
Type of Material, Not Tree Hole Characteristics Shapes Uropodina Mites’ Species Composition in Excavated Tree Holes


Abstract
1. Introduction
2. Materials and Methods
2.1. Study Area
2.2. Data Collection
2.3. Tree Hole Contents’ Analysis
- -
- wood debris—loose or soft wood debris in different stages of decay;
- -
- bird nest remains—parts of bird nests consisting of different materials, such as moss, grass, leaves, feathers, and wool;
- -
- insect remains—predominantly bat guano and remains of woodpecker food. In two cases, this last category included also the remains of bumblebee and wasp nests with dead specimens. For the analysis of the presence of Chiropturopoda nidiphila only, we subdivided this category into holes containing only bat guano and holes containing only insect remains (other than bat guano).
2.4. Data Analysis
2.5. Statistical Analysis
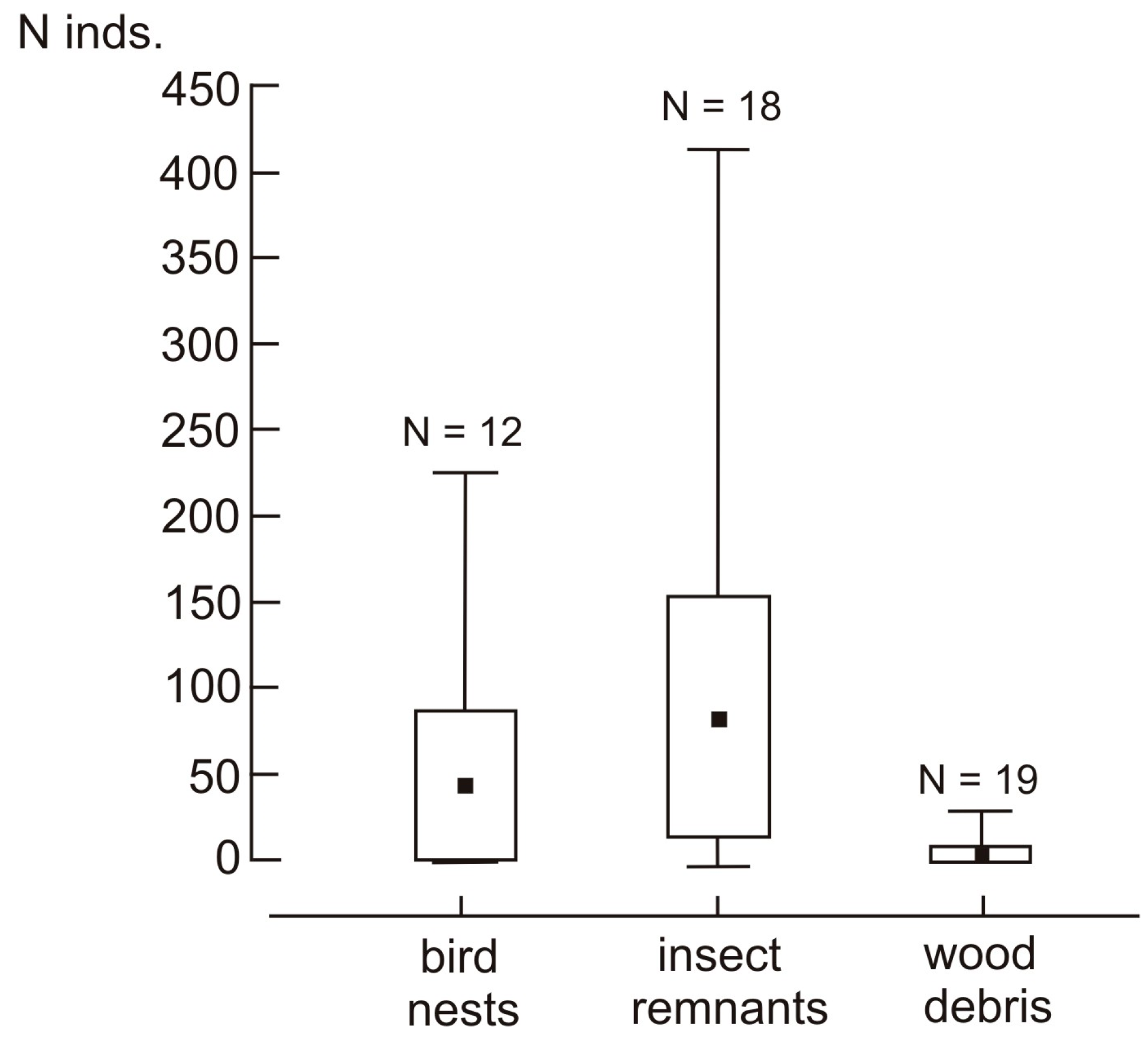
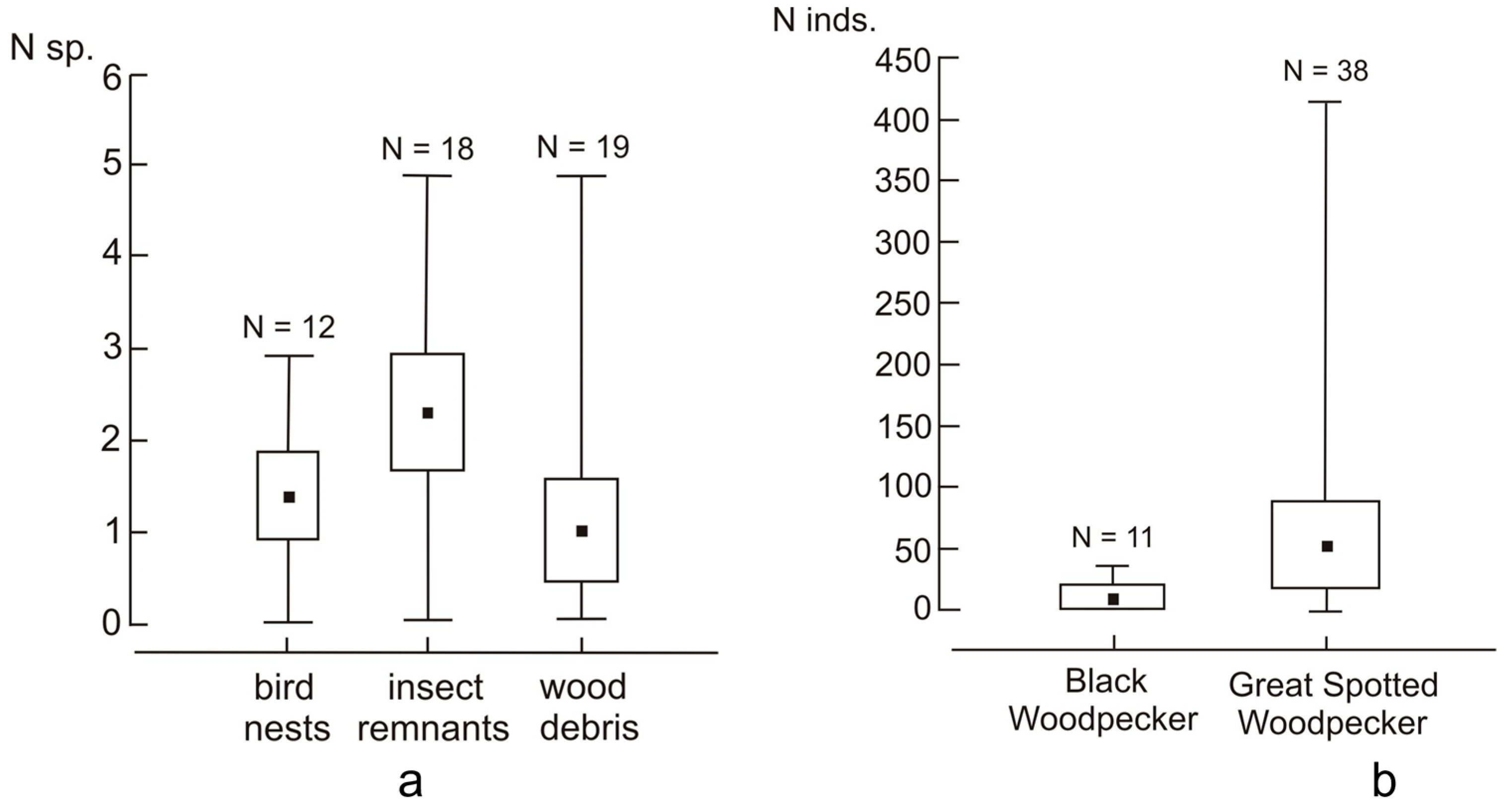
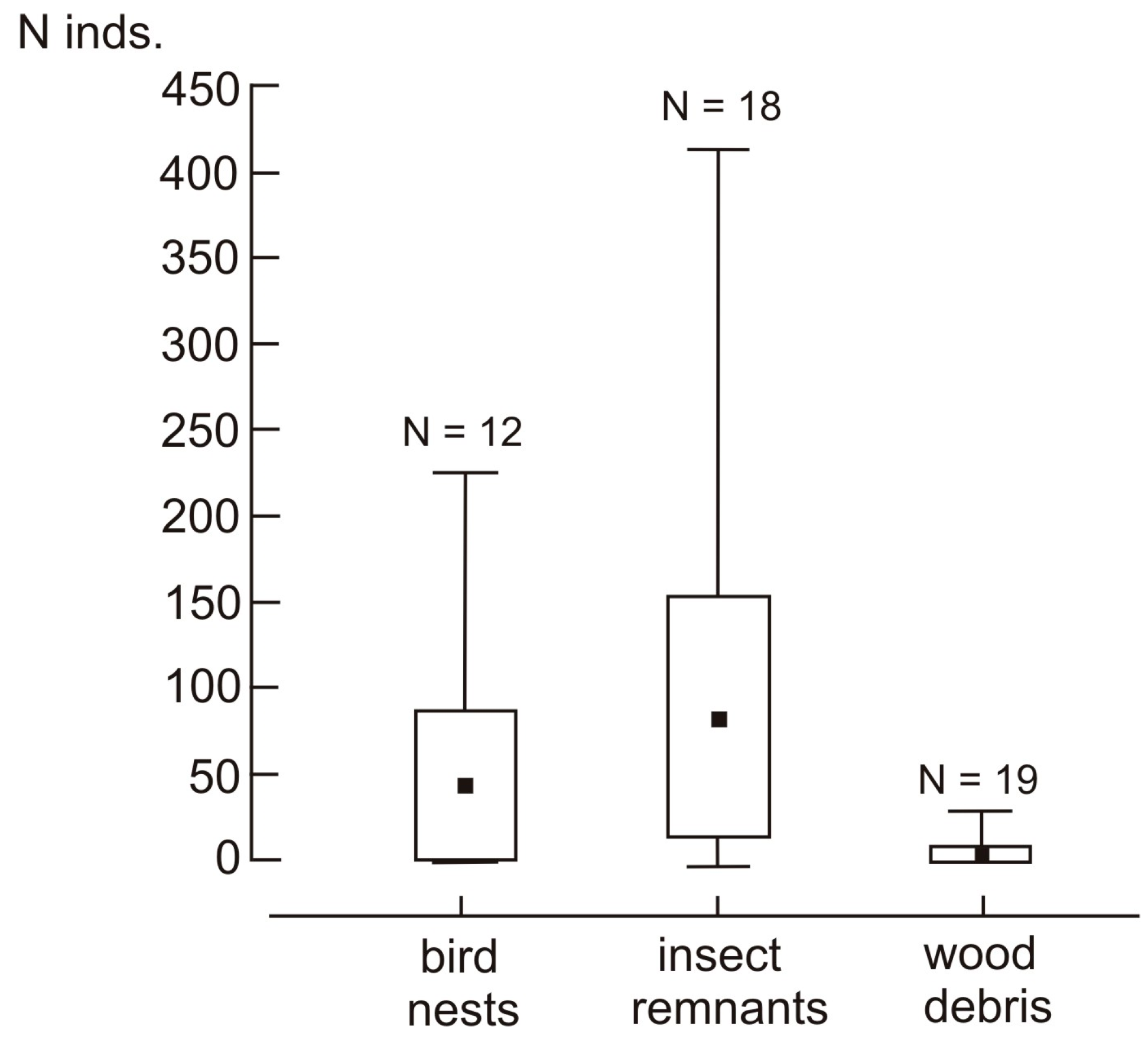
3. Results
3.1. Species Composition and the Structure of Assemblage
3.2. Effect of Tree Hole Characteristics and Type of Nest Material on the Structure of the Assemblage
3.3. Effect of Bat Guano Presence on the Occurrence of Ch. nidiphila
4. Discussion
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Pan, Y.; Birdsey, R.A.; Phillips, O.L.; Jackson, R.B. The structure, distribution, and biomass of the World’s forests. Annu. Rev. Ecol. Evol. Syst. 2013, 44, 593–622. [Google Scholar] [CrossRef]
- Rehm, J.; Lohmus, A. Tree cavities in forests—The broad distribution pattern of a keystone structure for biodiversity. For. Ecol. Manag. 2011, 262, 579–585. [Google Scholar] [CrossRef]
- Cockle, K.L.; Martin, K.; Wesołowski, T. Woodpeckers, decay, and the future of cavity-nesting vertebrate communities worldwide. Front. Ecol. Environ. 2011, 9, 377–382. [Google Scholar] [CrossRef]
