
Massive Bird Nest Losses: A Neglected Threat for Passerine Birds in Atlantic Forest Fragments from the Pernambuco Endemism Center

 ,
,  and
and
Abstract
1. Introduction
2. Materials and Methods
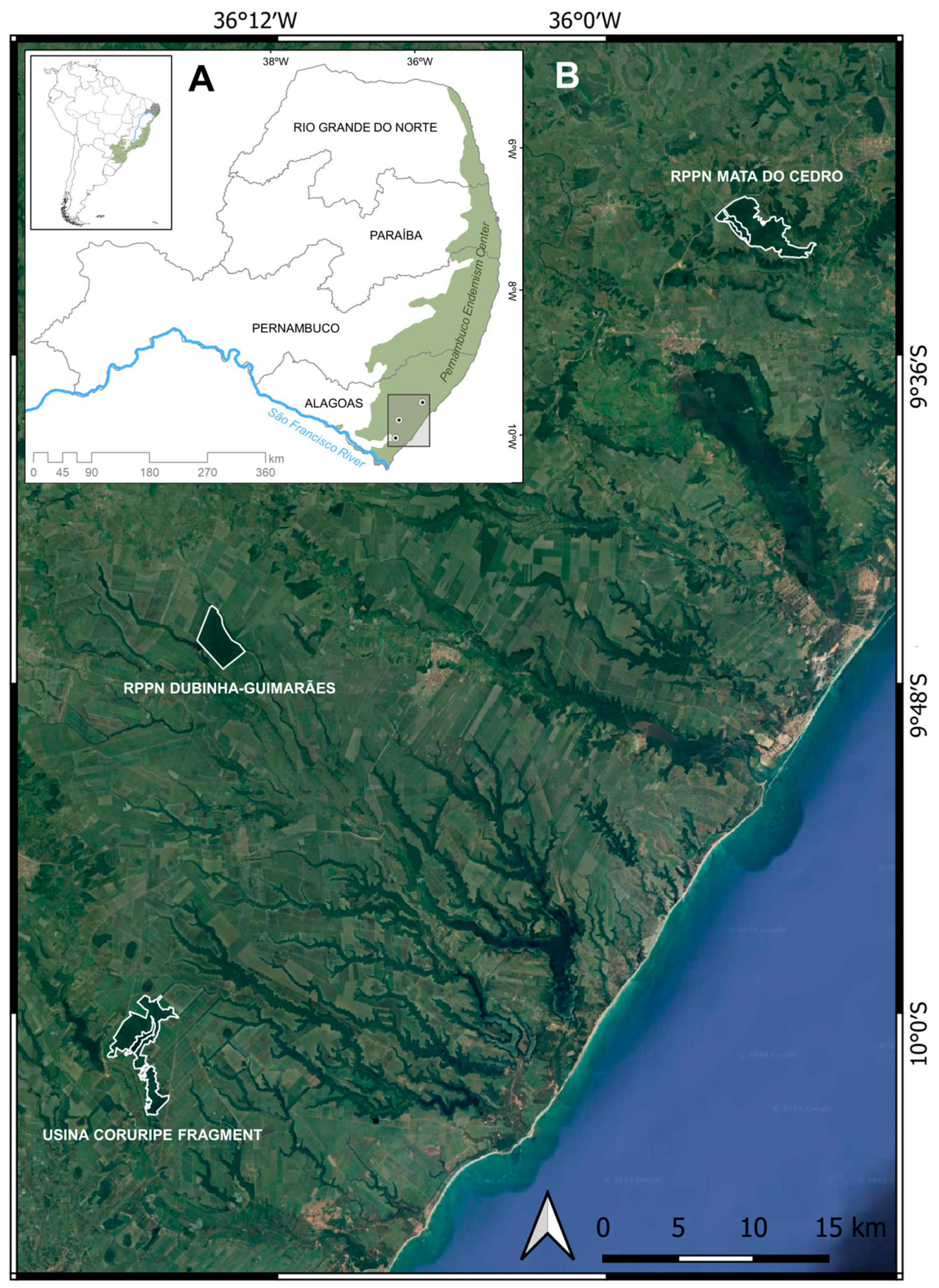
2.1. Study Areas
2.2. Nest Searches and Monitoring
2.3. Nest Survival Estimates
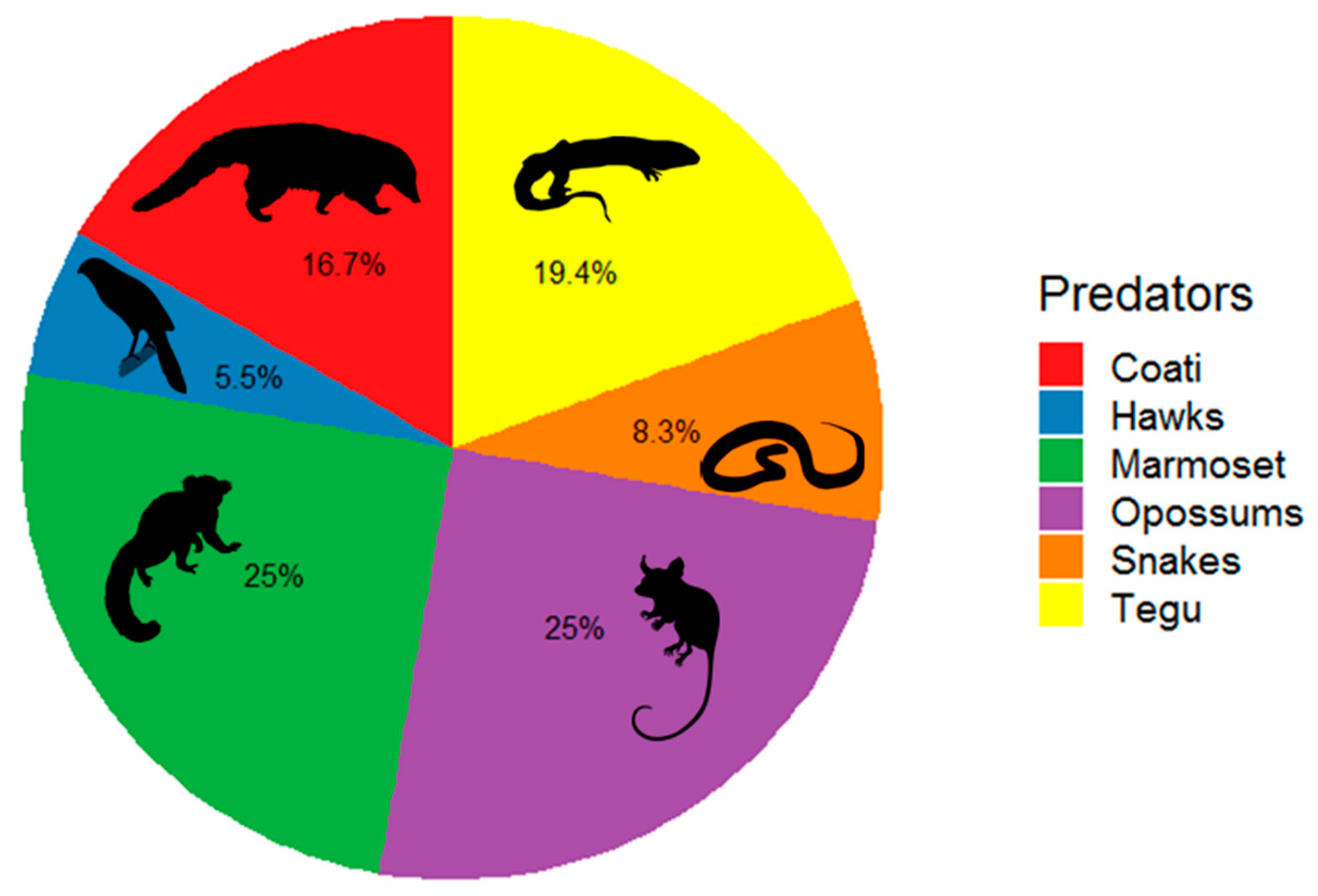
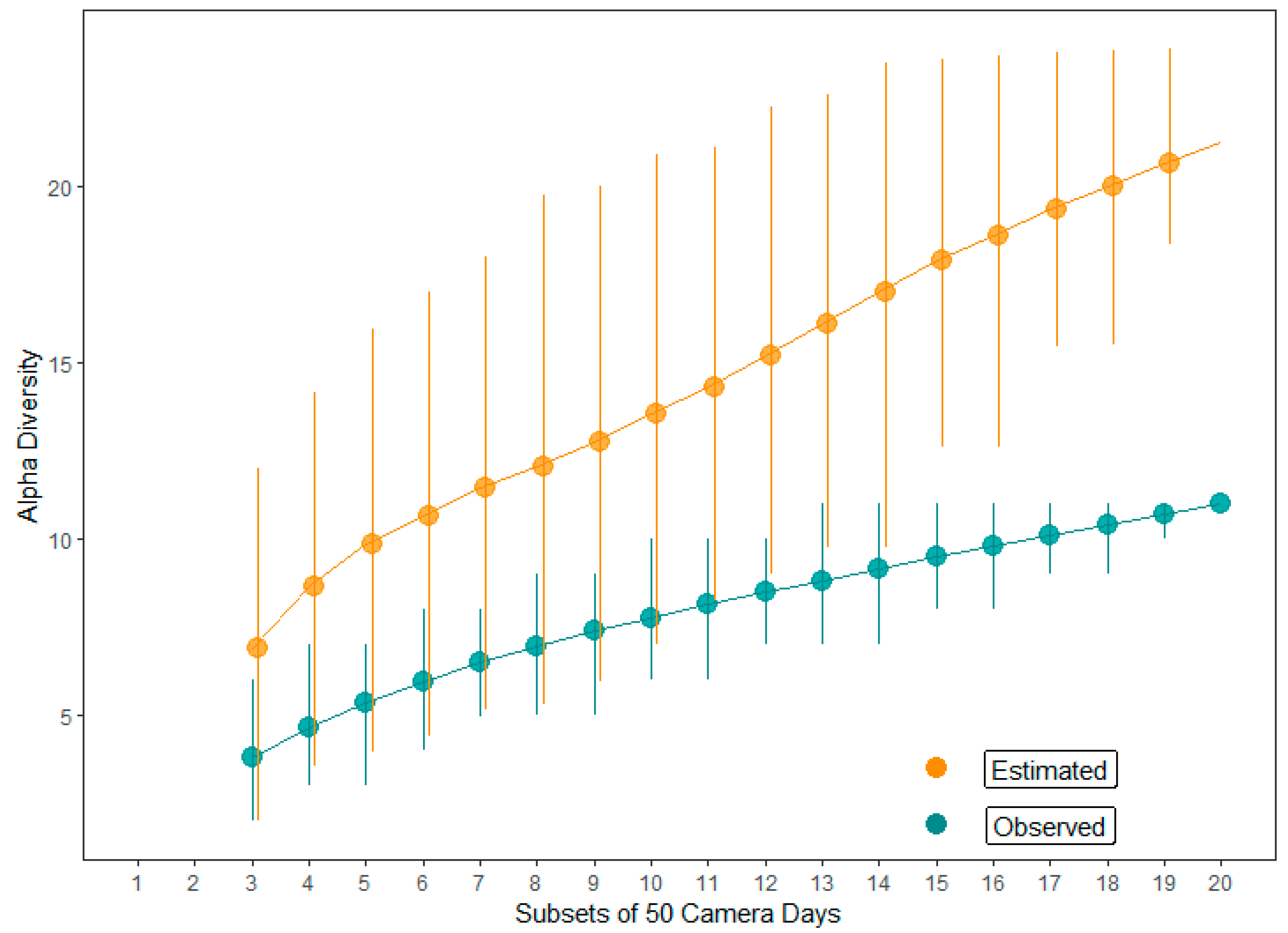
2.4. Nest Predator’s Identification and Alpha Diversity
3. Results
4. Discussion
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Crooks, K.R.; Burdett, C.L.; Theobald, D.M.; King, S.R.; Di Marco, M.; Rondinini, C.; Boitani, L. Quantification of habitat fragmentation reveals extinction risk in terrestrial mammals. Proc. Natl. Acad. Sci. USA 2017, 114, 7635–7640. [Google Scholar] [CrossRef]
- Liu, J.; Wilson, M.; Hu, G.; Liu, J.; Wu, J.; Yu, M. How does habitat fragmentation affect the biodiversity and ecosystem functioning relationship? Landsc. Ecol. 2018, 33, 341–352. [Google Scholar] [CrossRef]
- Costa-Araújo, R.; Regolin, A.L.; Martello, F.; Souza-Alves, J.P.; Hrbek, T.; Ribeiro, M.C. Occurrence and conservation of the vulnerable titi monkey Callicebus melanochir in fragmented landscapes of the Atlantic Forest hotspot. Oryx 2021, 55, 916–923. [Google Scholar] [CrossRef]
- Schlaepfer, D.R.; Braschler, B.; Rusterholz, H.P.; Baur, B. Genetic effects of anthropogenic habitat fragmentation on remnant animal and plant populations: A meta-analysis. Ecosphere 2018, 9, e02488. [Google Scholar] [CrossRef]
- Slater, H.D.; Gillingham, P.K.; Pratt, V.; Eaton, B.; Fletcher, S.; Abdullah, A.; Supriadi; Korstjens, A.H. Living on the edge: Forest edge effects on microclimate and terrestrial mammal activity in disturbed lowland forest in Sumatra, Indonesia. Oryx 2024, 58, 228–239. [Google Scholar] [CrossRef]
