
The Effects of Infectious Diseases on the Consequences of Interspecific Competition in Grassland Communities

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
1. Introduction
2. Materials and Methods
2.1. Spatial Competition Model without Infection
2.2. Modeling Disease Infection of Species Competition
3. Results
3.1. Stability Analysis
- (1)
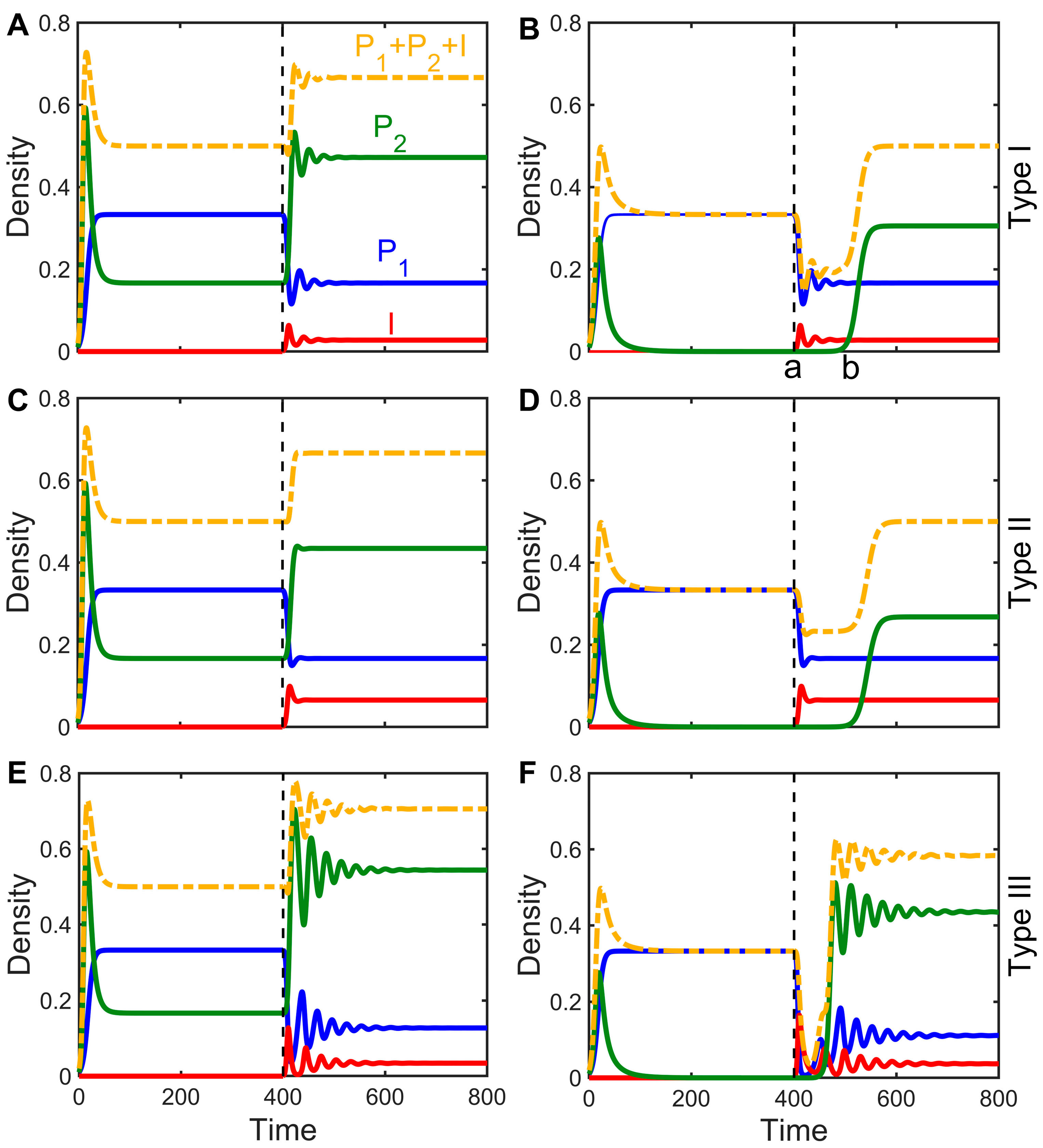
- The disease fails to spread and eventually disappears, at which point it is possible that the inferior competitor becomes extinct and that the superior competitor reaches its environmental carrying capacity of it is also possible that the inferior species successfully invades, so the superior and inferior species eventually coexist stably at a density of .
- (2)
- The disease successfully invades the host and eventually becomes endemic, at which point the inferior competitor may become competitively excluded and eventually bec Please confirm if the bold is unnecessary and can be removed. The following highlights are the same. ome extinct, or it may stably coexist with the susceptible and infected individuals at a density of (the steady-state values are different for each equation; see the supporting material for details).
- (3)
- Only Equation (4) has an extinction risk for the susceptible dominant species: When all hosts are infected, their numbers are close to an environmental capacity of , and the inferior competitor becomes extinct. In contrast, in Equations (2) and (3), the invasion of the pathogen does not directly lead to host extinction, and the equilibrium value for the susceptible individuals, , is always greater than zero.
