
Plant Diversity Distribution along an Urbanization Gradient and Relationships with Environmental Factors in Urban Agglomerations of Henan Province, China

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
1. Introduction
2. Materials and Methods
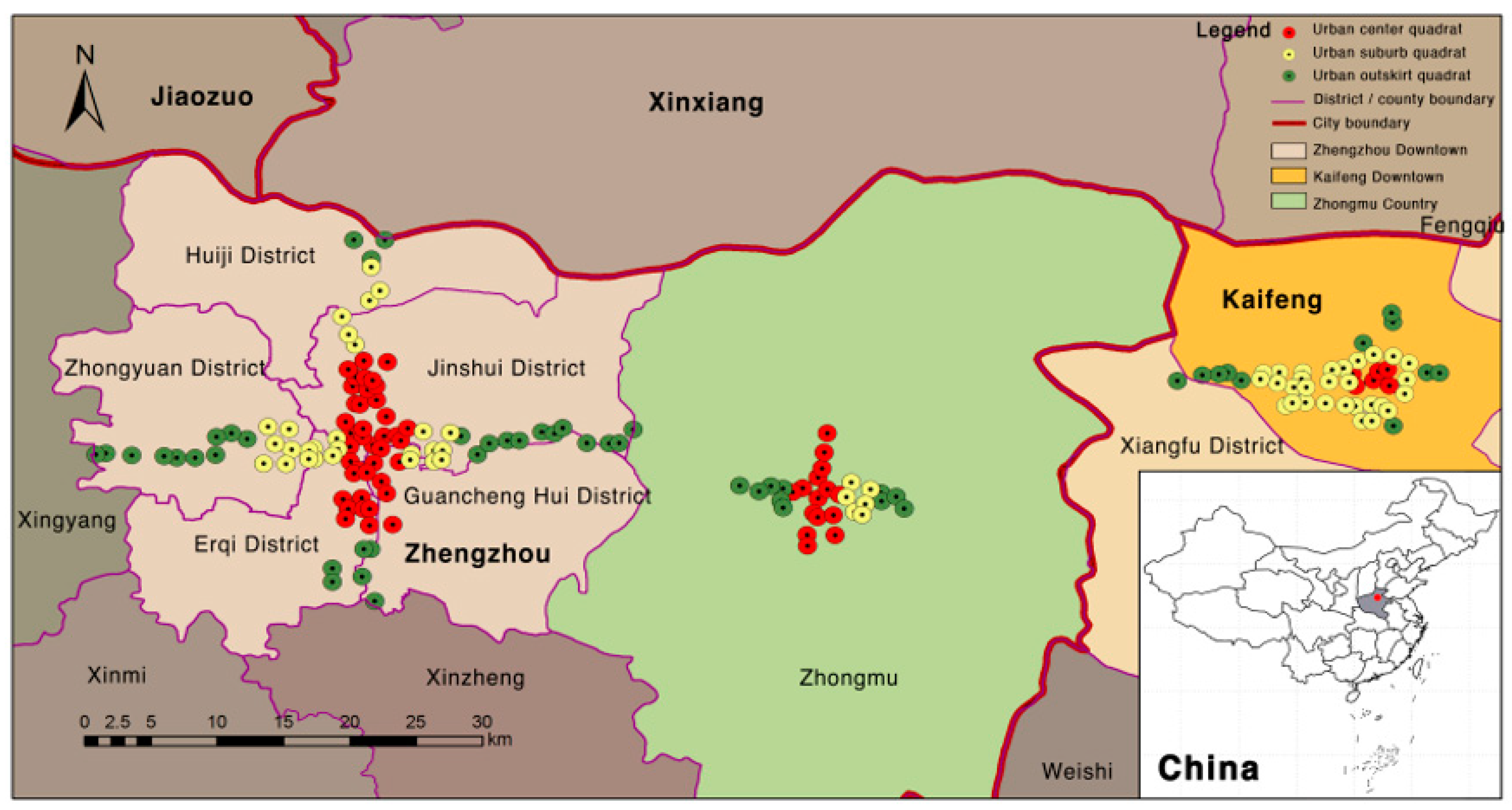
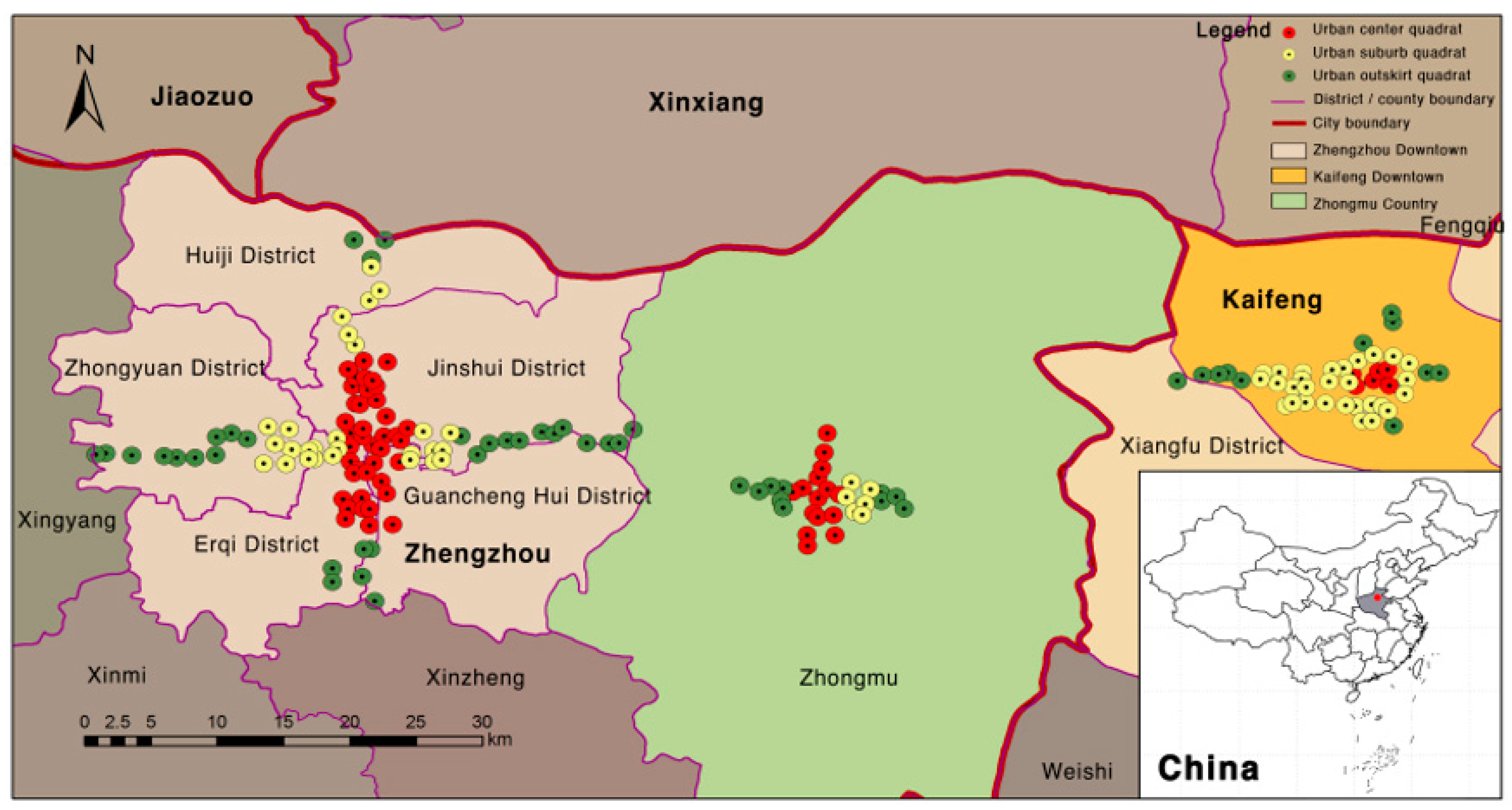
2.1. Study Area
2.2. Sampling Design and Plant Survey
2.3. Plant Diversity
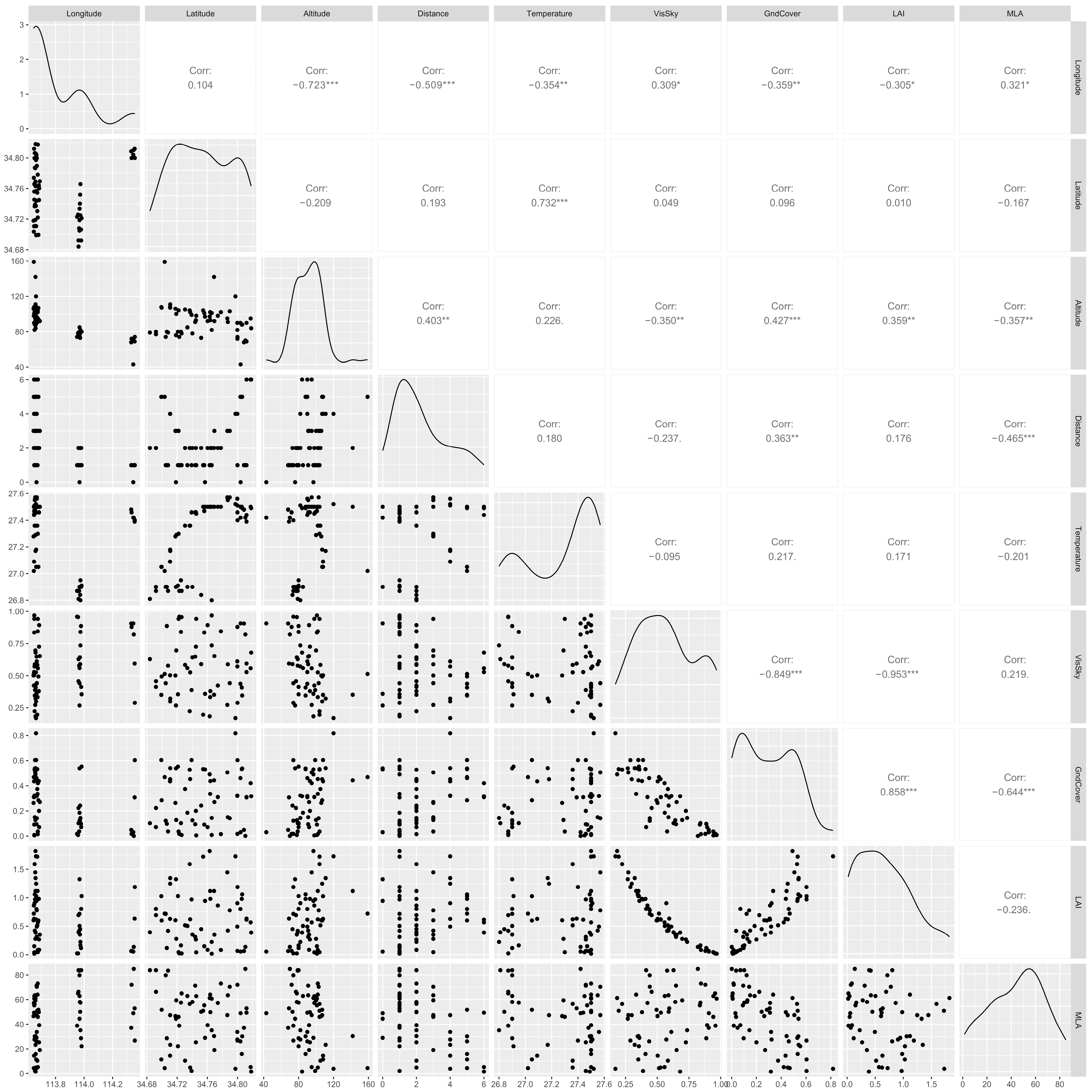
2.4. Environmental Data
2.5. Data Analyses
3. Results
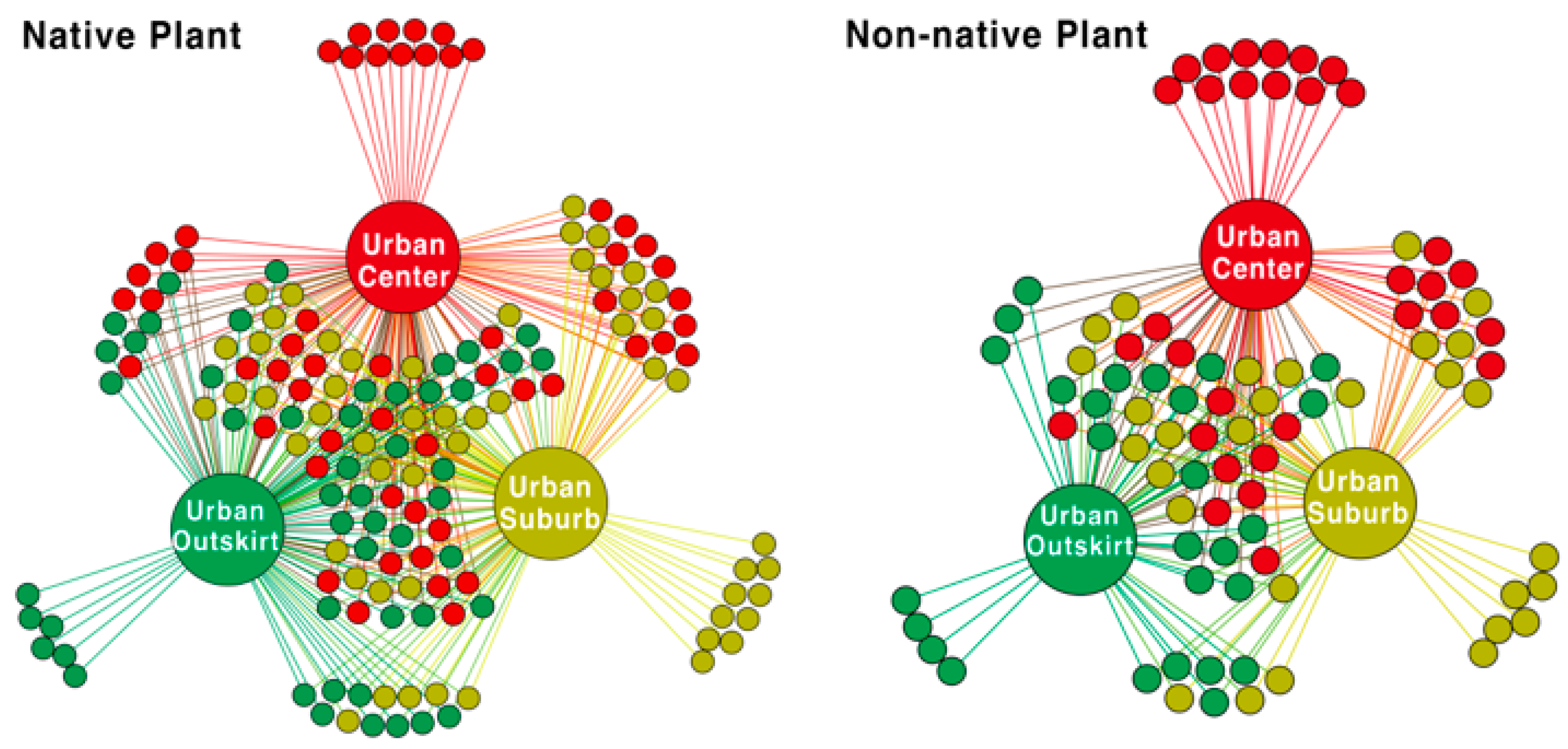
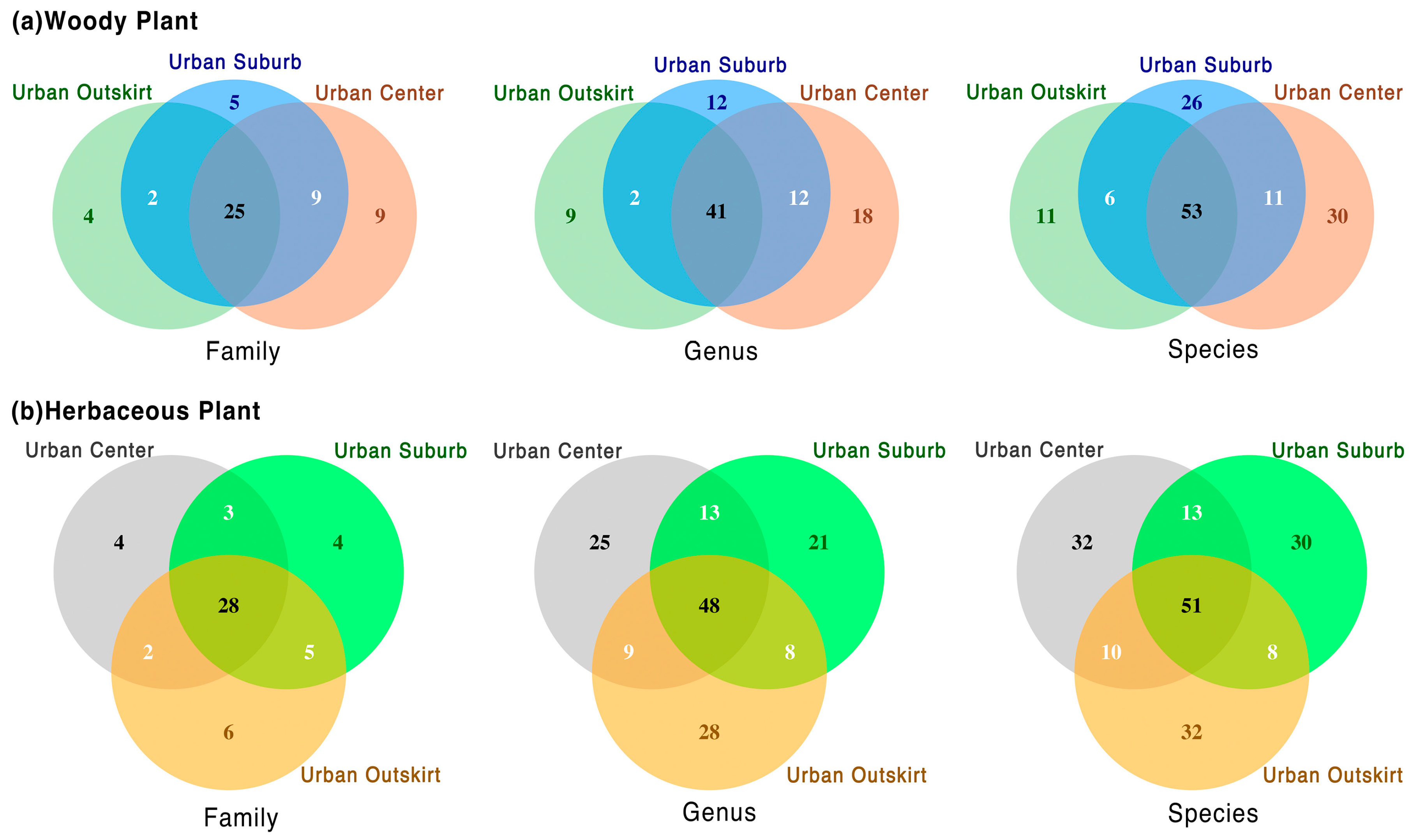
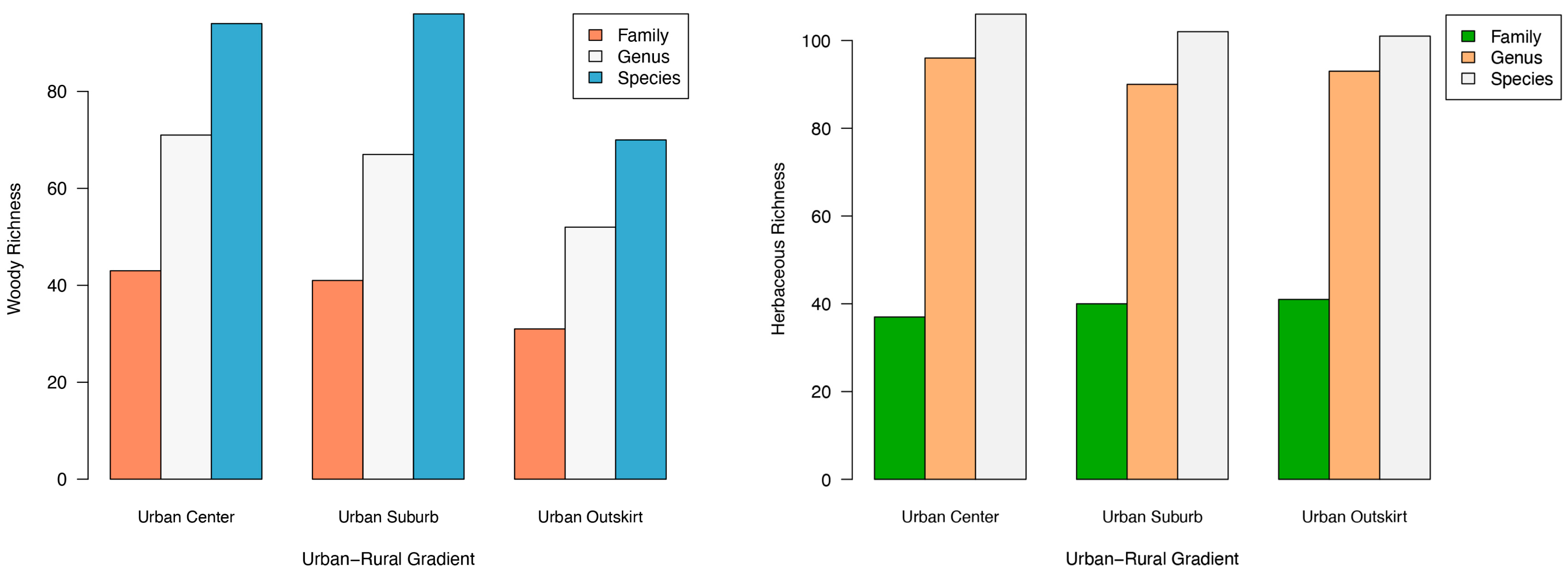
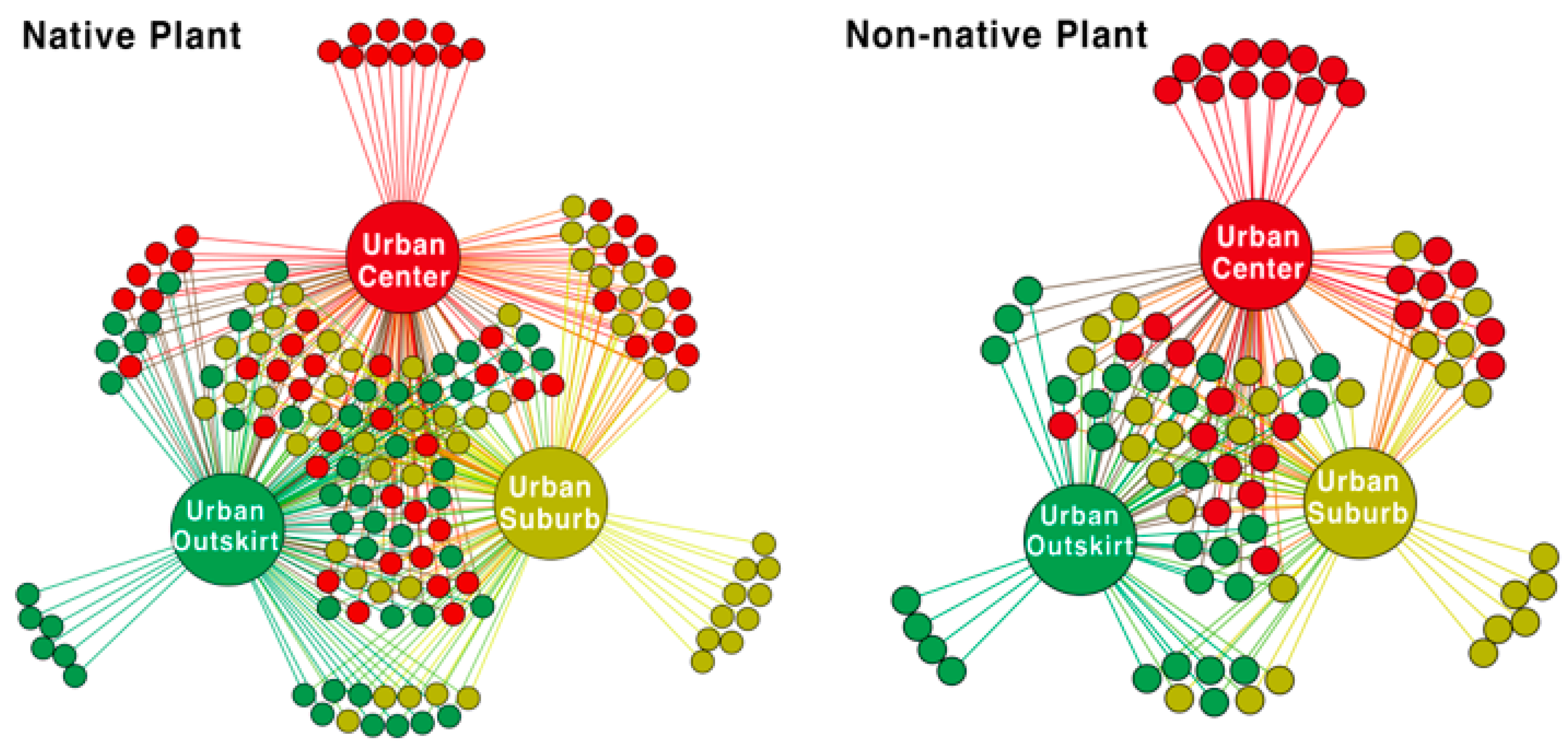
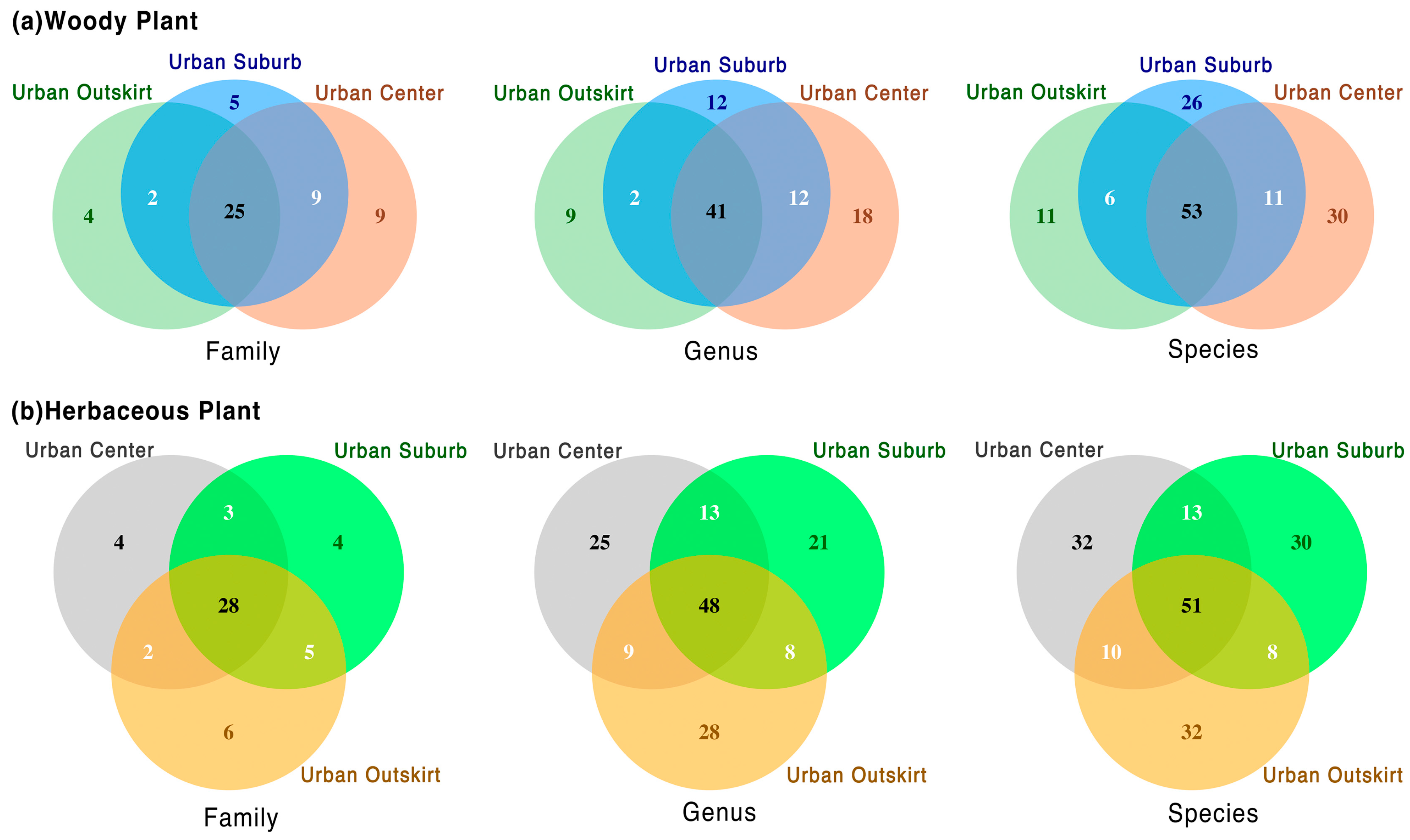
3.1. General Plant Species Composition
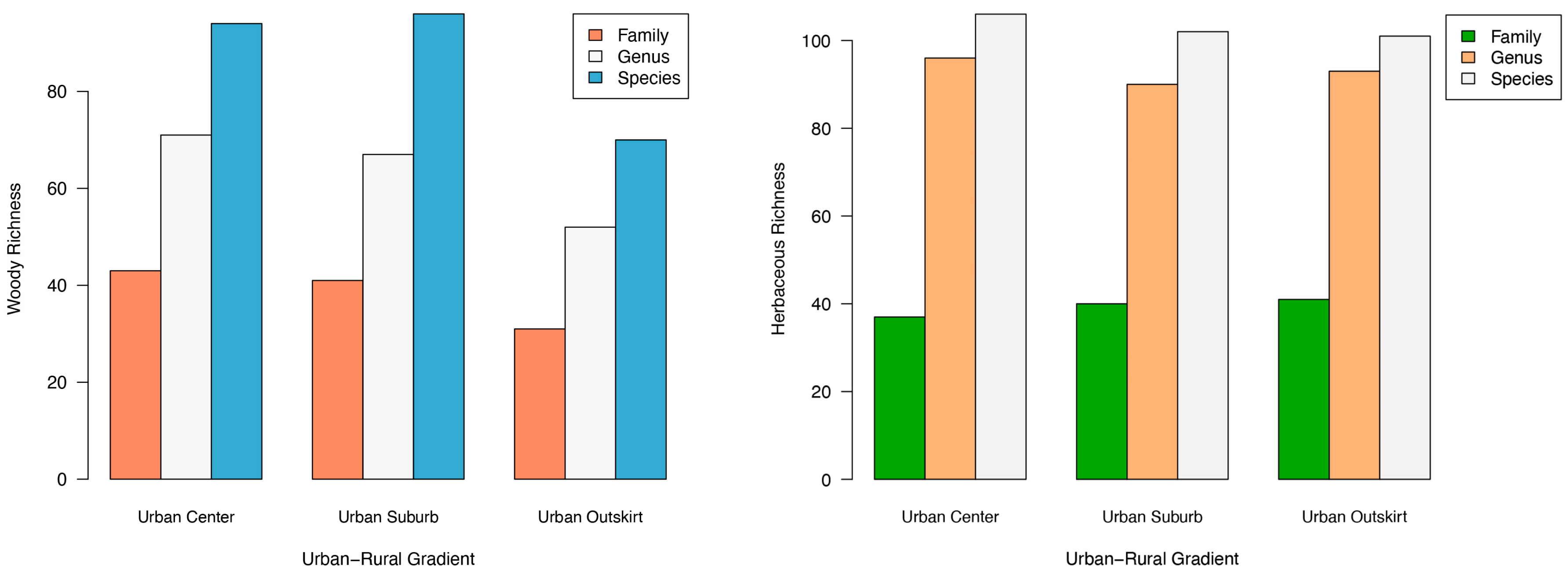
3.2. Different Plant Taxa at Various Gradients of Urbanization
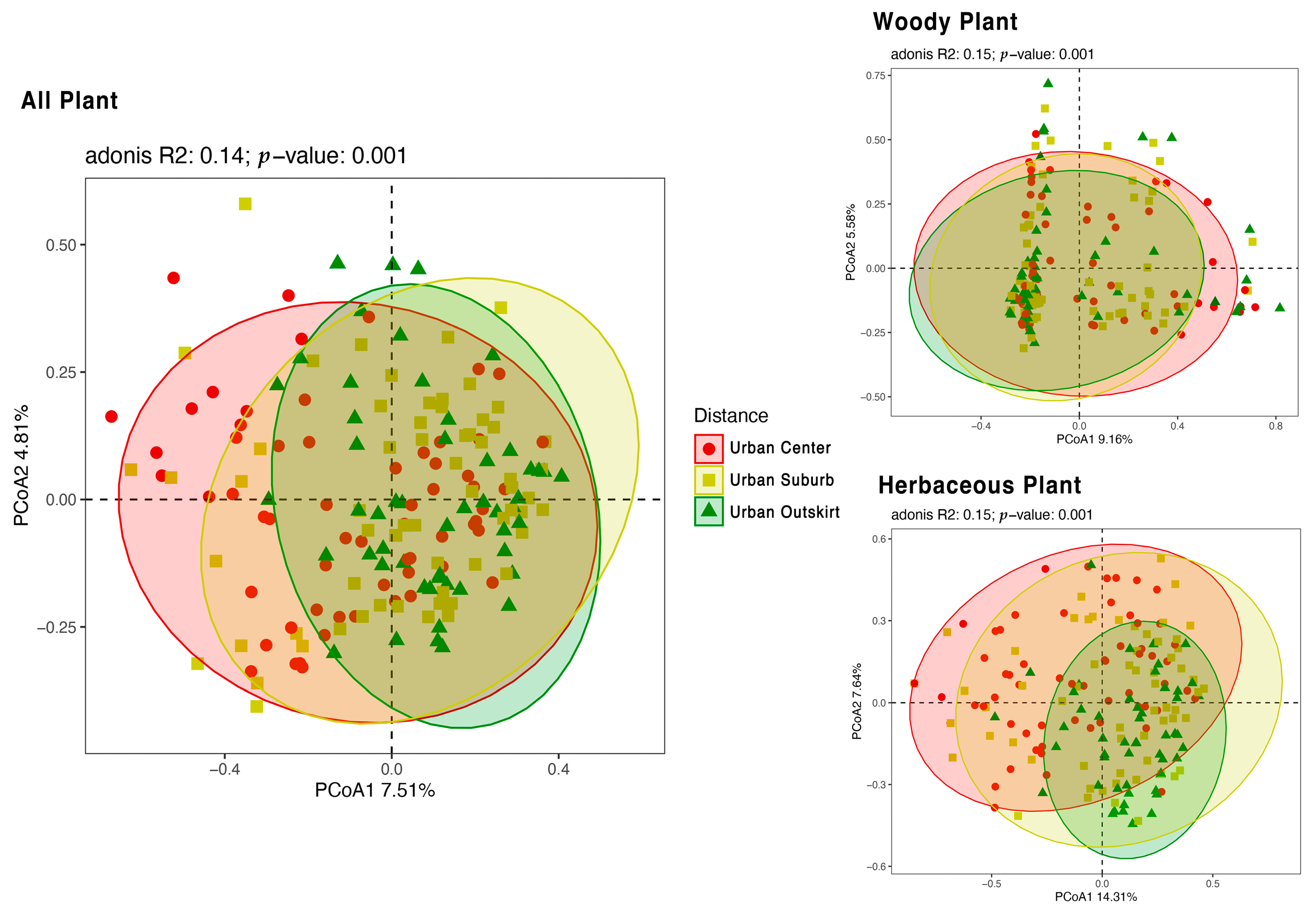
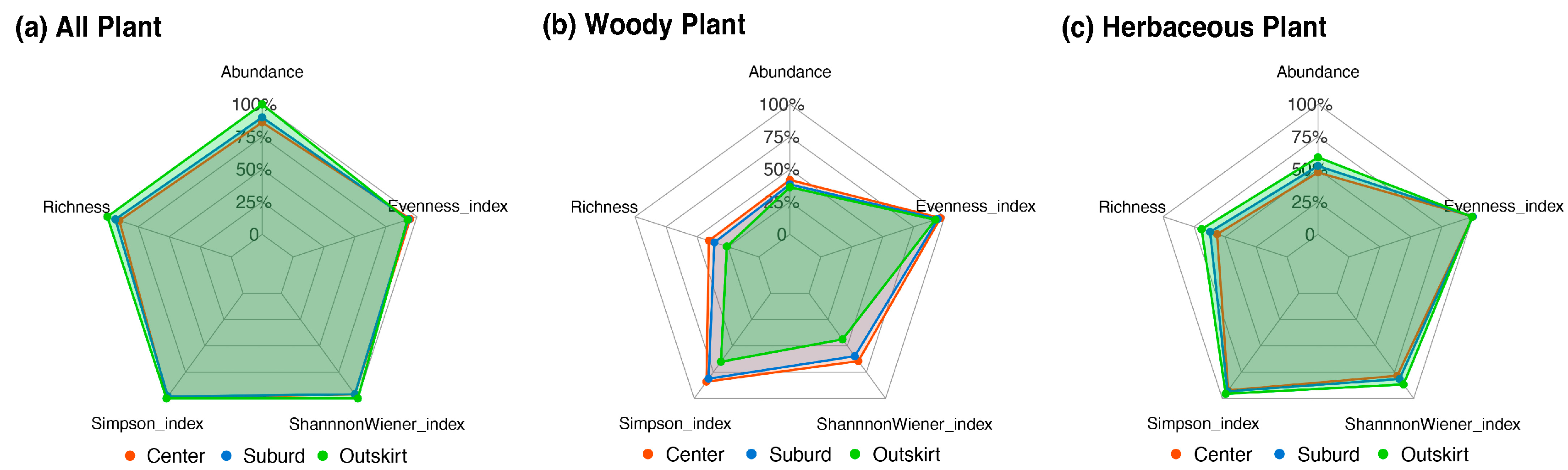
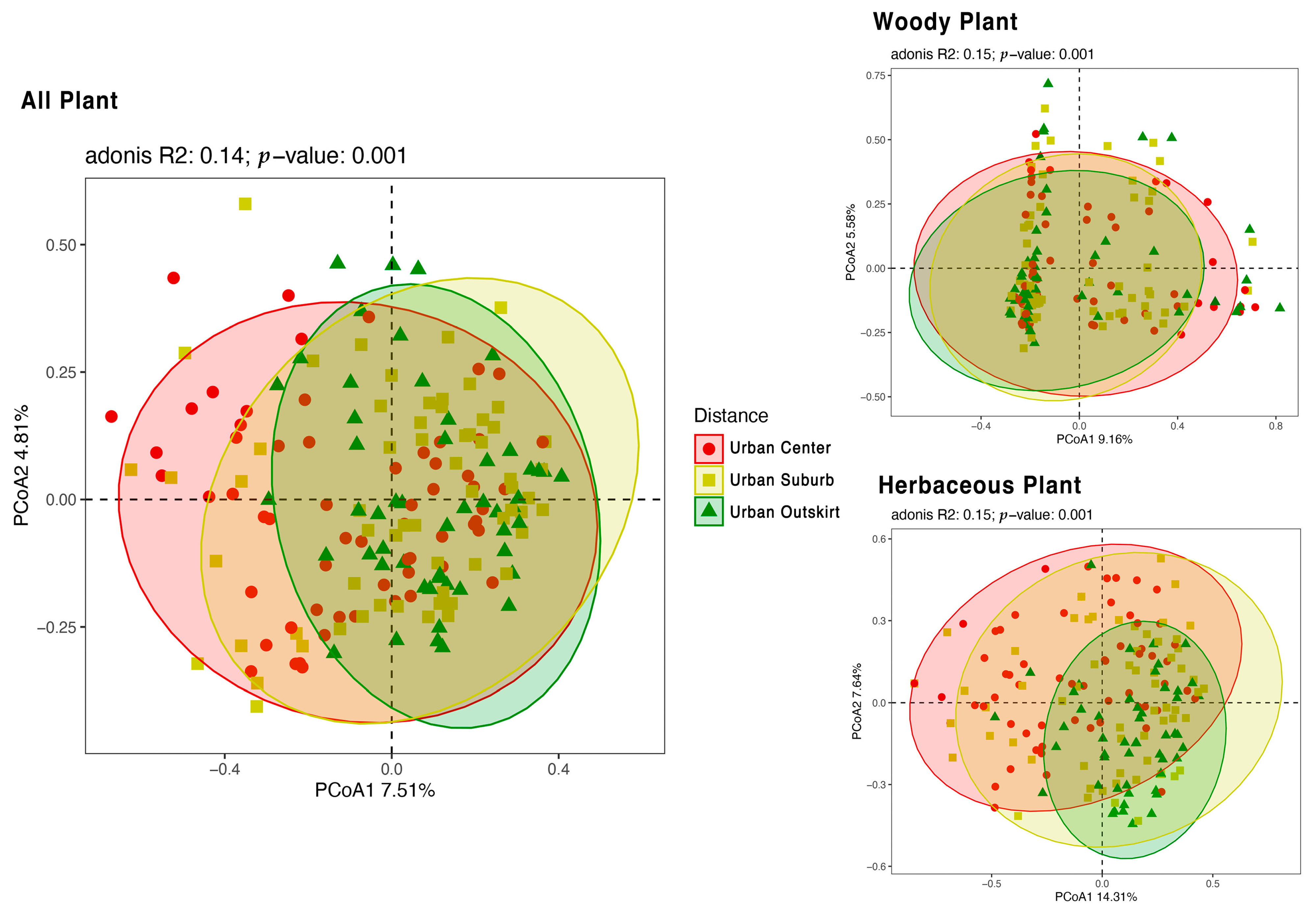
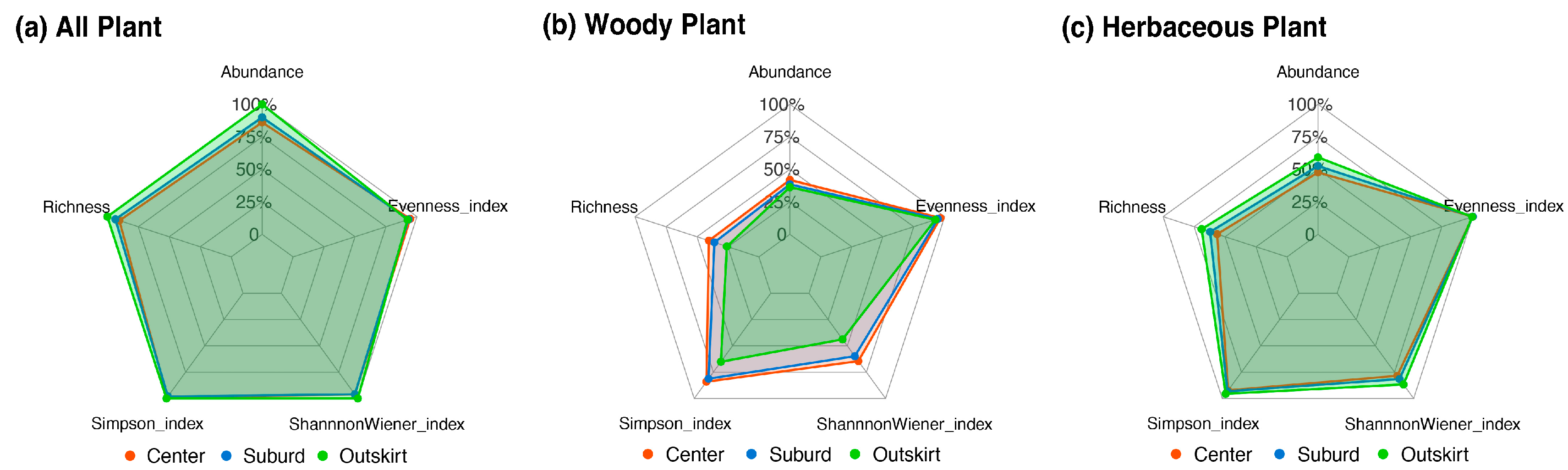
3.3. Different Plant Diversity among Various Gradients of Urbanization
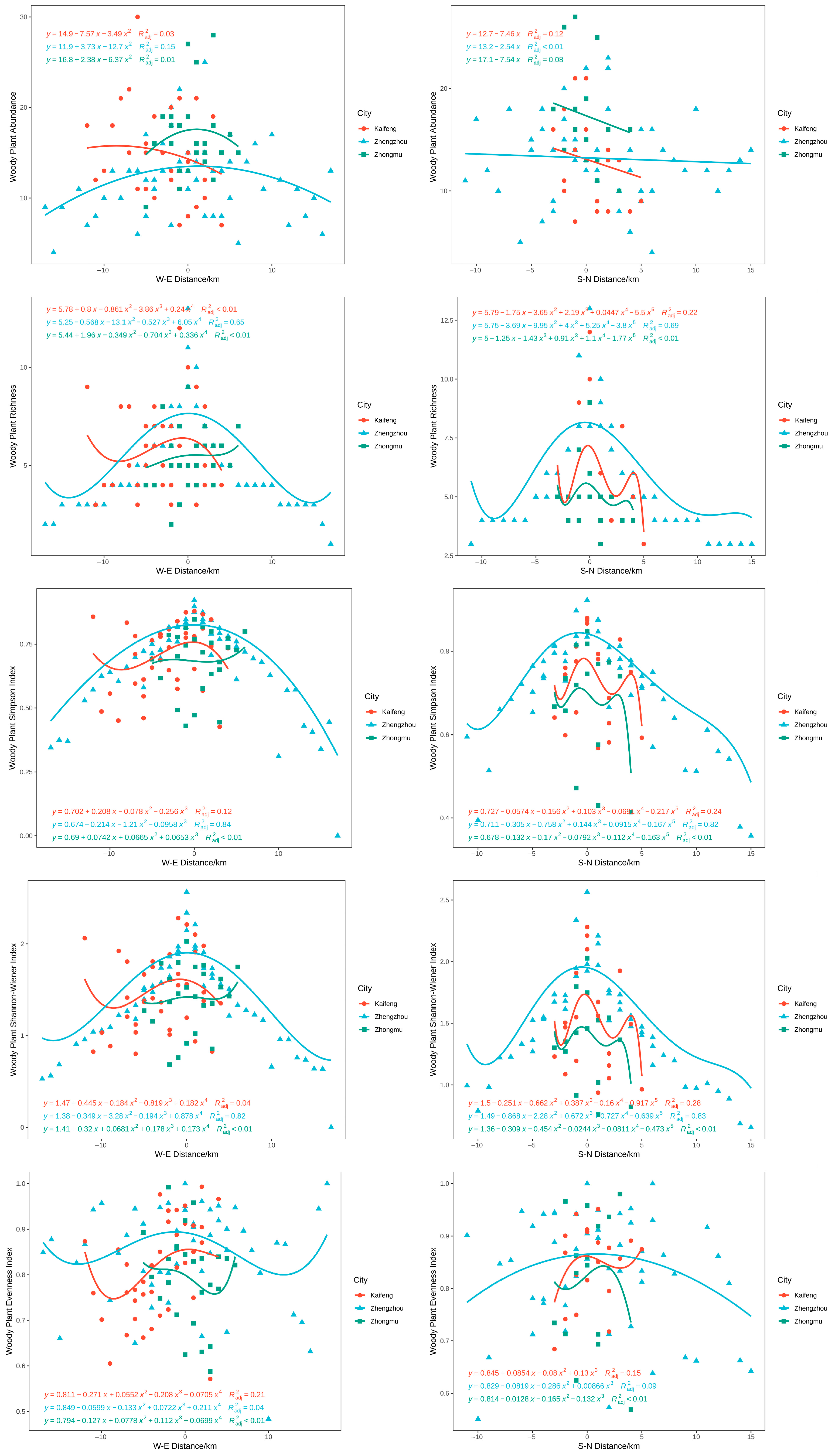
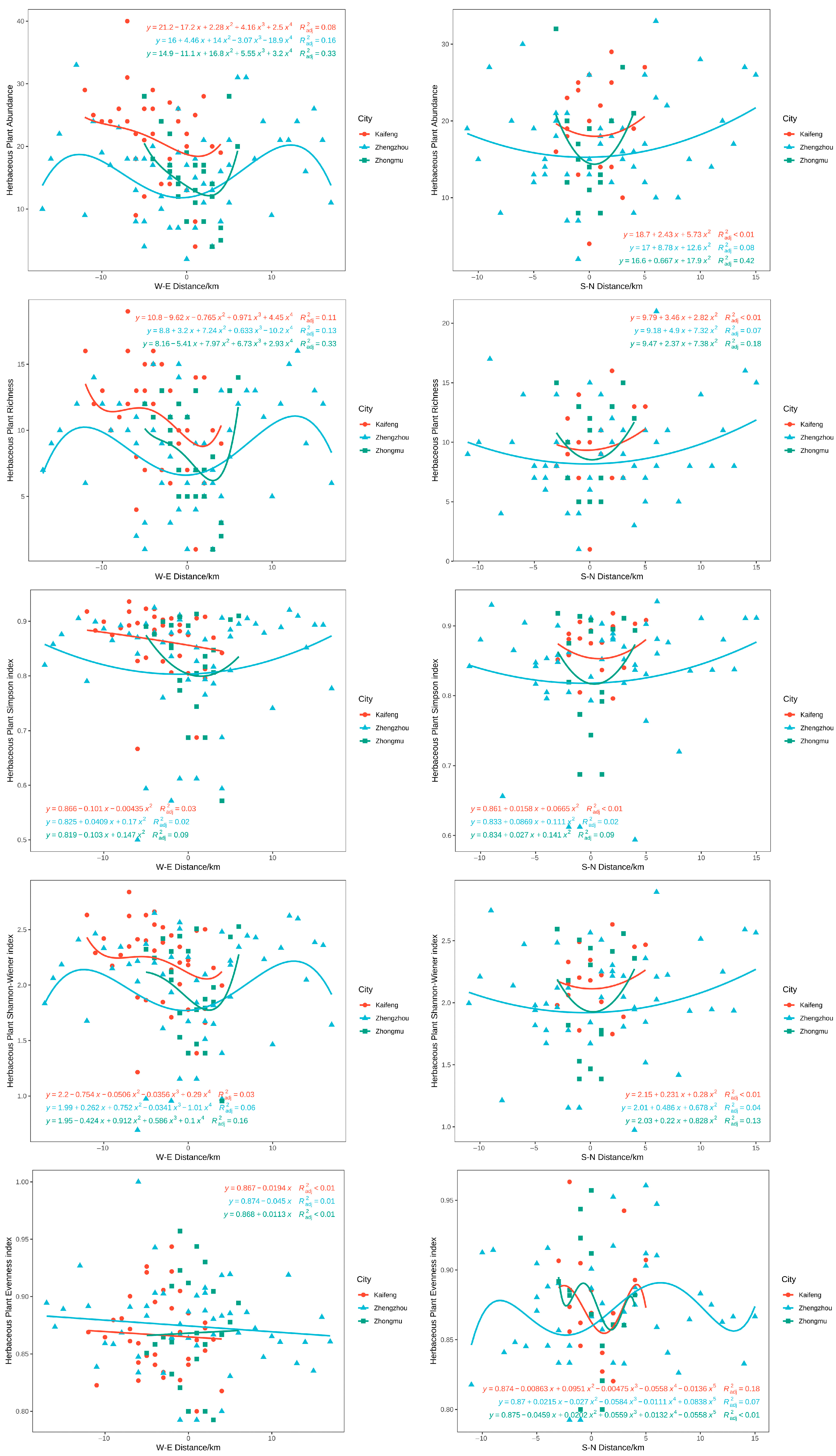
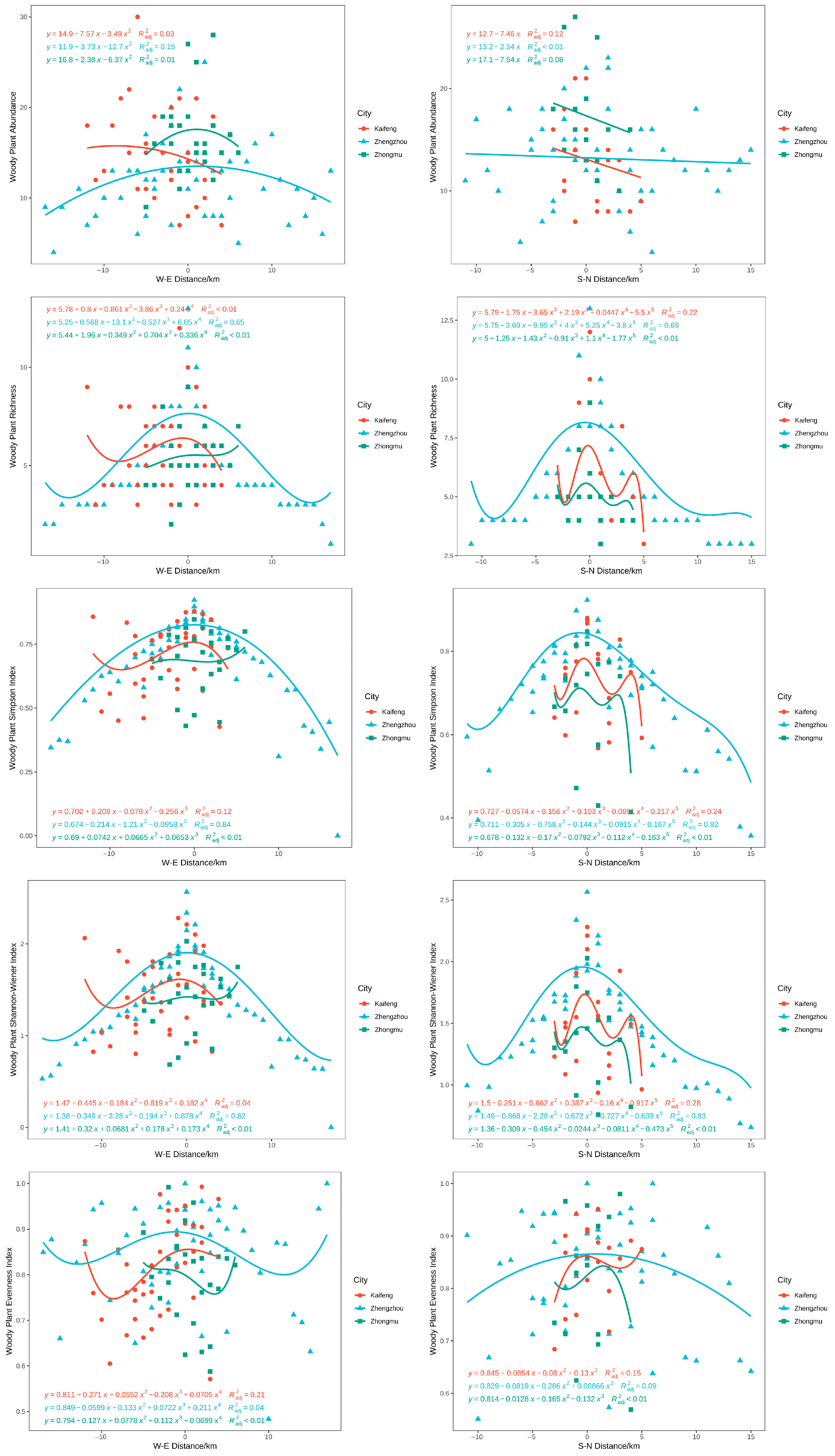
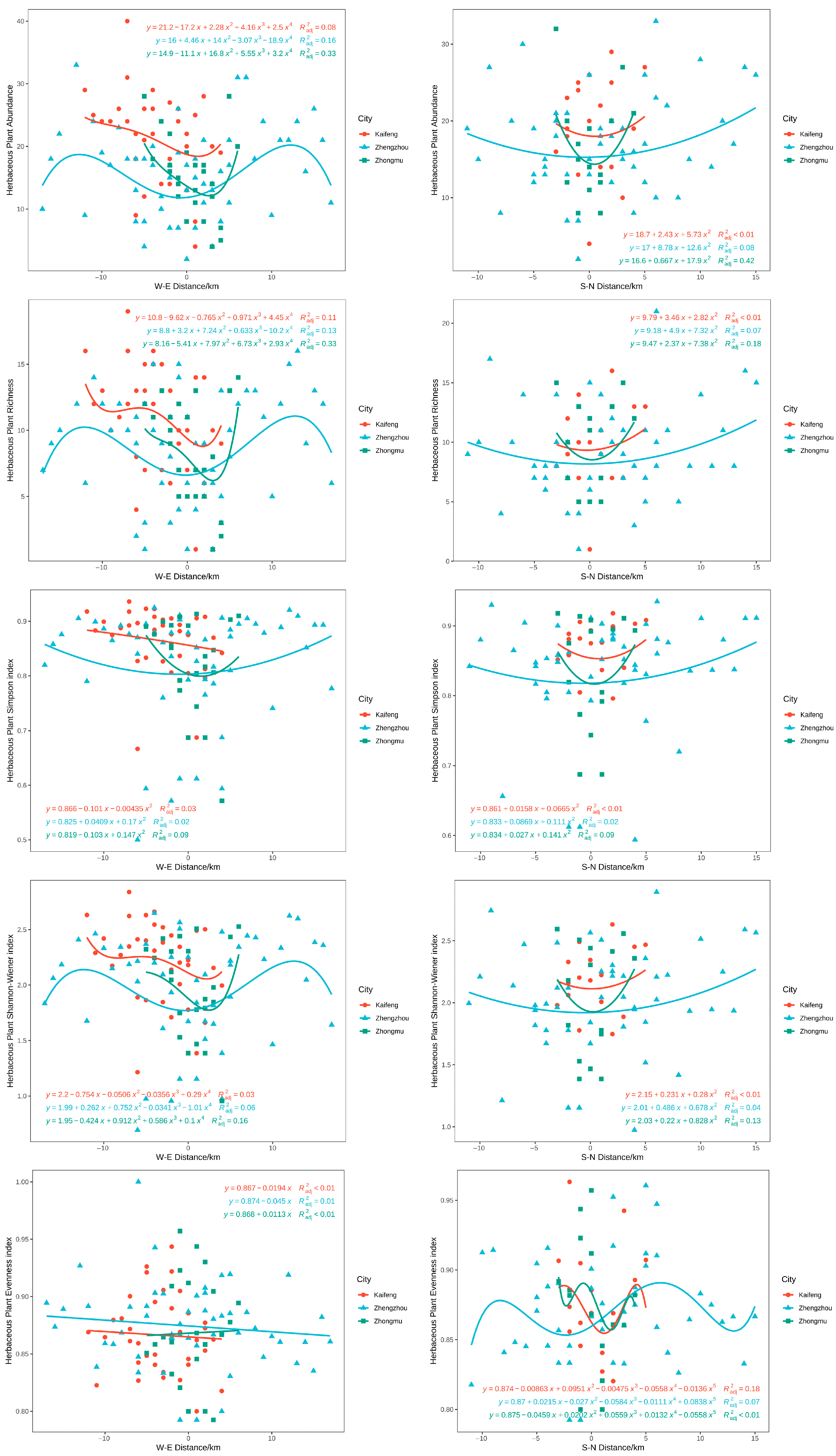
3.4. Distribution Pattern of Plant Diversity in Different Cities
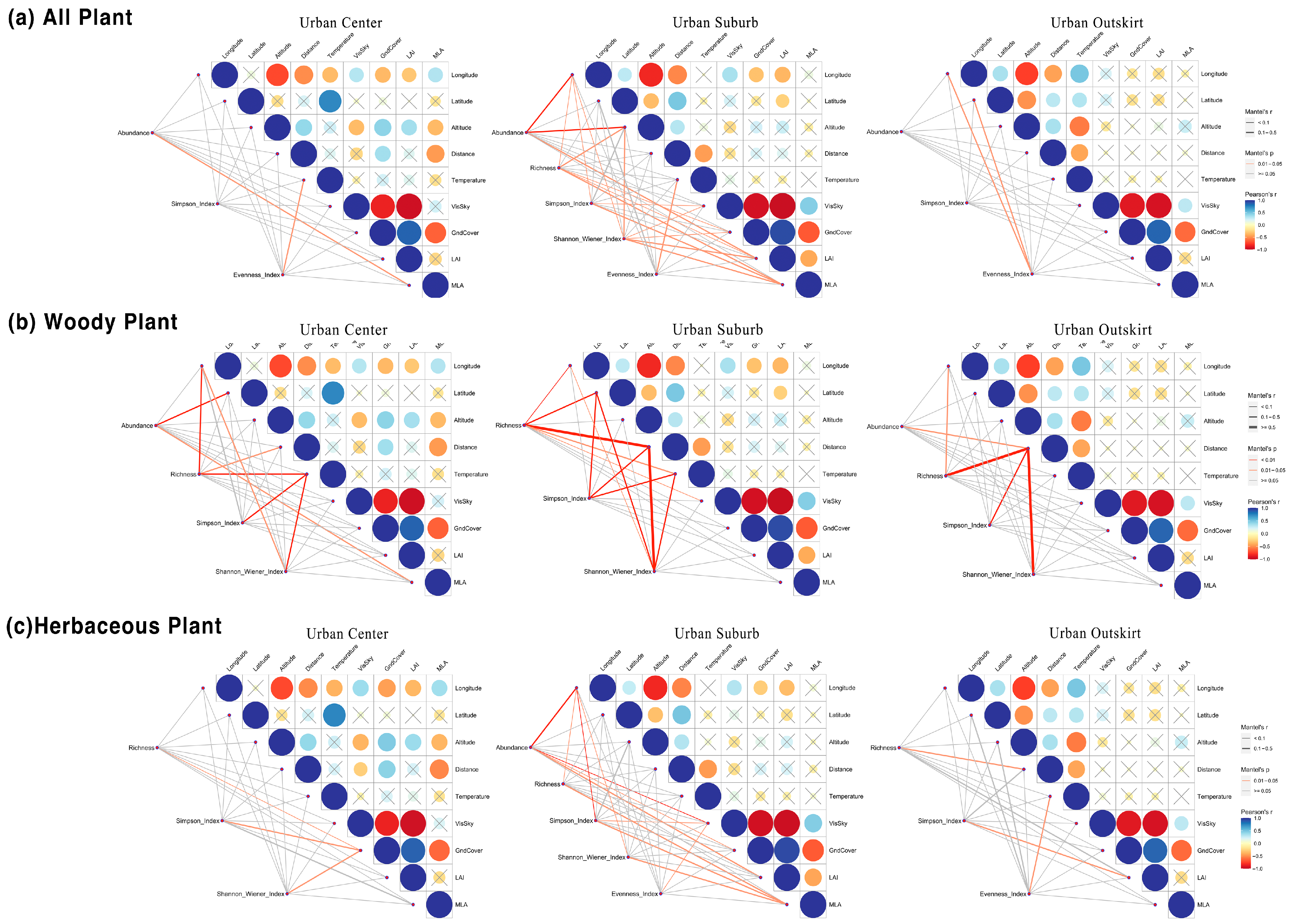
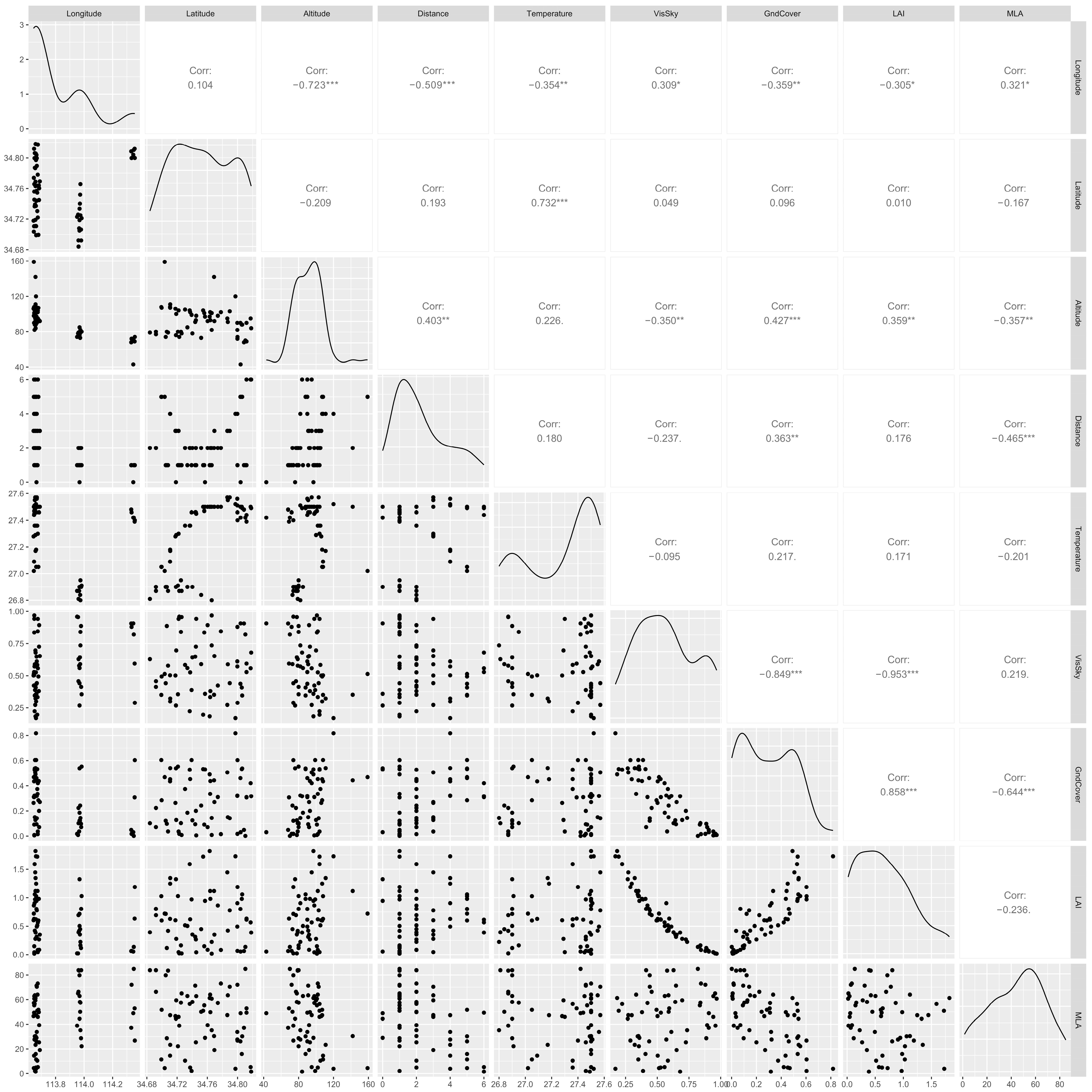
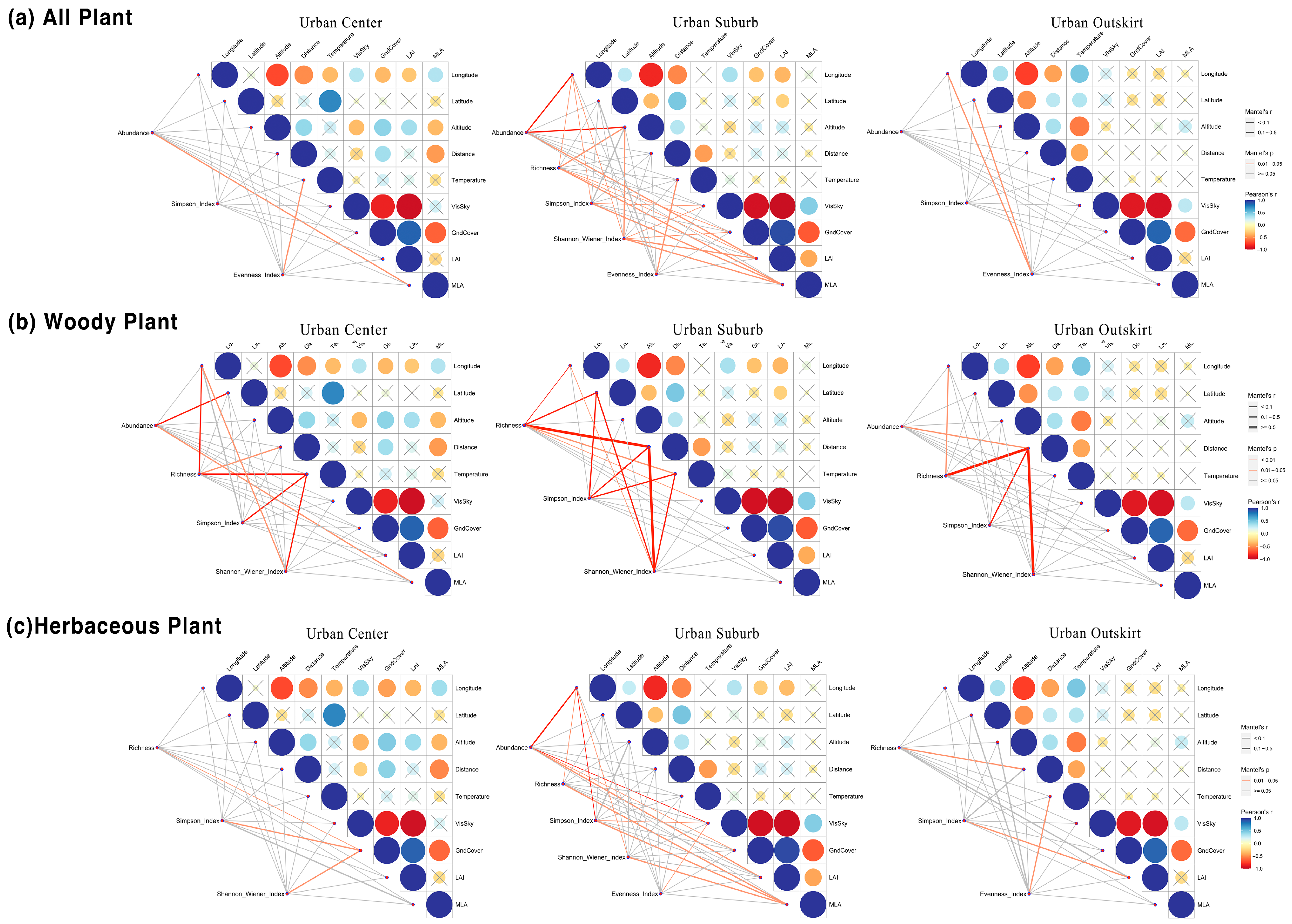
3.5. Effects of Environmental Factors on Plant Diversity under the Urbanization Gradient
4. Discussion
4.1. Plant Species Composition under the Urbanization Gradient
4.2. Multiple Patterns of Plant Species Diversity along the Urban-Rural Gradient
4.3. Effects of Urban Environmental Factors on Plant Diversity
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Díaz, J.G.; Gutiérrez-Bustillo, A.M.; Rojo, J. The phenological response of European vegetation to urbanisation is mediated by macrobioclimatic factors. Sci. Total Environ. 2023, 905, 167092. [Google Scholar] [CrossRef]
- Meng, L.; Mao, J.; Zhou, Y.; Richardson, A.D.; Lee, X.; Thornton, P.E.; Ricciuto, D.M.; Li, X.; Dai, Y.; Shi, X. Urban warming advances spring phenology but reduces the response of phenology to temperature in the conterminous United States. Proc. Natl. Acad. Sci. USA 2020, 117, 4228–4233. [Google Scholar] [CrossRef] [PubMed]