- Didham, R.K.; Tylianakis, J.M.; Gemmell, N.J.; Rand, T.A.; Ewers, R.M. Interactive effects of habitat modification and species invasion on native species decline. Trends Ecol. Evol. 2007, 22, 489–496. [Google Scholar] [CrossRef]
- Krosby, M.; Tewksbury, J.; Haddad, N.M.; Hoekstra, J. Ecological connectivity for a changing climate. Conserv. Biol. 2010, 24, 1686–1689. [Google Scholar] [CrossRef] [PubMed]
- Acharya, K.P.; Paudel, P.K.; Jnawali, S.R.; Neupane, P.R.; Köhl, M. Can forest fragmentation and configuration work as indicators of human–wildlife conflict? Evidences from human death and injury by wildlife attacks in Nepal. Ecol. Indic. 2017, 80, 74–83. [Google Scholar] [CrossRef]
- Rogers, C.; Caro, M. Song sparrows, top carnivores and nest predation: A test of the mesopredator release hypothesis. Oecologia 1998, 116, 227–233. [Google Scholar] [CrossRef]
- Robinson, W.D.; Sherry, T.W. Mechanisms of avian population decline and species loss in tropical forest fragments. J. Ornithol. 2012, 153, 141–152. [Google Scholar] [CrossRef]
- Stantial, M.L.; Cohen, J.B.; Darrah, A.J.; Farrell, S.L.; Maslo, B. The effect of top predator removal on the distribution of a mesocarnivore and nest survival of an endangered shorebird. Avian Conserv. Ecol. 2021, 16, 8. [Google Scholar] [CrossRef]
- Savidge, J.A. Extinction of an island forest avifauna by an introduced snake. Ecology 1987, 68, 660–668. [Google Scholar] [CrossRef]
- Bonnington, C.; Gaston, K.J.; Evans, K.L. Fearing the feline: Domestic cats reduce avian fecundity through trait-mediated indirect effects that increase nest predation by other species. J. Appl. Ecol. 2013, 50, 15–24. [Google Scholar] [CrossRef]
- Ballarini, Y.; Chaves, F.G.; Vecchi, M.B.; Alves, M.A.S. High rates of predation of the nests of two endemic antbirds of the Brazilian Atlantic Forest by invasive marmosets (Callithrix spp.). Ann. Zool. Fenn. 2021, 58, 31–40. [Google Scholar] [CrossRef]