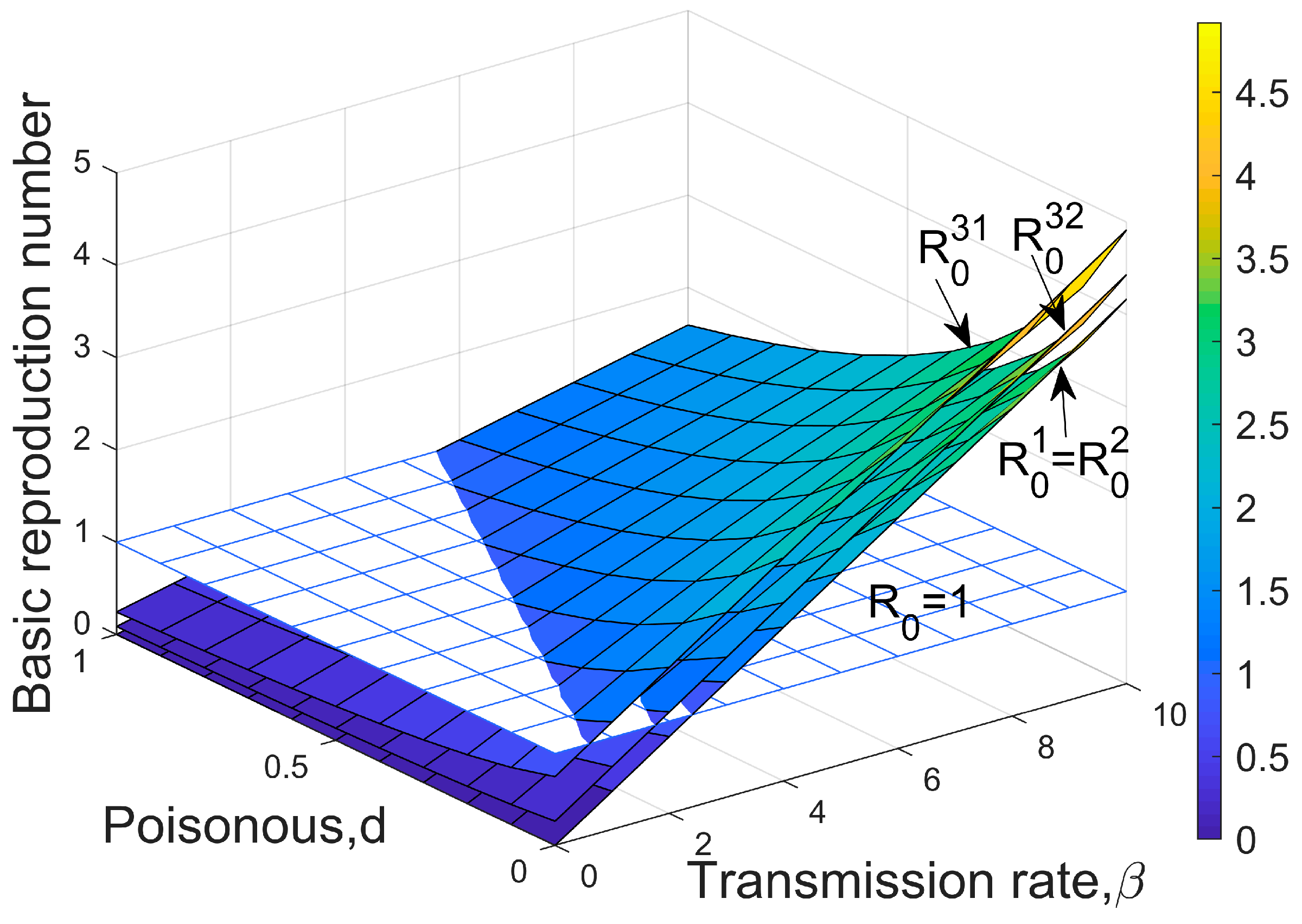
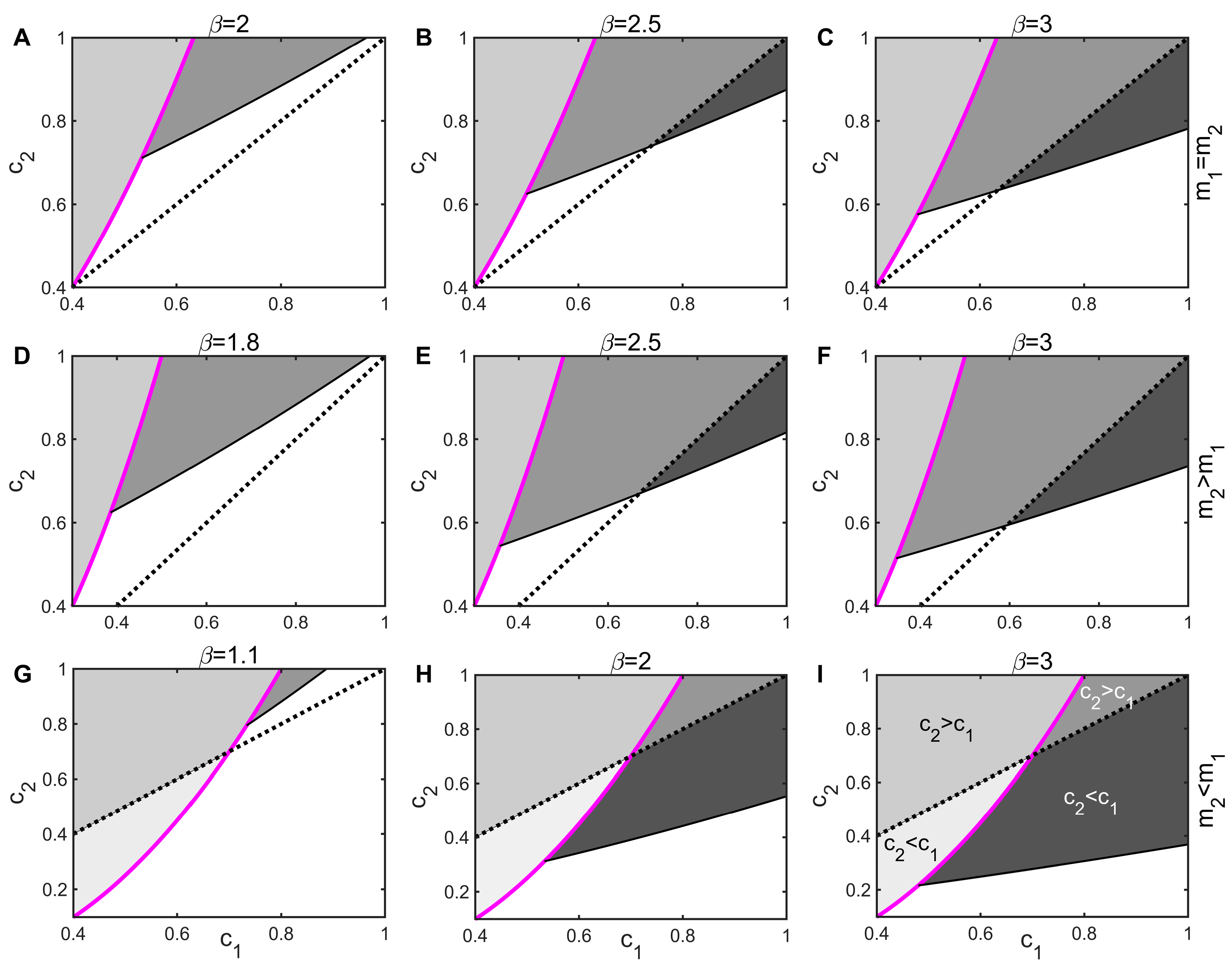
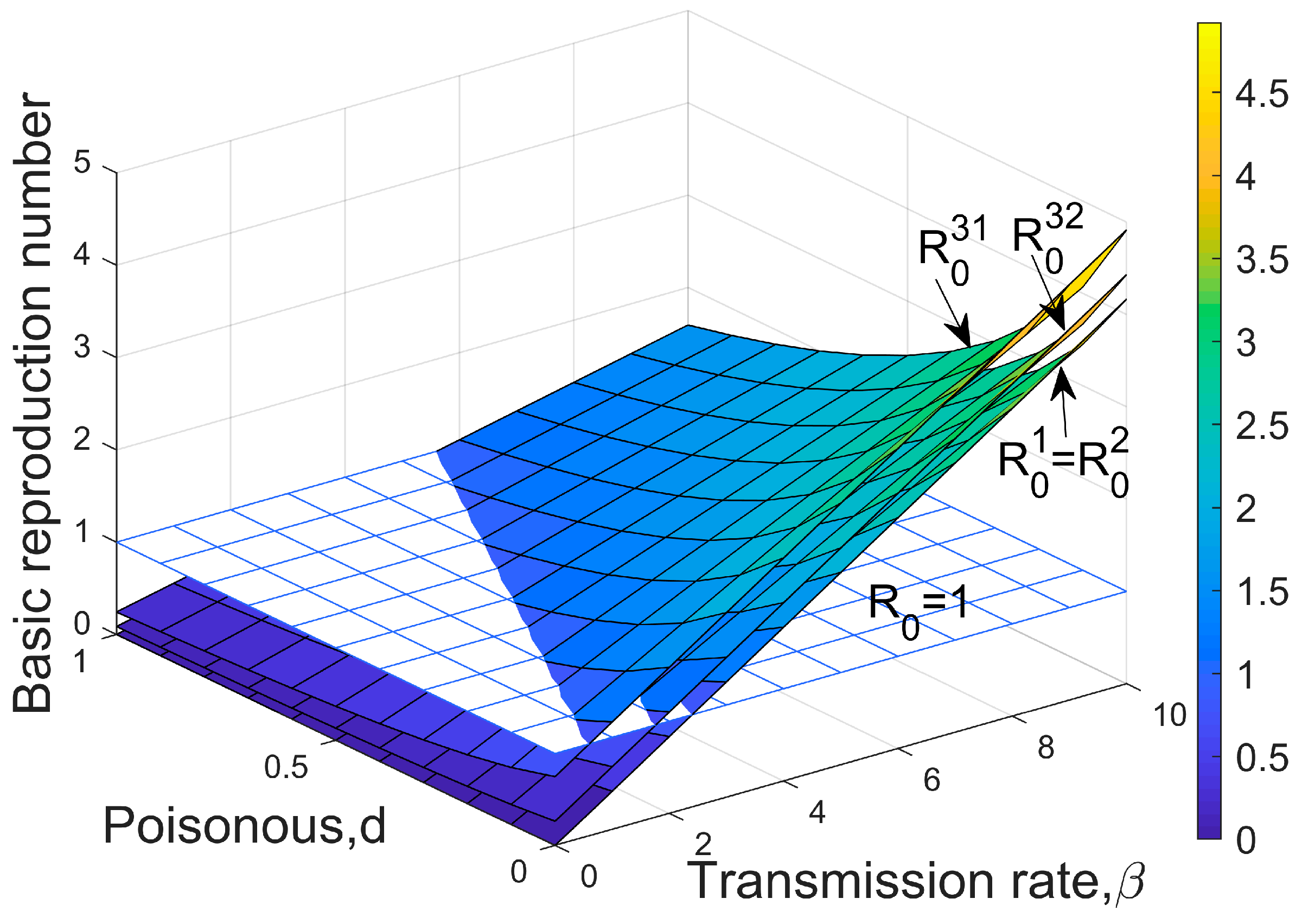
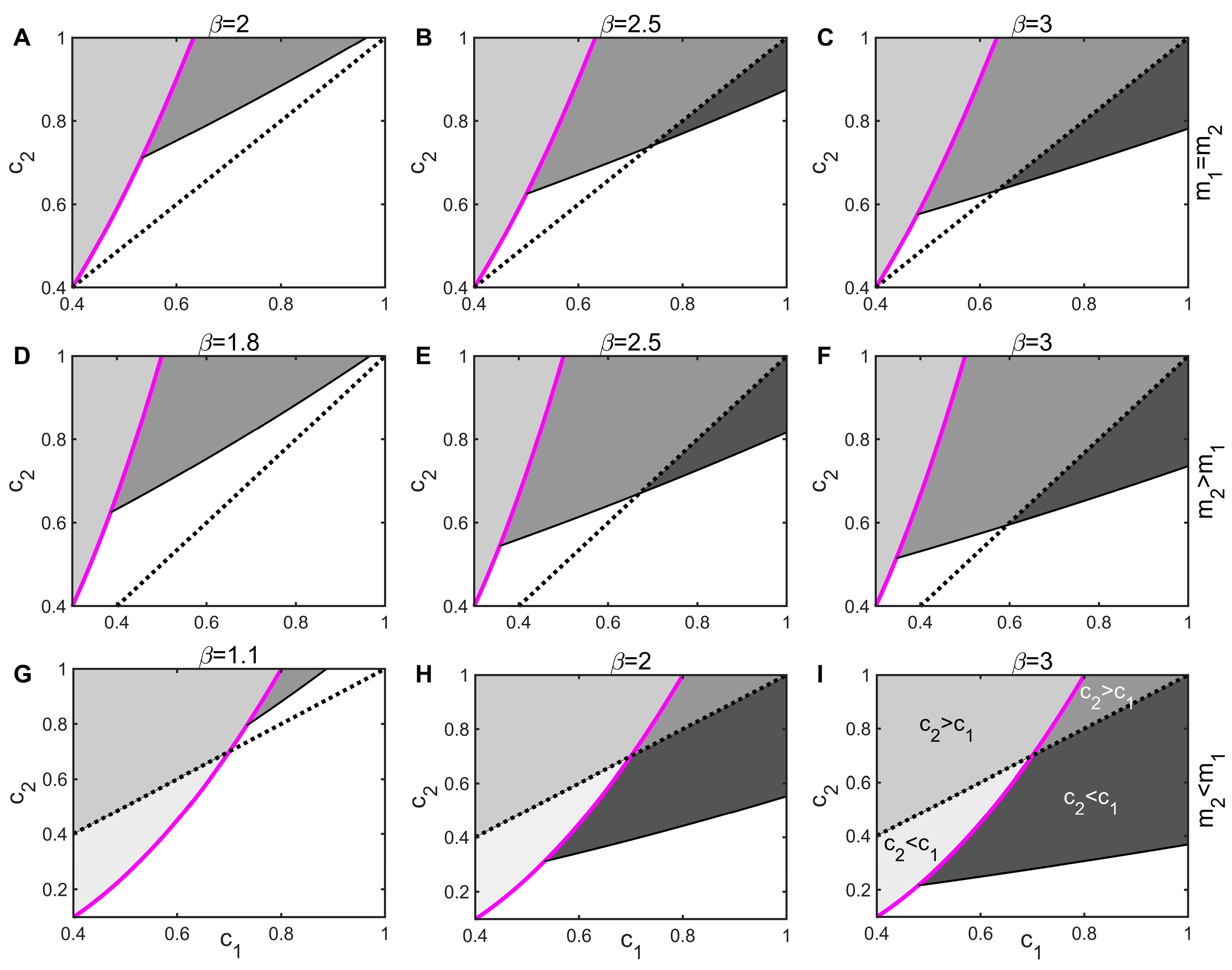
3.2. Threshold for Disease Invasion
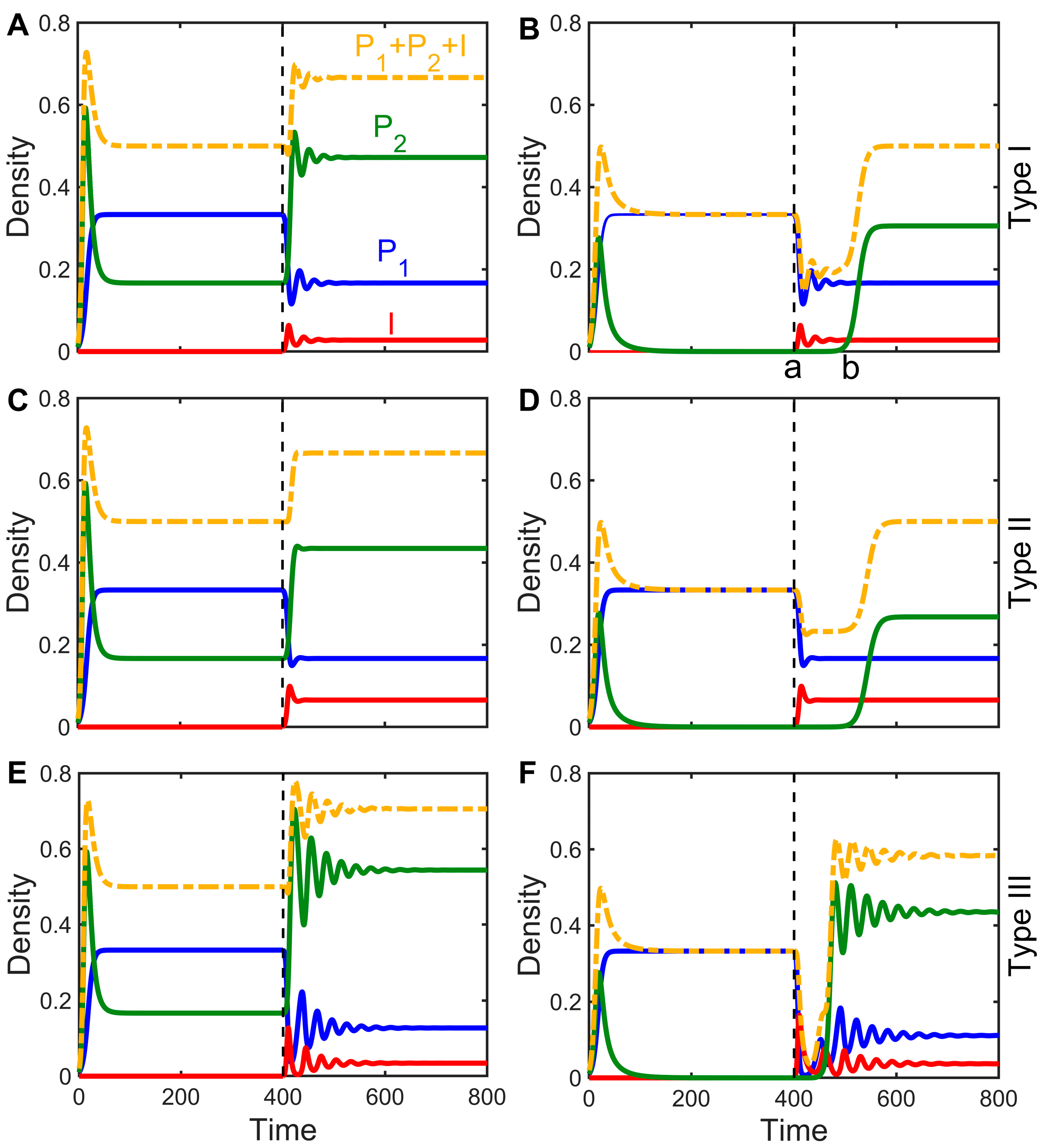
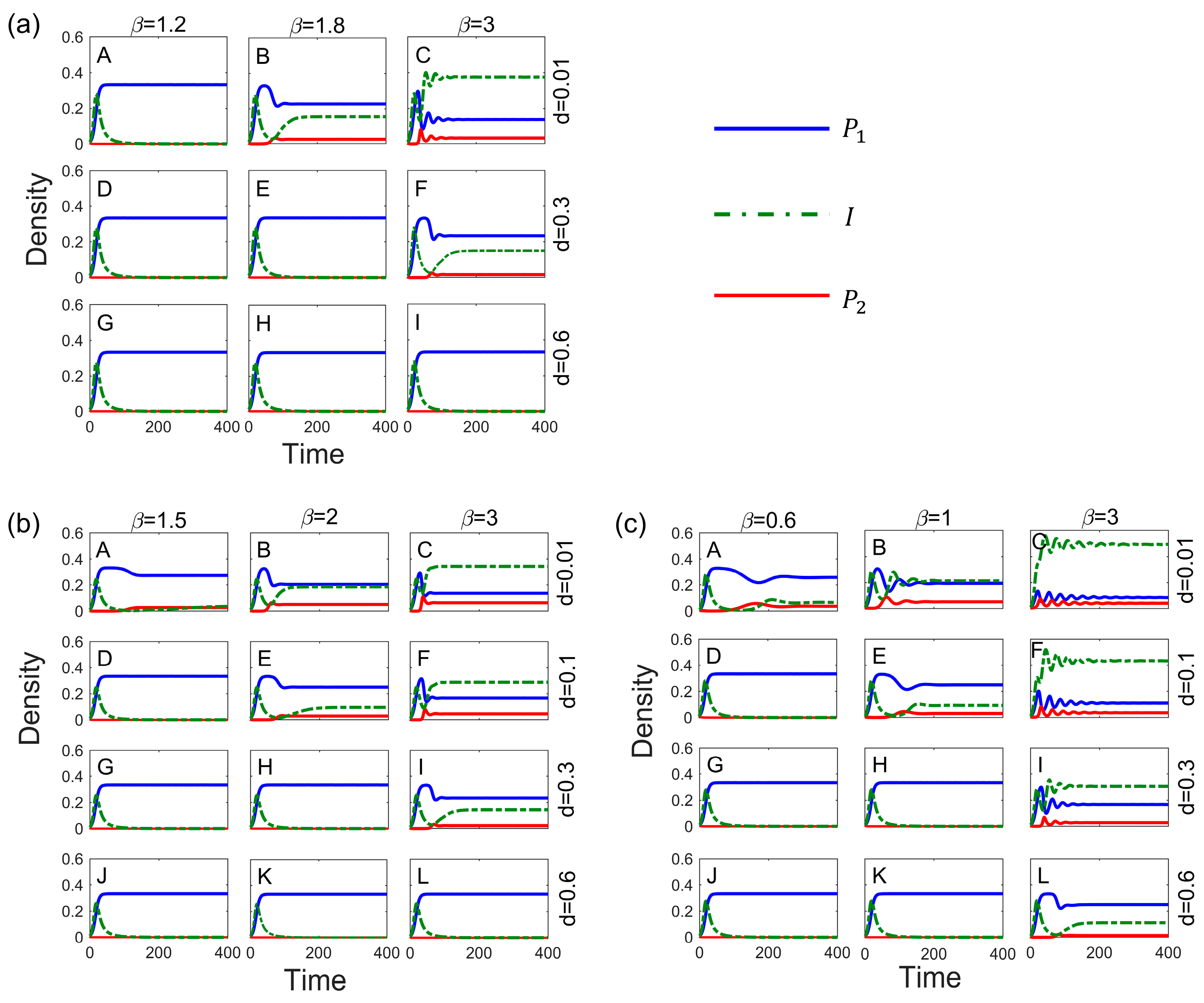
3.3. Impact of Infectious Diseases on Competitive Dynamics
