


Leaf Architecture in the Morphological Diversity of the Genus Prosopis in the Semi-Desert Area of Northeastern Mexico


 ,
,
Abstract
1. Introduction
2. Materials and Methods
2.1. Description of the Study Area
2.2. Description of Mesquite Species
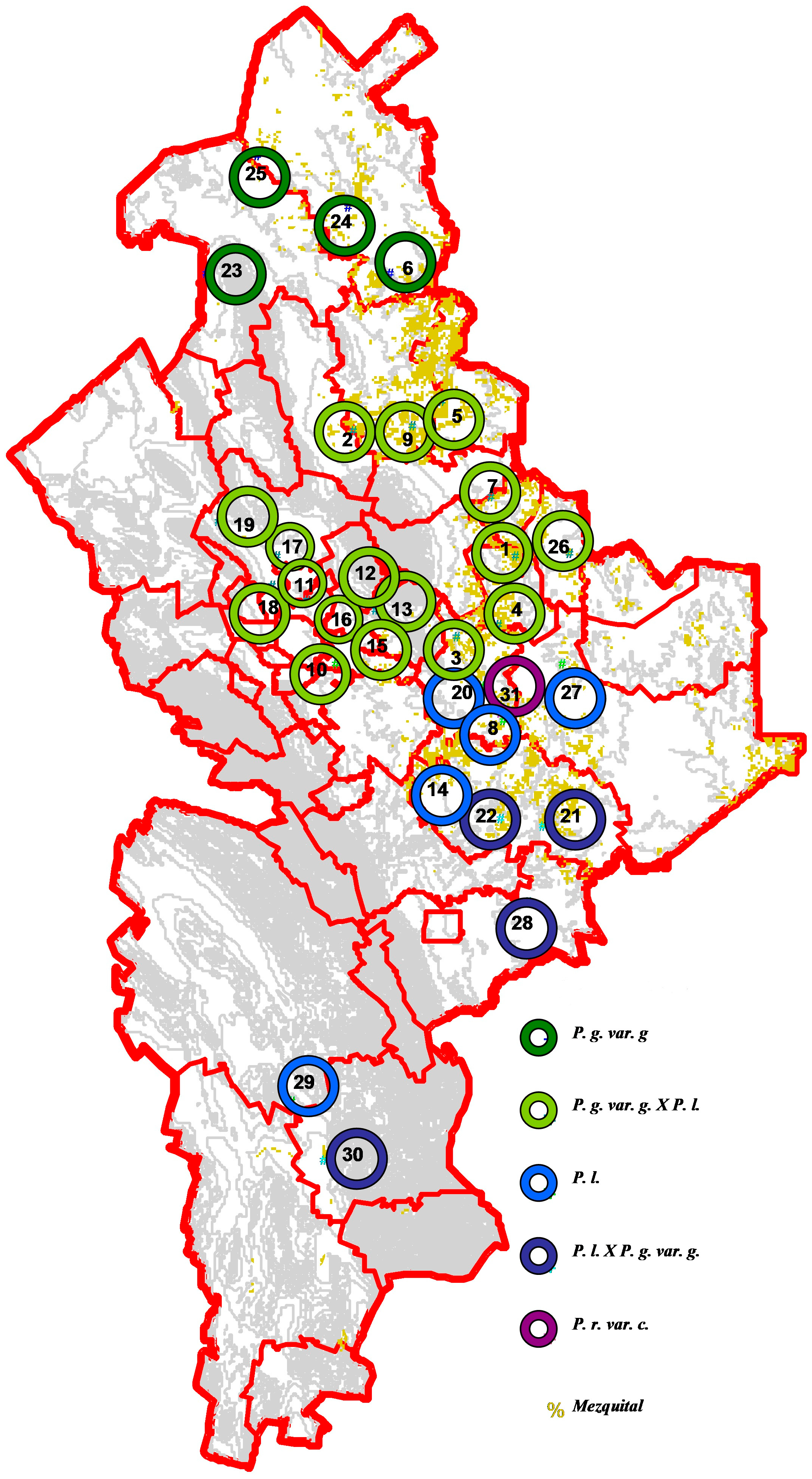
2.3. Sampling Design
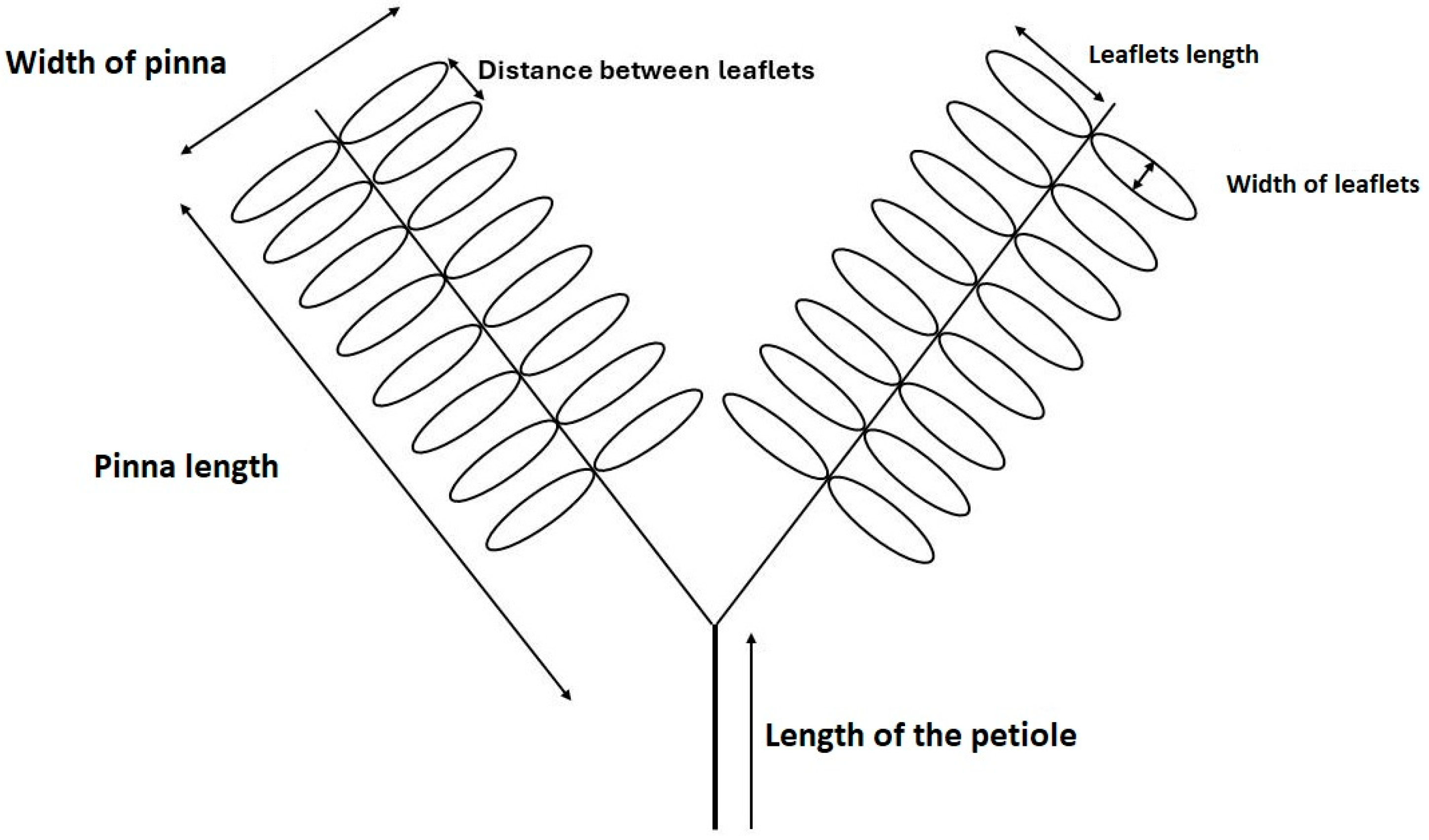
2.4. Study of Leaf Parameters
2.5. Statistical Design
3. Results
3.1. Traditional Taxonomic Identification
3.2. Analysis of Morphometric Parameters
3.3. Analysis of Variance on the Variability of Morphometric Parameters
3.4. Tukey Multiple Comparison of Means
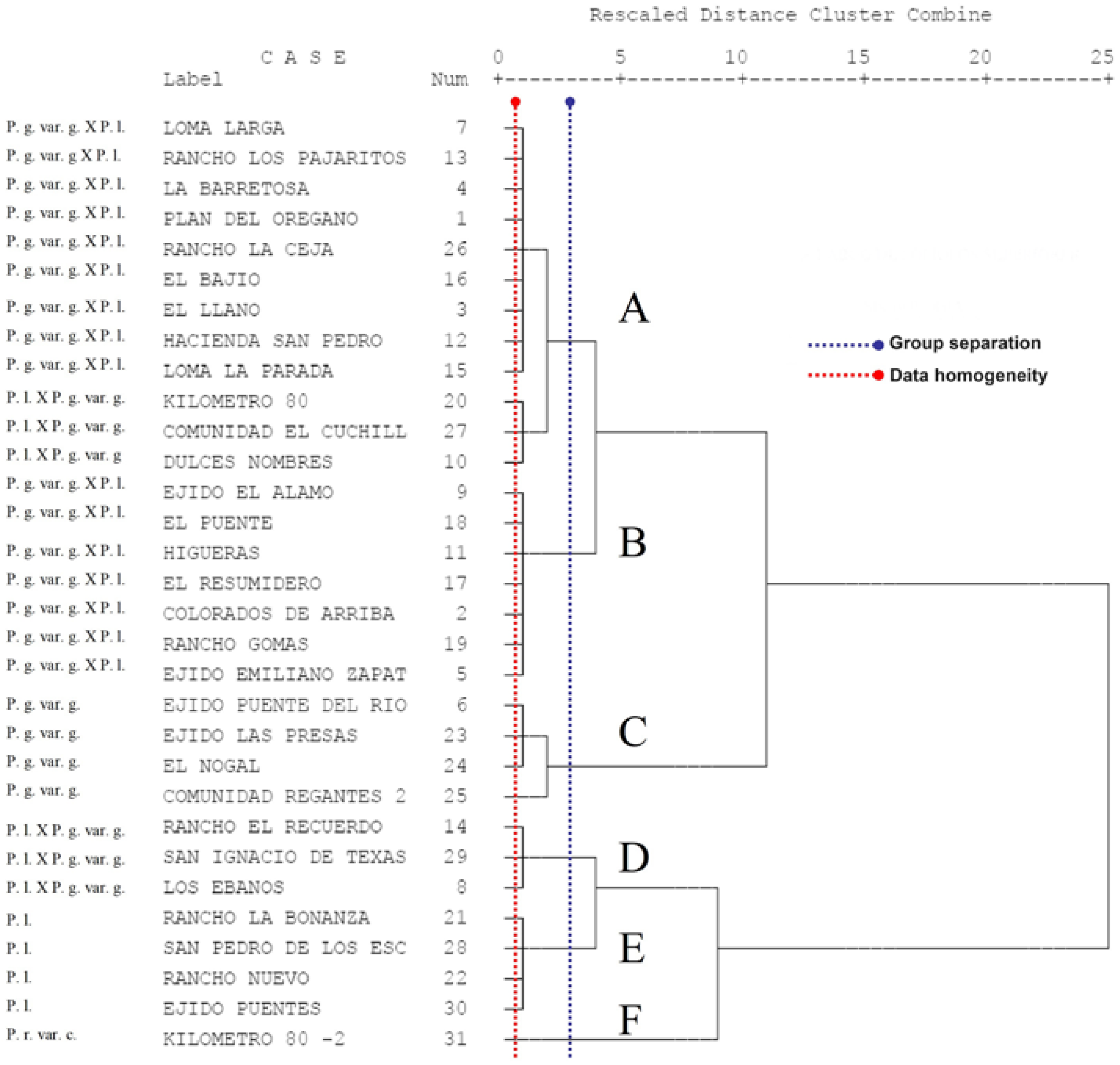
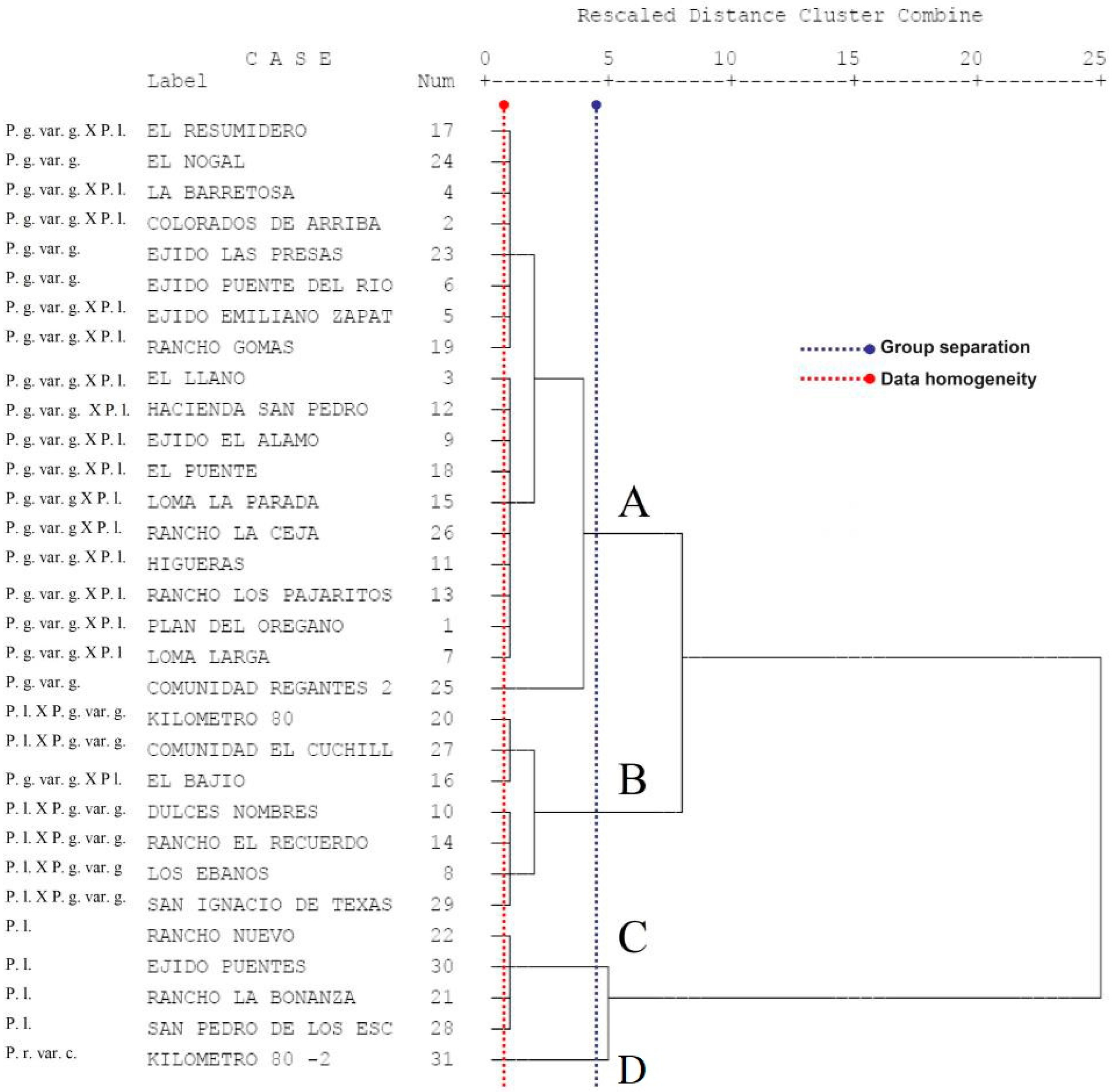
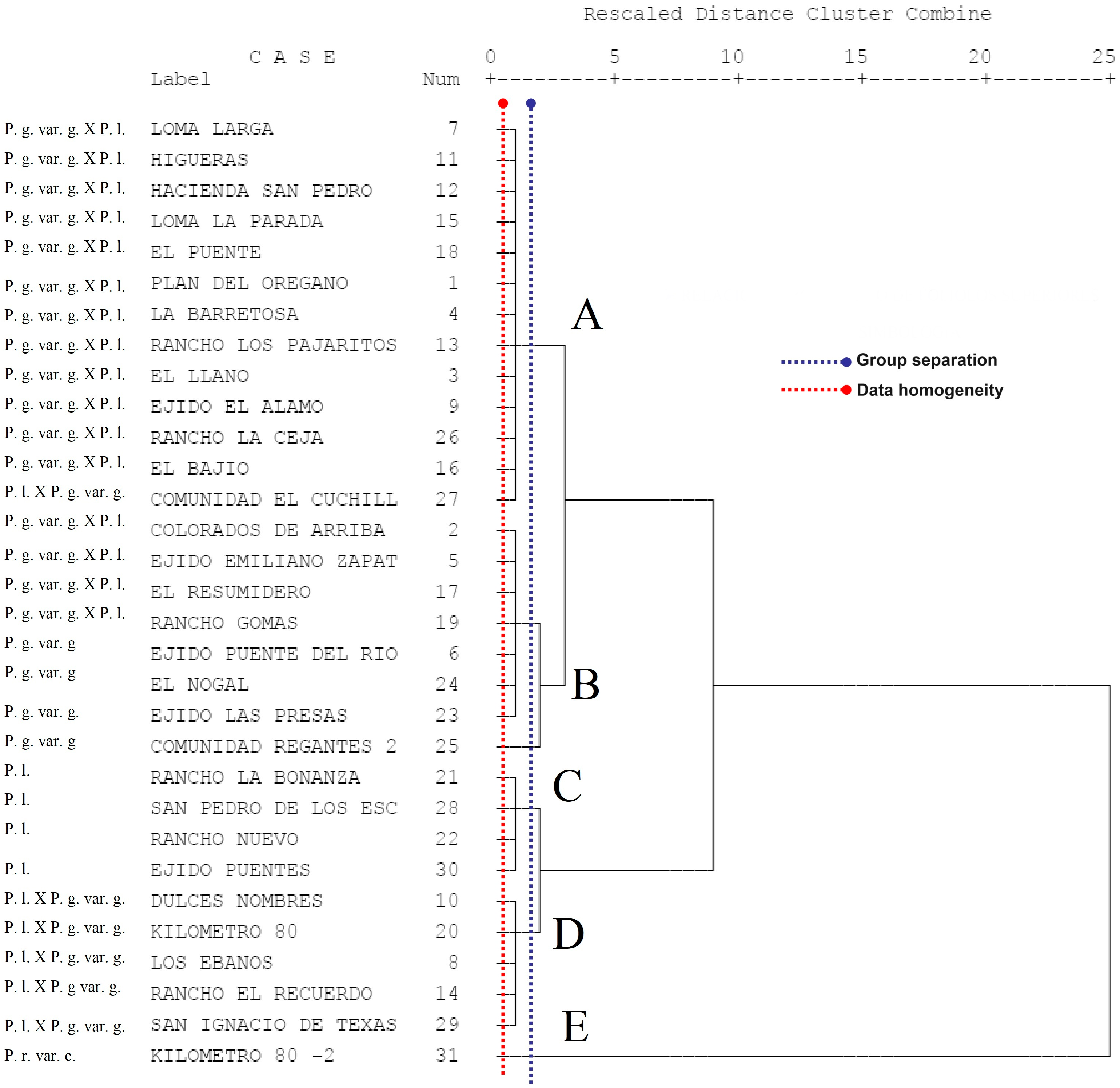
3.5. Grouping of Taxa Using Cluster Analysis
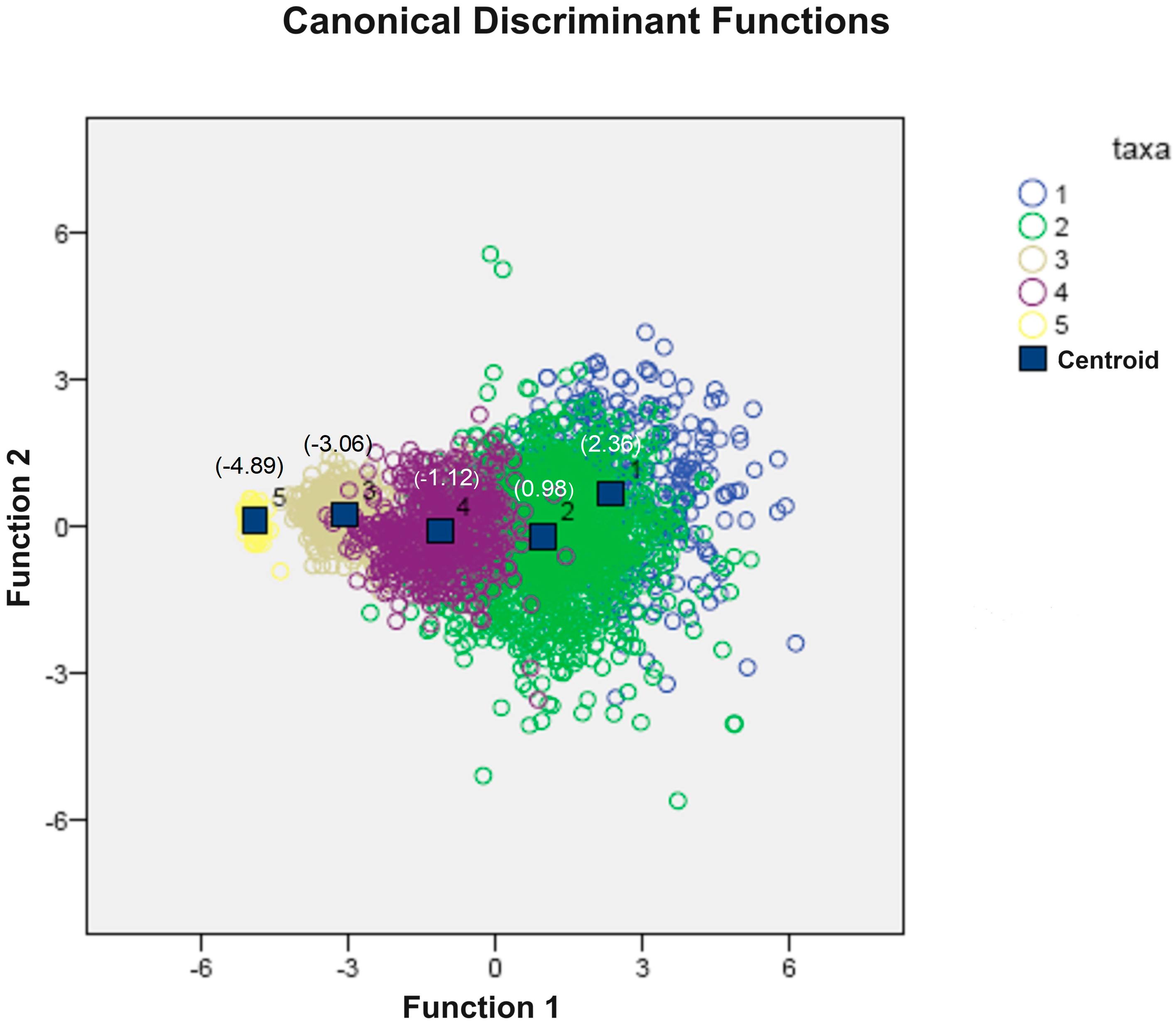
3.6. Discriminant Analysis
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Conflicts of Interest
References
- Sauceda, E.N.R.; Martínez, G.E.R.; Valverde, B.R.; Ruiz, R.M.; Hermida, M.D.L.C.C.; Torres, S.M.M.; Ruiz, H.H.P. Análisis técnico del árbol del mezquite (Prosopis laevigata Humb. & Bonpl. ex Willd.) en México. Ra Ximhai 2014, 10, 173–193. [Google Scholar]
- Golubov, J.; Mandujano, M.; Eguiarte, L.E. The paradox of mesquites (Prosopis spp.): Invading species of biodiversity enhancers? Bol. Soc. Bot. Mex. 2001, 69, 21–28. [Google Scholar] [CrossRef]
- Cates, G.; Rhoades, F. Prosopis laeves as a resource for insects. In Mezquite: Its Biology in Two Desert Ecosystem; Simpson, B.B., Ed.; Dowden, Hutchinson & Ross: Stroudsburg, PA, USA, 1977; pp. 61–83. [Google Scholar]
- Simpson, P.J. Datos sobre algunas características del mezquite (Prosopis laevigata) y su aprovechamiento en el Valle del Mezquital. In Mezquites y Huizaches; Instituto Mexicano de Recursos Naturales Renovables (IMRNR), México DF: Ciudad de México, México, 1977; p. 146. [Google Scholar]
- Langford, A.R. Uses of mezquite. In Literature on the Mezquite (Prosopis L.) of North America, and Annotated Bibliography; Shuster, J.L., Ed.; International Center for Arid and Semid-arid Studies, Texas Tech. University: Lubbock, TX, USA, 1969; pp. 20–22. [Google Scholar]
- Burkart, A. A monograph of the genus Prosopis (Mimosoideae). J. Arnold Arbor. 1976, 57, 450–525. [Google Scholar] [CrossRef]
- Graham, J.D. Morphological variation in the Mesquite (Prosopis, leguminosae) in the Lowlands of northeastern Mexico. Southwest. Nat. 1960, 5, 187–193. [Google Scholar] [CrossRef]
- Johnston, M.C. The North American mesquites. Prosopis sect. Algarobia (Leguminosae). Brittonia 1962, 14, 72–90. [Google Scholar] [CrossRef]
- Benson, L. The mesquites and screw-beans of the United States. Am. Jour. Bot. 1941, 28, 748–754. [Google Scholar]
- Bessega, C.; Vilardi, J.C.; Saidman, B.O. Genetic relationships among American species of the genus Prosopis (Mimosoideae, Leguminosae) inferred from ITS sequences: Evidence for long distance dispersal. J. Biogeogr. 2006, 33, 1905–1915. [Google Scholar] [CrossRef]
- Castillo, M.L.; Schaffner, U.; Van Wilgen, B.W.; Montaño, N.M.; Bustamante, R.O.; Cosacov, A.; Mathese, M.J.; Le Roux, J.J. Genetic insights into the globally invasive and taxonomically problematic tree genus Prosopis. AoB Plants 2021, 13, plaa069. [Google Scholar] [CrossRef] [PubMed]
- Trenchard, L.; Harris, P.; Smith, S.; Pasiecznik, N. A review of ploidy in the genus Prosopis (Leguminosae). Bot. J. Linn. Soc. 2008, 156, 425–438. [Google Scholar] [CrossRef]
- Fontana, M.L.; Pérez, V.R.; Luna, C.V. Características evolutivas en Prosopis spp.: Citogenética, genética e hibridaciones. Rodriguésia 2018, 69, 409–421. [Google Scholar]