- Dinsmore, S.J.; Gaines, E.P.; Pearson, S.F.; Lauten, D.J.; Castelein, K.A. Factors affecting Snowy Plover chick survival in a managed population. Condor 2017, 119, 34–43. [Google Scholar] [CrossRef][Green Version]
- Maina, G.G.; Jackson, W.M. Effects of fragmentation on artificial nest predation in a tropical forest in Kenya. Biol. Conserv. 2003, 111, 161–169. [Google Scholar] [CrossRef]
- Newmark, W.D.; Stanley, T.R. Habitat fragmentation reduces nest survival in an Afrotropical bird community in a biodiversity hotspot. Proc. Natl. Acad. Sci. USA 2011, 108, 1488–11493. [Google Scholar] [CrossRef] [PubMed]
- Visco, D.M.; Sherry, T.W. Increased abundance, but reduced nest predation in the chestnut-backed antbird in Costa Rican rainforest fragments: Surprising impacts of a pervasive snake species. Biol. Conserv. 2015, 188, 22–31. [Google Scholar] [CrossRef]
- Pontes, A.R.M.; Beltrão, A.C.M.; Normande, I.C.; Malta, A.D.J.R.; Silva Júnior, A.P.D.; Santos, A.M.M. Mass extinction and the disappearance of unknown mammal species: Scenario and perspectives of a biodiversity hotspot’s hotspot. PLoS ONE 2016, 11, e0150887. [Google Scholar]
- Dias, T.C.; Silveira, L.F.; Francisco, M.R. Spatiotemporal dynamics reveals forest rejuvenation, fragmentation, and edge effects in an Atlantic Forest hotspot, the Pernambuco Endemism Center, northeastern Brazil. PLoS ONE 2023, 18, e0291234. [Google Scholar] [CrossRef]
- Pereira, G.A.; Dantas, S.M.; Silveira, L.F.; Roda, A.S.; Albano, C.; Sonntag, F.A.; Leal, S.; Periquito, M.C.; Malacco, G.B.; Lees, A.C. Status of the globally threatened forest birds of northeast Brazil. Pap. Avulsos Zool. 2014, 54, 177–194. [Google Scholar] [CrossRef]