- Cockle, K.L.; Martin, K.; Robledo, G. Linking fungi, trees, and hole-using birds in a Neotropical tree-cavity network: Pathways of cavity production and implications for conservation. For. Ecol. Manag. 2012, 264, 210–219. [Google Scholar] [CrossRef]
- Van der Hoek, Y.; Gaona, G.V.; Martin, K. The diversity, distribution and conservation status of the tree-cavity-nesting birds of the world. Divers. Distrib. 2017, 23, 1120–1131. [Google Scholar] [CrossRef]
- Blanc, L.A.; Walters, J.R. Cavity-nest webs in a longleaf pine ecosystem. Condor 2008, 110, 80–92. [Google Scholar] [CrossRef]
- Trzcinski, M.K.; Cockle, K.L.; Norris, A.R.; Edworthy, M.; Wiebe, K.L.; Martin, K. Woodpeckers and other excavators maintain the diversity of cavity-nesting vertebrates. J. Anim. Ecol. 2022, 91, 1251–1265. [Google Scholar] [CrossRef] [PubMed]
- Wesołowski, T. “Lifespan” of woodpecker-made holes in a primeval temperate forest: A thirty year study. For. Ecol. Manag. 2011, 262, 1846–1852. [Google Scholar] [CrossRef]
- Cockle, K.L.; Trzcinski, M.K.; Wiebe, K.L.; Edworthy, A.B.; Martin, K. Lifetime productivity of tree cavities used by cavity-nesting animals in temperate and subtropical forests. Ecol. Appl. 2019, 29, e01916. [Google Scholar] [CrossRef] [PubMed]
- Gibbons, P.; Lindenmayer, D. Tree Hollows and Wildlife Conservation in Australia; CSIRO Publishing: Collingwood, Australia, 2002. [Google Scholar]
- Czeszczewik, D.; Walankiewicz, W.; Stańska, M. Small mammals in nests of cavity-nesting birds: Why should ornithologists study rodents? Can. J. Zool. 2008, 86, 286–293. [Google Scholar] [CrossRef]
- McComb, W.C.; Noble, R.E. Invertebrate use of natural tree cavities and vertebrate nest boxes. Am. Midl. Nat. 1982, 107, 163–172. [Google Scholar] [CrossRef]