- Ffolliott, P.F.; Thames, J.L. Manual Sobre Taxonomía de Prosopis en México, Perú y Chile; FAO: Roma, Italy, 1983. [Google Scholar]
- Gómez, L.F.; Poillon, J.S.; Abuín, M.M.C. Mezquites y Huizaches. In Algunos Aspectos de la Economía, Ecología Y Taxonomía de los Géneros Prosopis y Acacia en México; Instituto Mexicano de Recursos Naturales Renovables: Ciudad de México, Mexico, 1970; pp. 1–69. [Google Scholar]
- Gómez, M. Contenido de Carbohidratos en el Lignotuber de Cryptocarya alba (Mol.) Looser y Respuestas Morfológicas de los Rebrotes Producidos a Partir de el, Durante La Regeneración Postfuego, en el Matorral de Chile Central. Master’s Thesis, Facultad de Ciencias, Universidad de Chile, Santiago, Chile, 2003; 61p. [Google Scholar]
- Fahn, A. Plant Anatomy; Ediciones Pirámide S.A: Madrid, Spain, 1985; pp. 229–231. [Google Scholar]
- Silva, H.; Martínez, J.P.; Baginsky, C.; Pinto, M. Efecto del déficit hídrico en la Anatomía foliar de seis cultivares de poroto Phaseolus vulgaris. Rev. Chil. Hist. Nat. 1999, 72, 219–235. [Google Scholar]
- INEGI, Anuario Estadístico de Climas Nuevo León. Instituto Nacional de Estadística, Geografía e Informática, Aguascalientes, Mexico, 2002. Available online: https://www.inegi.org.mx/contenidos/productos/prod_serv/contenidos/espanol/bvinegi/productos/historicos/1334/702825157937/702825157937_1.pdf (accessed on 3 June 2023).
- Gámez, G.L. Análisis Genético del Mezquite (Prosopis spp.) por Medio de la Amplificación Enzimática al Azar de Polimorfismos Genómicos (RAPDs). Bachelor’s Thesis, Universidad Autónoma de Nuevo León, San Nicolás de los Garza, Mexico, 1995. [Google Scholar]
- González, E. Algunas Plantas Silvestres Comestibles en los Municipios de Mina, Linares y Dr. Arroyo. Bachelor’s Thesis, Facultad de Ciencias Biológicas, Universidad Autónoma de Nuevo León, San Nicolás de los Garza, Mexico, 1981. [Google Scholar]
- USDA. Agricultural Prices 2008 Summary. United States Department of Agicultura, 2009. Available online: http://plants.usda.gov/java/profile?symbol=PRGL2 (accessed on 3 June 2023).
- Villareal, Q.J.A. Vegetación del Municipio de los Ramones, Nuevo León, México. Bachelor’s Thesis, Facultad de ciencias Biológicas, Universidad Autónoma de Nuevo León, San Nicolás de los Garza, Mexico, 1979. [Google Scholar]
- Barneby, R.C. Atlas of North American Astragulus. Part I–II. Mem. N. Y. Bot. Gard. 1964, 13, 1–1188. [Google Scholar]
- Brizuela, M.M.; Burghardt, A.D.; Tanoni, D.; Palacios, R.A. Estudio de la Variación Morfológica en Tres Procedencias de Prosopis flexuosa y su Manifestación en Cultivo Bajo Condiciones Uniformes; Multiquenina, 009; Instituto Argentino de la Investigación de las Zonas Áridas: Mendoza, Argentina, 2000; pp. 7–15. [Google Scholar]
- Zar, J.H. Biostatistical Analysis, 3rd ed.; Prentice Hall, Inc.: Upper Saddle River, NJ, USA, 1996. [Google Scholar]
- Abrego, R.H.J. Estudio Fenológico del Mezquite (Prosopis spp. L.) en Cuatro Localidades del Estado de Nuevo León. Bachelor’s Thesis, Facultad de Agronomía, Universidad Autónoma de Nuevo León, San Nicolás de los Garza, Mexico, 1991. [Google Scholar]
- Galindo, A.S. Caracterización de la Variación en el Mezquite (Prosopis L.), y sus usos en el Altiplano Potosino. Bachelor’s Thesis, Facultad de Ciencias Biológicas, Universidad Autónoma de Nuevo León, San Nicolás de los Garza, Mexico, 1983. [Google Scholar]
- Leakey, R.; Last, T. Biology and patential of Prosopis especies in enviroments with particular reference to P. Cineraria. J. Arid. Environ. 1979, 3, 9–24. [Google Scholar] [CrossRef]