- Instituto Chico Mendes de Conservação da Biodiversidade (ICMBio). Livro Vermelho da Fauna Brasileira Ameaçada de Extinção: Volume III—Aves; ICMBio/MMA: Brasília, Brazil, 2018.
- Dickens, J.K.; Bitton, P.P.; Bravo, A.; Silveira, L.F. Species limits, patterns of secondary contact and a new species in the Trogon rufus complex (Aves: Trogonidae). Zool. J. Linn. Soc. 2021, 193, 499–540. [Google Scholar] [CrossRef]
- Lima, R.D.; Silveira, L.F.; Lemos, R.C.A.; Lobo-Araujo, L.W.; Andrade, A.B.; Francisco, M.R.; Efe, M.A. An annotated avian inventory of the Brazilian state of Alagoas, one of the world’s most threatened avifauna. Pap. Avulsos Zool. 2022, 62, e202262034. [Google Scholar] [CrossRef]
- Develey, P.F.; Phalan, B. Bird Extinctions in Brazil’s Atlantic Forest and How They Can Be Prevented. Front. Ecol. Evol. 2021, 9, e624587. [Google Scholar] [CrossRef]
- Mayfield, H. Nesting success calculated from exposure. Wilson Bull. 1961, 73, 255–261. [Google Scholar]
- Roda, S.A.; Santos, A.M.M. Avaliação de Fragmentos Florestais Para uma Possível Reintrodução do Mutum-de-Alagoas em seu Ambiente natural; Centro de Pesquisas Ambientais do Nordeste CEPAN: Recife, Brazil, 2005. [Google Scholar]
- Pereira, G.A.; Araújo, H.F.P.; Azevedo-Júnior, S.M. Distribution and conservation of three important bird groups of the Atlantic Forest in north-east Brazil. Braz. J. Biol. 2016, 76, 1004–1020. [Google Scholar] [CrossRef] [PubMed]
- Barros, A.H.C.; Araújo-Filho, J.C.; Silva, A.B.; Santiago, G.A.C.F. Climatologia do Estado de Alagoas; Boletín de Pesquisa e Desenvolvimento: Embrapa, Brasil, 2012. [Google Scholar]
- Studer, A.; Sousa, M.C.; Barcena-Goyena, B. The breeding biology and nest success of the Short-tailed Antthrush Chamaeza campanisona (Aves: Formicariidae) in the Atlantic rainforest of northeastern Brazil. Zoologia 2018, 35, e12906. [Google Scholar] [CrossRef]