- Nilsson, S.G.; Baranowski, R. Habitat predictability and the occurrence of wood beetles in old-growth beech forests. Ecography 1997, 20, 491–498. [Google Scholar] [CrossRef]
- Hanzelka, J.; Baroni, D.; Martikainen, P.; Eeva, T.; Laaksonen, T. Cavity-breeding birds create specific microhabitats for diverse arthropod communities in boreal forests. Biodivers. Conserv. 2023, 32, 3845–3874. [Google Scholar] [CrossRef]
- Hebda, G.; Kandziora, A.; Mitrus, S. Decomposition of nest material in tree holes and nest-boxes occupied by European Starlings Sturnus vulgaris: An experimental study. Acta Ornithol. 2017, 52, 119–125. [Google Scholar] [CrossRef]
- Maziarz, M.A.; Broughton, R.K.; Wesołowski, T. Microclimate in tree cavities and nest-boxes: Implications for hole-nesting birds. For. Ecol. Manag. 2017, 389, 306–313. [Google Scholar] [CrossRef]
- Mazgajski, T.D. Effect of old nest material on nest site selection and breeding parameters in secondary hole nesters—Review. Acta Ornithol. 2007, 42, 1–14. [Google Scholar] [CrossRef]
- Roy, L.; Bouvier, J.-C.; Lavigne, C.; Galès, M.; Buronfosse, T. Impact of pest control strategies on the arthropodofauna living in bird nests built in nestboxes in pear and apple orchards. Bull. Entomol. Res. 2013, 103, 458–465. [Google Scholar] [CrossRef] [PubMed]
- Boyes, D.H.; Lewis, O.T. Ecology of Lepidoptera associated with bird nests in mid-Wales, UK: Ecology of Lepidoptera associated with bird nests. Ecol. Entomol. 2019, 44, 1–10. [Google Scholar] [CrossRef]
- Krištofík, J.; Mašán, P.; Šustek, Z.; Nuhličková, S. Arthropods (Acarina, Coleoptera, Siphonaptera) in nests of hoopoe (Upupa epops) in Central Europe. Biologia 2013, 68, 155–161. [Google Scholar] [CrossRef]
- Cosandey, V.; Séchaud, R.; Béziers, P.; Chittaro, Y.; Sanchez, A.; Roulin, A. Nidicolous beetle species richness is driven by Barn Owl’s nests occupancy and landscape structure. J. Ornithol. 2021, 162, 857–864. [Google Scholar] [CrossRef]