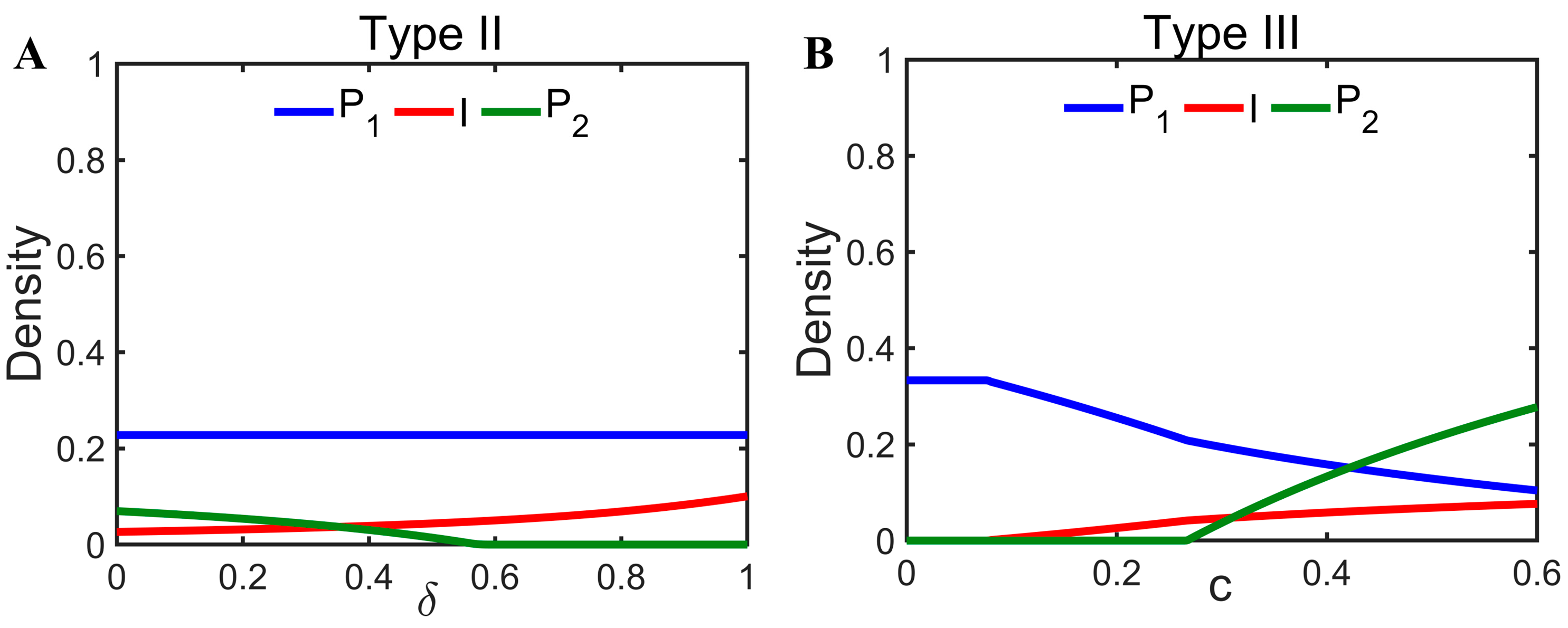
3.4. Mechanisms by Which Infectious Diseases Promote Coexistence
4. Discussion
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Appendix A
- (1)
- Equilibrium always exist and unstable, which means the dominant species can always invade the weaker competing population;
- (2)
- Equilibrium always exists and locally stable when , which represents the weaker species is rejected, only the dominant competitor continues to survive and reaches its environmental capacity ;
- (3)
- The disease-free equilibrium exists when , and it is locally stable if , which means the dominant and weaker species coexist stably, and the disease is not yet able to invade;
- (4)
- Equilibrium exists when , and is locally stable if , which represents the host is infected in a certain proportion, the disease persists for a long time, and the weaker species are rejected, eventually dying out.
