
Pleistocene Aquatic Refuges Support the East–West Separation of the Neotropical Catfish Trichomycterinae (Siluriformes: Trichomycteridae) and High Diversity in the Magdalena, Guiana, and Paraná-Paraguay Basins


Abstract
1. Introduction
2. Materials and Methods
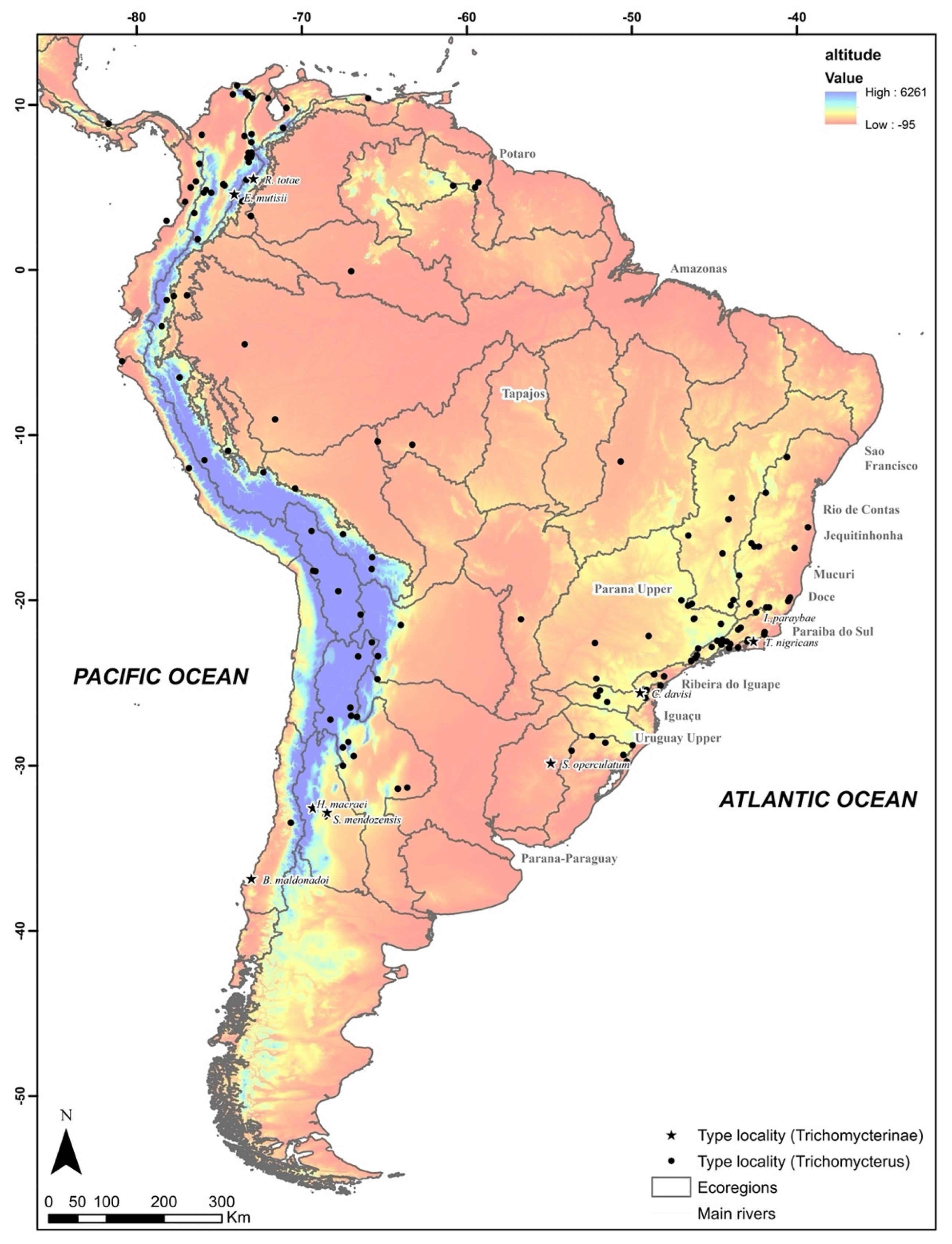
2.1. Mapping Patterns of Species Distribution
2.2. Sampling
2.3. Sequencing and Phylogenetic Analysis
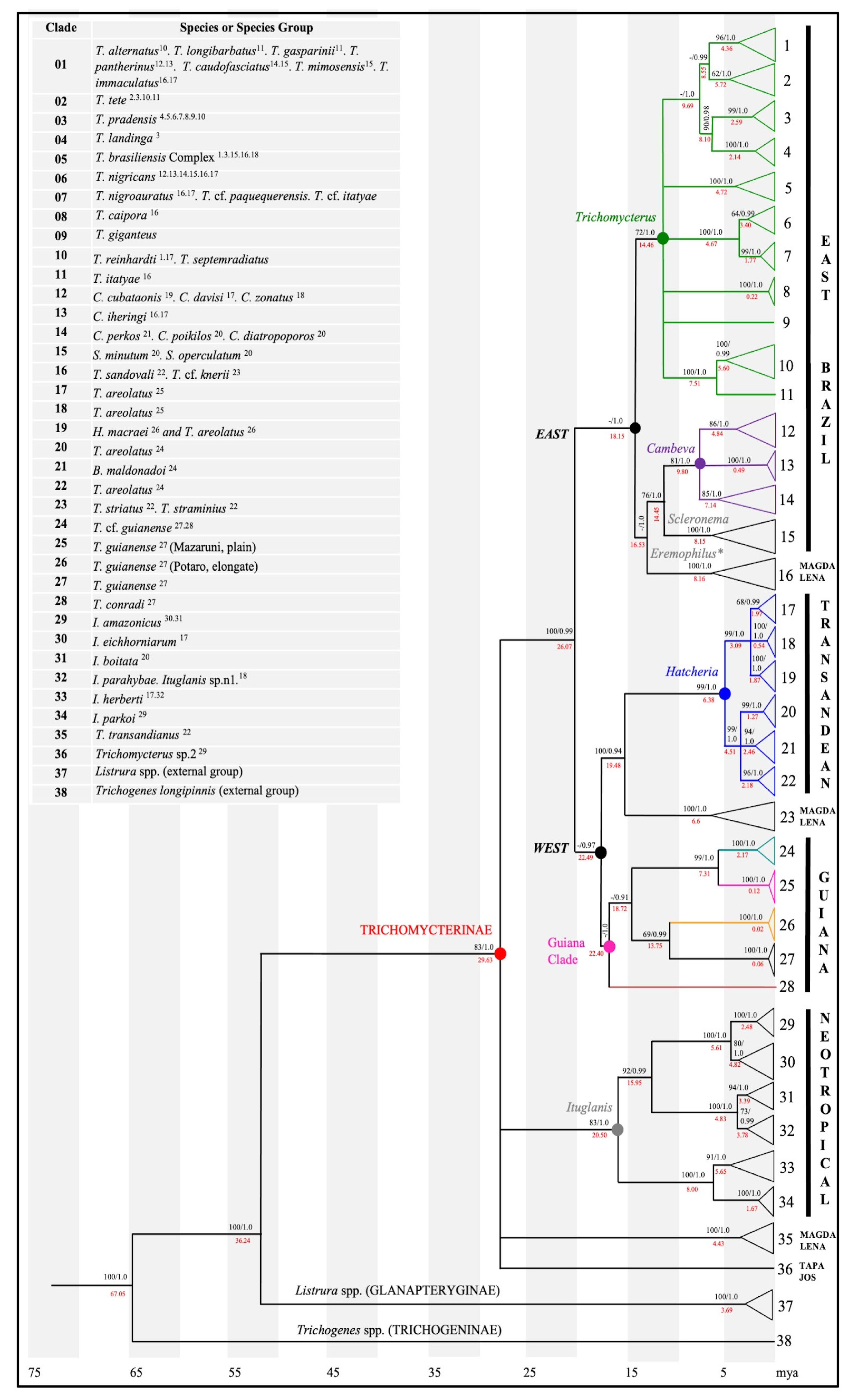
3. Results
3.1. Relationships among Major Clades
3.2. East Major Clade
3.2.1. Trichomycterus Sensu Stricto—Clade A
3.2.2. Cambeva—Clade B
3.2.3. Scleronema—Clade C
3.2.4. Eremophilus—Clade D
3.3. West Major Clade
Hatcheria—Clade E
3.4. Striatus—Clade F
3.5. Guiana—Clade G
3.6. Ituglanis—Clade H
3.7. Transandianus—Clade I
3.8. Tapajós—Clade J
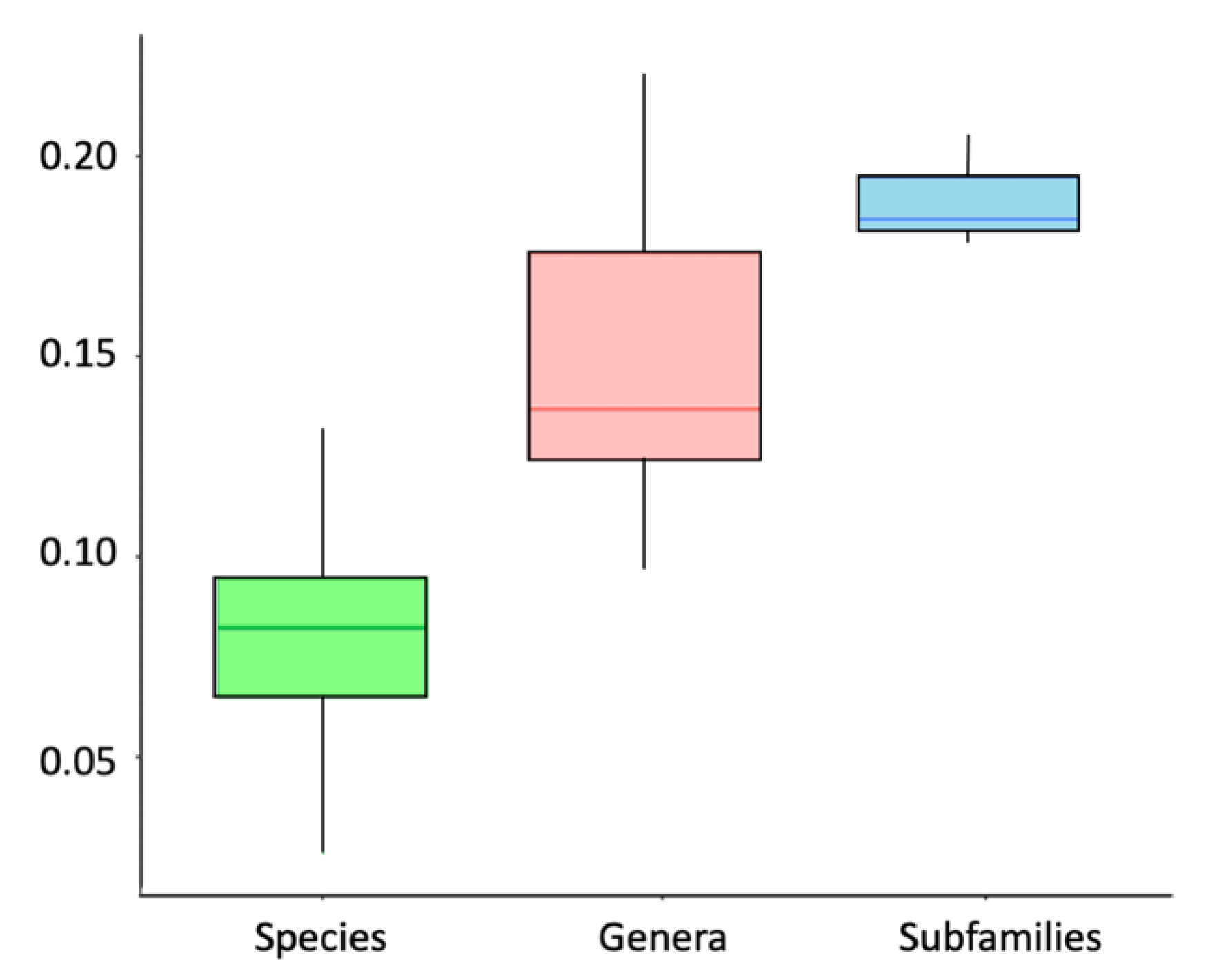
3.9. Referential Indices of Molecular Divergence for Trichomycterinae
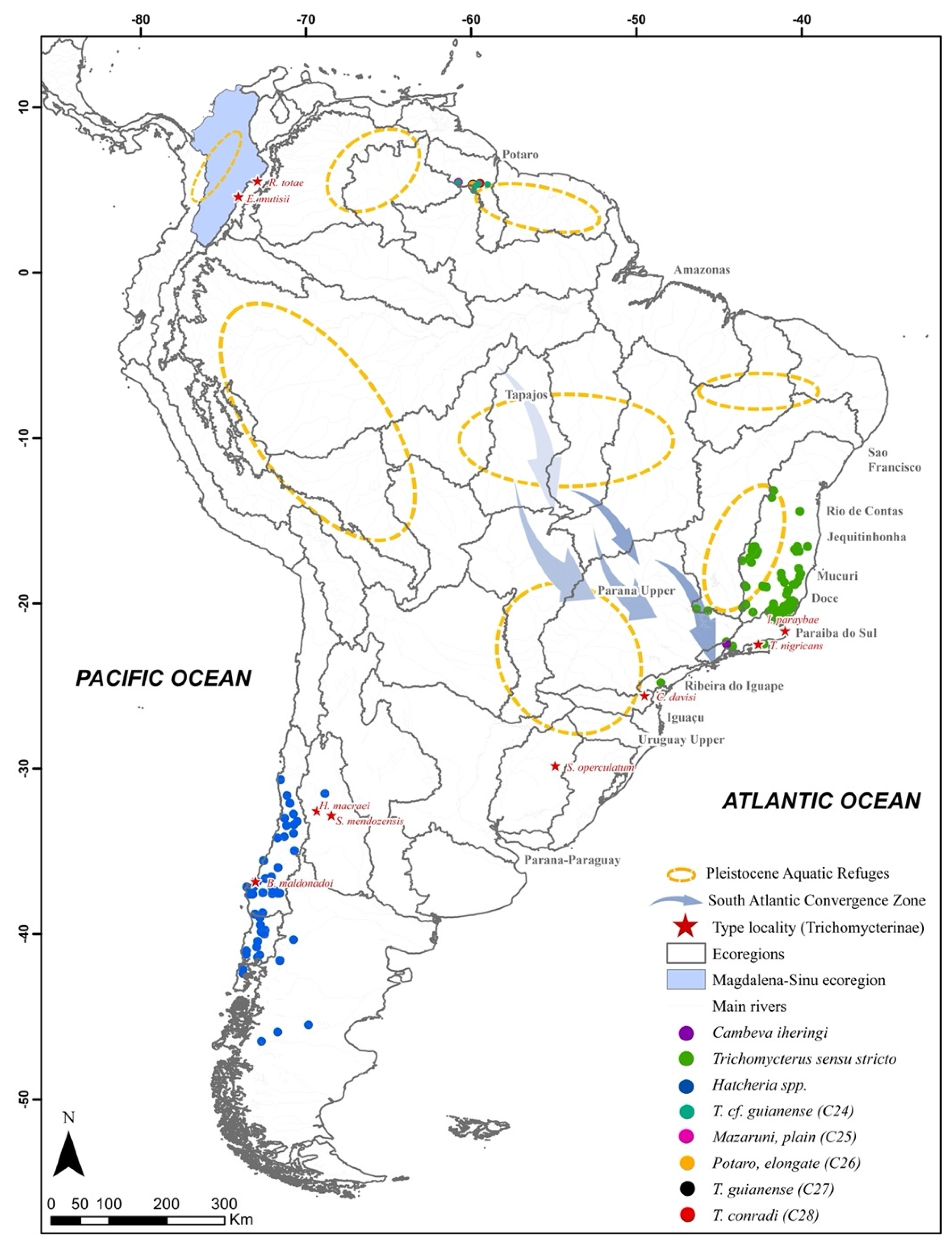
4. Discussion
4.1. The Major Clades in the Trichomycterinae
4.1.1. Clade Trichomycterus Sensu Stricto
4.1.2. Clades Cambeva and Scleronema
4.1.3. Clade Eremophilus
4.1.4. Clade Hatcheria
4.1.5. Clade Striatus
4.1.6. Clade Guiana
4.1.7. Clade Ituglanis
4.1.8. Clade Transandianus
4.1.9. Clade Tapajós
4.2. Efficiency of Cytochrome b
4.3. The Magdalena–Sinu Diversity
4.4. The Cladogenesis Processes under a Phylogeographic Perspective
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- FishBase. Available online: https://www.fishbase.org (accessed on 9 May 2023).
- De Pinna, M.C.C. Phylogenetic relationships of Neotropical Siluriformes (Teleostei: Ostariophysi); historical overview and synthesis of hypothesis. In Phylogeny and Classification of Neotropical Fishes; Malabarba, L.R., Reis, R.E., Vari, R.P., Lucena, Z.M., Lucena, C.A.S., Eds.; Edipucrs: Porto Alegre, Brazil, 1998; pp. 279–330. [Google Scholar]
- De Pinna, M.C.C.; Wosiacki, W.B. Family Trichomycteridae (pencil or parasitic catfishes). In Check List of the Freshwater Fishes of South and Central America; Reis, R.E., Kullander, S.O., Ferraris, C.J., Jr., Eds.; Edipucrs: Porto Alegre, Brazil, 2003; pp. 270–290. [Google Scholar]