- Jiménez, V.I.A. Dinámica de Comunidades Vegetales con Mezquite Prosopis glandulosa Torr. en la zona Metropolitana de monterrey, N. L., Medidas para su Manejo y Conservación. Bachelor’s Thesis, Universidad Autónoma de Nuevo León, San Nicolás de los Garza, Mexico, 2002; pp. 133–143. [Google Scholar]
- López, V.A.P. Morfología y Anatomía de la Hoja del Mezquite (Prosopis glandulosa Torr.) en Nueve Localidades del Centro y norte del Estado de Nuevo León, México. Bachelor’s Thesis, Universidad Autónoma de Nuevo León, San Nicolás de los Garza, Mexico, 2006; pp. 59–78. [Google Scholar]
- Rzedowski, J. Analisis de la Distribución Geográfica del Complejo Prosopis (Leguminosae-Mimosoideae) en Norteamérica. Acta Botánica Mex. 1988, 3, 7–19. [Google Scholar] [CrossRef]
- Galindo, A.S.; García, M.E.; Wendt, T.L.; González, C.F.V. Potencial de Hibridación Natural en el Mezquite (Prosopis Laevigata y Prosopis glandulosa var. torreyana, Leguminosae) de la Altiplanicie de San Luis Potosí; Acta Botánica Mexicana. No. 020; Instituto Nacional de Ecologia A. C.: Patzcuaro, Mexico, 1992; pp. 107–117. [Google Scholar]
- Solbring, O.T.; Bawa, K.; Carman, N.; Hunziker, J.H.; Naranjo, C.A.; Palacios, R.A.; Poggio, L.; Simpson, B.B. Patterns of variation. In Mesquite: Its Biology in Two Desert Ecosystems; Simpson, B., Ed.; Dowen, Hutchinson and Ross: Stroudsburg, PA, USA, 1977; pp. 44–60. [Google Scholar]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Type or Subtype | Weather Symbol | % on Surface |
|---|---|---|
| Very warm and warm semi-dry | BS1(h) | 17.92 |
| Very warm and warm dry | BS(h′) | 17.15 |
| Semi-warm dry | BSh | 13.27 |
| Semi-warm semi-humid with little rain all year round | ACx | 10.44 |
| Semi-warm semi-humid with rain in summer | ACw | 8.66 |
| Temperate dry | BSk | 7.87 |
| Temperate semi-dry | BS1k | 6.65 |
| Semi-warm semi-dry | BS1h | 5.77 |
| Semi-warm very dry | BWh | 4.96 |
| Temperate semi-humid with rains in summer | C(w) | 4.52 |
| Temperate semi-humid with little rain all year round | Cx | 2.52 |
| Semi-cold semi- humid with rains in summer | C(E)(w) | 0.24 |
| Semi-cold semi-humid with little rain all year round | C(E)(x) | 0.03 |
| Species | Distribution * | Characteristics | Synonymy |
|---|---|---|---|
| P. glandulosa | Mina [20] Linares and Dr. Arroyo [21] | Oblong leaflets, less than five times longer than widths or lengths of 15 to 20 mm. Right sheath with recurved appendage, sometimes curved, yellow to brown [14]. | Prosopis juliflora (Swartz) DC P. juliflora var. glandulosa Cockerell P. juliflora var. torreyana L. Benson [22] |
| P. glandulosa var. torreyana | Iturbide [20] | Leaflets less than five times longer than wide, herbaceous, relatively tender. They are usually trees. Distribution: coastal areas of Mexico [14]. | Prosopis juliflora (Sw.) DC. var. torreyana L.D. Benson Prosopis odorata Torr. & Frém. [22] |
| P. glandulosa var. glandulosa | Mina and China [20] | Evergreen tree 1–10(–20) m high; one pair of pinnae per leaf: six to fifteen pairs of leaflets per pinna, obovate, oblong to narrow, glabrous, 30–40 mm long, 10–30 mm wide [14]. | Algarobia glandulosa (Torrey) Cooper. Neltuma constricta (Sarg.) Britton & Rose Prosopis chilensis (Molina) Stuntz variety glandulosa (Torrey) Standley Prosopis juliflora (Sw.) DC. var. constricta Sarg. [23] |
| P. laevigata | Mina, Linares, China and Dr. Arroyo [20], Los Ramones [23] | Small leaves, shorter than or equal to the length of the inflorescence. Dry, thin, flexible and soft pod, compressed between the segments and whitish in color. Leaves with one or two pairs of pinnae and six to twenty pairs of leaflets. Distribution: northwest Mexico and the Baja California peninsula [14]. | Acacia laevigata Willd. Algorobia dulces Benth Mimosa laevigata (Wild.) Poiret Mimosa rotundata Sessé & Mocino Nelthuma laevigata (Wild.) Britton & Rose Neltuma attenuata Prosopis dulcis Kunth [23] |