- Aronson, M.F.J.; La Sorte, F.A.; Nilon, C.H.; Katti, M.; Goddard, M.A.; Lepczyk, C.A.; Warren, P.S.; Williams, N.S.G.; Cilliers, S.; Clarkson, B.; et al. A global analysis of the impacts of urbanization on bird and plant diversity reveals key anthropogenic drivers. Proc. R. Soc. B—Biol. Sci. 2014, 281, 20133330. [Google Scholar] [CrossRef]
- La Sorte, F.A.; Aronson, M.F.J.; Williams, N.S.G.; Celesti-Grapow, L.; Cilliers, S.; Clarkson, B.D.; Dolan, R.W.; Hipp, A.; Klotz, S.; Kuehn, I.; et al. Beta diversity of urban floras among European and non-European cities. Glob. Ecol. Biogeogr. 2014, 23, 769–779. [Google Scholar] [CrossRef]
- Zhou, L.M.; Dickinson, R.E.; Tian, Y.H.; Fang, J.Y.; Li, Q.X.; Kaufmann, R.K.; Tucker, C.J.; Myneni, R.B. Evidence for a significant urbanization effect on climate in China. Proc. Natl. Acad. Sci. USA 2004, 101, 9540–9544. [Google Scholar] [CrossRef] [PubMed]
- Wilby, R.L.; Perry, G.L.W. Climate change, biodiversity and the urban environment: A critical review based on London, UK. Prog. Phys. Geogr.—Earth Environ. 2006, 30, 73–98. [Google Scholar] [CrossRef]
- Zhao, S.; Da, L.; Tang, Z.; Fang, H.; Song, K.; Fang, J. Ecological consequences of rapid urban expansion: Shanghai, China. Front. Ecol. Environ. 2006, 4, 341–346. [Google Scholar] [CrossRef]
- Chen, Y.; Zhou, X.; Wang, Z.; Su, X.; Liu, F.; Tian, X.; Ye, Y.; Shao, Y.; Yuan, Z. Cd contamination determined assembly processes and network stability of AM fungal communities in an urban green space ecosystem. Sci. Total Environ. 2023, 899, 166372. [Google Scholar] [CrossRef]
- Jochner, S.; Menzel, A. Urban phenological studies—Past, present, future. Environ. Pollut. 2015, 203, 250–261. [Google Scholar] [CrossRef]
- Niemelä, J.; Breuste, J.H.; Guntenspergen, G.; McIntyre, N.E.; Elmqvist, T.; James, P. Urban Ecology: Patterns, Processes, and Applications; OUP Oxford: Oxford, UK, 2011. [Google Scholar]
- McDonnell, M.J.; Pickett, S.T. Ecosystem structure and function along urban-rural gradients: An unexploited opportunity for ecology. Ecology 1990, 71, 1232–1237. [Google Scholar] [CrossRef]
- McDonnell, M.J.; Hahs, A.K. The use of gradient analysis studies in advancing our understanding of the ecology of urbanizing landscapes: Current status and future directions. Landsc. Ecol. 2008, 23, 1143–1155. [Google Scholar] [CrossRef]