- Eschmeyer’s Catalog of Fishes: Genera, Species, References. Available online: http://researcharchive.calacademy.org/research/ichthyology/catalog/fishcatmain.asp (accessed on 10 May 2019).
- Sazima, I. Natural history of Trichogenes longipinnis, a threatened trichomycterid catfish endemic to Atlantic Forest streams in Southeast Brazil. Ichthyol. Explor. Freshw. 2004, 15, 49–60. [Google Scholar]
- Ochoa, L.E.; Roxo, F.F.; Do-Nascimiento, C.; Sabaj, M.H.; Datovo, A.; Alfaro, M.; Oliveira, C. Multilocus analysis of the catfish family Trichomycteridae (Teleostei: Ostariophysi: Siluriformes) supporting a monophyletic Trichomycterinae. Mol. Phylogenetics Evol. 2017, 115, 71–81. [Google Scholar] [CrossRef] [PubMed]
- Katz, A.M.; Barbosa, M.A.; Mattos, J.L.O.; Costa, W.J.E.M. Multigene analysis of the catfish genus Trichomycterus and description of a new South American trichomycterine genus (Siluriformes, Trichomycteridae). Zoosystematics Evol. 2018, 94, 557–566. [Google Scholar] [CrossRef]
- Hayes, M.H.; Paz, H.J.; Stout, C.C.; Werneke, D.C.; Armbruster, J.W. A Hotspot Atop: Rivers of the Guiana Highlands Hold High Diversity of Endemic Pencil Catfish (Teleostei: Ostariophysi: Siluriformes). Biol. J. Linn. Soc. 2020, 129, 862–874. [Google Scholar] [CrossRef]
- De Pinna, M.C.C. A new Sarcoglanidine catfish. phylogeny of its subfamily, and an appraisal of the phyletic status of the Trichomycterinae (Teleostei, Trichomycteridae). Am. Mus. Novit. 1989, 2950, 1–39. [Google Scholar]
- Uieda, V.S.; Castro, R.M.C. Coleta e fixação de peixes de riachos. In Ecologia de Peixes de Riachos; Caramaschi, E.P., Mazzoni, R., Peres-Neto, P.R., Eds.; Universidade Federal do Rio de Janeiro (Oecologia Brasiliensis): Rio de Janeiro, Brazil, 1999; Volume 6, pp. 1–22. [Google Scholar]
- Lucena, C.A.; Calegari, B.; Pereira, E.; Dallegrave, E. O uso de óleo de cravo na eutanásia de peixes. Bol. Soc. Bras. Ictiol. 2013, 105, 20–24. [Google Scholar]