| P.reptans var. cinerascens | Bustamante, Villaldama [24]. | Smooth pods with numerous compact regular whorls, united singly or only a few in a cluster [14]. | Prosopis cinerascens (A. Gray) Benth. Strombocarpa cinerascens A. Gray [22] |
| SAMPLING SITES | UTM COORDINATES | ||||
|---|---|---|---|---|---|
| LOCALITY | MUNICIPALITY | East | North | Elevation | |
| 1 | Plan del Orégano | Melchor Ocampo | 45°76′94″ | 28°83′106″ | 152 |
| 2 | Colorados de Arriba | Vallecillo | 40°09′06″ | 29°27′506″ | 239 |
| 3 | El Llano | Los Ramones | 43°70′24″ | 28°54′589″ | 193 |
| 4 | La Barretosa | Los Herreras | 45°19′90″ | 28°54′589″ | 166 |
| 5 | Ej. Emiliano Zapata | Parás | 43°16′51″ | 29°37′498″ | 164 |
| 6 | Ej. Puente del Río Salado | Anáhuac | 41°34′60″ | 29°82′484″ | 146 |
| 7 | Loma Larga | General Treviño | 44°88′58″ | 29°04′109″ | 147 |
| 8 | Los Ébanos | Los Ramones | 45°35′17″ | 28°24′931″ | 196 |
| 9 | Ejido El Álamo | Vallecillo | 42°10′68″ | 29°29′095″ | 195 |
| 10 | Dulces Nombres | Pesquería | 39°42′36″ | 28°44′953″ | 351 |
| 11 | Hacienda San Pedro | General Zuazua | 38°40′66″ | 28°67′217″ | 369 |
| 12 | Higueras | Higueras | 39°81′19″ | 28°72′672″ | 503 |
| 13 | Los Pajaritos | Doctor González | 40°88′54″ | 28°63′751″ | 423 |
| 14 | Rancho el Recuerdo | General Terán | 42°76′56″ | 28°06′597″ | 285 |
| 15 | Loma la Parada | Marín | 40°18’47″ | 28°56’144″ | 323 |
| 16 | El Bajío | Marín | 40°21′22″ | 28°58′136″ | 342 |
| 17 | El Resumidero | Salinas Victoria | 37°37′12″ | 28°82′558″ | 451 |
| 18 | El Puente | Salinas Victoria | 37°22′52″ | 28°72′246″ | 424 |
| 19 | Rancho Gomas | Salinas Victoria | 35°31′64″ | 28°95′485″ | 191 |
| 20 | Km80 ** | Los Ramones | 44°60′57″ | 28°37′409″ | 191 |
| 21 | Rancho La bonanza | General Terán | 46°71′87″ | 27°87′506″ | 217 |
| 22 | Rancho Nuevo | General Terán | 45°28′41″ | 27°90′393″ | 259 |
| 23 | Ejido las Presas | Lampazos | 34°84′05″ | 29°82′066″ | 341 |
| 24 | El Nogal | Anáhuac | 39°91′25″ | 30°05′741″ | 177 |
| 25 | Regantes 26 | Anáhuac | 36°71′40″ | 30°24′331″ | 228 |
| 26 | Rancho la Ceja | Los Aldama | 47°69′56″ | 28°83′426″ | 112 |
| 27 | China | China | 47°44′39″ | 28°45′014″ | 138 |
| 28 | San pedro de los Escobedos | Linares | 46°23′56″ | 27°60′631″ | 261 |
| 29 | San Ignacio de Texas | Galeana | 37°90′92″ | 26°90′496″ | 1684 |
| 30 | Ejido las Puentes | Aramberri | 39°06′06″ | 26°70′446″ | 1581 |
| 31 | Km80 ** | Los Ramones | 44°91′25″ | 28°50′504″ | 188 |
| NAME | ABBREVIATION | |
|---|---|---|
| TAXA 1 | Prosopis glandulosa var. glandulosa | P. g. var. g. |
| TAXA 2 | Prosopis glandulosa var. glandulosa x Prosopis laevigata | P. g. var. g. x P. l. |
| TAXA 3 | Prosopis laevigata | P. l. |
| TAXA 4 | Prosopis laevigata x Prosopis glandulosa var. glandulosa | P. l. x P. g. var. g. |
| TAXA 5 | Prosopis reptans var. cinerascens | P. r. var. c. |
| Site | Locality—Municipality | Taxa * | Length | Length/Width Ratio | Spacing | ||
|---|---|---|---|---|---|---|---|
| Middle | Upper | Middle | Upper | ||||
| 6 | Ejido Puente del Río Salado—Anáhuac | P.g. var. g. | 32.40 ± 0.43 | 26.87 ± 0.49 | 11.38 ± 0.11 | 9.69 ± 0.11 | 9.38 ± 0.19 |
| 23 | Ejido las Presas—Lampazos | P.g. var. g. | 31.50 ± 0.50 | 27.39 ± 0.50 | 11.15 ± 0.15 | 9.39 ± 0.13 | 8.58 ± 0.22 |
| 24 | El Nogal—Anáhuac | P.g. var. g. | 31.49 ± 0.32 | 28.18 ± 0.42 | 10.98 ± 0.15 | 9.76 ± 0.14 | 8.47 ± 0.14 |