- (5)
- When , the endemic disease equilibrium exists and is globally stable. It means that the disease becomes endemic at a certain level, while the two competing species coexist in a stable way and the equilibrium density does not change with the initial value of the population size.
- (1)
- Equilibrium , always exist and unstable;
- (2)
- always exists and locally stable when , ;
- (3)
- exists when , and stable if ;
- (4)
- Equilibrium exists when , and stable if , which means all hosts are infected and their numbers are close to the environmental capacity of , the inferior competitor becomes extinct.
- (5)
- exists when , and unstable if (the process see Supplementary Materials part III). where A0 = β(c1 − c + β), A1 = c1 (m1 + d) − β(c − m1 − d) − cm1, A2 = −c1 (m1 + d) + β(c1 − m1) + cm1 ;
- (6)
- exists if , , . And further, when , the equilibrium is locally asymptotically stable according to the Routh-Huriwitz criterion, where, , , , , , . The , and process see Supplementary Materials part III.
References
- Dixon, A.P.; Faber-Langendoen, D.; Josse, C.; Morrison, J.; Loucks, C.J. Distribution mapping of world grassland types. J. Biogeogr. 2014, 41, 2003–2019. [Google Scholar] [CrossRef]
- Chen, S.; Wang, W.; Xu, W.; Wang, Y.; Wan, H.; Chen, D.; Tang, Z.; Tang, X.; Zhou, G.; Xie, Z.; et al. Plant diversity enhances productivity and soil carbon storage. Proc. Natl. Acad. Sci. USA 2018, 115, 4027–4032. [Google Scholar] [CrossRef]
- Bai, Y.; Cotrufo, M.F. Grassland soil carbon sequestration: Current understanding, challenges, and solutions. Science 2022, 377, 603–608. [Google Scholar] [CrossRef]
- Bengtsson, J.; Bullock, J.M.; Egoh, B.; Everson, C.; Everson, T.; O’connor, T.; O’farrell, P.J.; Smith, H.G.; Lindborg, R. Grasslands—More important for ecosystem services than you might think. Ecosphere 2019, 10, e02582. [Google Scholar] [CrossRef]
- Zhao, Y.; Liu, Z.; Wu, J. Grassland ecosystem services: A systematic review of research advances and future directions. Landsc. Ecol. 2020, 35, 793–814. [Google Scholar] [CrossRef]
- Morens, D.M.; Folkers, G.K.; Fauci, A.S. The challenge of emerging and re-emerging infectious diseases. Nature 2004, 430, 242–249. [Google Scholar] [CrossRef]
- Pimm, S.L.; Jenkins, C.N.; Abell, R.; Brooks, T.M.; Gittleman, J.L.; Joppa, L.N.; Raven, P.H.; Roberts, C.M.; Sexton, J.O. The biodiversity of species and their rates of extinction, distribution, and protection. Science 2014, 344, 1246752. [Google Scholar] [CrossRef]
- Rúa, M.A.; McCulley, R.L.; Mitchell, C.E. Climate drivers, host identity and fungal endophyte infection determine virus prevalence in a grassland ecosystem. J. Ecol. 2014, 102, 690–699. [Google Scholar] [CrossRef]
- Jeger, M.J.; Salama, N.K.G.; Shaw, M.W.; Van Den Berg, F.; Van Den Bosch, F. Effects of plant pathogens on population dynamics and community composition in grassland ecosystems: Two case studies. Eur. J. Plant Pathol. 2014, 138, 513–527. [Google Scholar] [CrossRef]
- Semchenko, M.; Leff, J.W.; Lozano, Y.M.; Saar, S.; Davison, J.; Wilkinson, A.; Jackson, B.G.; Pritchard, W.J.; De Long, J.R.; Oakley, S.; et al. Fungal diversity regulates plant-soil feedbacks in temperate grassland. Sci. Adv. 2018, 11, eaau4578. [Google Scholar] [CrossRef]
- Prevéy, J.S.; Seastedt, T.R. Increased winter precipitation benefits the native plant pathogen Ustilago bullata that infects an invasive grass. Biol. Invasions 2015, 17, 3041–3047. [Google Scholar] [CrossRef]
- Mordecai, E.A. Pathogen impacts on plant communities: Unifying theory, concepts, and empirical work. Ecol. Monogr. 2011, 81, 429–441. [Google Scholar] [CrossRef]
- Baums, I.B.; Baker, A.C.; Davies, S.W.; Grottoli, A.G.; Kenkel, C.D.; Kitchen, S.A.; Kuffner, I.B.; LaJeunesse, T.C.; Matz, M.V.; Miller, M.W.; et al. Considerations for maximizing the adaptive potential of restored coral populations in the western Atlantic. Ecol. Appl. 2019, 29, e01978. [Google Scholar] [CrossRef] [PubMed]
- Roughgarden, J.; Gilbert, S.F.; Rosenberg, E.; Zilber-Rosenberg, I.; Lloyd, E.A. Holobionts as units of selection and a model of their population dynamics and evolution. Biol. Theor. 2018, 13, 44–65. [Google Scholar] [CrossRef]
- Barrett, L.G.; Kniskern, J.M.; Bodenhausen, N.; Zhang, W.; Bergelson, J. Continua of specificity and virulence in plant host–pathogen interactions: Causes and consequences. New Phytol. 2009, 183, 513–529. [Google Scholar] [CrossRef] [PubMed]
- Bever, J.D.; Mangan, S.A.; Alexander, H.M. Maintenance of plant species diversity by pathogens. Annu. Rev. Ecol. Evol. Syst. 2015, 46, 305–325. [Google Scholar] [CrossRef]