- Close, B.; Banister, K.; Baumans, V.; Bernoth, E.-M.; Bromage, N.; Bunyan, J.; Erhardt, W.; Flecknell, P.; Gregory, N.; Hackbarth, H.; et al. Recommendations for euthanasia of experimental animals Part 2 Report of a Working Party. Lab. Anim. 1997, 31, 1–32. [Google Scholar] [CrossRef]
- Bruford, M.W.; Hanotte, O.; Brookfield, J.F.Y.; Burke, T. Molecular Genetic Analysis of Populations: A Practical Approach; IRL Press: Oxford, UK, 1992; 315p. [Google Scholar]
- Villa-Verde, L.; Lazzarotto, H.; Costa, W.J.E.M.; Lima, S.M.Q. A new Glanapterygine catfish of the genus Listrura (Teleostei: Siluriformes: Trichomycteridae) from southeastern Brazil. Neotrop. Ichthyol. 2012, 10, 527–538. [Google Scholar] [CrossRef]
- Kearse, M.; Moir, R.; Wilson, A.; Stones-Havas, S.; Cheung, M.; Sturrock, S.; Buxton, S.; Cooper, A.; Markowitz, S.; Duran, C.; et al. Geneious Basic: An integrated and extendable desktop software platform for the organization and analysis of sequence data. Bioinformatics 2012, 28, 1647–1649. [Google Scholar] [CrossRef]
- Xia, X.; Xie, Z.; Salemi, M.; Chen, L.; Wang, Y. An index of substitution saturation and its application. Mol. Biol. Evol. 2003, 26, 1–7. [Google Scholar] [CrossRef]
- Xia, X. DAMBE5: A comprehensive software package for data analysis in molecular biology and evolution. Mol. Biol. Evol. 2013, 30, 1720–1728. [Google Scholar] [CrossRef]
- Kumar, S.; Stecher, G.; Tamura, K. MEGA7: Molecular Evolutionary Genetics Analysis version 7.0 for bigger datasets. Mol. Biol. Evol. 2016, 33, 1870–1874. [Google Scholar] [CrossRef]
- Darriba, D.; Taboada, G.L.; Doallo, R.; Posada, D. jModelTest 2: More models. new heuristics and parallel computing. Nat. Methods 2012, 9, 772. [Google Scholar]
- Bouckaert, R.; Vaughan, T.G.; Barido-Sottani, J.; Duchêne, S.; Fourment, M.; Gavryushkina, A.; Heled, J.; Jones, G.; Kühnert, D.; De Maio, N.; et al. BEAST 2.5: An advanced software platform for Bayesian evolutionary analysis. PLoS Comput. Biol. 2019, 15, e1006650. [Google Scholar] [CrossRef] [PubMed]
- Tracer Version v1. 5.0. Available online: http://tree.bio.ed.ac.uk/software/tracer (accessed on 5 May 2020).