| 25 | Regantes 26—Anáhuac | P.g. var. g. | 36.47 ± 0.36 | 30.77 ± 0.38 | 12.17 ± 0.13 | 10.58 ± 0.11 | 9.55 ± 0.17 |
| 1 | Plan del Orégano—Melchor Ocampo | P.g. var. g X P.l. | 26.02 ± 0.26 | 19.41 ± 0.31 | 9.88 ± 0.11 | 7.84 ± 0.10 | 7.47 ± 0.14 |
| 2 | Colorados de Arriba—Vallecillo | P.g. var. g. X P.l. | 29.15 ± 0.39 | 23.27 ± 0.38 | 11.08 ± 0.12 | 8.82 ± 0.11 | 7.91 ± 0.17 |
| 3 | El Llano—Los Ramones | P.g. var. g X P.l. | 24.69 ± 0.33 | 18.80 ± 0.35 | 10.23 ± 0.09 | 8.02 ± 0.09 | 6.77 ± 0.10 |
| 4 | La Barretosa—Los Herrera | P.g. var. g. X P.l. | 26.32 ± 0.36 | 19.90 ± 0.35 | 10.92 ± 0.12 | 8.10 ± 0.10 | 7.38 ± 0.12 |
| 5 | Ejido Emiliano Zapata—Parás | P.g. var. g. X P.l. | 30.15 ± 0.46 | 24.73 ± 0.39 | 10.78 ± 0.13 | 8.83 ± 0.10 | 8.38 ± 0.22 |
| 7 | Loma Larga—Gral. Treviño | P.g. var. g. X P.l. | 24.83 ± 0.28 | 19.90 ± 0.29 | 9.54 ± 0.09 | 7.78 ± 0.10 | 6.80 ± 0.10 |
| 9 | Ejido El Álamo—Vallecillo | P.g. var. g. X P.l. | 28.43 ± 0.41 | 21.16 ± 0.38 | 10.16 ± 0.11 | 8.04 ± 0.11 | 7.97 ± 0.15 |
| 10 | Dulces Nombres—Pesquería | P.g. var. g. X P.l. | 22.29 ± 0.29 | 17.26 ± 0.39 | 7.66 ± 0.09 | 6.04 ± 0.10 | 6.96 ± 0.22 |
| 11 | Hacienda San Pedro—Gral. Zuazua | P.g. var. g. X P.l. | 30.67 ± 0.38 | 22.40 ± 0.39 | 9.84 ± 0.12 | 7.79 ± 0.12 | 8.54 ± 0.16 |
| 12 | Higueras—Higueras | P.g. var. g. X P.l. | 28.27 ± 0.30 | 18.81 ± 0.36 | 10.27 ± 0.15 | 7.74 ± 0.14 | 7.59 ± 0.10 |
| 13 | Los Pajaritos—Dr. González | P.g. var. g. X P.l. | 25.64 ± 0.37 | 19.90 ± 0.39 | 9.81 ± 0.10 | 8.07 ± 0.10 | 7.34 ± 0.19 |
| 15 | Loma la Parada—Marín | P.g. var. g. X P.l. | 23.28 ± 0.31 | 18.62 ± 0.44 | 9.74 ± 0.09 | 7.90 ± 0.10 | 6.90 ± 0.14 |
| 16 | El Bajío—Marín | P.g. var. g. X P.l. | 23.86 ± 0.41 | 19.35 ± 0.39 | 8.69 ± 0.10 | 7.19 ± 0.10 | 7.27 ± 0.17 |
| 17 | El Resumidero—Salinas Victoria | P.g. var. g. X P.l. | 28.48 ± 0.30 | 22.79 ± 0.36 | 11.00 ± 0.14 | 8.92 ± 0.12 | 8.15 ± 0.17 |
| 18 | El Puente—Salinas Victoria | P.g. var. g. X P.l. | 27.36 ± 0.32 | 21.81 ± 0.34 | 10.04 ± 0.12 | 7.91 ± 0.09 | 7.94 ± 0.14 |
| 19 | Rancho Gomas—Salinas Victoria | P.g. var. g. X P.l. | 28.09 ± 0.43 | 23.90 ± 0.47 | 10.53 ± 0.09 | 8.89 ± 0.08 | 9.20 ± 0.24 |
| 26 | Rancho la Ceja—Los Aldama | P.g. var. g. X P.l. | 23.80 ± 0.31 | 19.43 ± 0.35 | 9.77 ± 0.13 | 7.98 ± 0.10 | 6.95 ± 0.15 |
| 21 | Rancho La Bonanza—Gral. Terán | P. l. | 9.19 ± 0.09 | 7.04 ± 0.09 | 5.31 ± 0.05 | 4.04 ± 0.05 | 2.24 ± 0.06 |
| 22 | Rancho Nuevo—Gral. Terán | P. l. | 11.45 ± 0.12 | 8.83 ± 0.11 | 5.87 ± 0.05 | 4.53 ± 0.04 | 2.73 ± 0.06 |
| 28 | San Pedro de los Escobedos—Linares | P. l. | 9.20 ± 0.13 | 7.07 ± 0.11 | 5.45 ± 0.05 | 3.99 ± 0.03 | 2.20 ± 0.05 |
| 30 | Ejido las Puentes—Aramberri | P. l. | 11.47 ± 0.19 | 9.21 ± 0.21 | 5.74 ± 0.07 | 4.46 ± 0.06 | 3.33 ± 0.08 |
| 8 | Los Ébanos—Los Ramones | P.l. X P. g. var. g. | 17.96 ± 0.29 | 13.41 ± 0.26 | 7.89 ± 0.07 | 5.76 ± 0.06 | 5.06 ± 0.12 |
| 14 | Rancho el Recuerdo—Gral. Terán | P.l. X P. g. var. g. | 16.69 ± 0.24 | 12.44 ± 0.25 | 7.60 ± 0.08 | 5.83 ± 0.08 | 4.98 ± 0.09 |
| 20 | Km80 **—Los Ramones | P.l. X P. g. var. g. | 19.36 ± 0.23 | 12.06 ± 0.21 | 7.08 ± 0.10 | 5.49 ± 0.09 | 4.72 ± 0.10 |
| 27 | China—China | P.l. X P. g. var. g. | 20.31 ± 0.23 | 15.07 ± 0.22 | 8.48 ± 0.06 | 6.27 ± 0.05 | 4.70 ± 0.09 |
| 29 | San Ignacio de Texas—Galeana | P.l. X P. g. var. g. | 14.79 ± 0.18 | 15.94 ± 0.21 | 8.56 ± 0.10 | 6.65 ± 0.08 | 4.90 ± 0.08 |
| 31 | Km80 **—Los Ramones | P.r. var. c. | 4.63 ± 0.04 | 2.76 ± 0.03 | 3.46 ± 0.04 | 1.59 ± 0.01 | 1.00 ± 0.02 |
| Variable | Source of Variation | Degrees of Freedom | Mean Squares | Values of F |
|---|---|---|---|---|