- Burton, M.L.; Samuelson, L.J.; Pan, S. Riparian woody plant diversity and forest structure along an urban-rural gradient. Urban Ecosyst. 2005, 8, 93–106. [Google Scholar] [CrossRef]
- Evans, B.S.; Reitsma, R.; Hurlbert, A.H.; Marra, P.P. Environmental filtering of avian communities along a rural-to-urban gradient in Greater Washington, DC, USA. Ecosphere 2018, 9, e02402. [Google Scholar] [CrossRef]
- Niemela, J.; Kotze, D.J. Carabid beetle assemblages along urban to rural gradients: A review. Landsc. Urban Plan. 2009, 92, 65–71. [Google Scholar] [CrossRef]
- Pickett, S.T.A.; Cadenasso, M.L.; Meiners, S.J. Ever since Clements: From succession to vegetation dynamics and understanding to intervention. Appl. Veg. Sci. 2009, 12, 9–21. [Google Scholar] [CrossRef]
- Zipperer, W.C.; Wu, J.; Pouyat, R.V.; Pickett, S.T. The application of ecological principles to urban and urbanizing landscapes. Ecol. Appl. 2000, 10, 685–688. [Google Scholar] [CrossRef]
- Luck, M.; Wu, J.G. A gradient analysis of urban landscape pattern: A case study from the Phoenix metropolitan region, Arizona, USA. Landsc. Ecol. 2002, 17, 327–339. [Google Scholar] [CrossRef]
- Whittaker, R.H.; Niering, W.A. Vegetation of the Santa Catalina Mountains, Arizona: A gradient analysis of the south slope. Ecology 1965, 46, 429–452. [Google Scholar] [CrossRef]
- Aronson, M.F.J.; Nilon, C.H.; Lepczyk, C.A.; Parker, T.S.; Warren, P.S.; Cilliers, S.S.; Goddard, M.A.; Hahs, A.K.; Herzog, C.; Katti, M.; et al. Hierarchical filters determine community assembly of urban species pools. Ecology 2016, 97, 2952–2963. [Google Scholar] [CrossRef]
- Guenard, G.; Legendre, P. Hierarchical Clustering with Contiguity Constraint in R. J. Stat. Softw. 2022, 103, 1–26. [Google Scholar] [CrossRef]
- Gödde, M.; Richarz, N.; Walter, B. Habitat conservation and development in the city of Düsseldorf, Germany. Urban Ecol. Basis Urban Plan. 1995, 163–171. [Google Scholar]
- Knapp, S.; Kuehn, I.; Schweiger, O.; Klotz, S. Challenging urban species diversity: Contrasting phylogenetic patterns across plant functional groups in Germany. Ecol. Lett. 2008, 11, 1054–1064. [Google Scholar] [CrossRef]
- Kuhn, I.; Brandl, R.; Klotz, S. The flora of German cities is naturally species rich. Evol. Ecol. Res. 2004, 6, 749–764. [Google Scholar]