- Ochoa, L.E.; Datovo, A.; Do-Nascimiento, C.; Roxo, F.F.; Sabaj, M.H.; Chang, J.; Melo, B.F.; Silva, G.S.C.; Foresti, F.; Alfaro, M.; et al. Phylogenomic analysis of trichomycterid catfishes (Teleostei: Siluriformes) inferred from ultraconserved elements. Sci. Rep. 2020, 10, 2697. [Google Scholar] [CrossRef]
- Hillis, D.; Bull, J. An empirical test of bootstrapping as a method for assessing confidence in phylogenetic analysis. Syst. Biol. 1993, 42, 182–192. [Google Scholar] [CrossRef]
- ESRI. Environmental Systems Research Institute. ArcGIS Desktop: Release 10. Redlands. CA: Environmental Systems Research Institute 2011. Available online: http://www.esri.com/software/arcgis/index.html (accessed on 10 April 2021).
- Kimura, M. A simple method for estimating evolutionary rate of base substitutions through comparative studies of nucleotide sequences. J. Mol. Evol. 1980, 16, 111–120. [Google Scholar] [CrossRef] [PubMed]
- R Core Team. R: A Language and Environment for Statistical Computing; Version 4.2.1; R Foundation for Statistical Computing: Vienna, Austria, 2022; Available online: https://www.R-project.org (accessed on 20 July 2022).
- Hardman, M. The phylogenetic relationships among non-diplomystid catfishes as inferred from mitochondrial cytochrome b sequences: The search for the ictalurid sister taxon (Otophysi: Siluriformes). Mol. Phylogenetics Evol. 2006, 41, 636–662. [Google Scholar] [CrossRef]
- Abell, R.; Thieme, L.M.; Revenga, C.; Bryer, M.; Kottelat, M.; Bogutskaya, N.; Coad, B.; Mandrak, N.; Balderas, S.C.; Bussing, W.; et al. Freshwater Ecoregions of the World: A New Map of Biogeographic Units for Freshwater Biodiversity Conservation. BioScience 2008, 58, 403–414. [Google Scholar] [CrossRef]
- Species 2000 & ITIS Catalogue of Life. Available online: www.catalogueoflife.org/col (accessed on 24 February 2020).
- Unmack, P.J.; Bennin, A.P.; Habit, E.M.; Victoriano, P.F.; Johnson, J.B. Impact of ocean barriers, topography, and glaciation on the phylogeography of the catfish Trichomycterus areolatus (Teleostei: Trichomycteridae) in Chile. Biol. J. Linn. Soc. Lond. 2009, 97, 876–892. [Google Scholar]
- Costa, W.J.E.M.; Katz, A. Integrative taxonomy supports high species diversity of south-eastern Brazilian mountain catfishes of the T. reinhardti group (Siluriformes: Trichomycteridae). Syst. Biodivers. 2021, 19, 601–621. [Google Scholar]
- Endersby, J. Lumpers and Splitters: Darwin, Hooker, and the Search for Order. Science 2009, 326, 1496–1499. [Google Scholar] [CrossRef] [PubMed]
- Hubert, N.; Renno, J.F. Historical biogeography of South American freshwater fishes. J. Biogeogr. 2006, 33, 1414–1436. [Google Scholar] [CrossRef]
- Arratia-F, G.; Chang-G, A.; Menu-Marque, S.; Rojas-M, G. About Bullockia gen. nov., Trichomycterus mendozensis n. sp, and Revision of the Family Trichomycteridae (Pisces, Siluriformes). Stud. Neotrop. Environ. 1978, 13, 157–194. [Google Scholar] [CrossRef]
- Ferraris, C.J. Checklist of catfishes, recent and fossil (Osteichthyes: Siluriformes), and catalogue of siluriform primary types. Zootaxa 2007, 1418, 1–628. [Google Scholar] [CrossRef]
- Arratia-F, G.; Huaquin, L. Morphology of the lateral line system and of the skin of diplomystid and certain primitive loricarioid catfishes and systematic and ecological considerations. Bonn. Zool. Monogr. 1995, 36, 1–110. [Google Scholar]
- Mesa-S, L.M.; Lasso, C.A.; Ochoa, L.E.; Do-Nascimiento, C. Trichomycterus rosablanca (Siluriformes, Trichomycteridae) a new species of hipogean catfish from the Colombian Andes. Biota Colomb. 2018, 19, 95–116. [Google Scholar]
- Arratia-F, G.; Menu-Marque, S. Revision of the freshwater catfishes of the genus Hatcheria (Siluriformes, Trichomycteridae) with commentaries on ecology and biogeography. Zool. Anz. 1981, 207, 88–111. [Google Scholar]
- Tchernavin, V.V. A revision of some Trichomycterinae based on material preserved in the British Museum (Natural History). Proc. Zool. Soc. Lond. 1944, 114, 234–275. [Google Scholar] [CrossRef]
- Eigenmann, C.H. The Pigidiidae, a family of South American catfishes. Mem. Carnegie Mus. 1918, 7, 259–398. [Google Scholar] [CrossRef]
- Farias, I.P.; Orti, G.; Sampaio, I.; Schneider, H.; Meyer, A. The cytochrome b gene as a phylogenetic marker: The limits of resolution for analyzing relationships among cichlid fishes. J. Mol. Evol. 2001, 53, 89–103. [Google Scholar] [CrossRef] [PubMed]
- Albert, J.S.; Tagliacollo, V.A.; Dagosta, F. Diversification of Neotropical Freshwater Fishes. Annu. Rev. Ecol. Evol. Syst. 2020, 8, 27–53. [Google Scholar] [CrossRef]
- Albert, J.S.; Carvalho, T.P. Neogene Assembly of Modern Faunas. In Historical Biogeography of Neotropical Freshwater Fishes; Albert, J.S., Reis, R.E., Eds.; University of California Press: Berkeley, CA, USA, 2011; pp. 119–136. [Google Scholar]
- Ramos, V.A.; Aleman, A. Tectonic of the Andes. In Tectonic Evolution of South America; Cordani, U.G., Milani, E.J., Thomaz-Filho, A., Campos, D.A., Eds.; Academia Brasileira de Ciências e Departamento Nacional da Produção Mineral (DNPM): Rio de Janeiro, Brazil, 2000; pp. 635–685. [Google Scholar]
- Lujan, N.K.; Conway, K.W. Life in the fast lane: A review of rheophily in freshwater fishes. In Extremophile Fishes; Riesch, R., Tobler, M., Plath, M., Eds.; Springer: Cham, Switzerland; New York, NY, USA, 2015; pp. 107–136. [Google Scholar]
- Jégu, M.; Santos, G.M. Révision du status de Myleus setiger Muller and Troschel, 1844 et de Myleus knerii (Steindachner, 1881) (Teleostei: Characidae: Serrasalminae) avec une description complémentaire des deux espèces. Cybium 2002, 26, 33–57. [Google Scholar]