| Petiole length | Between groups | 30 | 36,051.91 | 136.47 *** |
| Within the group | 6245 | 264.17 | ||
| Total | 6275 | |||
| Pinna width | Between groups | 30 | 53,651.75 | 591.38 *** |
| Within the group | 6245 | 90.72 | ||
| Total | 6275 | |||
| Pinna number | Between groups | 30 | 53,651.75 | 45.50 *** |
| Within the group | 6245 | 90.72 | ||
| Total | 6275 | |||
| Number of leaflets per pinna | Between groups | 30 | 9203.88 | 373.45 *** |
| Within the group | 6245 | 24.64 | ||
| Total | 6275 | |||
| Rachis length | Between groups | 30 | 89,633.42 | 248.71 *** |
| Within the group | 6245 | 360.39 | ||
| Total | 6275 | |||
| Lower leaflet length | Between groups | 30 | 8445.18 | 374.70 *** |
| Within the group | 6245 | 22.54 | ||
| Total | 6275 | |||
| Lower leaflet width | Between groups | 30 | 27.84 | 150.42 *** |
| Within the group | 6244 | 0.18 | ||
| Total | 6274 | |||
| Lower leaflet length/widthratio | Between groups | 30 | 866.64 | 336.33 *** |
| Within the group | 6244 | 2.57 | ||
| Total | 6274 | |||
| Middle leaflet length | Between groups | 30 | 13,108.13 | 655.73 *** |
| Within the group | 6245 | 19.99 | ||
| Total | 6275 | |||
| Middle leaflet width | Between groups | 30 | 37.05 | 212.04 *** |
| Within the group | 6245 | 0.17 | ||
| Total | 6275 | |||
| Middle leaflets Length/width ratio | Between groups | 30 | 956.32 | 440.65 *** |
| Within the group | 6245 | 2.17 | ||
| Total | 6275 | |||
| Upper leaflets length | Between groups | 30 | 9484.38 | 425.94 *** |
| Within the group | 6245 | 22.26 | ||
| Total | 6275 | |||
| Upper leaflets width | Between groups | 30 | 26.19 | 92.01 *** |
| Within the group | 6245 | 0.28 | ||
| Total | 6275 | |||
| Upper leaflet length/widthratio | Between groups | 30 | 843.55 | 470.15 *** |
| Within the group | 6245 | 1.79 | ||
| Total | 6275 | |||
| Lower leaflet spacing | Between groups | 30 | 494.72 | 259.41 *** |
| Within the group | 3107 | 1.90 | ||
| Total | 3137 | |||
| Middle leaflet spacing | Between groups | 30 | 560.84 | 284.62 *** |
| Within the group | 3107 | 1.97 | ||
| Total | 3137 | |||
| Upper leaflet spacing | Between groups | 30 | 693.32 | 251.14 *** |
| Within the group | 3107 | 2.76 | ||
| Total | 3137 |
| Length of Middle Leaflets | Length of Upper Leaflets | ||||||
|---|---|---|---|---|---|---|---|
| Site | Locality | Taxa | Mean | Site | Locality | Taxa | Mean |
| 31 | Km 80 ** | P.r. var. c. | 4.63 A * | 31 | Km 80 ** | P.r. var. c. | 2.76 A * |
| 21 | Rancho La Bonanza | P. l. | 9.18 B | 21 | Rancho La Bonanza | P. l. | 7.03 B |
| 28 | San pedro de los Escobedos | P. l. | 9.20 B | 28 | San pedro de los Escobedos | P. l. | 7.06 BC |
| 22 | Rancho Nuevo | P. l. | 11.45 C | 22 | Rancho Nuevo | P. l. | 8.83 CD |
| 30 | Ejido las Puentes | P. l. | 11.47 C | 30 | Ejido las Puentes | P. l. | 9.20 D |
| 29 | San Ignacio de Texas | P.l. X P. g. var. g. | 14.78 D | 29 | San Ignacio de Texas | P.l. X P. g. var. g. | 12.06 E |
| 14 | Rancho el Recuerdo | P.l. X P. g. var. g. | 16.68 E | 14 | Rancho el Recuerdo | P.l. X P. g. var. g. | 12.44 E |
| 8 | Los Ébanos | P.l. X P. g. var. g. | 17.96 EF | 8 | Los Ébanos | P.l. X P. g. var. g. | 13.40 EF |
| 20 | Km 80 ** | P.l. X P. g. var. g. | 19.36 FG | 20 | Km 80 ** | P.l. X P. g. var. g. | 15.06 FG |
| 27 | China | P.l. X P. g. var. g. | 20.30 G | 27 | China | P.l. X P. g. var. g. | 15.93 GH |
| 10 | Dulces Nombres | P.g. var. g. X P.l. | 22.29 H | 10 | Dulces Nombres | P.g. var. g. X P.l. | 17.26 HI |
| 15 | Loma la Parada | P.g. var. g. X P.l. | 23.28 HI | 15 | Loma la Parada | P.g. var. g. X P.l. | 18.62 IJ |
| 26 | Rancho la Ceja | P.g. var. g. X P.l. | 23.80 HI | 3 | El Llano | P.g. var. g. X P.l. | 18.79 IJ |