- Jaworski, T.; Gryz, J.; Krauze-Gryz, D.; Plewa, R.; Bystrowski, C.; Dobosz, R.; Horák, J. My home is your home: Nest boxes for birds and mammals provide habitats for diverse insect communities. Insect Conserv. Divers. 2022, 15, 461–469. [Google Scholar] [CrossRef]
- Rendell, W.B.; Verbeek, N.A.M. Are avian ectoparasites more numerous in nest boxes with old nest material? Can. J. Zool. 1996, 74, 1819–1825. [Google Scholar] [CrossRef]
- Eeva, T.; Andersson, T.; Berglund, Å.M.M.; Brommer, J.E.; Hyvönen, R.; Klemola, T.; Laaksonen, T.; Loukola, O.; Morosinotto, C.; Rainio, K.; et al. Species and abundance of ectoparasitic flies (Diptera) in pied flycatcher nests in Fennoscandia. Parasites Vectors 2015, 8, 648. [Google Scholar] [CrossRef] [PubMed]
- Nordberg, S. Biologisch-ökologische Untersuchungen über die Vogelnidicolen. Acta Zool. Fenn. 1936, 21, 1–168. [Google Scholar]
- Błoszyk, J.; Olszanowski, J. Materiały do znajomości fauny roztoczy gniazd i budek lęgowych ptaków. II. Różnice w liczebności i składzie gatunkowym populacji Uropodina (Acari, Anactinotrichida) budek lęgowych na Mierzei Wiślanej na podstawie dwuletnich obserwacji. Przegl. Zool. 1986, 30, 63–66. [Google Scholar]
- Napierała, A.; Maziarz, M.; Hebda, G.; Broughton, R.K.; Rutkowski, T.; Zacharyasiewicz, M.; Błoszyk, J. Lack of specialist nidicoles as a characteristic of mite assemblages inhabiting nests of the ground-nesting wood warbler, Phylloscopus sibilatrix (Aves: Passeriformes). Exp. Appl. Acarol. 2021, 84, 149–170. [Google Scholar] [CrossRef]
- Mašan, P. Mites of the Cohort Uropodina (Acarina, Mesostigmata) in Slovakia, 1st ed.; Annotationes Zoologicae et Botanicae: Bratislava, Slovakia, 2001. [Google Scholar]