- Studer, A.; Sousa, M.C.; Barcena-Goyena, B. Breeding biology and nesting success of the endemic Black-cheeked Gnateater (Conopophaga melanops). Stud. Neotrop. Fauna Environ. 2019, 54, 157–162. [Google Scholar] [CrossRef]
- Martin, T.E.; Geupel, G.R. Nest-monitoring plots: Methods for locating nests and monitoring success. J. Field Ornithol. 1993, 64, 507–519. [Google Scholar]
- Ribeiro-Silva, L.; Perrella, D.F.; Biagolini, C.H., Jr.; Zima, P.V.Q.; Piratelli, A.J.; Schlindwein, M.N.; Galetti Junior, P.M.; Francisco, M.R. Testing camera traps as a potential tool for detecting nest predation of birds in a tropical rainforest environment. Zoologia 2018, 35, e14678. [Google Scholar] [CrossRef]
- Johnson, D.H. Estimating nest success: The Mayfield method and an alternative. Auk 1979, 96, 651–661. [Google Scholar]
- Zimmer, K.; Isler, M.L. Plain Antvireo (Dysithamnus mentalis). In Birds of the World; del Hoyo, J., Elliott, A., Sargatal, J., Christie, D.A., de Juana, E., Eds.; Cornell Lab of Ornithology: Ithaca, NY, USA, 2020. [Google Scholar]
- Snow, D. Blue-backed Manakin (Chiroxiphia pareola). In Birds of the World; del Hoyo, J., Elliott, A., Sargatal, J., Christie, D.A., de Juana, E., Eds.; Cornell Lab of Ornithology: Ithaca, NY, USA, 2020. [Google Scholar]
- Koloff, J.; Mennill, D. Barred Antshrike (Thamnophilus doliatus). In Birds of the World; Schulenberg, T.S., Ed.; Cornell Lab of Ornithology: Ithaca, NY, USA, 2020. [Google Scholar]
- Costa, H.C.; Guedes, T.B.; Bérnils, R.S. Lista de répteis do Brasil: Padrões e tendências. Herpetol. Bras. 2021, 10, 109–279. [Google Scholar] [CrossRef]
- Pacheco, J.F.; Silveira, L.F.; Aleixo, A.; Agne, C.E.; Bencke, G.A.; Bravo, G.A.; Brito, G.R.R.; Cohn-Haft, M.; Maurício, G.N.; Naka, L.N.; et al. Annotated checklist of the birds of Brazil by the Brazilian Ornithological Records Committee—Second edition. Ornithol. Res. 2021, 29, 94–105. [Google Scholar] [CrossRef]