- Rodríguez-Tribaldos, V.; White, N.J.; Roberts, G.G.; Hoggard, M.J. Spatial and temporal uplift history of South America from calibrated drainage analysis. Geochem. Geophys. Geosystems 2017, 18, 2321–2353. [Google Scholar] [CrossRef]
- Ribeiro, A.C.; Riccomini, C.; Leite, J.A.D. Origin of the largest South American transcontinental water divide. Sci. Rep. 2018, 8, 17144. [Google Scholar] [CrossRef] [PubMed]
- Cherem, L.F.S.; Varajão, C.A.C.; Braucher, R.; Bourlés, D.; Salgado, A.A.R.; Varajão, A.C. Long-term evolution of denudational escarpments in southeastern Brazil. Geomorphology 2012, 173–174, 118–127. [Google Scholar] [CrossRef]
- Lima, S.M.Q.; Berbel-Filho, W.M.; Vasconcellos, A.V.; Lazoski, C.; Volpi, T.A.; Lazzarotto, H.; Russo, C.A.M.; Tatarenkov, A.; Avise, J.C.; Sole-Cava, A.M. Rio de Janeiro and other paleodrainages evidenced by the genetic structure of an Atlantic Forest catfish. J. Biogeogr. 2021, 48, 1475–1488. [Google Scholar] [CrossRef]
- Carvalho, L.M.V.; Jones, C.; Liebmann, B. The South Atlantic convergence zone: Intensity, form, persistence, and relationships with intraseasonal to interannual activity and extreme rainfall. J. Clim. 2004, 17, 88–108. [Google Scholar]
- Fernandez, L.; Osinaga, K. A new Trichomycterus (Siluriformes: Trichomycteridae) from Aguarague National Park of the Bolivian preandean region, with comments on relationships within of the genus. Environ. Biol. Fish. 2006, 75, 385–393. [Google Scholar]
- Costa, W.J.E.M. Description de huit nouvelles espèces du genre Trichomycterus (Siluriformes: Trichomycteridae), du Brésil oriental. Revue Française d’Aquariologie et Herpétologie 1992, 18, 101–110. [Google Scholar]
- Triques, M.L.; Vono, V. Three new species of Trichomycterus (Teleostei: Siluriformes: Trichomycteridae) from the Rio Jequitinhonha basin, Minas Gerais, Brazil. Ichthyol. Explor. Freshw. 2004, 15, 161–172. [Google Scholar]
- Buckup, P.A.; Menezes, N.A.; Ghazzi, M.A. Catálogo das Espécies de Peixes de Água Doce do Brasil; Série Livros 23; Museu Nacional: Rio de Janeiro, Brazil, 2007; 195p. [Google Scholar]
- Reis, V.J.C.; De Pinna, M.C.C. The type specimens of Trichomycterus alternatus (Eigenmann, 1917) and Trichomycterus zonatus (Eigenmann, 1918), with elements for future revisionary work (Teleostei: Siluriformes: Trichomycteridae). Zootaxa 2019, 4585, 100–120. [Google Scholar]
- Fernandez, L.; Vari, R.P. New Species of Trichomycterus from the Andes of Argentina with a redescription of Trichomycterus alterus (Siluriformes: Trichomycteridae). Copeia 2002, 3, 739–747. [Google Scholar] [CrossRef]
- Lezama, A.Q.; Triques, M.L.; Santos, P.S. Trichomycterus argos (Teleostei: Siluriformes: Trichomycteridae), a new species from the Doce River Basin, Eastern Brazil. Zootaxa 2012, 3352, 60–68. [Google Scholar] [CrossRef]
- Ardila Rodríguez, C.A.A. Cinco Nuevas Especies de Peces Trichomycterus Para la Región Caribe-Colombia; Departamento del Atlántico. Barranquilla 2016, 2, 1–26. [Google Scholar]
- Reis, V.J.C.; Santos, S.A.; De Pinna, M.C.C.; Pessali, T.C. An osteological CT-scan and description of a new species of Trichomycterus (Trichomycteridae: Siluriformes) from the Rio Doce drainage with remarkable similarities with Bullockia. J. Fish. Biol. 2019, 95, 918–931. [Google Scholar]
- Ferrer, J.; Malabarba, L.R. Taxonomic review of the genus Trichomycterus Valenciennes (Siluriformes: Trichomycteridae) from the laguna dos Patos system, Southern Brazil. Neotrop. Ichthyol. 2013, 11, 217–246. [Google Scholar]
- Castellanos-Morales, C.A.; Galvis, F. Las especies del género Trichomycterus (Siluriformes: Trichomycteridae) en Colombia. Bol. Cient. Mus. Hist. Nat. 2012, 16, 194–206. [Google Scholar]
- Fernandez, L.A. Redescription of the teleost Trichomycterus barbouri (Eigenmann, 1911), occurrence in Argentina and comparison with related species (Ostariophysi: Siluriformes: Trichomycteridae). Stud. Neotrop. Fauna Environ. 2000, 35, 27–33. [Google Scholar] [CrossRef]
- Arratia, G.; Menu-Marque, S. New catfishes of the genus Trichomycterus from the high Andes of South America (Pisces, Siluriformes) with remarks on distribution and ecology. Zool. Jahrb. Abt. Syst. 1984, 111, 493–520. [Google Scholar]
- Barbosa, M.A.; Costa, W.J.E.M. Seven new species of the catfish genus Trichomycterus (Teleostei: Siluriformes: Trichomycteridae) from southeastern Brazil and redescription of T. brasiliensis. Ichthyol. Explor. Freshw. 2010, 21, 97–122. [Google Scholar]
- Ardila-Rodríguez, C.A.A. Trichomycterus cachiraensis (Siluriformes: Trichomycteridae), nueva especie del rio cachira, cuencia del rio Magdalena, Colombia, Dahlia. Rev. Asoc. Colomb. Ictiol. 2008, 10, 33–41. [Google Scholar]
- Lima, S.M.Q.; Lazzarotto, H.; Costa, W.J.E.M. A new species of Trichomycterus (Siluriformes: Trichomycteridae) from lagoa Feia drainage, southeastern Brazil. Neotrop. Ichthyol. 2008, 6, 315–322. [Google Scholar] [CrossRef]
- Barbosa, M.A.; Costa, W.J.E.M. Validade, relações filogenéticas e redescrição de Eremophilus candidus Ribeiro, 1949 (Teleostei, Siluriformes, Trichomycteridae). Arq. Mus. Nac. 2003, 61, 179–188. [Google Scholar]
- De Pinna, M.C.C. Trichomycterus castroi, a new species of trichomycterid catfish (Teleostei: Silurifomes). Ichthyol. Explor. Freshw. 1992, 3, 89–95. [Google Scholar]
- Fernandez, L.; Vari, R.P. New Species of Trichomycterus (Teleostei: Siluriformes: Trichomycteridae) Lacking a Pelvic Fin and Girdle from the Andes of Argentina. Copeia 2000, 4, 990–996. [Google Scholar] [CrossRef]