- Błoszyk, J.; Bajerlein, D.; Gwiazdowicz, D.J.; Halliday, R.B.; Dylewska, M. Uropodine mite communities (Acari: Mesostigmata) in birds’ nests in Poland. Belg. J. Zool. 2006, 136, 145–153. [Google Scholar]
- Melekhina, E.N.; Korolev, A.N.; Selivanova, N.P. Oribatid Mites (Oribatida) Associated with Nests of Hollow Nesting Birds, on the Example of a Model Species, the European Pied Flycatcher (Ficedula hypoleuca), in the Taiga Forests of the European North-East of Russia. Diversity 2023, 15, 765. [Google Scholar] [CrossRef]
- Błoszyk, J.; Hebda, G.; Adamski, Z.; Zacharyasiewicz, M. Redescription of Chiropturopoda nidiphila Wiśniewski & Hirschmann (Acari: Uropodina) from a woodpecker’s tree holes, including all development stages and first notes on its ecology. Syst. Appl. Acarol. 2021, 26, 1867–1899. [Google Scholar] [CrossRef]
- Błoszyk, J. Geograficzne i Ekologiczne Zróżnicowanie Zgrupowań Roztoczy z Kohorty Uropodina (Acari: Mesostigmata) w Polsce: Uropodina Lasów Grądowych (Carpinion betuli), 1st ed.; Kontekst: Poznań, Poland, 1999. [Google Scholar]
- Napierała, A.; Błoszyk, J. Unstable microhabitats (merocenoses) as specific habitats of Uropodina mites (Acari: Mesostigmata). Exp. Appl. Acarol. 2013, 60, 163–180. [Google Scholar] [CrossRef] [PubMed]
- Bregetova, N.G.; Kaditje, B.A.; Petrova, A.D. Nagkogorta Mesostigmata, Kogorta Trchytina Tragardh, 1938, Kogorta Uropodina Kramer, 1881. In Opredelitel’ Obitajuscich v Pocve Klescej, Mesostigmata; Ghilarov, M.C., Bregetova, N.G., Eds.; Nauka: Leningrad, Russia, 1977; pp. 12–25, 621–691. [Google Scholar]
- Karg, W. Acari (Acarina), Milben Uunterordnung Parasitiformes (Anactinochaeta), Uropodina Kramer, Schildkotenmilben; Die Tirwelt Deutschlands; Gustav Fischer: Jena, Germany, 1989; Volume 67, 202p. [Google Scholar]
- Venables, W.N.; Ripley, B.D. Modern Applied Statistics with S, 4th ed.; Springer: New York, NY, USA, 2002. [Google Scholar]