- Quintela, F.M.; Rosa, C.A.; Feijó, A. Updated and annotated checklist of recent mammals from Brazil. An. Acad. Bras. Cienc. 2020, 92, e20191004. [Google Scholar] [CrossRef] [PubMed]
- Gotelli, N.J.; Colwell, R.K. Estimating species richness. In Biological Diversity: Frontiers in Measurement and Assessment; Magurran, A.E., McGill, B.J., Eds.; Oxford University Press: Oxford, UK, 2011. [Google Scholar]
- Tobler, M.W.; Carrillo-Percastegui, S.E.; Pitman, R.L.; Mares, R.; Powell, G. An evaluation of camera traps for inventorying large- and medium-sized terrestrial rainforest mammals. Anim. Conserv. 2008, 11, 169–178. [Google Scholar] [CrossRef]
- Arévalo-Sandi, A.R.; Gonçalves, A.L.S.; Onizawa, K.; Yabe, T.; Spironello, W.R. Mammal diversity among vertical strata and the evaluation of a survey technique in a central Amazonian forest. Pap. Avulsos Zool. 2021, 61, e20216133. [Google Scholar] [CrossRef]
- Morales-Martinez, D.M.; Atuesta-Dimiani, N.; Martínez-Medina, D.; Gutiérrez-Sanabria, D.R.; Rodríguez-Posada, M.E. Completeness of rapid assessments of medium and large mammal diversity in the northwestern Amazon in Colombia. Acta Amazon. 2021, 51, 224–233. [Google Scholar] [CrossRef]
- Burnham, K.P.; Overton, W.S. Robust estimation of population size when capture probabilities vary among animals. Ecology 1979, 60, 927–936. [Google Scholar] [CrossRef]
- Palmer, M.W. Estimating species richness: The second-order Jackknife reconsidered. Ecology 1991, 72, 1512–1513. [Google Scholar] [CrossRef]
- Oksanen, J.; Blanchet, F.G.; Friendly, M.; Kindt, R.; Legendre, P.; McGlinn, D.; Minchin, P.R.; O’Hara, R.B.; Simpson, G.L.; Solymos, P.; et al. Vegan: Community Ecology Package, R Package Version 2.5-7. 2020. Available online: https://CRAN.R-project.org/package=vegan (accessed on 15 December 2023).