- Jarosz, A.M.; Davelos, A.L. Effects of disease in wild plant populations and the evolution of pathogen aggressiveness. New Phytol. 1995, 129, 371–387. [Google Scholar] [CrossRef]
- van Dijk, L.J.; Ehrlén, J.; Tack, A.J. Direct and insect-mediated effects of pathogens on plant growth and fitness. J. Ecol. 2021, 109, 2769–2779. [Google Scholar] [CrossRef]
- Lin, Z.; Liu, H. How species diversity responds to different kinds of human-caused habitat destruction. Ecol. Res. 2006, 21, 100–106. [Google Scholar] [CrossRef]
- Hirzel, A.H.; Le, L.G. Habitat suitability modelling and niche theory. J. Appl. Ecol. 2008, 45, 1372–1381. [Google Scholar] [CrossRef]
- Levine, J.M.; Bascompte, J.; Adler, P.B.; Allesina, S. Beyond pairwise mechanisms of species coexistence in complex communities. Nature 2017, 546, 56–64. [Google Scholar] [CrossRef] [PubMed]
- Buck, J.C. Indirect effects explain the role of parasites in ecosystems. Trends Parasitol. 2019, 35, 835–847. [Google Scholar] [CrossRef] [PubMed]
- Chesson, P. Mechanisms of maintenance of species diversity. Annu. Rev. Ecol. Syst. 2000, 31, 343–366. [Google Scholar] [CrossRef]
- Chen, T.; Nan, Z.B. Progress in effects of phytopathogens on plant communities: A review. Acta Ecol. Sin. 2015, 35, 177–183. [Google Scholar] [CrossRef]
- Dai, D.; Xing, H.; Yang, J.R.; Liu, Y.J.; Cai, H.M.; Liu, Y. Advances in mechanisms of rare species maintenance and plant-soil feedback in plant communities. Biodivers. Sci. 2021, 29, 1687–1699. [Google Scholar] [CrossRef]
- Souvik, B.; Maia, M. A Predator-Prey-Disease Model with Immune Response in Infected Prey. J. Math Anal. Appl. 2014, 411, 297–313. [Google Scholar]
- Herbert, W. A Predator-Prey Model with Infected Prey. Theor. Popul. Biol. 2004, 66, 259–268. [Google Scholar]
- Mainul, H. A Predator-Prey Model with Disease in the Predator Species Only. Nonlin. Anal.-Real World Appl. 2010, 11, 2224–2236. [Google Scholar]
- Xuin, G. A Predator-Prey Model with Diseases in both Prey and Predator. Physica A 2013, 392, 5898–5906. [Google Scholar]
- Tilman, D. Competition and Biodiversity in Spatially Structured Habitats. Ecology 1994, 75, 2–16. [Google Scholar] [CrossRef]
- Libál, A.; Forgács, P.; Néda, Á.; Reichhardt, C.; Hengartner, N.; Reichhardt, C.J.O. Transition from susceptible-infected to susceptible-infected-recovered dynamics in a susceptible-cleric-zombie-recovered active matter model. Phys. Rev. E 2023, 107, 024604. [Google Scholar] [CrossRef] [PubMed]
- Baloda, S.; Sehrawat, S.K.; Yadav, B.S.; Ahlawat, V.P.; Singh, S. Present status of ber production and future thrusts in India—A review. Agric. Rev. 2012, 33, 256–264. [Google Scholar]
- Barbetti, M.J. The expression of resistance in subterranean clover (Trifolium subterraneum) to races 1 and 2 of Kabatiella caulivora is affected by inoculum pressure but not by combinations of the two races. Australas. Plant Pathol. 2007, 36, 318–324. [Google Scholar] [CrossRef]
- Veltman, B.; Harpaz, D.; Melamed, S.; Tietel, Z.; Tsror, L.; Eltzov, E. Whole-cell bacterial biosensor for volatile detection from Pectobacterium-infected potatoes enables early identification of potato tuber soft rot disease. Talanta 2022, 247, 123545. [Google Scholar] [CrossRef]
- Liu, M.; Shoukouhi, P.; Bisson, K.R.; Wyka, S.A.; Broders, K.D.; Menzies, J.G. Sympatric divergence of the ergot fungus, Claviceps purpurea, populations infecting agricultural and nonagricultural grasses in North America. Ecol. Evol. 2021, 11, 273–293. [Google Scholar] [CrossRef] [PubMed]
- Lind, E.M.; Borer, E.; Seabloom, E.; Adler, P.E.; Bakker, J.B. Life-history constraints in grassland plant species: A growth-defence trade-off is the norm. Ecol. Lett. 2013, 16, 513–521. [Google Scholar] [CrossRef] [PubMed]
- Liu, X.; Chen, L.; Zhou, S. The relationship between biodiversity and infectious disease: Progress, challenge and perspective. Biodivers. Sci. 2020, 28, 1376. [Google Scholar] [CrossRef]
- Keeling, M.J.; Rohani, P. Modeling Infectious Diseases in Humans and Animals; Princeton University Press: Princeton, NJ, USA, 2008. [Google Scholar]
- Hatcher, M.J.; Dick, J.T.; Dunn, A.M. Diverse effects of parasites in ecosystems: Linking interdependent processes. Front. Ecol. Environ. 2012, 10, 186–194. [Google Scholar] [CrossRef] [PubMed]
- Hudson, P.J.; Dobson, A.P.; Lafferty, K.D. Is a healthy ecosystem one that is rich in parasites? Trends Ecol. Evolut. 2006, 21, 381–385. [Google Scholar] [CrossRef]
- Wood, C.L.; Johnson, P.T. A world without parasites: Exploring the hidden ecology of infection. Front Ecol. Environ. 2015, 13, 425–434. [Google Scholar] [CrossRef]