- Brooks, M.E.; Kristensen, K.; van Benthem, K.J.; Magnusson, A.; Berg, C.W.; Nielsen, A.; Skaug, H.J.; Maechler, M.; Bolker, B.M. glmmTMB Balances Speed and Flexibility among Packages for Zero-inflated Generalized Linear Mixed Modeling. R J. 2017, 9, 378–400. [Google Scholar] [CrossRef]
- Hartig, F. DHARMa: Residual Diagnostics for Hierarchical (Multi-Level/Mixed) Regression Models. R Package Version 0.4.6. 2022. Available online: https://CRAN.R-project.org/package=DHARMa (accessed on 10 November 2023).
- R Core Team. R: A Language and Environment for Statistical Computing; R Foundation for Statistical Computing: Vienna, Austria, 2021; Available online: https://www.R-project.org/ (accessed on 10 November 2023).
- Błoszyk, J.; Hebda, G.; Kulczak, M.; Zacharyasiewicz, M.; Rutkowski, T.; Napierała, A. Communities of Uropodina (Acari: Mesostigmata) in Nest Boxes Inhabited by Dormice (Glis glis and Muscardinus avellanarius) and Differences in Percentages of Nidicoles in Nests of Various Hosts. Animals 2023, 13, 3567. [Google Scholar] [CrossRef]
- Palacios-Vargas, J.G.; Decu, V.; Iavorski, V.; Hutzu, M.; Juberthie, C. Acari Terestia. In Encyklopedia Biospeologicai; Juberthie, C., Decu, V., Eds.; Socieate de Biospeologie: Bucarest, Romania, 1998; Volume 2, pp. 929–952. [Google Scholar]
- Błoszyk, J.; Rutkowski, T.; Wojtaszyn, G.; Książkiewicz-Parulska, Z.; Zacharyasiewicz, M.; Napierała, A. Leiodinychus orbicularis (CL Koch, 1839) in bat boxes in Poland. Eur. J. Biol. Res. 2020, 10, 150–155. [Google Scholar] [CrossRef]
- Błoszyk, J.; Wendzonka, J.; Kulczak, M.; Lubińska, K.; Napierała, A. Bird nesting boxes as a specific artificial microenvironment increasing biodiversity of mites from the suborder Uropodina (Acari: Mesostigmata): A case study of Bory Tucholskie National Park. Exp. Appl. Acarol. 2023, submitted.