- Alencar, A.R.; Costa, W.J.E.M. Description of two new species of the catfish genus Trichomycterus from southeastern Brazil (Siluriformes: Trichomycteridae). Zootaxa 2004, 744, 1–8. [Google Scholar]
- Lasso, C.A.; Provenzano, F. Dos nuevas especies de bagres del género Trichomycterus (Siluriformes: Trichomycteridae) de la Gran Sabana, Escudo de las Guayanas, Venezuela. Rev. Biol. Trop. 2003, 50, 1139–1149. [Google Scholar]
- Wosiacki, W.B.; De Pinna, M.C.C. Trichomycterus igobi, a new catfish species from the rio Iguaçu drainage: The largest head in Trichomycteridae (Siluriformes: Trichomycteridae). Neotrop. Ichthyol. 2008, 6, 17–23. [Google Scholar] [CrossRef]
- Rizzato, P.P.; Costa, E.P.D., Jr.; Trajano, E.; Bichuette, M.E. Trichomycterus dali: A new highly troglomorphic catfish (Silurifomes: Trichomycteridae) from Serra da Bodoquena, Mato Grosso do Sul State, Central Brazil. Neotrop. Ichthyol. 2011, 9, 477–491. [Google Scholar] [CrossRef]
- Bockmann, F.A.; Casatti, L.; De Pinna, M.C.C. A new species of trichomycterid catfish from the Rio Paranapanema basin, southeastern Brazil (Teleostei: Siluriformes), with comments on the phylogeny of the family. Ichthyol. Explor. Freshw. 2004, 15, 225–242. [Google Scholar]
- Barbosa, M.A. Description of two new species of the catfish genus Trichomycterus (Teleostei: Siluriformes: Trichomycteridae) from the coastal river basins, southeastern Brazil. Vertebr. Zool. 2013, 63, 269–275. [Google Scholar]
- Lima, S.M.Q.; Costa, W.J.E.M. Trichomycterus giganteus (Siluriformes: Loricarioidea: Trichomycteridae): A new catfish from the Rio Guandu basin, southeastern Brazil. Zootaxa 2005, 761, 1–6. [Google Scholar] [CrossRef]
- Fernández, L.; Schaefer, S.A. New Trichomycterus (Siluriformes: Trichomycteridae) from an offshore Island of Colombia. Copeia 2005, 1, 68–76. [Google Scholar]
- Do-Nascimiento, C.; Prada-Pedreros, S.; Guerrero-Kommritz, J. Trichomycterus venulosus (Steindachner, 1915), a junior synonym of Eremophilus mutisii Humboldt, 1805 (Siluriformes: Trichomycteridae) and not an extinct species. Neotrop. Ichthyol. 2014, 12, 707–715. [Google Scholar]
- Wosiacki, W.B. A new species of Trichomycterus (Siluriformes: Trichomycteridae) from south Brazil and redescription of T. iheringi (Eigenmann). Zootaxa 2005, 1040, 49–64. [Google Scholar]
- Fernández, L.; Vari, R.P. New species of Trichomycterus from the Andean Cordillera of Argentina (Siluriformes: Trichomycteridae). Copeia 2009, 1, 195–202. [Google Scholar]
- Caramaschi, E.P.; Caramaschi, U. Taxonomic Status of the Trichomycterid Catfish Trichomycterus itatiayae. Copeia 1991, 1, 222–224. [Google Scholar] [CrossRef]
- Barbosa, M.A.; Costa, W.J.E.M. Description of a new species of catfish from the upper rio Paraíba do Sul basin, south-eastern Brazil (Teleostei: Siluriformes: Trichomycteridae) and re-description of Trichomycterus itatiayae. Int. J. Ichthyol. 2008, 14, 176–186. [Google Scholar]
- Wosiacki, W.B.; Oyakawa, O.T. Two new species of the catfish genus Trichomycterus (Siluriformes: Trichomycteridae) from the rio Ribeira de Iguape Basin, Southeastern Brazil. Neotrop. Ichthyol. 2005, 3, 465–472. [Google Scholar]
- Barbosa, M.A.; Costa, W.J.E.M. Trichomycterus macrophthalmus (Teleostei: Siluriformes: Trichomycteridae), a new species of catfish from the Paraíba do Sul River basin, southeastern Brazil. Vertebr. Zool. 2012, 62, 79–82. [Google Scholar] [CrossRef]
- Barbosa, M.A.; Costa, W.J.E.M. Description of a new species of the catfish genus Trichomycterus (Teleostei: Siluriformes: Trichomycteridae) from the rio Paraíba do Sul basin, southeastern Brazil. Vertebr. Zool. 2010, 60, 193–197. [Google Scholar] [CrossRef]
- Bockmann, F.A.; Sazima, I. Trichomycterus maracaya, a new catfish from the upper rio Paraná, southeastern Brazil (Siluriformes: Trichomycteridae), with notes on the T. brasiliensis species complex. Neotrop. Ichthyol. 2004, 2, 61–74. [Google Scholar]
- Wosiacki, B.W.; Garavello, J.C. Five new species of Trichomycterus from the rio Iguaçu (rio Paraná Basin), southern Brazil (Siluriformes: Trichomycteridae). Ichthyol. Explor. Freshw. 2004, 15, 1–16. [Google Scholar]
- Fernandez, L.; Chuquihuamani, R.Q. Species of Trichomycterus (Siluriformes: Trichomycteridae) from the Andean Cordillera of Perú, with comments on relationships within the genus. Zootaxa 2007, 1545, 49–57. [Google Scholar]
- Reis, V.J.C.; dos Santos, S.A.; Britto, M.R.; Volpi, T.A.; De Pinna, M.C.C. Iterative taxonomy reveals a new species of Trichomycterus Valenciennes 1832 (Siluriformes, Trichomycteridae) widespread in rio Doce basin: A pseudocryptic of T. immaculatus. J. Fish Biol. 2020, 97, 1607–1623. [Google Scholar]
- Fernandez, L.; Vari, R.P. New Species of Trichomycterus (Teleostei: Siluriformes) from the Andean Cordillera of Argentina and the Second Record of the Genus in Thermal Waters. Copeia 2012, 4, 631–636. [Google Scholar] [CrossRef]
- Ardila-Rodriguez, C.A. Trichomycterus nietoi sp. nov. (Siluriformes: Trichomycteridae) Una nueva especie de pez del Río Guachaca Sierra Nevada de Santa Marta, Departamento del Magdalena, Colombia. Barranquilla 2014, 4, 1–24. [Google Scholar]
- Costa, W.J.E.M.; Katz, A.M.; Mattos, J.L.O.; Amorim, P.F.; Mesquita, B.O.; Vilardo, P.J.; Barbosa, M.A. Historical review and redescription of three poorly known species of the catfish genus Trichomycterus from south-eastern Brazil (Siluriformes: Trichomycteridae). J. Nat. Hist. 2020, 53, 2905–2928. [Google Scholar]