- Rosa, D.C.P.; Brocardo, C.R.; Rosa, C.; Castro, A.B.; Norris, D.; Fadini, R. Species-rich but defaunated: The case of medium and large-bodied mammals in a sustainable use protected area in the Amazon. Acta Amazon. 2021, 51, 323–333. [Google Scholar] [CrossRef]
- Ponce-Martins, M.; Lopes, C.K.M.; Carvalho-Jr, E.A.B.; Castro, F.M.R.; Paulac, M.J.; Pezzuti, J.C.B. Assessing the contribution of local experts in monitoring Neotropical vertebrates with camera traps, linear transects and track and sign surveys in the Amazon. Perspect. Ecol. Conserv. 2022, 20, 303–313. [Google Scholar] [CrossRef]
- Lim, S.J.; Han, S.H.; Kim, K.Y.; Hong, S.; Park, Y.C. Relative abundance of mammals and estimation of minimum trapping effort using camera traps in Jangsudae, Seoraksan National Park. Mamm. Study 2023, 48, 171–179. [Google Scholar] [CrossRef]
- Develey, P.F.; Goerck, J.M. Important Bird Areas Americas—Priority Sites for Biodiversity Conservation; BirdLife Conservation Series No. 16; BirdLife International: Cambridge, UK, 2009. [Google Scholar]
- Marini, M.A. Nesting success of birds from Atlantic Forest fragments. Rev. Bras. Ornitol. 2017, 25, 77–83. [Google Scholar] [CrossRef]
- Zima, P.V.Q.; Perrella, D.F.; Biagolini, C.H., Jr.; Ribeiro-Silva, L.; Francisco, M.R. Breeding behavior of the Atlantic Forest endemic Blue Manakin (Chiroxiphia caudata). Wilson J. Ornithol. 2017, 129, 53–61. [Google Scholar] [CrossRef]
- Lima, A.M.X.; Roper, J.J. Population dynamics of the black-cheeked gnateater (Conopophaga melanops, Conopophagidae) in southern Brazil. J. Trop. Ecol. 2009, 25, 605–613. [Google Scholar] [CrossRef]
- Studer, A.; Sousa, M.C.; Barcena-Goyena, B. Reproduction and nest success of the Scalloped Antbird, Myrmoderus ruficauda (Passeriformes: Thamnophilidae), in an Atlantic rainforest of northeastern Brazil. Atual. Ornitol. 2017; 199, 33–37. [Google Scholar]
- Dinsmore, S.J.; White, G.C.; Knopf, F.L. Advanced techniques for modeling avian nest survival. Ecology 2002, 83, 3476–3488. [Google Scholar] [CrossRef]
- Coelho, C.G.; Falótico, T.; Izar, P.; Mannu, M.; Resende, B.D.; Siqueira, J.O.; Ottoni, E.B. Social learning strategies for nut-cracking by tufted capuchin monkeys (Sapajus spp.). Anim. Cogn. 2015, 18, 911–919. [Google Scholar] [CrossRef] [PubMed]
- Zima, P.V.Q.; Perrella, D.F.; Francisco, M.R. The influence of egg presence and eggshell colour in the attraction of visually oriented predators to nests of a tropical forest bird. Ibis 2021, 163, 1080–1086. [Google Scholar] [CrossRef]
- Cockle, K.L.; Bodrati, A.; Lammertink, M.; Bonaparte, E.B.; Ferreyra, C.; Di Sallo, F.G. Predators of bird nests in the Atlantic Forest of Argentina and Paraguay. Wilson J. Ornithol. 2016, 128, 120–131. [Google Scholar] [CrossRef]
- Londoño, G.A.; Gomez, J.P.; Sánchez-Martínez, M.A.; Levey, D.J.; Robinson, S.K. Changing patterns of nest predation and predator communities along a tropical elevation gradient. Ecol. Lett. 2022, 26, 609–620. [Google Scholar] [CrossRef]
- Duron, Q.; Bourguet, E.; Meringgo, H.D.; Millon, A.; Vidal, E. Invasive rats strengthen predation pressure on bird eggs in a South Pacific island rainforest. Curr. Zool. 2017, 63, 583–590. [Google Scholar] [CrossRef] [PubMed]