- Ferenc, H.; Skoracki, M. Stan zbadania roztoczy z rodziny Spinturnicidae (Acari: Mesostigmata) w Polsce. Wiad. Parazytol. 2000, 46, 433–438. [Google Scholar] [PubMed]
- Baker, A.S.; Craven, J.S. Checklist of the mites (Arachnida: Acari) associated with bats (Mammalia: Chiroptera) in the British Isles. Syst. Appl. Acarol. Spec. Publ. 2003, 14, 1–20. [Google Scholar] [CrossRef]
- Haitlinger, R.; Łupicki, D. Arthropods (Acari, Siphonaptera, Heteroptera, Psocoptera) associated with Nyctalus noctula (Schreber, 1774) (Chiroptera: Vespertilionidae) in Southern Poland. Wiad. Parazytol. 2008, 54, 123–130. [Google Scholar] [PubMed]
- Léger, C. Bat parasites (Acari, Anoplura, Cestoda, Diptera, Hemiptera, Nematoda, Siphonaptera, Trematoda) in France (1762–2018): A literature review and contribution to a checklist. Parasite 2020, 27, 61. [Google Scholar] [CrossRef]
- Orlova, M.V.; Stanyukovich, M.K.; Orlov, O.L. Gamasid Mites (Mesostigmata: Gamasina) Parasiting Bats (Chiroptera: Rhinolophida, Vespertilionidae, Molossidae) of Palearctic Boreal Zone (Russia and Adjacent Countries); Babenko, A.S., Ed.; National Research Tomsk Stae University, Biological Institute, Russian Academy of Science, Zoological Institute, Publishing House of Tomsk State Uniwersity: Tomsk, Russia, 2015; pp. 3–149. [Google Scholar]
- Krivolutsky, D.A.; Lebedeva, N.V. Oribatid Mites (Oribatei, Acariformes) in Bird Plumage; ABF Publ.: Moscow, Russia, 2003. (In Russian) [Google Scholar]
- Krivolutsky, D.A.; Lebedeva, N.V. Oribatid Mites (Oribatei, Acariformes) in Bird Feathers: Passerines. Acta Zool. Litu. 2004, 14, 19–38. [Google Scholar] [CrossRef]
- Lebedeva, N.V. Oribatid mites transported by birds to polar islands—A review. Berichte Polar Meeresforsch. Rep. Polar Mar. Res. 2012, 640, 152–161. [Google Scholar]
- Günther, E.; Hellmann, M. Development and new tenants of holes of spotted woodpeckers (Dendrocopos) in the “Swift-forest” in the Harz Mountains (Sachsen-Anhalt). Results of twenty years investigations of the use of natural tree holes. Orn. Jber. Mus. Heine. 2005, 23, 103–122, (In German with English Summary). [Google Scholar]


{kind=link}
{kind=link}
| Species by Which the Tree Holes Were Excavated | N | Tree Species | Mean Height above the Ground [m], (Range) | Mean Depth of the Hole Chamber [cm], (Range) | Holes Containing Bat Guano |
|---|---|---|---|---|---|
| Great Spotted Woodpecker | 38 | Pine 28; Larch 2; Birch 2; Oak 6 | 5.5 (2–10) | 25 (9–39) | 9 |
| Black Woodpecker | 11 | Pine 8; Beech 2; Birch 1 | 8.6 (1.5–12) | 25 (6–48) | 0 |
| Species | N | F | M | D | P | L | D% | Nps | F% |
|---|---|---|---|---|---|---|---|---|---|
| Leiodinychus orbicularis (C. L. Koch, 1839) | 1322 | 379 | 293 | 459 | 166 | 25 | 60.5 | 27 | 55 |
| Chiropturopoda nidiphila Wiśniewski et Hirschmann, 1983 | 709 | 221 | 259 | 147 | 75 | 7 | 32.4 | 18 | 37 |
| Apionoseius infirmus (Berlese, 1887) | 46 | 16 | 10 | 19 | 0 | 1 | 2.1 | 10 | 20 |
| Nenteria banatica Feider et Hutu, 1971 | 44 | 24 | 13 | 7 | 0 | 0 | 2.0 | 2 | 4 |
| Uroobovella obovata (Canestrini et Berlese, 1884) | 28 | 17 | 9 | 2 | 0 | 0 | 1.3 | 8 | 16 |
| Pulchellaobovella pyriformis (Berlese, 1920) | 18 | 6 | 8 | 4 | 0 | 0 | 0.8 | 3 | 6 |
| Uropolyaspis hamulifera Berlese, 1904 | 6 | 3 | 2 | 1 | 0 | 0 | 0.3 | 2 | 4 |
| Oplitis paradoxa (Canestrini et Berlese, 1884) | 5 | 4 | 1 | 0 | 0 | 0 | 0.2 | 2 | 4 |
| Oodinychus ovalis (C. L. Koch, 1839) | 4 | 3 | 0 | 0 | 1 | 0 | 0.2 | 2 | 4 |
| Pulchellaobovella pulchella (Berlese, 1904) | 2 | 2 | 0 | 0 | 0 | 0 | 0.1 | 2 | 4 |