- Kowarik, I. On the role of alien species in urban flora and vegetation. In Urban Ecology: An International Perspective on the Interaction between Humans and Nature; Springer: Berlin/Heidelberg, Germany, 2008; pp. 321–338. [Google Scholar]
- Ranta, P.; Viljanen, V. Vascular plants along an urban-rural gradient in the city of Tampere, Finland. Urban Ecosyst. 2011, 14, 361–376. [Google Scholar] [CrossRef]
- Stenchly, K.; Lippmann, S.; Waongo, A.; Nyarko, G.; Buerkert, A. Weed species structural and functional composition of okra fields and field periphery under different management intensities along the rural-urban gradient of two West African cities. Agric. Ecosyst. Environ. 2017, 237, 213–223. [Google Scholar] [CrossRef]
- Tian, Z.; Song, K.; Da, L. Distribution patterns and traits of weed communities along an urban-rural gradient under rapid urbanization in Shanghai, China. Weed Biol. Manag. 2015, 15, 27–41. [Google Scholar] [CrossRef]
- Vakhlamova, T.; Rusterholz, H.-P.; Kanibolotskaya, Y.; Baur, B. Changes in plant diversity along an urban-rural gradient in an expanding city in Kazakhstan, Western Siberia. Landsc. Urban Plan. 2014, 132, 111–120. [Google Scholar] [CrossRef]
- Walker, J.S.; Grimm, N.B.; Briggs, J.M.; Gries, C.; Dugan, L. Effects of urbanization on plant species diversity in central Arizona. Front. Ecol. Environ. 2009, 7, 465–470. [Google Scholar] [CrossRef]
- Wang, M.; Li, J.; Kuang, S.; He, Y.; Chen, G.; Huang, Y.; Song, C.; Anderson, P.; Lowicki, D. Plant Diversity along the Urban-Rural Gradient and Its Relationship with Urbanization Degree in Shanghai, China. Forests 2020, 11, 171. [Google Scholar] [CrossRef]
- Cariñanos, P.; Casares-Porcel, M. Urban green zones and related pollen allergy: A review. Some guidelines for designing spaces with low allergy impact. Landsc. Urban Plan. 2011, 101, 205–214. [Google Scholar] [CrossRef]
- Aronson, M.F.J.; Handel, S.N.; La Puma, I.P.; Clemants, S.E. Urbanization promotes non-native woody species and diverse plant assemblages in the New York metropolitan region. Urban Ecosyst. 2015, 18, 31–45. [Google Scholar] [CrossRef]
- Cui, Y.C.; Song, K.; Guo, X.Y.; van Bodegom, P.M.; Pan, Y.J.; Tian, Z.H.; Chen, X.S.; Wang, J.; Da, L.J. Phylogenetic and functional structures of plant communities along a spatiotemporal urbanization gradient: Effects of colonization and extinction. J. Veg. Sci. 2019, 30, 341–351. [Google Scholar] [CrossRef]