- Stringer, A.P.; Linklater, W. Everything in moderation: Principles of parasite control for wildlife conservation. BioScience 2014, 64, 932–937. [Google Scholar] [CrossRef]
- Lafferty, K.D.; Allesina, S.; Arim, M.; Briggs, C.J.; Leo, G.D.; Dobson, A.P.; Dunne, J.A.; Johnson, P.T.J.; Kuris, A.M.; Marcogliese, D.J.; et al. Parasites in food webs: The ultimate missing links. Ecol. Lett. 2008, 11, 533–546. [Google Scholar] [CrossRef] [PubMed]
- Tompkins, D.M.; White, A.R.; Boots, M. Ecological replacement of native red squirrels by invasive greys driven by disease. Ecol. Lett. 2003, 6, 189–196. [Google Scholar] [CrossRef]
- Bowers, R.G.; Turner, J. Community structure and the interplay between interspecific infection and competition. J. Theor. Biol. 1997, 187, 95–109. [Google Scholar] [CrossRef]
- Han, L.; Pugliese, A. Epidemics in two competing species. Nonlin. Anal.-Real World Appl. 2009, 10, 723–744. [Google Scholar] [CrossRef]
- Han, L.; Ma, Z.; Shi, T. An SIRS epidemic model of two competitive species. Math. Comput. Model. 2003, 37, 87–108. [Google Scholar] [CrossRef]
- Venturino, E. The effects of diseases on competing species. Math. Biosci. 2001, 174, 111–131. [Google Scholar] [CrossRef] [PubMed]
- Van den Driessche, P.; Zeeman, M.L. Disease Induced Oscillations between Two Competing Species. SIAM J. Appl. Dyn. Syst. 2004, 3, 601–619. [Google Scholar] [CrossRef]
- Fang, D.X.; Wan, X.; Mao, W.Q.; Zhang, F. Effect of pathogen infection on resource competition between species: A study based on resource competition theory. Acta Ecol. Sin. 2023, 43, 7109–7117. [Google Scholar]
- Bravo de la Parra, R.; Marvá, M.; Sánchez, E.; Sanz, L. Discrete models of disease and competition. Discret. Dyn. Nat. Soc. 2017, 2017, 5310837. [Google Scholar] [CrossRef]
- Bravo de la Parra, R.; Marvá, M.; Sánchez, E.; Sanz, L. A discrete model of competing species sharing a parasite. Discret. Dyn. Nat. Soc. 2017, 5, 565–569. [Google Scholar] [CrossRef]
- Torchin, M.E.; Lafferty, K.D.; Dobson, A.P.; McKenzie, V.J.; Kuris, A.M. Introduced species and their missing parasites. Nature 2003, 421, 628–630. [Google Scholar] [CrossRef]
- Assaf, T.A.; Beyschlag, W.; Isselstein, J. The relationship between plant diversity and productivity in natural and in managed grasslands. Appl. Ecol. Environ. Res. 2011, 9, 157–166. [Google Scholar] [CrossRef]
- Loreau, M.; Hector, A. Partitioning selection and complementarity in biodiversity experiments. Nature 2001, 412, 72–76. [Google Scholar] [CrossRef] [PubMed]
- Tilman, D.; Reich, P.B.; Knops, J.; Wedin, D.; Mielke, T.; Lehman, C. Diversity and productivity in a long-term grassland experiment. Science 2001, 294, 843–845. [Google Scholar] [CrossRef] [PubMed]
- Mahaut, L.; Fort, F.; Violle, C.; Freschet, G.T. Multiple facets of diversity effects on plant productivity: Species richness, functional diversity, species identity and intraspecific competition. Funct. Ecol. 2020, 34, 287–298. [Google Scholar] [CrossRef]
- Vera, M.C.; Marvá, M.; García-Garrido, V.J.; Escalante, R. The Beddington–DeAngelis Competitive Response: Intra-Species Interference Enhances Coexistence in Species Competition. Mathematics 2024, 12, 562. [Google Scholar] [CrossRef]
- Carpenter, S.R.; Kitchell, J.F.; Hodgson, J.R. Cascading trophic interactions and lake productivity. BioScience 1985, 35, 634–639. [Google Scholar] [CrossRef]
- Zhang, F.; Cao, M.; Shi, L.; Wang, R.W. Disease-Induced Cooperation Mitigates Populations Against Decline: The Cascade Effect of Cooperation Evolution. Front. Ecol. Evolut. 2022, 10, 758659. [Google Scholar] [CrossRef]


Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Chen, Y.; Liu, Y.; Liu, X.; Zhang, Z.; Zhang, F. The Effects of Infectious Diseases on the Consequences of Interspecific Competition in Grassland Communities. Diversity 2024, 16, 231. https://doi.org/10.3390/d16040231
Chen Y, Liu Y, Liu X, Zhang Z, Zhang F. The Effects of Infectious Diseases on the Consequences of Interspecific Competition in Grassland Communities. Diversity. 2024; 16(4):231. https://doi.org/10.3390/d16040231
Chicago/Turabian StyleChen, Yanli, Yanping Liu, Xiaoni Liu, Zhengzhong Zhang, and Feng Zhang. 2024. "The Effects of Infectious Diseases on the Consequences of Interspecific Competition in Grassland Communities" Diversity 16, no. 4: 231. https://doi.org/10.3390/d16040231
APA StyleChen, Y., Liu, Y., Liu, X., Zhang, Z., & Zhang, F. (2024). The Effects of Infectious Diseases on the Consequences of Interspecific Competition in Grassland Communities. Diversity, 16(4), 231. https://doi.org/10.3390/d16040231