scholar] [CrossRef]
- Maggi, V.; Colucci, R.R.; Scoto, F.; Giudice, G.; Randazzo, L. Ice caves in Italy. In Ice Caves; Elsevier: Amsterdam, The Netherland, 2018; pp. 399–423. [Google Scholar]
- Sivelli, M.; Vianelli, M. Abissi delle Alpi Apuane: Guida Speleologica; Società Speleologica Italiana: Bologna, Italy, 1982. [Google Scholar]
- Perșoiu, A. Ice caves climate. In Ice Caves; Elsevier: Amsterdam, The Netherlands, 2018. [Google Scholar]
- Iepure, S. Chapter 7—Ice Cave Fauna. In Ice Caves; Elsevier: Amsterdam, The Netherlands, 2018. [Google Scholar]
- Avesani, D.; Latella, L. Spatio-temporal distribution of the genus Chionea (Diptera, Limoniidae) in the Buso del Valon ice cave and other caves in the Lessini Mountains (Northern Italy). Boll. Mus. Civ. St. Nat. Verona 2016, 40, 11–16. [Google Scholar]
- Latella, L.; Zorzin, R.; Annichini, G.; Pellegrini, B. Il progetto “Buso del Vallon” negli Alti Lessini Veronesi. Speleo. Ven. 2019, 23, 175–179. [Google Scholar]
- Petri, I.; Ballarin, F.; Latella, L. Seasonal abundance and spatio-temporal distribution of the troglophylic harvestman Ischyropsalis ravasinii (Arachnida, Opiliones, Ischyropsalididae) in the Buso del Valon ice cave, Eastern Italian Prealps. Subt. Biol. 2022, 42, 151–164. [Google Scholar] [CrossRef]
- Latella, L. Analisi preliminare della fauna sotterranea delle Dolomiti di Brenta (TN). In Proceedings of the 17th Regional Symposium of Speleology of Trentino Alto Adige 2021, Trento, Italy, 20–21 November 2022; GS SAT: Trento, Italy, 2022. [Google Scholar]
- Marazzi, S. Atlante Orografico Delle Alpi—SOIUSA; Priuli and Verlucca: Torino, Italy, 2005. [Google Scholar]
- Pasa, A. Carsismo e Idrografia Carsica nel Gruppo del M. Baldo e nei Lessini Veronesi. C.N.R.; Centro Studi Geografia Fisica, Ricerche Sulla Morfologia e Idrografiacarsica, 5; Tipografia Mareggiani: Bologna, Italy, 1954; p. 150. [Google Scholar]
- Zorzin, R.; Agostini, L.; Montecchi, M.C.; Torri, P.; Accorsi, C.A. Tanella cave (Monte Baldo—Verona, Italy): A record of environmental data on the last glacial period. Acta Carsol. 2011, 40, 189–205. [Google Scholar] [CrossRef]
- Di Cuia, R.; Riva, A.; Scifoni, A.; Moretti, A.; Spötl, C.; Caline, B. Dolomite characteristics and diagenetic model of the Calcari Grigi Group (Asiago Plateau, Southern Alps–Italy): An example of multiphase dolomitization. Sedimentology 2011, 58, 1347–1369. [Google Scholar] [CrossRef]
- Cui, J.; Tian, L.; Gerlein-Safdi, C.; Qu, D. The influence of memory, sample size effects, and filter paper material on online laser-based plant and soil water isotope measurements. Rapid Commun. Mass Spectrom. 2017, 31, 509–522. [Google Scholar] [CrossRef]
- Penna, D.; Stenni, B.; Šanda, M.; Wrede, S.; Bogaard, T.A.; Michelini, M.; Fischer, B.M.C.; Gobbi, A.; Mantese, N.; Zuecco, G.; et al. Technical Note: Evaluation of between-sample memory effects in the analysis of δ2H and δ18O of water samples measured by laser spectroscopes. Hydrol. Earth Syst. Sci. 2012, 16, 3925–3933. [Google Scholar] [CrossRef]
- Oksanen, J.; Blanchet, F.G.; Kindt, R.; Legendre, P.; O’hara, R.B.; Simpson, G.L.; Solymos, P.; Stevens, M.H.H.; Wagner, H. Vegan: Community Ecology Package. 2022 R Package Version 2.6-4. Available online: https://CRAN.R-project.org/package=vegan (accessed on 20 March 2024).