- Yang, Y.; Fujihara, M.; Li, B.; Yuan, X.; Hara, K.; Da, L.; Tomita, M.; Zhao, Y. Structure and diversity of remnant natural evergreen broad-leaved forests at three sites affected by urbanization in Chongqing metropolis, Southwest China. Landsc. Ecol. Eng. 2014, 10, 137–149. [Google Scholar] [CrossRef]
- Grimm, N.B.; Faeth, S.H.; Golubiewski, N.E.; Redman, C.L.; Wu, J.; Bai, X.; Briggs, J.M. Global change and the ecology of cities. Science 2008, 319, 756–760. [Google Scholar] [CrossRef] [PubMed]
- Lopez, B.E.; Urban, D.; White, P.S. Testing the effects of four urbanization filters on forest plant taxonomic, functional, and phylogenetic diversity. Ecol. Appl. 2018, 28, 2197–2205. [Google Scholar] [CrossRef] [PubMed]
- Palma, E.; Catford, J.A.; Corlett, R.T.; Duncan, R.P.; Hahs, A.K.; McCarthy, M.A.; McDonnell, M.J.; Thompson, K.; Williams, N.S.G.; Vesk, P.A. Functional trait changes in the floras of 11 cities across the globe in response to urbanization. Ecography 2017, 40, 875–886. [Google Scholar] [CrossRef]
- Ramalho, C.E.; Laliberte, E.; Poot, P.; Hobbs, R.J. Complex effects of fragmentation on remnant woodland plant communities of a rapidly urbanizing biodiversity hotspot. Ecology 2014, 95, 2466–2478. [Google Scholar] [CrossRef]
- Chalker-Scott, L. Nonnative, noninvasive woody species can enhance urban landscape biodiversity. Arboric. Urban For. 2015, 41, 173–186. [Google Scholar] [CrossRef]
- Helden, A.J.; Stamp, G.C.; Leather, S.R. Urban biodiversity: Comparison of insect assemblages on native and non-native trees. Urban Ecosyst. 2012, 15, 611–624. [Google Scholar] [CrossRef]
- Liu, D.; Semenchuk, P.; Essl, F.; Lenzner, B.; Moser, D.; Blackburn, T.M.; Cassey, P.; Biancolini, D.; Capinha, C.; Dawson, W. The impact of land use on non-native species incidence and number in local assemblages worldwide. Nat. Commun. 2023, 14, 2090. [Google Scholar] [CrossRef]
- Lin, J.; Wang, Q.; Li, X. Socioeconomic and spatial inequalities of street tree abundance, species diversity, and size structure in New York City. Landsc. Urban Plan. 2021, 206, 103992. [Google Scholar] [CrossRef]