- Ardila Rodriguez, C.A. Trichomycterus maldonadoi (Siluriformes, Trichomycteridae), especie nueva de la cuenca alta del rio Sinú, Colombia. Dahlia 2011, 11, 13–21. [Google Scholar]
- Alencar, A.R.; Costa, W.J.E.M. Trichomycterus pauciradiatus, a new catfish species from the upper rio Paraná basin. southeastern Brazil (Siluriformes: Trichomycteridae). Zootaxa 2006, 1269, 43–49. [Google Scholar]
- Sarmento-Soares, L.M.; Zanata, A.M.; Martins-Pinheiro, R.F. Trichomycterus payaya, new catfish (Siluriformes: Trichomycteridae) from headwaters of rio Itapicuru. Bahia. Brazil. Neotrop. Ichthyol. 2011, 9, 261–271. [Google Scholar]
- Datovo, A.; Carvalho, M.; Ferrer, J. A new species of the catfish genus Trichomycterus from the La Plata River basin, southern Brazil, with comments on its putative phylogenetic position (Siluriformes: Trichomycteridae). Zootaxa 2012, 3327, 33–44. [Google Scholar]
- Barbosa, M.A.; Azevedo-Santos, V.M. A new species of the catfish genus Trichomycterus (Teleostei: Siluriformes: Trichomycteridae) from the rio Paraná basin, southeastern Brazil. Vertebr. Zool. 2012, 62, 357–362. [Google Scholar]
- Barbosa, M.A.; Costa, W.J.E.M. Trichomycterus potschi (Siluriformes: Trichomycteridae): A new species of trichomycterid catfish from coastal streams of southeastern Brazil. Ichthyol. Explor. Freshw. 2003, 14, 281–287. [Google Scholar]
- Sarmento-Soares, L.M.; Martins-Pinheiro, R.F.; Aranda, A.T.; Chamon, C.C. Trichomycterus pradensis. a new catfish from southern Bahia coastal rivers. northeastern Brazil (Siluriformes: Trichomycteridae). Ichthyol. Explor. Freshw. 2005, 16, 289–302. [Google Scholar]
- Fernandez, L.; Vari, R.P. New species of Trichomycterus from mid elevation localities of northwestern Argentina (Siluriformes: Trichomycteridae). Copeia 2004, 4, 876–882. [Google Scholar]
- Barbosa, M.A.; Costa, W.J.E.M. Trichomycterus puriventris (Teleostei: Siluriformes: Trichomycteridae), a new species of catfish from the rio Paraíba do Sul basin, southeastern Brazil. Vertebr. Zool. 2012, 62, 155–160. [Google Scholar] [CrossRef]
- Bichuette, M.E.; Rizzato, P.P. A new species of cave catfish from Brazil, Trichomycterus rubbioli sp.n., from Serra do Ramalho karstic area, São Francisco River basin, Bahia State (Silurifomes: Trichomycteridae). Zootaxa 2012, 3480, 48–66. [Google Scholar]
- Ardila-Rodriguez, C.A.A. Trichomycterus ruitoquensis (Siluriformes: Trichomycteridae) una nueva especie de pez de la cuenca alta del río Lebrija, Departamento de Santander-Colombia. Barranquilla 2007, 3, 1–20. [Google Scholar]
- Castellanos-Morales, C.A. Trichomycterus sketi: A new species of subterranean catfish (Siluriformes: Trichomycteridae) from the Andean Cordillera of Colombia. Biota Colombiana 2011, 11, 33–41. [Google Scholar]
- Barbosa, M.A.; Costa, W.J.E.M. Description of a new species of the catfish genus Trichomycterus (Teleostei: Siluriformes: Trichomycteridae) from the rio de Contas basin, northeastern Brazil. Vertebr. Zool. 2011, 61, 307–312. [Google Scholar]
- Fernandez, L.; Miranda, G. A catfish of the genus Trichomycterus from a thermal stream in southern South America (Teleostei. Siluriformes. Trichomycteridae), with comments on relationships within the genus. J. Fish. Biol. 2007, 71, 1303–1316. [Google Scholar] [CrossRef]
- Wosiacki, W.B. New species of the catfish Trichomycterus (Siluriformes: Trichomycteridae) from the headwaters of the rio São Francisco basin, Brazil. Zootaxa 2004, 592, 1–12. [Google Scholar] [CrossRef]
- Ferrer, J.; Malabarba, L.R. A new Trichomycterus lacking pelvic fins and pelvic girdle with a very restricted range in Southern Brazil (Siluriformes: Trichomycteridae). Zootaxa 2011, 2912, 59–67. [Google Scholar]
- Castellanos-Morales, C.A. Trichomycterus uisae: A new species of hypogean catfish (Siluriformes: Trichomycteridae) from the northeastern Andean Cordillera of Colombia. Neotrop. Ichthyol. 2008, 6, 307–314. [Google Scholar] [CrossRef]
- Fernandez, L.; Schaefer, S.A. Trichomycterus yuska, a new species from high elevations of Argentina (Siluriformes: Trichomycteridae). Ichthyol. Explor. Freshw. 2003, 14, 353–360. [Google Scholar]
- Ferrer, J.; Malabarba, L.R. Systematic revision of the Neotropical catfish genus Scleronema (Siluriformes:Trichomycteridae), with descriptions of six new species from Pampa grasslands. Neotrop. Ichthyol. 2020, 18, e190081. [Google Scholar] [CrossRef]
- Lima, S.M.Q.; Vasconcellos, A.V.; Berbel-Filho, W.M.; Lazoski, C.; Russo, C.A.; Sazima, I.; Solé-Cava, A.M. Effects of Pleistocene climatic and geomorphological changes on the population structure of the restricted-range catfish Trichogenes longipinnis (Siluriformes: Trichomycteridae). Syst. Biodivers. 2016, 14, 155–170. [Google Scholar]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| % | A | B | C | D | E | F | G | H | I | J | K | L |
|---|---|---|---|---|---|---|---|---|---|---|---|---|
| A | 0.067 | 0.016 | 0.017 | 0.022 | 0.021 | 0.020 | 0.019 | 0.022 | 0.023 | 0.029 | 0.034 | 0.031 |
| B | 0.103 | 0.054 | 0.018 | 0.017 | 0.021 | 0.020 | 0.020 | 0.020 | 0.022 | 0.028 | 0.033 | 0.030 |
| C | 0.111 | 0.113 | 0.051 | 0.022 | 0.020 | 0.019 | 0.019 | 0.022 | 0.024 | 0.028 | 0.035 | 0.029 |
| D | 0.129 | 0.105 | 0.120 | 0.072 | 0.026 | 0.024 | 0.022 | 0.025 | 0.025 | 0.029 | 0.033 | 0.035 |
| E | 0.125 | 0.122 | 0.110 | 0.135 | 0.032 | 0.018 | 0.017 | 0.022 | 0.022 | 0.026 | 0.032 | 0.037 |
| F | 0.128 | 0.123 | 0.105 | 0.131 | 0.101 | 0.050 | 0.016 | 0.019 | 0.021 | 0.025 | 0.029 | 0.031 |