- Vanderwerf, E.A.; Smith, D.G. Effects of alien rodent control on demography of the O’ahu ‘Elepaio, an endangered Hawaiian forest bird. Pac. Conserv. Biol. 2002, 8, 73–81. [Google Scholar] [CrossRef]
- Jansen, W.P. Rat Rattus control at nests of the endangered kakapo Strigops habroptilus on Codfish Island, New Zealand. Conserv. Evid. 2005, 2, 1–2. [Google Scholar]




{kind=link}
{kind=link}
{kind=link}
| Bird Species | Total | Depredated | Failed | Successful | Apparent Survival |
|---|---|---|---|---|---|
| Columbidae | |||||
| Leptotila sp. | 1 | 1 | 0% | ||
| Geotrygon montana | 1 | 1 | 0% | ||
| Caprimulgidae | |||||
| Antrostomus rufus | 2 | 1 | 1 | 50% | |
| Nyctidromus albicollis | 5 | 3 | 2 | 40% | |
| Trochilidae | |||||
| Chlorestes notata | 5 | 3 | 1 | 1 | 20% |
| Thamnophilidae | |||||
| Myrmotherula axillaris | 4 | 2 | 2 | 50% | |
| Dysithamnus mentalis | 23 | 20 | 2 | 1 | 4.3% |
| Thamnophilus aethiops | 10 | 6 | 2 | 2 | 20% |
| Conopophagidae | |||||
| Conopophaga melanops | 12 | 11 | 0 | 1 | 8.3% |
| Pipridae | |||||
| Chiroxiphia pareola | 10 | 8 | 1 | 1 | 10% |
| Ceratopipra rubrocapilla | 1 | 1 | 0% | ||
| Tityridae | |||||
| Schiffornis turdina | 2 | 2 | 0% | ||
| Platyrinchidae | |||||
| Platyrinchus mystaceus | 2 | 2 | 0% | ||
| Rhynchocyclidae | |||||
| Leptopogon amaurocephalus | 1 | 1 | 0 | 0 | 0% |
| Hemitriccus griseipectus | 5 | 1 | 2 | 2 | 40% |
| Bird Species | S. merianae | Chironius sp. | D. corais | Philodryas sp. | B. nitidus | M. semitorquatus | D. aurita | G. emiliae | M. demerarae | C. jacchus | N. nasua |
|---|---|---|---|---|---|---|---|---|---|---|---|
| Leptotila sp. | 1 | ||||||||||
| A. rufus | 1 | ||||||||||
| N. albicollis | 2 | ||||||||||
| C. notata | 1 | ||||||||||
| M. axillaris | 1 | ||||||||||
| D. mentalis | 2 | 1 | 1 | 1 | 1 | 6 | 1 | ||||
| T. aethiops | 1 | 1 | 1 | ||||||||
| C. melanops | 2 | 2 | 1 | 2 | |||||||
| C. pareola | 1 | 1 | 2 | ||||||||
| S. turdina | 1 | 1 | |||||||||
| L. amaurocephalus | 1 | ||||||||||
| Total | 7 | 1 | 1 | 1 | 1 | 1 | 1 | 6 | 2 | 9 | 6 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Lobo-Araújo, L.W.; Costa, M.C.; Silveira, L.F.; Francisco, M.R. Massive Bird Nest Losses: A Neglected Threat for Passerine Birds in Atlantic Forest Fragments from the Pernambuco Endemism Center. Diversity 2024, 16, 207. https://doi.org/10.3390/d16040207
Lobo-Araújo LW, Costa MC, Silveira LF, Francisco MR. Massive Bird Nest Losses: A Neglected Threat for Passerine Birds in Atlantic Forest Fragments from the Pernambuco Endemism Center. Diversity. 2024; 16(4):207. https://doi.org/10.3390/d16040207
Chicago/Turabian StyleLobo-Araújo, Lahert William, Mariellen Cristine Costa, Luís Fábio Silveira, and Mercival Roberto Francisco. 2024. "Massive Bird Nest Losses: A Neglected Threat for Passerine Birds in Atlantic Forest Fragments from the Pernambuco Endemism Center" Diversity 16, no. 4: 207. https://doi.org/10.3390/d16040207
APA StyleLobo-Araújo, L. W., Costa, M. C., Silveira, L. F., & Francisco, M. R. (2024). Massive Bird Nest Losses: A Neglected Threat for Passerine Birds in Atlantic Forest Fragments from the Pernambuco Endemism Center. Diversity, 16(4), 207. https://doi.org/10.3390/d16040207