- Dansgaard, W. Stable isotopes in precipitation. Tellus 1964, 16, 436–468. [Google Scholar] [CrossRef]
- Zuecco, G.; Marchina, C. Hydrological Data from Ressi Experimental Catchment (Italy). 2021. Available online: https://osf.io/n24dg/ (accessed on 12 March 2024).
- Marchina, C.; Zuecco, G.; Chiogna, G.; Bianchini, G.; Carturan, L.; Comiti, F.; Engel, M.; Natali, C.; Borga, M.; Penna, D. Alternative methods to determine the δ2H-δ18O relationship: An application to different water types. J. Hydrol. 2020, 587, 124951. [Google Scholar] [CrossRef]
- Zuecco, G.; Marchina, C.; Gelmini, Y.; Amin, A.; van Meerveld, H.J.; Penna, D.; Borga, M. Ressi experimental catchment: Ecohydrological research in the Italian pre-Alps. Hydrol. Process. 2021, 35, e14095. [Google Scholar] [CrossRef]
- Plachter, H. Cave-dwelling flies in Central Europe: Adaptation to environment, especially to low temperatures (Diptera, Nematocera: Trichoceridae et Sciaridae). Oecologia 1983, 58, 367–372. [Google Scholar] [CrossRef]
- Lauga-Reyrel, F.; Lauga, J. Collembola of Cold Pyrenean habitats. Eur. J. Soil Biol. 1995, 31, 217–229. [Google Scholar]
- Dahal, C.; Krzemińska, E.; Baez, M. Trichoceridae. In Hjorth Andersen MCT (Coordinator) Catalogo de los Diptera de España, Portugal y Andorra (Insecta); Monografias SEA 8: Zaragoza, Spain, 2002; p. 82. [Google Scholar]
- Mauries, J.P. Osellasoma, un remarquable nouveau genre de Diplopode cavernicole du Monte Baldo (Trentin) (Myriapoda: Diplopoda: Craspedosomida: Neoatractosomidae). Studi trentini di scienze naturali. Acta Biol. 1984, 61, 189–196. [Google Scholar]
- Zacharda, M.; Isaia, M.; Piva, E. New troglobitic species of the genus Troglocheles (Acari: Prostigmata: Rhagidiidae) from caves in northern Italy and Austria, with a key to adult species of the genus. J. Nat. Hist. 2011, 45, 641–666. [Google Scholar] [CrossRef]
- Humphreys, W.F. Relict fauna and their derivation. In Ecosystems of the World; Wilkens, H., Culver, D.C., Humphreys, W.F., Eds.; Subterranean Ecosystems; Elsevier: Amsterdam, The Netherlands, 2000; Volume 30, pp. 417–432. [Google Scholar]
- Latella, L.; Verdari, N.; Gobbi, M. Distribution of terrestrial cave-dwelling arthropods in two adjacent Prealpine Italian areas with different glacial histories. Zool. Stud. 2012, 51, 1113–1121. [Google Scholar]
- Lacelle, D.; Lauriol, B.; Clark, I.D. Formation of seasonal ice bodies and associated cryogenic carbonates in Caverne de l’Ours, Quebec, Canada: Kinetic isotope effects and pseudo-biogenic crystal structures. J. Cave. Karst. Stud. 2009, 71, 48–62. [Google Scholar]


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Class | Order | Family | Genus/Species/Subspecies | Status | Tac. St 1 | Tac. St 2 | Tac. St 3 | Sie. St 1 | Sie. St 2 | Sie. 3 |
|---|---|---|---|---|---|---|---|---|---|---|
| Arachnida | Trombidiformes | Rhagidiidae | Gen. sp. | Tb | 1 | 1 | 1 | 0 | 0 | 0 |
| Arachnida | Trombidiformes | Fam. | Gen. sp. | 1 | 1 | 0 | 0 | 0 | 1 | |
| Diplopoda | Chordeumatida | Neoatractosomatidae | Osellasoma caoduroi Mauries, 1984 | Tb | 1 | 1 | 1 | 0 | 0 | 0 |