| 16 | El Bajío | P.g. var. g. X P.l. | 23.86 HI | 12 | Higueras | P.g. var. g. X P.l. | 18.80 IJ |
| 3 | El Llano | P.g. var. g. X P.l. | 24.69 IJ | 16 | El Bajío | P.g. var. g. X P.l. | 19.35 J |
| 7 | Loma Larga | P.g. var. g. X P.l. | 24.82 IJ | 1 | Plan del Orégano | P.g. var. g. X P.l. | 19.41 JK |
| 13 | Los Pajaritos | P.g. var. g. X P.l. | 25.64 J | 26 | Rancho la Ceja | P.g. var. g. X P.l. | 19.43 JK |
| 1 | Plan del Orégano | P.g. var. g X P.l. | 26.02 JK | 7 | Loma Larga | P.g. var. g. X P.l. | 19.89 JK |
| 4 | La Barretosa | P.g. var. g. X P.l. | 26.31 JK | 13 | Los Pajaritos | P.g. var. g. X P.l. | 19.89 JK |
| 18 | El Puente | P.g. var. g. X P.l. | 27.36 KL | 4 | La Barretosa | P.g. var. g. X P.l. | 19.90 JK |
| 19 | Rancho Gomas | P.g. var. g. X P.l. | 28.09 LM | 9 | Ejido El Álamo | P.g. var. g. X P.l. | 21.15 KL |
| 12 | Higueras | P.g. var. g. X P.l. | 28.27 LM | 18 | El Puente | P.g. var. g. X P.l. | 21.80 LM |
| 9 | Ejido El Álamo | P.g. var. g. X P.l. | 28.43 LM | 11 | Hacienda San Pedro | P.g. var. g. X P.l. | 22.39 LMN |
| 17 | El Resumidero | P.g. var. g. X P.l. | 28.48 LMN | 17 | El Resumidero | P.g. var. g. X P.l. | 22.78 LMN |
| 2 | Colorados de Arriba | P.g. var. g. X P.l. | 29.15 MNO | 2 | Colorados de Arriba | P.g. var. g. X P.l. | 23.27 MNO |
| 5 | Ej. Emiliano Zapata | P.g. var. g. X P.l. | 30.14 NOP | 19 | Rancho Gomas | P.g. var. g. X P.l. | 23.89 NO |
| 11 | Hacienda San Pedro | P.g. var. g. X P.l. | 30.66 OP | 5 | Ej. Emiliano Zapata | P.g. var. g. X P.l. | 24.72 O |
| 24 | El Nogal—Anáhuac | P.g. var. g. | 31.49 PQ | 6 | Ej. Puente del Río Salado | P.g. var. g. | 26.86 P |
| 23 | Ejido las Presas—Lampazos | P.g. var. g. | 31.49 PQ | 23 | Ejido las Presas | P.g. var. g. | 27.38 P |
| 6 | Ej. Puente del Río Salado | P.g. var. g. | 32.40 PQ | 24 | El Nogal | P.g. var. g. | 28.17 P |
| 25 | Regantes 26—Anáhuac | P.g. var. g. | 36.47 R | 25 | Regantes 26 | P.g. var. g. | 30.70 Q |
| Spacing of Middle Leaflets | Length/Width Ratio of Middle Leaflets | Length/Width Ratio of Upper Leaflets | ||||||
|---|---|---|---|---|---|---|---|---|
| Site | Locality | Mean | Site | Locality | Mean | Site | Locality | Mean |
| 31 | Km80 ** | 1.00 A * | 31 | Km80 ** | 3.45 A * | 31 | Los Ramones | 1.58 A * |
| 28 | San pedro de los Escobedos | 2.19 B | 21 | Rancho La bonanza | 5.30 B | 28 | Linares | 3.98 B |
| 21 | Rancho La bonanza | 2.24 B | 28 | San pedro de los Escobedos | 5.45 BC | 21 | General Terán | 4.03 BC |
| 22 | Rancho Nuevo | 2.72 BC | 30 | Ejido las Puentes | 5.73 BC | 30 | Aramberri | 4.46 BC |
| 30 | Ejido las Puentes | 3.33 C | 22 | Rancho Nuevo | 5.86 C | 22 | General Terán | 4.52 C |
| 20 | Km 80 ** | 4.70 D | 29 | San Ignacio de Texas | 7.07 D | 29 | Galeana | 5.49 D |
| 29 | San Ignacio de Texas | 4.71 D | 14 | Rancho el Recuerdo | 7.60 DE | 8 | Los Ramones | 5.75 DE |
| 27 | China | 4.90 D | 10 | Dulces Nombres | 7.65 E | 14 | General Terán | 5.82 DEF |
| 14 | Rancho el Recuerdo | 4.97 D | 8 | Los Ébanos | 7.89 E | 10 | Pesquería | 6.03 EF |
| 8 | Los Ébanos | 5.05 E | 20 | Km80 ** | 8.48 F | 20 | Los Ramones | 6.27 FG |
| 3 | El Llano | 6.77 E | 27 | China | 8.56 F | 27 | China | 6.65 G |
| 7 | Loma Larga | 6.79 E | 16 | El Bajío | 8.69 F | 16 | Marín | 7.18 H |
| 15 | Loma la Parada | 6.89 EF | 7 | Loma Larga | 9.53 G | 12 | Higueras | 7.74 I |
| 26 | Rancho la Ceja | 6.94 EF | 15 | Loma la Parada | 9.73 GH | 7 | General Treviño | 7.78 I |
| 10 | Dulces Nombres | 6.96 EF | 26 | Rancho la Ceja | 9.77 GH | 11 | General Zuazua | 7.78 I |
| 16 | El Bajío | 7.77 EFG | 13 | Los Pajaritos | 9.80 GH | 1 | Melchor Ocampo | 7.84 I |