| Urodiaspis tecta (Kramer, 1876) | 1 | 1 | 0 | 0 | 0 | 0 | <0.1 | 1 | 2 |
| Pseudouropoda sp. | 1 | 0 | 1 | 0 | 0 | 0 | <0.1 | 1 | 2 |
| Total | 2186 | 676 | 596 | 639 | 242 | 33 | 100.0 | 49 | 100.0 |
| Species | N | Insect Remains | Bird Nests | Wood Debris |
|---|---|---|---|---|
| Leiodinychus orbicularis (C. L. Koch, 1839) | 1322 | 383 (12) | 883 (8) | 56 (7) |
| Chiropturopoda nidiphila Wiśniewski et Hirschmann, 1983 | 709 | 622 (10) | 58 (1) | 29 (7) |
| Apionoseius infirmus (Berlese, 1887) | 46 | 32 (7) | 13 (2) | 1 (1) |
| Nenteria banatica Feider et Hutu, 1971 | 44 | 44 (2) | 0 | 0 |
| Uroobovella obovata (Canestrini et Berlese, 1884) | 28 | 21 (4) | 2 (2) | 5 (2) |
| Pulchellaobovella pyriformis (Berlese, 1920) | 18 | 17 (2) | 1 (1) | 0 |
| Uropolyaspis hamulifera Berlese, 1904 | 6 | 3 (1) | 0 | 3 (1) |
| Oplitis paradoxa (Canestrini et Berlese, 1884) | 5 | 3 (1) | 0 | 2 (1) |
| Oodinychus ovalis (C. L. Koch, 1839) | 4 | 3 (1) | 1 (1) | 0 |
| Pulchellaobovella pulchella (Berlese, 1904) | 2 | 0 | 2 (2) | 0 |
| Urodiaspis tecta (Kramer, 1876) | 1 | 0 | 1 (1) | 0 |
| Pseudouropoda sp. | 1 | 1 (1) | 0 | 0 |
| Total | 2186 | 1129 | 961 | 96 |
| Variable | Estimate | SE | Cl 2.5% | Cl 97.5% |
|---|---|---|---|---|
| Intercept | 0.935 | 0.410 | 0.117 | 1.727 |
| Black woodpecker tree hole | −0.424 | 0.309 | −1.067 | 0.151 |
| Hole depth | 0.007 | 0.014 | −0.021 | 0.034 |
| Material volume | 0.000 | 0.001 | −0.002 | 0.001 |
| Type of material: | ||||
| bird nest | −0.528 | 0.290 | −1.124 | 0.023 |
| wood debris | −0.984 | 0.306 | −1.602 | −0.397 |
| Variable | Estimate | SE | Cl 2.5% | Cl 97.5% |
|---|---|---|---|---|
| Intercept | 3.587 | 0.806 | 1.794 | 5.461 |
| Black woodpecker tree hole | −1.357 | 0.550 | −2.504 | −0.056 |
| Hole depth | 0.044 | 0.026 | −0.012 | 0.105 |
| Material volume | <−0.000 | 0.001 | −0.003 | 0.003 |
| Type of material | ||||
| bird nest | −0.784 | 0.566 | −1.982 | 0.460 |
| wood debris | −2.953 | 0.564 | −4.087 | −1.838 |
| Variable | Estimate | SE | CI 2.5% | CI 97.5% |
|---|---|---|---|---|
| Intercept | –2.398 | 1.044 | –4.445 | –0.351 |
| Type of material: | ||||
| bat guano | 3.651 | 1.317 | 1.070 | 6.231 |
| insect remains | 1.705 | 1.261 | –0.767 | 4.177 |
| wood debris | 1.859 | 1.148 | –0.390 | 4.108 |
| Variable | Estimate | SE | Cl 2.5% | Cl 97.5% |
|---|---|---|---|---|
| Intercept | −4.930 | 1.075 | −7.037 | −2.822 |
| Type of material: | ||||
| bat guano | 2.752 | 1.066 | 0.664 | 4.841 |
| insect remains | 1.700 | 1.152 | −0.558 | 3.958 |
| wood debris | 2.279 | 1.068 | 0.187 | 4.371 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Hebda, G.; Błoszyk, J. Type of Material, Not Tree Hole Characteristics Shapes Uropodina Mites’ Species Composition in Excavated Tree Holes. Diversity 2024, 16, 114. https://doi.org/10.3390/d16020114
Hebda G, Błoszyk J. Type of Material, Not Tree Hole Characteristics Shapes Uropodina Mites’ Species Composition in Excavated Tree Holes. Diversity. 2024; 16(2):114. https://doi.org/10.3390/d16020114
Chicago/Turabian StyleHebda, Grzegorz, and Jerzy Błoszyk. 2024. "Type of Material, Not Tree Hole Characteristics Shapes Uropodina Mites’ Species Composition in Excavated Tree Holes" Diversity 16, no. 2: 114. https://doi.org/10.3390/d16020114
APA StyleHebda, G., & Błoszyk, J. (2024). Type of Material, Not Tree Hole Characteristics Shapes Uropodina Mites’ Species Composition in Excavated Tree Holes. Diversity, 16(2), 114. https://doi.org/10.3390/d16020114