| Collembola | Poduromorpha | Onychiuridae | Deutheraphorura n. sp. | Tb | 1 | 1 | 0 | 0 | 0 | 0 |
| Collembola | Poduromorpha | Onychiuridae | Onychiuroides n.sp. | Tb | 0 | 0 | 0 | 1 | 1 | 1 |
| Collembola | Poduromorpha | Onychiuridae | Hymenaphorura sp. | 0 | 1 | 0 | 0 | 0 | 0 | |
| Collembola | Poduromorpha | Hypogastruridae | Ceratophysella cf macrocantha | 0 | 1 | 1 | 0 | 0 | 0 | |
| Collembola | Poduromorpha | Hypogastruridae | Ceratophysella bengtssoni (Agren, 1904) | 0 | 0 | 1 | 0 | 0 | 0 | |
| Collembola | Poduromorpha | Neanuridae | Pseudachorudina alpina Stach, 1949 | Cy | 1 | 0 | 0 | 0 | 0 | 0 |
| Collembola | Entomobryomorpha | Isotomiidae | Folsomia nigrimaculata Najt, 1981 | Cy | 1 | 0 | 0 | 0 | 0 | 0 |
| Collembola | Entomobryomorpha | Isotomiidae | Desoria n. sp. | Cy | 0 | 0 | 0 | 1 | 1 | 1 |
| Collembola | Entomobryomorpha | Tomoceridae | Pogonognathellus flavescens (Tullberg, 1871) | 0 | 0 | 0 | 0 | 0 | 1 | |
| Collembola | Entomobryomorpha | Entomobryidae | Gen. sp. | 0 | 0 | 1 | 0 | 0 | 0 | |
| Collembola | Entomobryomorpha | Entomobryidae | Lepidocyrtus sp. | 0 | 1 | 0 | 0 | 0 | 0 | |
| Collembola | Entomobryomorpha | Entomobryidae | Pseudosinella concii Gising, 1950 | Tb | 0 | 0 | 0 | 0 | 1 | 1 |
| Collembola | Neelipleona | Neelidae | Megalothorax carpaticus Papáč and Kováč, 2013 | 0 | 0 | 0 | 0 | 1 | 0 | |
| Insecta | Coleoptera | Staphylinidae | Gen. sp. | 0 | 1 | 0 | 1 | 0 | 0 | |
| Insecta | Coleoptera | Staphylinidae | Pselaphinae Gen. sp. | 0 | 0 | 0 | 1 | 0 | 0 | |
| Insecta | Coleoptera | Leiodidae | Halberria n. sp. | Tb | 1 | 1 | 1 | 0 | 0 | 0 |
| Insecta | Coleoptera | Cucujidae | Gen. sp. | 1 | ||||||
| Insecta | Diptera | Trichoceridae | Trichocera maculipennis Meigen, 1818 | Cy | 1 | 1 | 1 | 1 | 1 | 1 |
| Insecta | Diptera | Limoniidae | Chionea araneoides (Dalman, 1816) | Cy | 0 | 0 | 1 | 0 | 1 | 1 |
| Thermal Parameter | Sieson Shaft Base | Sieson Bottom | Taccole Shaft Base | Taccole Bottom |
|---|---|---|---|---|
| Reference period | 9/14/22–5/16/23 | 9/14/22–5/16/23 | 9/13/20–5/8/21 9/11/21–12/15/21 | 9/13/20–5/8/21 9/11/21–12/15/21 |
| Tmin/Tmax | −5.5/4.2 °C | −2.9/4.3 °C | −3.1/1.2 °C | −2.5/1.0 °C |
| Tavg ± Tsd | −0.9 ± 1.3 °C | −0.4 ± 0.8 °C | −0.7 ± 1.0 °C | −0.5 ± 0.7 °C |
| Tmed | −0.8 °C | −0.5 °C | −0.3 °C | −0.2 °C |
| Taug | na | na | 0.14 ± 0.16 °C | 0.01 ± 0.03 °C |
| Freezing index * | 163 | 161 | 192 | 181 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Latella, L.; Brighenti, S. Exploring Ice Cave Biodiversity in Northeastern Italy. Diversity 2024, 16, 364. https://doi.org/10.3390/d16070364
Latella L, Brighenti S. Exploring Ice Cave Biodiversity in Northeastern Italy. Diversity. 2024; 16(7):364. https://doi.org/10.3390/d16070364
Chicago/Turabian StyleLatella, Leonardo, and Stefano Brighenti. 2024. "Exploring Ice Cave Biodiversity in Northeastern Italy" Diversity 16, no. 7: 364. https://doi.org/10.3390/d16070364
APA StyleLatella, L., & Brighenti, S. (2024). Exploring Ice Cave Biodiversity in Northeastern Italy. Diversity, 16(7), 364. https://doi.org/10.3390/d16070364