| 13 | Los Pajaritos | 7.33 EFG | 11 | Hacienda San Pedro | 9.83 GH | 15 | Marín | 7.89 I |
| 4 | La Barretosa | 7.38 EFG | 1 | Plan del Orégano | 9.88 GH | 18 | Salinas Victoria | 7.90 I |
| 1 | Plan del Orégano | 7.46 EFGH | 18 | El Puente | 10.03 GHI | 26 | Los Aldama | 7.98 I |
| 12 | Higueras | 7.59 FGH | 9 | Ejido El Álamo | 10.15 HI | 3 | Los Ramones | 8.02 I |
| 2 | Colorados de Arriba | 7.90 GHI | 3 | El Llano | 10.22 HIJ | 9 | Vallecillo | 8.04 I |
| 18 | El Puente | 7.94 GHI | 12 | Higueras | 10.26 HIJ | 13 | Doctor González | 8.07 I |
| 9 | Ejido El Álamo | 7.97 GHI | 19 | Rancho Gomas | 10.52 IJK | 4 | Los Herreras | 8.10 J |
| 17 | El Resumidero | 8.14 GHI | 5 | Ej. Emiliano Zapata | 10.77 JKL | 2 | Vallecillo | 8.81 J |
| 5 | Ej. Emiliano Zapata | 8.37 HI | 4 | La Barretosa | 10.91 KLM | 5 | Parás | 8.83 J |
| 24 | El Nogal | 8.47 I | 24 | El Nogal | 10.97 KLM | 19 | Salinas Victoria | 8.88 JK |
| 11 | Hacienda San Pedro | 8.53 IJ | 17 | El Resumidero | 11.00 KLM | 17 | Salinas Victoria | 8.91 JK |
| 23 | Ejido las Presas | 8.58 IJ | 2 | Colorados de Arriba | 11.07 KLM | 23 | Lampazos | 9.38 KL |
| 19 | Rancho Gomas | 9.20 JK | 23 | Ejido las Presas | 11.14 LM | 6 | Anáhuac | 9.69 L |
| 6 | Ej. Puente del Río Salado | 9.37 K | 6 | Ej. Puente del Río Salado | 11.37 M | 24 | Lampazos | 9.76 L |
| 25 | Anáhuac | 9.54 K | 25 | Regantes 26-Anáhuac | 12.16 N | 25 | Regantes 26 - Anáhuac | 10.57 M |
| Contrast of Functions | Wilk’s Lambda | Chi-Squared | Degrees of Freedom | Sig. |
|---|---|---|---|---|
| 1 to 4 | 0.196 | 5105.635 | 20 | 0.000 |
| 2 to 4 | 0.895 | 345.854 | 12 | 0.000 |
| 3 to 4 | 0.975 | 79.802 | 6 | 0.000 |
| 4 | 0.997 | 10.851 | 2 | 0.000 |
| Taxa | Predicted Membership Group * | Total | |||||
|---|---|---|---|---|---|---|---|
| 1 | 2 | 3 | 4 | 5 | |||
| Count | 1 | 194 | 205 | 0 | 1 | 0 | 400 |
| 2 | 110 | 1371 | 0 | 123 | 0 | 1604 | |
| 3 | 0 | 2 | 422 | 30 | 0 | 454 | |
| 4 | 0 | 145 | 45 | 390 | 0 | 580 | |
| 5 | 0 | 0 | 3 | 0 | 97 | 100 | |
| % | 1 | 48.5 | 51.3 | 0.0 | 0.3 | 0.0 | 100 |
| 2 | 6.9 | 85.5 | 0.0 | 7.7 | 0.0 | 100 | |
| 3 | 0.0 | 0.4 | 93.0 | 6.6 | 0.0 | 100 | |
| 4 | 0.0 | 25.0 | 7.8 | 67.2 | 0.0 | 100 | |
| 5 | 0.0 | 0.0 | 3.0 | 0.0 | 97.0 | 100 | |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Foroughbakhch Pournavab, R.; Ngangyo Heya, M.; Castillo Gonzalez, E.A.; Rocha Estrada, A.; Salas Cruz, L.R.; Alvarado Vázquez, M.A. Leaf Architecture in the Morphological Diversity of the Genus Prosopis in the Semi-Desert Area of Northeastern Mexico. Diversity 2024, 16, 351. https://doi.org/10.3390/d16060351
Foroughbakhch Pournavab R, Ngangyo Heya M, Castillo Gonzalez EA, Rocha Estrada A, Salas Cruz LR, Alvarado Vázquez MA. Leaf Architecture in the Morphological Diversity of the Genus Prosopis in the Semi-Desert Area of Northeastern Mexico. Diversity. 2024; 16(6):351. https://doi.org/10.3390/d16060351
Chicago/Turabian StyleForoughbakhch Pournavab, Rahim, Maginot Ngangyo Heya, Emmanuel Adan Castillo Gonzalez, Alejandra Rocha Estrada, Lidia Rosaura Salas Cruz, and Marco Antonio Alvarado Vázquez. 2024. "Leaf Architecture in the Morphological Diversity of the Genus Prosopis in the Semi-Desert Area of Northeastern Mexico" Diversity 16, no. 6: 351. https://doi.org/10.3390/d16060351
APA StyleForoughbakhch Pournavab, R., Ngangyo Heya, M., Castillo Gonzalez, E. A., Rocha Estrada, A., Salas Cruz, L. R., & Alvarado Vázquez, M. A. (2024). Leaf Architecture in the Morphological Diversity of the Genus Prosopis in the Semi-Desert Area of Northeastern Mexico. Diversity, 16(6), 351. https://doi.org/10.3390/d16060351