- Peng, Y.; Mi, K.; Wang, H.; Liu, Z.; Lin, Y.; Sang, W.; Cui, Q. Most suitable landscape patterns to preserve indigenous plant diversity affected by increasing urbanization: A case study of Shunyi District of Beijing, China. Urban For. Urban Green. 2019, 38, 33–41. [Google Scholar] [CrossRef]
- Cao, Y.; Natuhara, Y. Effect of urbanization on vegetation in riparian area: Plant communities in artificial and semi-natural habitats. Sustainability 2019, 12, 204. [Google Scholar] [CrossRef]
- Alue, B.A.; Salleh Hudin, N.; Mohamed, F.; Mat Said, Z.; Ismail, K. Plant Diversity along an Urbanization Gradient of a Tropical City. Diversity 2022, 14, 1024. [Google Scholar] [CrossRef]
- Rebelo, A.; Holmes, P.; Dorse, C.; Wood, J. Impacts of urbanization in a biodiversity hotspot: Conservation challenges in Metropolitan Cape Town. S. Afr. J. Bot. 2011, 77, 20–35. [Google Scholar] [CrossRef]
- Marcacci, G.; Westphal, C.; Rao, V.S.; Kumar, S.S.; Tharini, K.; Belavadi, V.V.; Nölke, N.; Tscharntke, T.; Grass, I. Urbanization alters the spatiotemporal dynamics of plant–pollinator networks in a tropical megacity. Ecol. Lett. 2023, 26, 1951–1962. [Google Scholar] [CrossRef] [PubMed]
- Muratet, A.; Porcher, E.; Devictor, V.; Arnal, G.; Moret, J.; Wright, S.; Machon, N. Evaluation of floristic diversity in urban areas as a basis for habitat management. Appl. Veg. Sci. 2008, 11, 451–460. [Google Scholar] [CrossRef]
- Benayas, R.; José, M.; Scheiner, S.M. Plant diversity, biogeography and environment in Iberia: Patterns and possible causal factors. J. Veg. Sci. 2002, 13, 245–258. [Google Scholar] [CrossRef]
- Chesson, P. Mechanisms of maintenance of species diversity. Annu. Rev. Ecol. Syst. 2000, 31, 343–366. [Google Scholar] [CrossRef]
- Hale, S.E.; Edwards, C. Comparison of film and digital hemispherical photography across a wide range of canopy densities. Agric. For. Meteorol. 2002, 112, 51–56. [Google Scholar] [CrossRef]
- Oksanen, J.; Blanchet, F.; Kindt, R.; Legendre, P.; Minchin, P.; O’Hara, R.; Simpson, G.; Solymos, P.; Stevens, M.; Wagner, H. Vegan: Community Ecology Package. Available online: https://cran.r-project.org/web/packages/vegan/vegan.pdf (accessed on 8 June 2022).
- Villanueva, R.A.M.; Chen, Z.J. ggplot2: Elegant Graphics for Data Analysis; Taylor & Francis: Abingdon, UK, 2019. [Google Scholar]
- Hollister, E.B.; Engledow, A.S.; Hammett, A.J.M.; Provin, T.L.; Wilkinson, H.H.; Gentry, T.J. Shifts in microbial community structure along an ecological gradient of hypersaline soils and sediments. ISME J. 2010, 4, 829–838. [Google Scholar] [CrossRef]