| G | 0.124 | 0.124 | 0.106 | 0.128 | 0.097 | 0.098 | 0.064 | 0.020 | 0.020 | 0.026 | 0.031 | 0.033 |
| H | 0.140 | 0.128 | 0.137 | 0.151 | 0.128 | 0.126 | 0.124 | 0.095 | 0.023 | 0.027 | 0.035 | 0.035 |
| I | 0.143 | 0.134 | 0.131 | 0.150 | 0.120 | 0.129 | 0.117 | 0.137 | 0.031 | 0.028 | 0.034 | 0.040 |
| J | 0.156 | 0.153 | 0.152 | 0.154 | 0.131 | 0.136 | 0.147 | 0.157 | 0.165 | n/c | 0.039 | 0.040 |
| K | 0.179 | 0.175 | 0.192 | 0.181 | 0.176 | 0.162 | 0.176 | 0.191 | 0.185 | 0.201 | 0.046 | 0.041 |
| L | 0.177 | 0.176 | 0.168 | 0.190 | 0.192 | 0.171 | 0.184 | 0.191 | 0.221 | 0.205 | 0.205 | 0.000 |
| Group | Minimum | Mean | Maximum | Mean (Intra) |
|---|---|---|---|---|
| Trichomycterus s.s. | 4.3 | 8.2 | 11.3 | 8.5 |
| Cambeva | 6.3 | 8.6 | 10.9 | |
| Hatcheria | 2.5 | 6.3 | 11.5 | |
| Guiana genus | 4.8 | 9.2 | 11 | |
| Ituglanis | 4.3 | 10.4 | 13.1 | |
| Genera into Trichomycterinae | 9.7 | 12.9 | 15.7 | 5.61 |
| Subfamilies (3) | 17.8 | 18.9 | 20.5 | 7.2 |
| Genus/Clade | Subclade | Species | Distribution |
|---|---|---|---|
| Trichomycterus (A) | 1 | Trichomycterus alternatus, T. longibarbatus, T. pantherinus, T. caudofasciatus, T. immaculatus, T. mimosensis | Contas, Jequitinhonha, Barra Seca, São Mateus, Doce, Timbui, Piraque-Açu, Santa Maria da Vitoria, Jucu, itapemirim, itabapoana, Paraiba do Sul, Ribeira do Iguape |
| 2 | Trichomycterus tete | Contas, Jequitinhonha, Piraque-Açu | |
| 3 | Trichomycterus pradensis | Buranhem, Jucuruçu, Mucuri, itaunas, Barra Seca, Doce, Jucu | |
| 4 | Trichomycterus landinga | Jequitinhonha | |
| 5 | Trichomycterus brasiliensis, T. brunoi, T. candidus, T. macrotrichopterus, T. pirabitira, T. rubiginosus | Itabapoana. Jequitinhonha. São Francisco. Parana | |
| 6 | Trichomycterus nigricans | Santa Maria da Vitoria. Jucu, itapemirim, itabapoana. Paraiba do Sul | |
| 7 | Trichomycterus nigroauratus, T. cf. paquequerensis, T. cf. itatyae | Paraiba do Sul. Parana | |
| 8 | Trichomycterus caipora | Paraiba do Sul | |
| 9 | Trichomycterus giganteus | Baia de Guanabara | |
| 10 | Trichomycterus reinhardti, T. septemradiatus | Parana. São Francisco | |
| 11 | Trichomycterus itatyae | Paraiba do Sul | |
| Cambeva (B) | 12 | Cambeva cubataonis, C. davisi, C. zonatus | Itapocu, Ribeira do Iguape, Paranapanema |
| 13 | Cambeva iheringi | Paraiba do Sul, Parana | |
| 14 | Cambeva perkos, C. poikilos, C. diatropoporos | Jacui, Uruguay | |
| Scleronema (C) | 15 | Scleronema minutum, S. operculatum | Uruguay |
| D | 16 | Trichomycterus sandovali, T. cf. knerii | Magdalena, Orinoco |
| Hatcheria (E) | 17 | Trichomycterus areolatus | Transandinean (South) |
| 18 | Trichomycterus areolatus | Transandinean (South) | |
| 19 | Hatcheria macraei, Trichomycterus areolatus | Transandinean (South) | |
| 20 | Trichomycterus areolatus | Transandinean (South) | |
| 21 | Bullockia maldonadoi | Transandinean (South) | |
| 22 | Trichomycterus areolatus | Transandinean (South) | |
| F | 23 | Trichomycterus striatus, T. straminius | Magdalena |
| Guiana Clade (G) | 24 | Trichomycterus cf. guianense | Potaro, Guyana |
| 25 | Trichomycterus guianense (Mazaruni, plain) | Potaro, Guyana | |
| 26 | Trichomycterus guianense (Potaro, elongate) | Potaro, Guyana | |
| 27 | Trichomycterus guianense | Potaro, Guyana | |
| 28 | Trichomycterus conradi | Potaro, Guyana | |
| Ituglanis (H) | 29 | Ituglanis parkoi | Tapajós–Amazonas basin |
| 30 | Ituglanis herberti | Araguaia–Tocantins and Pirai–Paraná basins | |
| 31 | Ituglanis boitata | Jacui-Laguna dos Patos basin | |
| 32 | Ituglanis parahybae, ituglanis sp.n1 | Macabu and Ribeira do Iguape basins | |
| 33 | Ituglanis amazonicus | Mamoré–Madeira basin | |
| 34 | Ituglanis eichhorniarum | Taquari and Aquidauana –Paraguay, Paraguay–Paraná basins | |
| I | 35 | Trichomycterus transandianus | Magdalena |
| J | 36 | Trichomycterus sp.2 (Tapajos) | Tapajos, Amazonas |
| Listrura (K) | 37 | Listrura nematopteryx, L. picinguabae | Neotropical |
| Trichogenes (L) | 38 | Trichogenes longipinnis | Rio de Janeiro and São Paulo States |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Volpi, T.d.A.; Monjardim, M.; Sarmento-Soares, L.M.; Fagundes, V. Pleistocene Aquatic Refuges Support the East–West Separation of the Neotropical Catfish Trichomycterinae (Siluriformes: Trichomycteridae) and High Diversity in the Magdalena, Guiana, and Paraná-Paraguay Basins. Diversity 2023, 15, 929. https://doi.org/10.3390/d15080929
Volpi TdA, Monjardim M, Sarmento-Soares LM, Fagundes V. Pleistocene Aquatic Refuges Support the East–West Separation of the Neotropical Catfish Trichomycterinae (Siluriformes: Trichomycteridae) and High Diversity in the Magdalena, Guiana, and Paraná-Paraguay Basins. Diversity. 2023; 15(8):929. https://doi.org/10.3390/d15080929
Chicago/Turabian StyleVolpi, Thais de Assis, Marina Monjardim, Luisa M. Sarmento-Soares, and Valéria Fagundes. 2023. "Pleistocene Aquatic Refuges Support the East–West Separation of the Neotropical Catfish Trichomycterinae (Siluriformes: Trichomycteridae) and High Diversity in the Magdalena, Guiana, and Paraná-Paraguay Basins" Diversity 15, no. 8: 929. https://doi.org/10.3390/d15080929
APA StyleVolpi, T. d. A., Monjardim, M., Sarmento-Soares, L. M., & Fagundes, V. (2023). Pleistocene Aquatic Refuges Support the East–West Separation of the Neotropical Catfish Trichomycterinae (Siluriformes: Trichomycteridae) and High Diversity in the Magdalena, Guiana, and Paraná-Paraguay Basins. Diversity, 15(8), 929. https://doi.org/10.3390/d15080929