- Wang, H.-F.; MacGregor-Fors, I.; Lopez-Pujol, J. Warm-temperate, immense, and sprawling: Plant diversity drivers in urban Beijing, China. Plant Ecol. 2012, 213, 967–992. [Google Scholar] [CrossRef]
- Capuana, M. A review of the performance of woody and herbaceous ornamental plants for phytoremediation in urban areas. Iforest-Biogeosci. For. 2020, 13, 139–151. [Google Scholar] [CrossRef]
- Gregg, J.W.; Jones, C.G.; Dawson, T.E. Urbanization effects on tree growth in the vicinity of New York City. Nature 2003, 424, 183–187. [Google Scholar] [CrossRef]
- Cochard, A.; Pithon, J.; Braud, F.; Beaujouan, V.; Bulot, A.; Daniel, H. Intraspecific trait variation in grassland plant communities along urban-rural gradients. Urban Ecosyst. 2019, 22, 583–591. [Google Scholar] [CrossRef]
- Yang, J.; Luo, X.; Lu, S.; Yang, Y.; Yang, J. Effects of compositional and configurational heterogeneity of the urban matrix on the species richness of woody plants in urban remnant forest patches. Landsc. Ecol. 2022, 37, 619–632. [Google Scholar] [CrossRef]
- Coban, S.; Yener, S.D.; Bayraktar, S. Woody plant composition and diversity of urban green spaces in Istanbul, Turkey. Plant Biosyst. 2021, 155, 83–91. [Google Scholar] [CrossRef]
- Cochard, A.; Pithon, J.; Jagaille, M.; Beaujouan, V.; Pain, G.; Daniel, H. Grassland plant species occurring in extensively managed road verges are filtered by urban environments. Plant Ecol. Divers. 2017, 10, 217–229. [Google Scholar] [CrossRef]
- Ramalho, C.E.; Laliberte, E.; Poot, P.; Hobbs, R. Effects of fragmentation on the plant functional composition and diversity of remnant woodlands in a young and rapidly expanding city. J. Veg. Sci. 2018, 29, 285–296. [Google Scholar] [CrossRef]
- Concepción, E.D.; Obrist, M.K.; Moretti, M.; Altermatt, F.; Baur, B.; Nobis, M.P. Impacts of urban sprawl on species richness of plants, butterflies, gastropods and birds: Not only built-up area matters. Urban Ecosyst. 2015, 19, 225–242. [Google Scholar] [CrossRef]
- Chang, C.-R.; Chen, M.-C.; Su, M.-H. Natural versus human drivers of plant diversity in urban parks and the anthropogenic species-area hypotheses. Landsc. Urban Plan. 2021, 208, 104023. [Google Scholar] [CrossRef]
- Bayón, Á.; Godoy, O.; Maurel, N.; van Kleunen, M.; Vilà, M. Proportion of non-native plants in urban parks correlates with climate, socioeconomic factors and plant traits. Urban For. Urban Green. 2021, 63, 127215. [Google Scholar] [CrossRef]
- Li, D.; Barve, N.; Brenskelle, L.; Earl, K.; Barve, V.; Belitz, M.W.; Doby, J.; Hantak, M.M.; Oswald, J.A.; Stucky, B.J.; et al. Climate, urbanization, and species traits interactively drive flowering duration. Glob. Chang. Biol. 2021, 27, 892–903. [Google Scholar] [CrossRef] [PubMed]
- Arruda, L.J.; Ranieri, B.D.; Cheib, A.L.; Morellato, L.P.C.; Fernandes, G.W.; Negreiros, D. Phenological behavior of herbaceous and woody species in the highly threatened Ironstone Rupestrian Grasslands. S. Afr. J. Bot. 2021, 140, 135–142. [Google Scholar] [CrossRef]
- Zhou, W.; Huang, G.; Pickett, S.T.A.; Wang, J.; Cadenasso, M.L.; McPhearson, T.; Grove, J.M.; Wang, J. Urban tree canopy has greater cooling effects in socially vulnerable communities in the US. One Earth 2021, 4, 1764–1775. [Google Scholar] [CrossRef]
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Qi, R.; Zhou, X.; Li, Z.; Ye, Y.; Yuan, Z.; Liu, F.; Shao, Y.; Wei, D.; Chen, Y. Plant Diversity Distribution along an Urbanization Gradient and Relationships with Environmental Factors in Urban Agglomerations of Henan Province, China. Diversity 2024, 16, 53. https://doi.org/10.3390/d16010053
Qi R, Zhou X, Li Z, Ye Y, Yuan Z, Liu F, Shao Y, Wei D, Chen Y. Plant Diversity Distribution along an Urbanization Gradient and Relationships with Environmental Factors in Urban Agglomerations of Henan Province, China. Diversity. 2024; 16(1):53. https://doi.org/10.3390/d16010053
Chicago/Turabian StyleQi, Rui, Xiayan Zhou, Zihao Li, Yongzhong Ye, Zhiliang Yuan, Fengqin Liu, Yizhen Shao, Dongwei Wei, and Yun Chen. 2024. "Plant Diversity Distribution along an Urbanization Gradient and Relationships with Environmental Factors in Urban Agglomerations of Henan Province, China" Diversity 16, no. 1: 53. https://doi.org/10.3390/d16010053
APA StyleQi, R., Zhou, X., Li, Z., Ye, Y., Yuan, Z., Liu, F., Shao, Y., Wei, D., & Chen, Y. (2024). Plant Diversity Distribution along an Urbanization Gradient and Relationships with Environmental Factors in Urban Agglomerations of Henan Province, China. Diversity, 16(1), 53. https://doi.org/10.3390/d16010053